L
Journal of Experimental Marine Biology and Ecology 251 (2000) 205–225
www.elsevier.nl / locate / jembe
Effect of reproduction on escape responses and muscle
¨
metabolic capacities in the scallop Chlamys islandica Muller
1776
*
Katherina B. Brokordt , John H. Himmelman, Helga E. Guderley
´ ´
Departement de Biologie and GIROQ(Groupe Interuniversitaire de Recherches Oceanographiques du
´ ´ ´ ´
Quebec), Universite Laval, Cite Universitaire, Ste.-Foy, Quebec, Canada G1K 7P4 Received 15 January 2000; received in revised form 27 March 2000; accepted 15 April 2000
Abstract
In scallops, gametogenesis leads to mobilization of glycogen and proteins from the adductor muscle towards the gonad. This mobilization is likely to diminish the metabolic capacities of the adductor muscle and thereby the scallops’ escape response. We examined the escape response in terms of number of valve claps until exhaustion, rate of clapping and the recovery during and after valve closure in adult scallops, Chlamys islandica, sampled at different stages in the reproductive cycle (immature, mature, before and after spawning). In parallel, we measured muscle glycogen, protein and phosphoarginine contents, the oxidative capacity of mitochondria isolated from the adductor muscle and levels of muscle enzymes which are active during exercise and recovery. The
21
number of claps (24–26), rate of clapping (|13 claps min ) and phosphoarginine and arginine
kinase levels were similar during the different reproductive stages. All immature scallops responded to restimulation immediately after opening their valves, while only 62% of mature, 82% of prespawned and 38% of spawned scallops responded. Immature animals completely recovered their initial swimming capacity within 4 h of opening their valves, but mature, prespawned and spawned scallops needed 18, 12 and 18 h, respectively. Overall phasic adductor muscle from mature, prespawned and spawned animals showed decreased glycogen phosphorylase, phospho-fructokinase, pyruvate kinase (except for prespawned), octopine dehydrogenase and citrate synthase levels, a deterioration of the oxidative capacity of mitochondria and a marked decrease in glycogen content compared to immature scallops. Therefore, during gonadal maturation and spawning, C. islandica did not change its clapping capacity, but slowed its recuperation from exhausting burst exercise, both during and after valve closure, likely due to the decreased metabolic capacity of the adductor muscle. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Chlamys islandica; Escape response; Muscle; Reproduction; Scallop
*Corresponding author. Tel.:11-418-656-2131; fax:11-418-656-2339.
E-mail address: [email protected] (K.B. Brokordt).
1. Introduction
Gametogenesis represents a period of high energy demand, particularly for organisms with broadcast spawning, and when external food supplies are limited, gamete production occurs at the expense of biochemical components in somatic tissues (Calow, 1985). In bivalves and fish, muscle is one of the tissues most affected during gonadal maturation, with both protein and glycogen levels decreasing (Shevechenko, 1972; reviewed by Barber and Blake, 1991; Von der Decken, 1992). Such mobilization of macromolecules could decrease muscle metabolic capacities.
In scallops the adductor muscle is one of the largest soft tissues and while its primary role is the movement of the valves during escape responses or swimming, it also serves as a major site of reserve deposition. In the bay scallop Argopecten irradians
concentricus, 63–99% of total energy gain by the gonad during gametogenesis is
thought to come from the adductor muscle (Epp et al., 1988). In this species, gametogenesis occurs mainly at the expense of muscle protein, however, in other scallops the substrate most mobilized is glycogen (reviewed by Barber and Blake, 1991;
´ ´
Martınez, 1991; Martınez and Mettifogo, 1998).
How far the reproductive cycle affects biochemical reserves may depend on the timing of gonadal proliferation in relation to food availability (Shafee, 1981). In the black scallop Chlamys varia, during the spring reproduction, when food is abundant, only carbohydrate reserves are used for gonadal development, while during the autumn reproduction, when food is less abundant, all reserves (protein, lipid and glycogen) decrease with gonadal development (Shafee, 1981). In fishes, such as the salmon,
Oncorhynchus nerka and Salmo salar, not only are lipid reserves from muscle mobilized
during gonadal maturation and their non-feeding spawning migration, but the levels of enzymes and contractile proteins are also affected (Mommsen et al., 1980; Olin and von der Decken, 1987). In the roach Rutilus rutilis, swimming activity is markedly reduced during gonadal maturation (Koch and Wieser, 1983), either due to the metabolic costs of gonad maturation or to decreases in muscle metabolic capacities. A decrease of muscle metabolic capacities after gonadal maturation and spawning could reduce an animal’s locomotor ability and thereby the capacity to escape predators.
Among bivalves, scallops are well known for their swimming ability (Brand, 1991). They swim by jet propulsion using a succession of claps consisting of alternate adduction and abduction of the valves (Olson and Marsh, 1993). Swimming is powered by the adductor muscle, principally by the large phasic portion and to a lesser extent by the smaller tonic (catch) muscle (de Zwaan et al., 1980).
The physiological and biochemical aspects of the escape response (valve clapping and valve closure) are best understood in the giant scallop Placopecten magellanicus (Thompson et al., 1980; de Zwaan et al., 1980; Livingstone et al., 1981) and in the bay scallop Argopecten irradians concentricus (Chih and Ellington, 1983, 1986). In
Placopecten magellanicus, the main source of ATP during valve clapping is arginine
Once the valves have opened, the full restoration of arginine phosphate pools and elimination of octopine is achieved aerobically (Livingstone et al., 1981). Similarly, in
Argopecten irradians concentricus intense valve clapping is initially supported by
arginine phosphate and only toward the end do anaerobic glycolysis and octopine accumulation intervene (Chih and Ellington, 1983, 1986). Mitochondria isolated from the adductor muscle of the tropical scallop, Euvola ziczac, seem adapted for a role in recovery metabolism given their increased affinity for pyruvate at the pH values likely to occur in muscle fibers after intense clapping (Guderley et al., 1995). Oxidative capacities and respiratory control ratios of mitochondria isolated from the adductor muscle are lower during the first of two spawnings than during other periods (Boadas et al., 1997), suggesting an impact of reproduction.
We reasoned that changes in muscle metabolic capacities or in the levels of energetic reserves during the reproductive cycle of adult scallops could modify the escape response or recuperation from an exhausting escape response. We predicted that before gonadal maturation, scallops would show a stronger escape response and would recuperate faster from exhausting burst exercise given the higher muscle metabolic capacities and energy reserves the scallops should have at this time.
