www.elsevier.nlrlocateraqua-online
The influence of nocturnal vs. diurnal feeding under
winter conditions on growth and feed conversion of
ž
/
European sea bass Dicentrarchus labrax, L.
M. Azzaydi
), F.J. Martınez, S. Zamora, F.J. Sanchez-Vazquez,
´
´
´
J.A. Madrid
Department of Physiology and Pharmacology, Faculty of Biology, UniÕersity of Murcia, 30100 Murcia, Spain
Accepted 23 July 1999
Abstract
European sea bass display predominantly nocturnal self-feeding patterns in winter and early spring, and this has been further examined by comparing the effect of night and day feeding on growth and feed conversion. To this end, three feeding regimes were compared: nocturnal
Ž . Ž . Ž .
automatic-feeding NF , diurnal automatic-feeding DF and free access to self-feeders SF .
Ž
Under NF, feed was supplied in two meals differing in size pre-dawn and post-dusk, 33.33:
.
66.67% of daily feed ration, respectively . In DF feed was supplied in three equally-sized meals
Žmorning, afternoon and evening, 33.33: 33.33: 33.33% of daily feed ration . The experiment was.
carried out from January to April. Under SF, sea bass showed a phase inversion of daily feeding rhythms, which changed from an initial nocturnal to diurnal by the end of the experiment. Feeding
Ž . Ž .
strategies affected specific growth rate SGR and feed conversion ratio FCR , the highest SGR
Ž
and the lowest FCR being obtained with SF and NF treatments SGR: 0.26"0.01 in NF vs.
.
0.19"0.01 in DF; FCR: 2.65"0.08 in NF vs. 3.73"0.17 in DF . The results demonstrate that
feeding sea bass by night in winter, when they show nocturnal behaviour, may improve their growth and feed efficiency.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Feeding rhythms; Growth; Feed conversion; Self-feeding; Automatic-feeding; Sea bass
1. Introduction
Knowledge about fish feeding habits is of importance in fish farming, because feeding regime may have consequences for both growth efficiency and feed wastage.
)Corresponding author. Tel.:q34-968-364931; fax:q34-968-363963; e-mail: [email protected] 0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
Intensively farmed fish are often fed regularly throughout the day on predetermined
Ž
daily feed rations, but fish display daily cycles of feeding Kadri et al., 1991; Alanara,
¨ ¨
1992a,b; Boujard and Leatherland, 1992a,b; Eriksson and Alanara, 1992; Boujard, 1995;¨ ¨
.
Jobling et al., 1995 , so this feeding practice may result in under-feeding when the fish are hungry and in over-feeding at times of low appetite.
The matching of feed supply to natural feeding rhythms may lead to improved growth
ŽNoeske et al., 1981; Sundararaj et al., 1982; Noeske and Spieler, 1984; Noeske-Hallin et al., 1985; Kerdchuen and Legendre, 1991; Boujard et al., 1995; Gelineau et al., 1996;
´
.
Azzaydi et al., 1998, 1999 , but feeding preferences may change with the seasons. Consequently, feeding time which results in best growth does not necessarily remain
Ž
unchanged throughout the year Landless, 1976; Noeske and Spieler, 1984; Boujard and
.
Leatherland, 1992a; Fraser et al., 1993; Smith et al., 1993 .
Sea bass, which are widely cultivated in the Mediterranean region, displays flexibility
Ž
in its feeding rhythms Anthouard et al., 1993; Sanchez-Vazquez et al., 1994, 1995;
´
´
.
Boujard et al., 1996 . Recently, it has been reported that sea bass show phase inversion in their feeding rhythms on a seasonal basis: fish which were diurnal in summer and autumn, changed to nocturnal in winter, and returned to being diurnal in later spring
ŽSanchez-Vazquez et al., 1998 .
´
´
.In the present paper we tested whether feeding sea bass by night in winter improves their growth performance. To this end, we compared the growth and feed conversion of sea bass fed by automatic systems which either mimicked the natural winter feeding
Ž .
rhythm nocturnal feeding, NF or in which feed was supplied as three equally
Ž .
sized-meals provided during daylight hours diurnal feeding, DF . A third treatment in
Ž .
which fish had free access to a self-feeder SF was used to study self-feeding patterns.