To evaluate these hypotheses, we compared the escape response and capacity for recuperation from exhausting exercise for adult Iceland scallops, Chlamys islandica, sampled at different stages in the annual reproductive cycle (immature, mature, before and after spawning). In parallel, we determined the effect of these reproductive stages on the energetic reserves in the phasic adductor muscle (arginine phosphate, glycogen and proteins). Given that in fish muscle, starvation leads to more extensive mobilization of sarcoplasmic than structural (insoluble) proteins (Beaulieu and Guderley, 1998), we quantified both fractions. To assess the impact of the reproductive cycle on muscle metabolic capacities, we measured muscle levels of the glycolytic enzymes, glycogen phosphorylase (GP), pyruvate kinase (PK), phosphofructokinase (PFK), octopine dehydrogenase (ODH) and arginine kinase (AK), as well as the mitochondrial enzyme, citrate synthase (CS). We measured the oxidative capacities, substrate preferences, respiratory control ratios and CS levels of mitochondria isolated from the phasic adductor muscle at different reproductive stages. By profiling these enzyme activities and mitochondrial capacities we sought to assess the capacity of enzymes likely to supply ATP for contraction (AK and glycolytic enzymes (GP, PFK, PK and ODH)), recuperation during valve closure (glycolytic enzymes) and recuperation after reopening of the valves (CS and mitochondrial capacities).
2. Materials and methods
2.1. Sampling and maintenance of scallops
ˆ ˆ
spawning (prespawned) and after spawning (spawned). Reproductive stage was de-termined using qualitative and quantitative criteria. As qualitative criteria we used the shape of the gonad and the degree of vascularization by secondary genital ducts which were observable through the gonadal epithelium (David Arsenault, personal communica-tion). Histological classification used the gonadal developmental stages described for
´
Chlamys islandica by Thorarinsdottir (1993). As quantitative criteria, we determined the
gonadosomatic index corrected for the size of the scallop ((gonadal mass3total tissue
21 21
mass )3shell height ), as described by Bonardelli and Himmelman (1995). We also evaluated the lipid and protein content of the gonads on the different sampling dates.
At each sampling period, we determined the shell height and wet mass of the gonad, muscle and remaining soft tissues of 20 scallops. One part of the phasic muscle was rapidly excised, quickly frozen (|1 min) using a freeze clamping press (Gagnon et al.,
1998) prechilled in liquid nitrogen and stored at 2708C for determination of enzyme activities and arginine phosphate concentrations. The remainder was frozen at 2208C for protein and glycogen determinations. Part of the gonad was frozen at 2208C for
´ lipid and protein determinations. Ten animals were transported live to Universite Laval in Quebec City where they were maintained in sea water aquaria and fed with Isochrysis
galbana and Chaetoceros gracilis for at least one day, before determinations of the
respiratory capacity of muscle mitochondria.
The remaining 48 scallops were acclimated during 2 days in holding tanks with running sea water pumped from 10 m in depth at Havre St. Pierre, and then used to evaluate escape responses. Despite the variations in ambient temperature, at the time of measurements of escape responses in the laboratory water temperatures were con-sistently 5–68C.
2.2. Environmental conditions
Bottom temperature was recorded using a Sealog-T thermograph (Vemco Inc., Halifax, N.S.), which was anchored on the bottom in the scallop bed. Phytoplankton abundance was quantified periodically from water samples collected with a Niskin bottle from 1 m above the scallop bed. From each sample, chlorophyll a determinations were made using the spectrophotometric method as described by Parsons et al. (1984). 2.3. Evaluation of escape responses
When the scallop, Chlamys islandica, is stimulated with its natural predator, the seastar Leptasterias polaris (Himmelman, 1991), the escape response is highly stereotyped and reproducible. It begins with a series of valve claps and if the stimulus is maintained until fatigue, the scallop closes its valves firmly and remains closed for a certain period, after which the valves slowly reopen.
were restimulated (in the same fashion as initially) at set times after reopening of the valves. One group was restimulated after 2 min, and the others 2, 4, 6, 12 or 18 h after reopening the valves. Again, the number of claps was determined for each animal. 2.4. Metabolite assays
2.4.1. Muscle phosphoarginine
Deproteinization of the samples followed the method described by Lamprecht and Trautschold (1974). The neutralized perchloric acid extracts were used immediately for the determination of phosphoarginine, in an assay analogous to the phosphocreatine assay described by Lamprecht et al. (1974).
2.4.2. Muscle glycogen
The concentration of glycogen was determined by enzymatic hydrolysis with amyloglucosidase as described by Keppler and Decker (1974).
2.4.3. Muscle and gonad protein
Muscle proteins were separated into sarcoplasmic and myofibrillar fractions according to the method of Bates and Millard (1983) as modified by Somero and Childress (1990). Muscle and gonad protein concentrations were determined using the bicinchoninic acid method of Smith et al. (1985) with bovine serum albumin as the standard.
2.4.4. Gonadal lipid
The lipid concentration in the gonad was estimated gravimetrically after extraction with chloroform–methanol according to the method of Folch et al. (1957), except that after the centrifugations the sediment was resuspended for an additional extraction. 2.5. Enzyme assays
Samples of phasic adductor muscle were homogenized on ice, using a Polytron (Brinkman Instruments; Rexdale, Ontario, Canada), in 10 vol of 50 mM imidazole–HCl, 2 mM EDTA-Na2 (ethylene dinitrilotetraacetic acid), 5 mM EGTA (ethyleneglycol tetraacetic acid), 1 mM dithiothreitol, 0.1% Triton X-100, pH 6.6 for PK, ODH and AK, pH 7.2 for GP, PFK and CS. One part of the homogenates was centrifuged at 10 0003g
at 48C for 15 min for the assay of GP and PFK, and another at 6003g at 48C for 15 min for the assay of CS.
Assay temperature was controlled at 68C with a circulating refrigerating water bath (Haake D8). Enzyme activity was measured using a UV/ VIS spectrophotometer (Beckman / DU 640) to follow the absorbance changes of NAD(P)H at 340 nm, with the exception of CS which was monitored at 412 nm to detect the transfer of sulfydryl groups from CoASH to 5,59-dithio-bis(2-nitro)benzoic acid (DTNB). The molar extinction coefficients used for NAD(P)H and DTNB were 6.22 and 13.6, respectively. All assays were run in duplicate and the specific activities were expressed in
21 21
Conditions for enzyme assays (except for CS) were adapted from conditions used by de
21
Zwaan et al. (1980) for P. magellanicus, as follows (all concentrations in mmol l ): 2.5.1. Glycogen phosphorylase (EC 2.4.1.1)
21
Imidazole–HCl 50, KH PO2 4 80, Mg-acetate 5, EDTA 2.5, 10 mg ml glycogen (omitted for the control), AMP 0.8, AMP cyclic 0.5, NADP 0.6, glucose-1,6-diphosphate 0.004, excess glucose-6-phosphate dehydrogenase and phosphoglucomutase, pH 7.5. 2.5.2. Phosphofructokinase (EC 2.7.1.11)
Tris–HCl 50, KCl 50, Mg-acetate 5, fructose-6-phosphate 1 (omitted for the control), fructose-2,6-diphosphate 0.08, ATP 1, AMP 0.8, NADH 0.2, excess aldolase, glycerol-3-phosphate dehydrogenase and trioseglycerol-3-phosphate isomerase, pH 7.5.
2.5.3. Pyruvate kinase (EC 2.7.1.40)
Imidazole–HCl 50, MgSO4 13, KCl 100, phosphoenolpyruvate 5 (omitted for the control), ADP 5, NADH 0.2, excess lactate dehydrogenase, pH 6.6.
2.5.4. Arginine kinase (EC 2.7.3.3)
Imidazole–HCl 50, MgCl 5, phosphoarginine 5 (omitted for the control), glucose 10,2
ADP 0.4, NADP 0.6, excess hexokinase and glucose-6-phosphate dehydrogenase, pH 6.6.
2.5.5. Octopine dehydrogenase
Imidazole–HCl 50, EDTA-Na 2, EGTA 5, KCN 1, sodium pyruvate 5 (omitted for2
the control), arginine–HCl 6, NADH 0.2, pH 6.6. 2.5.6. Citrate synthase (EC 4.1.3.7)
Tris–HCl 75, oxaloacetate 0.3 (omitted for the control), DTNB 0.1, acetyl CoA 0.2, pH 8.0.
2.6. Mitochondrial assays
2.6.1. Mitochondrial isolation and polarographic measurements
All procedures followed Guderley et al. (1995) as adapted from Ballantyne and Moon (1985), except that media were prepared in nanopure water. Mitochondrial oxygen uptake was measured in a water-jacketed respiration cell (Cameron Instruments) using a Clarke-type electrode (Yellow Springs Instrument Comp., Yellow Springs, OH). The electrode was connected to a chart recorder (Linear, model 0585, Canlab Comp.) by an oxymeter (Cameron Instruments, Port Aransas, TX). The respiration chamber was maintained at 68C by a circulating refrigerated water bath. The values of oxygen concentration were calculated from the data of Graham (1987) for physiological buffers using the atmospheric pressure on the day of assay. Around 1.3 mg mitochondrial protein was added to 1 ml of assay medium. The reaction medium contained (in
21
Preliminary studies indicated that glutamate, malate, pyruvate and succinate at final
21
concentrations of 30, 6, 0.9 and 24 mmol l , respectively, gave maximal rates. To attain
21
maximal rates of pyruvate oxidation, ‘sparking’ levels of malate (0.6 mmol l ), which by themselves did not support significant rates of oxygen uptake, were required. To measure maximal rates of respiration we added ADP at a final concentration of 0.6
21
mmol l . The respiratory control ratio (RCR) was calculated from the ratio of the state 3 rate (in presence of ADP) to state 4 rate (when all ADP had been phosphorylated) (Estabrook, 1967).
To estimate the maximal aerobic capacity of the phasic adductor muscle, we measured the level of CS in the mitochondrial suspensions and, using the specific activity of CS determined for the same muscle, carried out the following conversion:
21 21 21 21 21
(nmol O min2 ?CS U )CS U g 5nmol O g2 min
where the first factor is mitochondrial oxygen uptake expressed per unit CS in the mitochondrial preparation and the second factor is the CS activity in the muscle homogenates.
2.6.2. Protein concentrations
Mitochondrial protein concentrations were determined using the bicinchoninic acid method of Smith et al. (1985) with BSA as the standard. The concentration of BSA in the resuspension medium was subtracted to establish the concentration of mitochondrial protein.
2.7. Chemicals
All biochemicals were from Boehringer Mannheim Co. (Montreal, Canada) or Sigma Chemical Co. (St. Louis, MO). All other chemicals were analytical grade.
2.8. Statistical analysis
Data were analyzed using a one-way ANOVA to test the null hypotheses of no differences between reproductive stages (Sokal and Rohlf, 1981). Normality was tested using a Shapiro–Wilk’s test (SAS, 1991) and homogeneity of variances using a Levene test (Snedecor and Cochran, 1989). Multiple pairwise comparisons (Tukey) were used to test for specific differences when the ANOVAs indicated significant differences (SAS, 1991). Comparisons between the proportions of scallops, for each reproductive stage, that responded to restimulation 2 min after opening their valves were made using
2
x -tests (Sokal and Rohlf, 1981).
3. Results
3.1. Reproductive stages
with animals sampled in mid-July and early August (Fig. 1). The scallops sampled in mid-July and early August did not differ in their gonadal indices or in the lipid and protein contents of the gonads. As the latter sample was taken only one week before spawning began, we identified the groups sampled in mid-July and those sampled in early August as mature and prespawned scallops, respectively. The scallops sampled in mid-August showed a marked decrease of the gonadal index and of the lipid and protein contents in their gonads, indicating that animals had spawned (Fig. 1). Our classification of these reproductive stages was supported by histological observations of gonadal sections from the scallops sampled in the four periods.
3.2. Environmental conditions
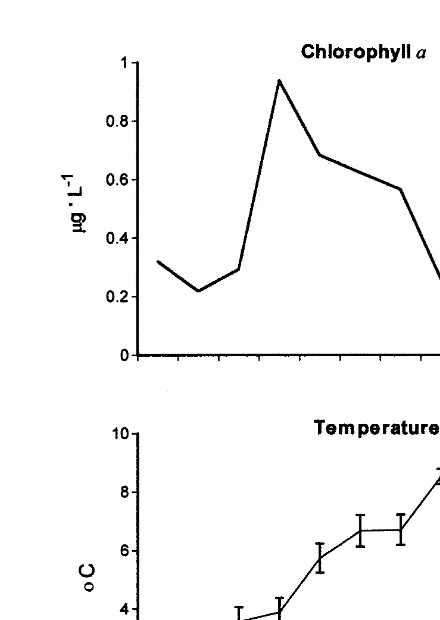
Phytoplankton abundance fluctuated considerably during the study. In general, during
21
maturation scallops had more phytoplankton available (|0.5 mg l of chlorophyll a)
than before and after spawning, where phytoplankton availability decreased (|0.3
21
mg l , Fig. 2).