2. Materials and methods
Ž .
A total of 180 European sea bass Dicentrarchus labrax, L. hatched in captivity
ŽCULMAREX, Aguilas, Murcia, Spain , were used in the experiment which was carried.
out from January to April. The fish, which had an initial body weight of 59.3"1.2 g
Žmean"SEM , were divided into 12 groups with 15 fish in each. Each group was.
Ž .
placed into a 460 1 experimental tank supplied with running seawater 37‰ salinity . Throughout the experiment, the fish were exposed to natural photoperiod and water
Ž . Ž .
temperature, which ranged from 10.25 h January to 12.50 h April , and from 14.18C
ŽJanuary to 16.8. 8C AprilŽ . ŽFig. 2 . Photoperiod light level and water temperature. Ž . Ž
were continuously registered every 15 min by means of a portable data logger Logit SL,
.
DCP Microdevelopments and SCC Research, UK .
Ž .
The fish were fed a commercial pellet feed EWOS, 2 mm pellet containing 49% protein, 12% fat, 6.8% moisture, 11% ash and 21.2% NFE. Each tank was provided with
Ž .
an electronic self-feeder which delivered approximately 8–9 pellets about 0.4 g of feed
Ž
each time a fish activated a trigger located 3 cm below the water surface Sanchez-
´
.
Vazquez et al., 1994 . Feeders were checked every other day, the feed remaining was
´
weighed and the feed reservoir refilled.All groups of fish were initially given free access to the self-feeders for 33 days. Fish
Ž
. Ž .
treatment . In the first treatment, sea bass had free access to self-feeders SF treatment connected to a computer which recorded the number of trigger activations in 10 min bins. In the second and the third treatments, sea bass were fed using
automatically-tri-Ž .
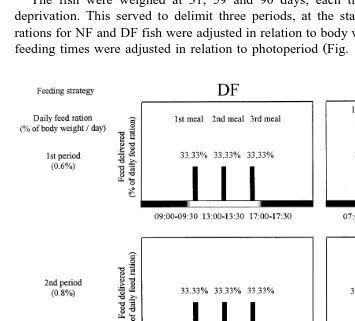
ggered feeders according to fixed schedules Fig. 1: NF and DF . NF fish were fed in
Ž .
accordance with winter self-feeding rhythms Sanchez-Vazquez et al., 1998 whereas
´
´
DF fish were fed three equally-sized meals. An electronic feeder activated automatically by a timer was used, there being a pause between each feed delivery. In the NF and DF treatments, the daily rations were established in accordance with tables provided by theŽ .
feed manufacturer EWOS .
The fish were weighed at 31, 59 and 90 days, each time after one day of feed deprivation. This served to delimit three periods, at the start of which the daily feed rations for NF and DF fish were adjusted in relation to body weight and temperature and
Ž .
feeding times were adjusted in relation to photoperiod Fig. 1 .
Table 1
Effect of feeding regime on growth performance
Ž . Ž . Ž .
Means with different letters in the same line in the same row for CV are significantly different ANOVA,
.
Tukey HSD, mean"SEM .
The following parameters were calculated for each period and for the duration of the
Ž . wŽ Ž . Ž
entire experiment: Specific growth rate SGR s In final biomass yIn initial
.. x Ž .
biomass rdays =100 Feed conversion ratio FCR sweight feed deliveredrbiomass
Ž . Ž
gain Initial and final Coefficient of Variation CVŽi or f. s standard deviation of the
.
individualŽi or f.weightrmeanŽi or f.weight =100
Ž
Treatment means were compared by one-way ANOVA STATGRAPHICS 7.0
.
Statistical Graphics, Cambridge, MA, 1993 after log-transformation of indices and percentages. When statistically significant differences were detected, Tukey’s multiple range test was used a posteriori. CV and CV were compared by a sign-test. In all tests,i i the significance level was set at P-0.05.