During gonadal maturation, bottom temperature was higher and more variable than during spawning (Fig. 2). During July and until early August, temperature fluctuated around 68C. During spawning, it dropped to |48C and remained low until the end of
August.
3.3. Escape response
When Chlamys islandica were stimulated with the seastar Leptasterias polaris, they responded with a series of|26 claps in a period of |2 min. There was no difference in
the mean number of claps performed by the scallops at different reproductive stages
21
(Table 1). The clapping rate of 13 claps min remained relatively stable, except for
21
prespawning scallops for which the rate was 17 claps min (Table 1).
The clapping response was followed by a period of |30 min where the animals
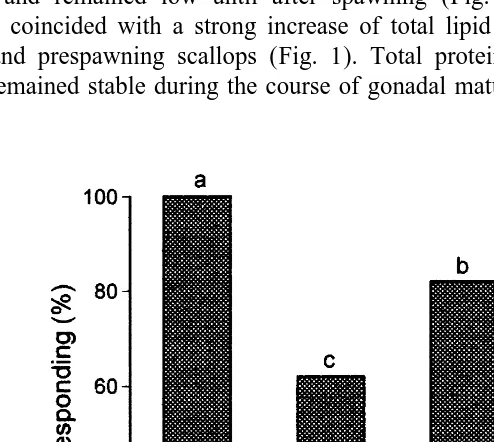
remained closed and refractory to further stimulation. This period was similar between reproductive stages, with the exception of mature animals which spent less time closed (Table 1). To evaluate the extent of recovery during valve closure, scallops were restimulated 2 min after they reopened their valves, to measure the proportion of initial claps performed during restimulation and the proportion of animals that responded to stimulation. Spawned scallops could only do 11% of the initial claps, compared to the other reproductive stages which responded with 25–35% of their initial claps (Table 2). Interestingly, all immature animals responded after restimulation with the seastar, whereas after gonadal maturation only 62% of mature and 82% of prespawning animals responded (Fig. 3). Spawned animals markedly decreased their responsiveness as only 38% reacted 2 min after reopening their valves.
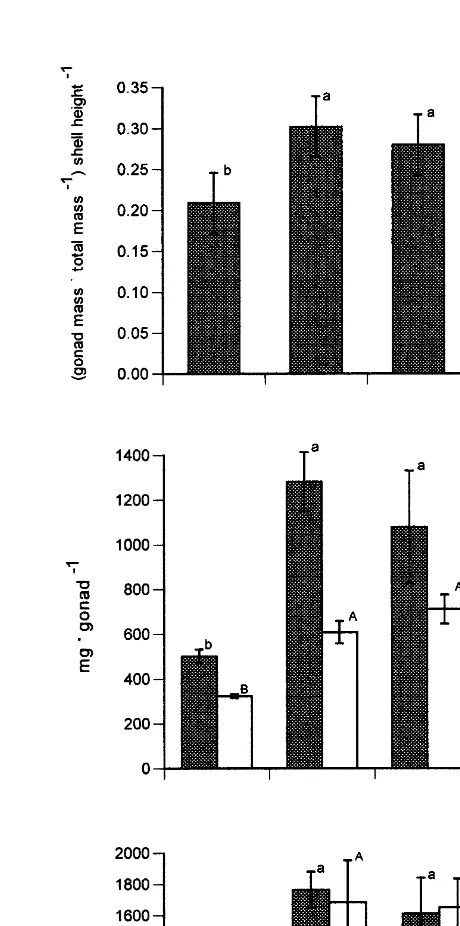
Fig. 1. Gonadosomatic index and total content of lipid and protein in gonads of female and male Chlamys
21 ˆ
Fig. 2. Variations in temperature (8C) and chlorophyll a (mg l ) 1 m above the Chlamys islandica bed, at Ile ˆ
du Fantome Mingan Islands from May to August 1996. Values represent means6S.E. Arrows indicates the times when scallops were sampled. P-S5prespawned.
C. islandica did not change its clapping capacity, but markedly slowed its rate of recuperation from exhausting escape responses. Both the recuperation during valve closure and that occurring once valves reopened were diminished.
Table 1
Mean number (S.E.) of claps, clapping rate and time spent with the valves closed after exhaustive exercise for the scallop Chlamys islandica at different reproductive stages
Reproductive Number Clapping rate Time spent n
21
stage of claps (claps min ) closed (min)
Immature 26 (1) 13 (1) 33.2 (2.7) 40
Mature 25 (1) 13 (1) 23.2 (1.7)** 48
Pre-spawned 30 (2) 17 (1)** 34.1 (3.4) 29
Spawned 24 (1) 13 (1) 28.8 (3.3) 48
Table 2
Mean (S.E.) proportion (%) of initial claps recovered at different time intervals after exhausting escape response for the scallop Chlamys islandica at different stages in gonadal development
Recuperation time Immature Mature Prespawned Spawned
with valves open
a
2 min 34.8 (2.7) 24.6 (3.6) 28.1 (6.7) 10.6 (5.8)*
2 h 73.3 (5.5) 54.2 (7.1) 72.7 (3.4) 69.5 (11.5)
4 h 95.2 (6.3)** 60.5 (6.3) 60.9 (9.8) 70.1 (8.9)
6 h 103.0 (4.1) 63.0 (7.0) 78.1 (6.2) 70.8 (4.8)
12 h 50.0 (10.7) 102.0 (3.9)** 81.9 (5.5)
18 h 100.6 (4.7)** 105.2 (8.8)**
a
Recovery efficiency during valve closure.
* Comparisons between reproductive stages for 2 min (P,0.05); ** Statistically different from the earlier test (P,0.05), indicate that recovery was completed.
3.4. Biochemical composition of muscle at different reproductive stages
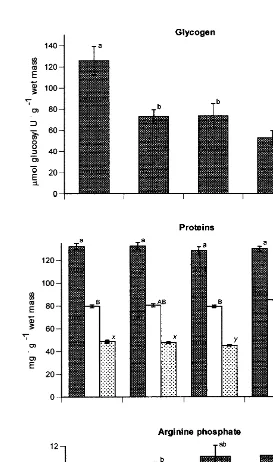
The concentration of glycogen in the phasic muscle decreased markedly as the gonad matured and remained low until after spawning (Fig. 4). The decrease in muscle glycogen coincided with a strong increase of total lipid and protein in the gonads of mature and prespawning scallops (Fig. 1). Total protein concentration in the phasic muscle remained stable during the course of gonadal maturation and spawning (Fig. 4).
Fig. 3. Proportion of Chlamys islandica at different reproductive stages that responded to restimulation with the seastar, 2 min after the end of glycolytic recuperation (closed valves) (n58). Bars sharing the same letters
2
21
Fig. 4. Concentration of glycogen (mmol glucosyl U g wet mass), total, structural and sarcoplasmic proteins
21 21
However, structural protein oscillated and the level of sarcoplasmic protein fell slightly before spawning. Muscle arginine phosphate gradually increased during gonadal maturation and was highest before and after spawning (Fig. 4).
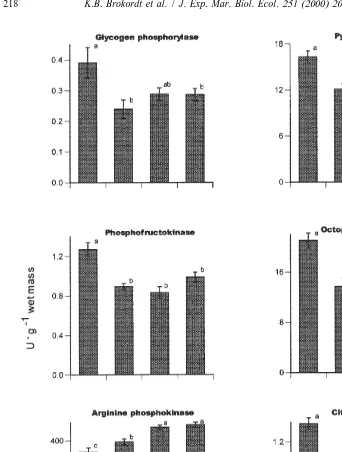
3.5. Muscle enzyme levels at different reproductive stages
With the exception of AK, the enzymes showed similar fluctuations in activity during gonadal maturation and spawning (Fig. 5). The glycolytic enzymes, GP, PFK, PK and ODH, and the mitochondrial enzyme, CS, showed their highest levels in immature scallops and levels declined with gonadal maturation. PK recovered its initial levels before spawning, but showed its lowest level after spawning. Much like the levels of arginine phosphate, AK activity increased during the course of gonadal maturation, reaching a maximum before spawning (Fig. 5).
Thus, the levels of enzymes that participate in the recovery of the adductor muscle from exhausting escape responses decreased during gonadal maturation and spawning. In contrast, the enzyme which generates most of the ATP required for the escape response gradually increased its activity during gonadal maturation and spawning.
3.6. Oxidative capacities of mitochondria and adductor muscle during the
reproductive cycle
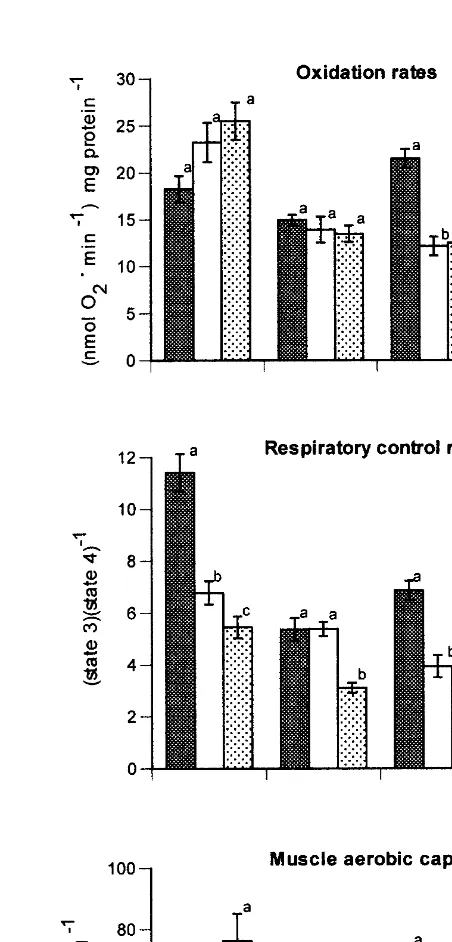
Rates of mitochondrial oxygen uptake and respiratory coupling varied with substrate and reproductive stage (Fig. 6). Glutamate and succinate gave highest oxidation rates at most times. Rates of glutamate and malate oxidation changed little during the reproductive cycle. In contrast, mitochondrial rates of pyruvate and succinate oxidation decreased considerably after gonadal maturation and remained low after spawning (Fig. 6). For all substrates, the respiratory control ratio (RCR) decreased during the reproductive cycle (Fig. 6). The RCR values for glutamate and pyruvate oxidation were lower in mitochondria from mature and spawned scallops, whereas those for the oxidation of malate and succinate declined only after spawning.
Total muscle oxidative capacities were calculated from mitochondrial rates of substrate oxidation, mitochondrial CS levels and muscle CS levels. Reproductive status markedly affected the muscle’s capacity for oxidizing pyruvate and succinate, with decreases occurring with maturation and spawning (Fig. 6). As pyruvate generated from octopine is metabolized during recuperation, the total capacity for pyruvate oxidation is most pertinent to evaluation of the capacity for recuperation from exhausting escape responses. Overall, the mitochondrial oxidative capacities, total muscle aerobic capacity and activities of enzymes involved in glycolytic and aerobic phases of recovery followed virtually the same pattern, a decrease in capacity with maturation and spawning.
4. Discussion
21
Fig. 5. Enzyme activities (U g wet mass) in the phasic adductor muscle of the scallop Chlamys islandica at different reproductive stages. Values represent means6S.E. (n57–12). Means sharing the same letters were not significantly different (P,0.01) as determined by Tukey multiple comparisons.
21 21
Fig. 6. Maximal oxidation rates (nmol O min2 mg mitochondrial protein ), respiratory control ratios (state 21 21
adductor muscle and in the organismal capacity to recover from exhausting escape responses. Both gonadal maturation and spawning decreased these parameters.
The decrease of the recovery efficiency with gonadal maturation and spawning occurred both during valve closure and after valves had reopened. The time scallops spent with their valves closed after exhausting exercise was not markedly affected by the reproductive stage. Only mature scallops spent less time closed. Given that prolonged valve closure is powered exclusively by the tonic (catch) part of the adductor muscle (de Zwaan et al., 1980), the time scallops spent closed could be related to the metabolic capacity of this section. However, the energetic cost of valve closure is only 1% of that of valve claps (de Zwaan et al., 1980). Therefore, the metabolic capacities of the tonic muscle should not strongly affect its capacity to maintain the valves closed and limitations may arise from the requirements of other tissues including the phasic adductor muscle. Since we did not measure the energetic status and metabolic capacities of the tonic muscle, studies of this muscle are required to confirm this interpretation.
The extent of recovery during valve closure could have been affected by its duration. However, spawned scallops showed the least recovery, although they spent the same time closed as immature and prespawned animals, which had the highest percentage of recovery (Table 2, Fig. 3). The negative impact of gonadal maturation and spawning on recovery during valve closure may be due to the lower levels of the glycolytic enzymes. Reliance upon glycolysis is likely since, in analogy with the responses of P.
magel-lanicus, the scallops probably consumed most of their oxygen reserves during valve
clapping and took up no oxygen during valve closure (Thompson et al., 1980). GP and PFK are important sites of metabolic control in the adductor muscle of Argopecten
irradians concentricus (Chih and Ellington, 1986) and ODH is used during the
restoration of ATP and arginine phosphate pools when oxygen supplies are depleted or absent as during valve closure (Livingstone et al., 1981). The fact that the activities of these enzymes are diminished after gametogenesis and spawning in C. islandica, could explain the reductions in glycolytic recovery.