3. Results
Actograms of feeding activity of SF sea bass along with temperature and photoperiod are shown in Fig. 2. Group A was nocturnal during the two first months of the experiment, before changing to diurnal. Group B changed from being nocturnal to diurnal at the end of the first month of the experiment. Group C was mainly diurnal, although during the first month of the experiment fish fed both in the photophase and the scotophase. The results obtained from one group of SF fish and one of DF fish were deleted because of unsolved technical problems with the self-feeding device.
Feed, demanded by SF fish and supplied to NF and DF fish, together with biomass increase, SGR, FCR and CV are shown in Table 1. Feed demanded by SF fish was significantly lower than that supplied to NF and DF fish.
Biomass gain was significantly greater in the NF than in the DF treatment and the poorest SGR and FCR were recorded for DF fish. CV was calculated to ascertain the influence of feeding regime on the range of fish weights. In all treatments, CV decreased during the course of the experiment.
Ž .
Fig. 2. Actograms of self-feeding records of 3 groups of 15 sea bass groups A, B and C . For convenient
Ž .
visualization, the data have been double plotted 48 h time scale , each point representing the percentage of
Ž .
daily requests for feed in 10 min bins. The evolution of mean daily temperature solid line and photoperiod
Ž .
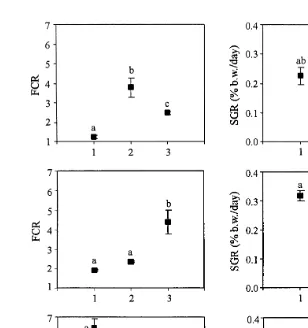
Fig. 3. Evolution of SGR and FCR during the three experimental periods in sea bass fed with the three feeding
Ž . Ž .
regimes. SF: sea bass having free access to the self-feeder Ns3 ; NF: nocturnal automatic-feeding Ns4 ;
Ž . Ž
DF: diurnal automatic-feeding Ns3 . Means with different letters are significantly different ANOVA,
.
Tukey HSD, P-0.05, mean"SEM .
Changes in SGR and FCR through the three periods of the experiment are shown in Fig. 3. In the DF treatment, the poorest FCR and SGR were obtained in the first period, while during the second and third period, FCR decreased and SGR increased. In the contrary, in the NF treatment, FCR and SGR were better in the first and second period than in the third period. Under SF treatment, no such a tendency was detected across the different experimental periods, although FCR increased in the second period, coinciding with the time of most phase inversions.
4. Discussion
ŽFig. 2 . These results agree with those reported previously for self-feeding sea bass. ŽAnthouard et al., 1993; Begout-Anras, 1995; Boujard et al., 1996; Madrid et al., 1997;
´
.
Azzaydi et al., 1998, 1999; Sanchez-Vazquez et al., 1998 and point to a seasonal
´
´
Ž .
change in feeding behaviour involving phase inversion Sanchez-Vazquez et al., 1998 .
´
´
One group of fish in the SF treatment which did not show well defined dual behaviour, something which is at odds with previous observations, where all groups ofŽ
sea bass showed a phase inversion in their daily feeding patterns Sanchez-Vazquez et
´
´
.
al., 1998 . However, not all groups of fish can be expected to display simultaneous changes in feeding behaviour.
The lowest biomass increase, SGR and poorest FCR were observed among fish in the DF treatment. Nevertheless, the evolution of growth performance during the experiment deserves detailed comment. Our results of FCR in treatment DF are in agreement with
Ž .
those described by Barnabe 1986 under winter environmental conditions. However, the
´
SGR and FCR seemed worse than those previously reported for sea bass of similar sizeŽ
and under high temperature Divanach et al., 1993; Boujard et al., 1996; Azzaydi et al.,
. Ž .
1998a or, of larger size and under the same temperature Anthouard et al., 1993 . Although comparisons between experiments must be made with caution, these
discrep-Ž .
ancies may be due to the following reasons: a the temperature conditions used in this
Ž
trial, which were far from the ‘‘optimum’’ estimated for feeding sea bass Barnabe,
´
. Ž .