In Placopecten magellanicus, the complete restoration of the phosphoarginine pool after exhaustive swimming requires aerobic metabolism (Livingstone et al., 1981). For the phasic muscle of C. islandica, the levels of CS, the mitochondrial capacity (and RCR values) for pyruvate oxidation and the total muscle aerobic capacity decreased after gonadal maturation and remained low after spawning. These reductions could explain why mature and spawned scallops needed 14 h more to recover their initial clapping capacity than immature animals (Table 2). In the adductor muscle of the tropical scallop,
Euvola ziczac, mitochondrial capacities for pyruvate oxidation were lower during the
first spawning, which occurred after a period of low food availability, than during other periods (Boadas et al., 1997). As pyruvate is the principal substrate to be oxidized after muscular activity (Chih and Ellington, 1986), a reduced capacity for pyruvate oxidation could slow the aerobic recovery of mature and spawned scallops from exhausting escape responses.
animals would have been feeding at higher levels than immature or prespawning animals, thereby reducing the aerobic scope for recuperation. The metabolic costs of gonadal maturation would have exacerbated this effect. In accordance with this interpretation, prespawning scallops partially regained the capacity to recover from exercise, compared to mature animals (Table 2, Fig. 3). However, spawning decreased the percentage of aerobic recuperation even more than gonadal maturation (Table 2, Fig. 3). As increased metabolic rates due to feeding and gonadal maturation would not explain this decrease, reductions in muscle metabolic capacity are a more likely explanation for the slowed rates of recuperation. In Argopecten irradians concentricus spawning leads to a state of negative energy balance and generally poor physiological condition, as indicated by a loss of carbon 14 from all body components (Barber and Blake, 1985). Assays of oxygen uptake at rest and during aerobic recovery would ascertain the extent to which gonadal maturation and feeding reduce the aerobic scope for recuperation.
Changes in the composition of body components should indicate which biochemical components contribute to energy metabolism during gametogenesis (Barber and Blake, 1991). In C. islandica, even though food availability was high during the accumulation of gonadal lipid and protein (Fig. 2), muscle glycogen decreased strongly while muscle protein changed little (Figs. 1 and 4). Therefore, muscle glycogen is likely a major precursor for lipid and protein synthesis in the gonad. This agrees with patterns found for C. islandica in Norway (Sundet and Vahl, 1981; Sundet and Lee, 1984); for Chlamys
varia (Shafee, 1981) and Placopecten magellanicus (Robinson et al., 1981). However,
in some scallops, such as Argopecten irradians concentricus, muscle protein is also used to support gonadal maturation after the depletion of glycogen (Barber and Blake, 1981). In this subspecies, physiological indices, such as O / NH3 and CO / O2 2 (respiratory quotient), indicate that gametogenesis is initially supported by lipid. Then, during the cytoplasmic growth of oocytes, gametogenesis is supported by muscle glycogen, which is converted to lipid. Finally, near and after spawning, metabolism is fueled by protein (Barber and Blake, 1985). In contrast, in A. irradians irradians muscle carbohydrates make a negligible contribution to gonadal development, rather gametogenesis occurs mainly at the expense of adductor muscle protein, which accounts for 63–99% of gonadal buildup in the spring (Epp et al., 1988). Total protein in the adductor muscle of
C. islandica did not change during gonadal maturation and spawning (Fig. 4). As gonadal maturation is completed during spring and summer, when chlorophyll a concentrations are the highest (Spence and Steven, 1974; Arsenault and Himmelman, 1998), external food supplies may decrease the need to mobilize muscle protein.
salmon, Oncorhynchus nerka, muscle enzymes show different rates of catabolism than the general categories of soluble and insoluble proteins (Mommsen et al., 1980). Whereas we do not know why scallops sacrifice the enzymes that facilitate recuperation of muscle performance, we suggest this may be a consequence of glycogen mobilization. It has been suggested that glycolytic enzymes are attached to glycogen particles (Hubbard and Cohen, 1989; Lam, 1990; Vardanis, 1990). In skeletal muscle of rabbit, GP accumulates concomitantly with glycogen (Francois et al., 1992). Thus, a decrease in glycogen levels could reduce binding sites for glycolytic enzymes. Once enzymes are in solution in the cytoplasm, they could be more susceptible to degradation. The concomitant decline of glycogen and glycolytic enzymes with gonadal maturation supports such a mechanism.
Contrary to our initial premise, arginine phosphate, the principal fuel for muscle contraction during the escape response, increased after gonadal maturation and after spawning (Fig. 4). Although the arginine phosphate levels we measured are lower than those previously obtained (possibly as our technique for sampling the muscle was slower than that of other workers), the relative levels should be reliable, as we applied the same sampling technique to each reproductive stage. Moreover, AK, the enzyme that converts arginine phosphate to ATP and arginine, also increased after gametogenesis and its levels remained high after spawning (Fig. 5). These results may partly explain why the number of claps did not change during reproduction of C. islandica (Table 1).
The fact that mature and spawned scallops need more time to recuperate after exhausting burst exercise may have ecological significance. After gonadal maturation and spawning, scallops could be more susceptible to predation and to capture by commercial fisheries. This is particularly true since many scallop species are captured by trawling or dragging (Joll, 1989; C. islandica in the northern Gulf of St. Lawrence), where their vulnerability to capture will depend on their capacity to escape and recuperate. This is particularly true since trawls or drags may be used repeatedly in the same area of the scallop bed.
Interindividual differences in the locomotor performance of vertebrates can be due to physiological changes or morphological variability (Garland, 1984; Walsberg et al., 1986; Bennett et al., 1989; Kolok, 1992); to size, age (Garenc et al., 1999), thermal acclimation (Elliott, 1991; Johnson and Bennett, 1995) or reproductive stage (Koch and Wieser, 1983; Calow, 1985; James and Johnston, 1998). Many of these factors modify
¨
muscle metabolic capacities (Sanger, 1993). In invertebrates, the evidence for inter-individual differences in locomotor performance is limited, nevertheless, there are studies, particularly for scallops, that show that swimming performance changes with morphology (Gould, 1971) and size or age (Winter and Hamilton, 1985; Joll, 1989; Manuel and Dadswell, 1991). Most of the differences have been attributed to hydrodynamic changes with size (Manuel and Dadswell, 1991). Our study demonstrates a marked effect of reproduction on the recuperation of clapping capacity after exhaustive exercise and on muscle metabolic capacity in scallops.
Acknowledgements
support and valuable comments on the manuscript. We also thank Martin Lafrance and Martin Giasson for technical assistance. This work was supported by operating grants from the NSERC to HEG and JHH. KBB was recipient of scholarships from the
´ ´ ´
Gouvernement du Quebec, Departement de Biologie de l’Universite Laval and GIROQ. [SS]
References
¨
Arsenault, D.J., Himmelman, J.H., 1998. Spawning of the Iceland scallop (Chlamys islandica Muller, 1776) in the northern Gulf of St. Lawrence and its relationship to temperature and phytoplankton abundance. Veliger 41, 180–185.
Ballantyne, J.S., Moon, T.W., 1985. Hepatopancreas mitochondria from Mytilus edulis: substrate preferences and effects of pH and osmolarity. Mar. Biol. 87, 239–244.
Barber, B.J., Blake, N.J., 1981. Energy storage and utilization in relation to gametogenesis in Argopecten
irradians concentricus (Say). J. Exp. Mar. Biol. Ecol. 52, 121–134.
Barber, B.J., Blake, N.J., 1985. Substrate catabolism related to reproduction in the bay scallop Argopecten
irradians concentricus, as determined by O / N and RQ physiological indexes. Mar. Biol. 87, 13–18.
Barber, B.J., Blake, N.J., 1991. Reproductive physiology. In: Shumway, S.E. (Ed.), Scallops: Biology, Ecology and Aquaculture. Elsevier, Amsterdam, pp. 377–428.
Bates, P.C., Milllard, D.J., 1983. Myofibrillar protein turnover: synthesis rates of myofibrillar and sarcoplasmic protein fractions in different muscles and the changes observed during postnatal development and in response to feeding and starvation. Biochem. J. 214, 587–592.
Beaulieu, M.A., Guderley, H., 1998. Changes in qualitative composition of white muscle with nutritional status of Atlantic cod, Gadus morhua. Comp. Biochem. Physiol. A. 121, 135–141.
Bennett, A.F., Garland, T., Else, P.L., 1989. Individual correlation of morphology, muscle mechanics and locomotion in a salamander. Am. J. Physiol. 256, R1200–R1208.
Boadas, M.A., Nusetti, O., Mundarain, F., Lodeiros, C., Guderley, H., 1997. Seasonal variation in the properties of muscle mitochondria from the tropical scallop Euvola (Pecten) ziczac. Mar. Biol. 128, 247–255.
Bonardelli, J.C., Himmelman, J.H., 1995. Examination of assumptions critical to body component indices: application to the giant scallop Placopecten magellanicus. Can. J. Fish. Aquat. Sci. 52, 2457–2469. Brand, A.R., 1991. Scallop ecology: distribution and behaviour. In: Shumway, S.E. (Ed.), Scallops: Biology,
Ecology and Aquaculture. Elsevier, Amsterdam, pp. 517–584.
Calow, P., 1985. Adaptive aspects of energy allocation. In: Tyler, P., Calow, P. (Eds.), Fish Energetics: New Perspectives. The Johns Hopkins University Press, Maryland, pp. 13–32.
Chih, P.C., Ellington, W.S., 1983. Energy metabolism during contractile activity and environmental hypoxia in the phasic adductor muscle of the bay scallop Argopecten irradians concentricus. Physiol. Zool. 56, 623–631.
Chih, P.C., Ellington, W.S., 1986. Control of glycolysis during contractile activity in the phasic adductor muscle of the bay scallop, Argopecten irradians concentricus: identification of potential sites of regulation and a consideration of the control of octopine dehydrogenase activity. Physiol. Zool. 59, 563–573. Elliott, J.M., 1991. Tolerance and resistance to thermal stress in juvenile Atlantic salmon, Salmo salar. Freshw.
Biol. 25, 61–70.
Epp, J., Bricelj, V., Malouf, R., 1988. Seasonal partitioning and utilization of energy reserves in two age classes of the bay scallop Argopecten irradians irradians. J. Exp. Mar. Biol. Ecol. 121, 113–136.
Estabrook, R.W., 1967. Mitochondrial respiratory control and the polarographic measurement of ADP:O ratios. Methods Enzymol. 10, 41–47.
Folch, J.M., Lee, S.M., Sloane-Stanley, G.H., 1957. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, 497–509.
Gagnon, R., Lehoux, E., Guderley, H., 1998. Manually operated freeze clamping press. Anal. Biochem. 260, 251–252.
Garenc, C., Couture, P., Laflamme, M.A., Guderley, H., 1999. Metabolic correlates of burst swimming capacity of juvenile and adult threespine stickleback (Gasterosteus aculeatus). J. Comp. Physiol. B 169, 113–122.
Garland, Jr. T., 1984. Physiological correlates of locomotory performance in a lizard: an allometric approach. Am. J. Physiol. 247, R806–R815.
Gould, S.J., 1971. Muscular mechanics and the ontogeny of swimming in scallops. Paleontology 14, 61–94. Graham, M., 1987. The solubility of oxygen in physiological salines. Fish Physiol. Biochem. 4, 1–4. Guderley, H.E., Rojas, F.M., Nusetti, O.A., 1995. Metabolic specialization of mitochondria from scallop phasic
muscle. Mar. Biol. 122, 409–416.
Himmelman, J.H., 1991. Diving observations of subtidal communities in the northern Gulf of St. Lawrence. In: Therriault, J.C. (Ed.), The Gulf of St. Lawrence Small Ocean or Big Estuary. Can. Spec. Publ. Fish. Aquat. Sci, Vol. 113, pp. 319–332.
Hubbard, M.J., Cohen, P., 1989. Regulation of protein phosphatase 1-G from rabbit skeletal muscle: 1. Phosphorylation by cyclic AMP-dependent protein kinase at site 2 releases catalytic subunit from glycogen-bound holoenzyme. Eur. J. Biochem. 186, 701–710.
James, R.S., Johnston, I.A., 1998. Influence of spawning on swimming performance and muscle contractile properties in the short-horn sculpin. J. Fish Biol. 53, 485–501.
Johnson, T.P., Bennett, A.F., 1995. The thermal acclimation of burst escape performance in fish: an integrated study of molecular and cellular physiology and organismal performance. J. Exp. Biol. 198, 2165–2175. Joll, L.M., 1989. Swimming behaviour of the saucer scallop Amusium balloti (Mollusca: Pectinidae). Mar.
Biol. 102, 299–305.