1986 ; b the feed losses due to reward level for each trigger activation, used in our
Ž .
experiment 8–9 pellets for 15 fish , which probably exceeded the consumption capacity
Ž .
of sea bass due to low feeding motivation. For instance, Alanara 1996 reported a
¨ ¨
strong relationship between the level of reward and temperature conditions in rainbow trout.Fish of the SF group grew better than those of the DF group, which is in agreement
Ž .
with previous findings for sea bream Divanach et al., 1986; Kentouri et al., 1993 ,
Ž . Ž .
rainbow trout Alanara, 1992a and sea bass Azzaydi et al., 1998, 1999 : feeding fish in
¨ ¨
Ž
phase with their natural feeding rhythms may improve growth performance see reviews
.
Boujard and Leatherland, 1992a; Jobling et al., 1995; Thorpe and Cho, 1995 , a view corroborated by our results, in which SF and NF fish showed similar growth and feed conversion.
Ž .
Although growth was better in sea bass fed by night NF treatment than by day
ŽDF , the poor overall FCR and SGR Table 1 observed for fish in the DF treatment. Ž .
Ž .
were mainly due a poor performance during the first period of the experiment Fig. 3 . Perhaps this period might be regarded as a learning or adaptation period, because feeding times were shifted from those regarded as natural, whereas fish in the NF treatment were exposed to predetermined meal cycle matched to the natural feeding
Ž . Ž .
rhythm. The poorest growth of NF fish third period and DF fish first period coincided with the diurnalism and nocturnalism of SF fish, respectively. These findings
Ž .
are in broad agreement with those of Boujard 1995 who observed a reduction in the
Ž .
DF, in the time of feeding inversions from diurnal to nocturnal. The ability of restricted feeding schedules to induce phase inversions in feeding rhythms in sea bass was
Ž .
previously reported Sanchez-Vazquez et al., 1995 .
´
´
Ž .
The CVSf were significantly smaller than CVSi P-0.01 in all treatments, indicat-ing that all groups became more homogeneous with the passage of time. This is in accordance with previous findings for groups of sea bass submitted to different feeding
Ž
strategies and fed using the same level of reward for each trigger activation 8–9 pellets
. Ž .
for 15 fish Azzaydi et al., 1998, 1999 .
Sea bass shift their daily feeding patterns from nocturnal to diurnal in winter and early spring, and feeding sea bass by night, in winter, may result in improved growth performance. Although the flexibility of sea bass feeding habits enabled fish to adapt to diurnal feeding, one cost of adaptation to feeding out of phase with the natural feeding rhythms seem to be reduced growth. Further work is needed to assess any benefits there might be in nocturnal feeding of sea bass under farming conditions.
5. Further reading
Anthouard et al., 1986; Hidalgo et al., 1988
Acknowledgements
This research was supported by grants from the ‘‘Comision Interministerial de
´
Ž .
Ciencia y Tecnologıa’’
´
CICYT, no. AGF95-0459 to J.A. Madrid and from theŽ
‘‘Comunidad Autonoma de la Region de Murcia’’
´
CARM, FI-con 96r9 to J.A..
Madrid . The authors wish to acknowledge Dr. Benjamın Garcıa and other staff from the
´
´
‘‘Centro de Recursos Marinos y Humedales del Litoral de San Pedro del Pinatar’’ for their kind support and assistance during the experiments. We are also grateful to Culmarex for supplying the sea bass and the ‘‘Agencia Espanola de Cooperacion˜
´
Ž .
Internacional ICMAMPD ’’, for the financial aid received by M. Azzaydi.
References
Ž
Alanara, A., 1992a. Demand-feeding as a self-regulating feeding system for rainbow trout Oncorhynchus¨ ¨
.
mykiss in net-pens. Aquaculture 108, 347–356.
Alanara, A., 1992b. The effect of time-restricted demand-feeding on feeding activity, growth and feed¨ ¨
Ž .
conversion in rainbow trout Oncorhynchus mykiss . Aquaculture 108, 357–368.
Ž .
Alanara, A., 1996. The use of self-feeders in rainbow trout Oncorhynchus mykiss production. Aquaculture¨ ¨ 145, 1–20.