Keppler, D., Decker, K., 1974. Glycogen determination with amyloglucosidase. In: Bergmeyer, H.U. (Ed.). Methods of Enzymatic Analysis, Vol. 3. Academic Press, New York, pp. 1127–1131.
Koch, F., Wieser, W., 1983. Partitioning of energy in fish: can reduction of swimming activity compensate for the cost of production? J. Exp. Biol. 107, 141–146.
Kolok, A.S., 1992. Morphological and physiological correlates with swimming performance in juvenile largemouth bass. Am. J. Physiol. 263, R1042–R1048.
Lam, R., 1990. The nature of cytoplasmic vacuoles in chordoma cells: a correlative enzyme and electron microscopic histochemical study. Pathol. Res. Pract. 186, 642–650.
Lamprecht, W., Trautschold, I., 1974. Adenosine triphosphate. Determination with hexokinase and glucose-6-phosphate dehydrogenase. In: Bergmeyer, H.U. (Ed.). Methods of Enzymatic Analysis, Vol. 4. Academic Press, New York, pp. 2101–2110.
Lamprecht, W., Stein, P., Wieser, H., 1974. Creatine phosphate. In: Bergmeyer, H.U. (Ed.). Methods of Enzymatic Analysis, Vol. 4. Academic Press, New York, pp. 1777–1781.
Livingstone, D.R., de Zwaan, A., Thompson, R.J., 1981. Aerobic metabolism, octopine production and phosphoarginine as sources of energy in the phasic and catch adductor muscles of the giant scallop
Placopecten magellanicus during swimming and the subsequent recovery period. Comp. Biochem. Physiol.
70B, 35–44.
Manuel, J.L., Dadswell, M.L., 1991. Swimming behaviour of juvenile giant scallop Placopecten magellanicus, in relation to size and temperature. Can. J. Zool. 69, 2250–2254.
´
Martınez, G., 1991. Seasonal variation in biochemical composition of three size classes of the Chilean scallop,
Argopecten purpuratus Lamarck, 1819. Veliger 34, 335–343.
´
Martınez, G., Mettifogo, L., 1998. Mobilization of energy from adductor muscle for gametogenesis of the scallop, Argopecten purpuratus Lamarck. J. Shellfish Res. 17, 113–116.
Mommsen, T.P., French, C.J., Hochachka, P.W., 1980. Sites and patterns of protein and amino acid utilization during the spawning migration of salmon. Can. J. Zool. 58, 1785–1799.
Olin, T., von der Decken, A., 1987. Estrogen treatment and its implication on vitellogenin and myosin synthesis in salmon (Salmo salar). Physiol. Zool. 60, 346–351.
Olson, J.M., Marsh, R.L., 1993. Contractile properties of the striated adductor muscle in the bay scallop
Argopecten irradians at several temperatures. J. Exp. Biol. 176, 175–193.
Robinson, W.E., Wehling, W.E., Morse, M.P., McLeod, G.C., 1981. Seasonal changes in soft-body component indices and energy reserves in the Atlantic deep-sea scallop, Placopecten magellanicus. Fish. Bull. 79, 449–458.
¨
Sanger, A.M., 1993. Limits to the acclimation of fish muscle. Rev. Fish Biol. Fish. 3, 1–15.
SAS, 1991. In: SAS / STAT User’s Guide, release 6.03 Edition. SAS Institute Press, Cary, NC, p. 1028. Shafee, M.S., 1981. Seasonal changes in the biochemical composition and calorific content of the black scallop
Chlamys varia (L.) from Lanvec, Bay of Brest. Oceanol. Acta 4, 331–341.
Shevechenko, V.V., 1972. The dynamics of protein and fat content in the organs and tissues of the north sea haddock Melanogrammus aeglefinus in the process of seasonal growth and gonad maturation. J. Ichthyol. 12, 830–842.
Shumway, S.E., Barter, J., Stahlnecker, J., 1988. Seasonal changes in oxygen consumption of the giant scallop,
Placopecten magellanicus (Gmelin). J. Shellfish Res. 7, 77–82.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Matha, A.K., Garther, F.H., Provenzano, M.D., Fujimoto, E.K., Goeke, N.M., Olson, B.J., Klenk, D.C., 1985. Measurements of protein using bicinchoninic acid. Anal. Biochem. 150, 76–85.
Snedecor, G.W., Cochran, W.G. (Eds.), 1989. Statistical Methods. Iowa State University Press, Ames, Iowa, p. 503.
Sokal, R.R., Rohlf, F.J. (Eds.), 1981. Biometry. W.H. Freeman, New York, p. 859.
Somero, G.N., Childress, J.J., 1990. Scaling of ATP-supplying enzymes, myofibrillar proteins and buffering capacity in fish muscle: relationship to locomotory habit. J. Exp. Biol. 149, 319–333.
Spence, C., Steven, D.M., 1974. Seasonal variation of the chlorophyll a: pheopigment ratio in the Gulf of St. Lawrence. J. Fish. Res. Bd Can. 31, 1263–1268.
Sundet, J.H., Vahl, O., 1981. Seasonal changes in dry weight and biochemical composition of the tissues of sexually mature and immature Iceland scallops Chlamys islandica. J. Mar. Biol. Assoc. UK 61, 1001–1010. Sundet, J.H., Lee, J.B., 1984. Seasonal variations in gamete development in the Iceland scallop Chlamys
islandica. J. Mar. Biol. Assoc. UK 64, 411–416.
Thompson, R.J., Livingstone, D.R., de Zwaan, A., 1980. Physiological and biochemical aspects of valve snap and valve closure responses in the giant scallop Placopecten magellanicus. I. Physiology. J. Comp. Physiol. 137, 97–104.
´ ¨ ¨
Thorarinsdottir, G.G., 1993. The Iceland scallop, Chlamys islandica (O.F. Muller), in Breidafjodur, west Iceland. II. Gamete development and spawning. Aquaculture 110, 87–96.
Vardanis, A., 1990. Fractionation of particulate glycogen and bound enzymes using high-performance liquid chromatography. Anal. Biochem. 187, 115–119.
Von der Decken, A., 1992. Physiological changes in skeletal muscle by maturation-spawning of non-migrating female Atlantic salmon, Salmo salar. Comp. Biochem. Physiol. 101B, 299–301.
Walsberg, G.E., Lean, M.S., Hillman, S.S., 1986. Individual variation in maximum aerobic capacity: cardiovascular and enzymatic correlates in Rana catesbiana. J. Exp. Zool. 239, 1–5.
Winter, M.A., Hamilton, P.V., 1985. Factors influencing swimming in bay scallops, Argopecten irradians (Lamarck, 1819). J. Exp. Mar. Biol. Ecol. 88, 227–242.