Anthouard, M., Desportes, C., Kentouri, M., Divanach, P., Paris, J., 1986. Etude des modeles comportemen-` taux manifestes au levier par Dicentrarchus labrax, Diplodus sargus, Puntazzo puntazzo, Sparus aurata et´
Ž . w
Lithognathus mormyrus poissons teleosteens , places dans une situation de nourissage auto-controle A´ ´ ´ ´ ˆ ´ study of behavioural patterns of Dicentrarchus labrax, Diplodus sargus, Puntazzo puntazzo, Sparus aurata
Ž . x
and Lithognathus mormyrus Teleostei Fishes in a self-feeding situation . Biol. Behav. 11, 97–110.
Ž
Anthouard, M., Divanach, P., Kentouri, M., 1993. An analysis of feeding activities of sea bass Dicentrarchus
.
Azzaydi, M., Madrid, J.A., Zamora, S., Sanchez-Vazquez, F.J., Martınez, F.J., 1998. Effect of three feeding´ ´ ´
Ž .
strategies automatic, ad libitum demand-feeding and time-restricted demand-feeding on feeding rhythms
Ž .
and growth in European sea bass Dicentrarchus labrax L. . Aquaculture 163, 285–296.
Azzaydi, M., Martınez, F.J., Zamora, S., Sanchez-Vazquez, F.J., Madrid, J.A., 1999. Effect of meal size´ ´ ´
Ž .
modulation on growth and feeding rhythms in European sea bass Dicentrarchus labrax L. . Aquaculture 170, 253–266.
w x Ž .
Barnabe, G., 1986. L’elevage du loup y de la daurade Sea bass and sea bream farming . In: Barnabe, G. Ed. ,´ ´ ´ Aquaculture. Technique and Documentation Lavoisier, Paris, France, 2, 627–666.
Begout-Anras, M.L., 1995. Demand-feeding behaviour of sea bass kept in ponds: diel and seasonal patterns´ and influences of environmental factors. Aquacult. Int. 3, 186–195.
Boujard, T., 1995. Diel rhythms of feeding activity in the European catfish, Silurus glanis. Physiol. Behav. 58, 641–645.
Boujard, T., Leatherland, J.F., 1992a. Circadian rhythms and feeding time in fishes. Environ. Biol. Fish 35, 109–131.
Boujard, T., Leatherland, J.F., 1992b. Demand-feeding behaviour and diel pattern activity in Oncorhynchus
mykiss held under different photoperiod regimes. J. Fish Biol. 40, 535–544.
Boujard, T., Gelineau, A., Corraze, G., 1995. Time of a single daily meal influences growth performance in
Ž .
rainbow trout Oncorhynchus mykiss . Aquacult. Res. 26, 341–349.
Boujard, T., Jourdan, M., Kentouri, M., Divanach, P., 1996. Diel feeding activity and the effect of time-restricted self-feeding on growth and feed conversion in European sea bass. Aquaculture 139, 117–127.
Divanach, P., Kentouri, M., Dewavrin, G., 1986. Sur le servage et l’evolution des performances biologiques´ d’alevins de daurades, Sparus auratus, provenant d’elevage extensif, apres remplacement des nourisseurs´ `
w
en continu par des distributeurs libre-service The weaning and the development of biological performance of extensively reared sea bream, Sparus auratus, fry after replacing continuous feeders by self-feeding
x
distributors . Aquaculture 52, 21–29.
Divanach, P., Kentouri, M., Charalambakis, G., Pouget, F., Sterioti, A., 1993. Comparison of growth
Ž .
performance of six Mediterranean fish species reared under intensive farming conditions in Crete Greece
Ž .
in raceways with use of self-feeders. In: Barnabe, G., Kestemont, P. Eds. , Production, Environment and´ Quality. EAS Special Publication, Ghent, Belgium, pp. 285–297.
Eriksson, L.O., Alanara, A., 1992. Timing of feeding behavior in salmonids. In: Thorpe, J.E., Huntingford,¨ ¨
Ž .
F.A. Eds. , The Importance of Feeding Behavior for the Efficient Culture of Salmonid Fishes. World Aquaculture Workshops World Aquaculture Society, Baton Rouge, LA, 2, pp. 41–48.
Fraser, N.H.C., Metcalfe, N.B., Thorpe, J.E., 1993. Temperature-dependant switch between diurnal and nocturnal foraging in salmon. Proc. R. Soc. London 252, 135–139.
Gelineau, A., Mambrini, M., Leatherland, J.F., Boujard, T., 1996. Effect of feeding time on hepatic nucleic´ acid, plama T3, T4 and GH concentrations in rainbow trout. Physiol. Behav. 59, 1061–1067.
Hidalgo, F., Kentouri, M., Divanach, P., 1988. Sur l’utilization du self-feeder comme outil d’epreuve w
nutritionelle du loup, Dicentrarchus labrax, L. Resultats preliminaires avec la methionine The utilisation´ ´ ´ of a self-feeder as a tool for the nutritional study of sea bass, Dicentrarchus labrax — preliminary results
x
with methionine . Aquaculture 68, 177–190.
Jobling, M., Arnesen, A.M., Baardvik, B.M., Christiansen, J.S., Jørgensen, E.H., 1995. Monitoring feeding behaviour and food intake: methods and applications. Aquacult. Nutr. 1, 131–143.
Kadri, S., Metcalfe, N.B., Huntingford, F.A., Thorpe, J.E., 1991. Daily feeding rhythms of Atlantic salmon in sea cages. Aquaculture 92, 219–224.
Kentouri, M., Divanach, P., Mainget, E., 1993. Comparaison de l’efficacite-cout de trois techniques d’alimen-´ ˆ w
tation de la daurade Sparus aurata en elevage intensif en bassin Effect of three feeding regimes on cost´ x
effectiveness in sea-bream Sparus aurata intensively reared in tanks . In Barnabe, G., Kestemont, P.´
ŽEds. , Production Environment and Quality. EAS Special Publication, Ghent, Belgium, pp. 273–283..
Kerdchuen, N., Legendre, M., 1991. Influence de la frequence et de la periode de nourrissage sur la croissance´
Ž .w
et l’efficacite alimentaire d’un silure africain, Heterobranchus longifilis Teleostei, Clariidae Influence of´ feeding frequency and feeding time on growth and feed efficiency in African catfish Heterobranchus
Ž .x
longifilis Teleostei, Clariidae . Aquat. Living Resour. 4, 241–248.
Madrid, J.A., Azzaydi, M., Zamora, S., Sanchez-Vazquez, F.J., 1997. Continuous recording of uneaten food´ ´ pellets and demand-feeding activity: a new approach to studying feeding rhythms in fish. Physiol. Behav. 62, 689–695.
Noeske, T.A., Erickson, D., Spieler, R.E., 1981. The time of day goldfish receive a single daily meal affects growth. J. World Maric. Soc. 12, 73–77.
Noeske, T.A., Spieler, R.E., 1984. Circadian feeding time affects growth of fish. Trans. Am. Fish. Soc. 113, 540–544.
Noeske-Hallin, T.A., Spieler, R.E., Parker, N.C., Suttle, M.A., 1985. Feeding time differentially affects fattening and growth of channel catfish. J. Nutr. 115, 1228–1232.
Sanchez-Vazquez, F.J., Martınez, M., Zamora, S., Madrid, J.A., 1994. Design and performance of an accurate´ ´ ´ demand feeder for the study of feeding behaviour in sea bass, Dicentrarchus labrax L. Physiol. Behav. 56, 789–794.
Sanchez-Vazquez, F.J., Madrid, J.A., Zamora, S., 1995. Circadian rhythms of feeding activity in sea bass,´ ´
Dicentrarchus labrax L.: dual phasing capacity of diel demand-feeding pattern. J. Biol. Rhythms 10,
256–266.
Sanchez-Vazquez, F.J., Azzaydi, M., Martinez, F.J., Zamora, S., Madrid, J.A., 1998. Annual rhythms of´ ´ demand-feeding activity in sea bass: evidence of a seasonal phase inversion of the diel feeding pattern. Chronobiol. Int. 15, 607–622.
Smith, I.P., Metcalfe, N.B., Huntingford, F.A., Kadri, S., 1993. Daily and seasonal patterns in the feeding
Ž .
behaviour of Atlantic salmon Salmo salar L. in sea cage. Aquaculture 117, 165–178.
Sundararaj, B., Nath, P., Halberg, F., 1982. Circadian meal timing in relation to lighting schedule optimizes catfish body weight gain. J. Nutr. 112, 1085–1097.