Ž .
Aquaculture 182 2000 249–260
www.elsevier.nlrlocateraqua-online
Bacterial additives that consistently enhance rotifer
growth under synxenic culture conditions
1. Evaluation of commercial products and pure
isolates
P.A. Douillet
)The UniÕersity of Texas at Austin, Marine Science Institute, 1300 Port Street, Port Aransas, TX 78373, USA Accepted 20 July 1999
Abstract
Ž .
Axenic rotifers Brachionus plicatilis Muller were cultured under aseptic conditions; they¨
Ž .
were fed either a bacteria-free artificial diet AD , or axenic Isochrysis galbana, or a combination of axenic Chlorella minutissima and the bacteria-free AD. The medium was inoculated with commercial bacterial additives or cultured strains of marine bacteria. The highest improvements in
Ž .
growth rate GR of rotifer populations were obtained with laboratory grown bacteria. Addition of Ž .
an Alteromonas strain and an unidentified Gram negative strain B3 consistently enhanced rotifer GR in all experiments, and under all feeding regimes in comparison with control cultures inoculated with microbial communities present in seawater, or maintained bacteria-free. None of the other isolates or commercial products were consistent in their enhancement of rotifer
production.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Rotifer; Brachionus plicatilis; Isochrysis galbana
1. Introduction
The rotifer Brachionus plicatilis has become a valuable and, in many cases indis-pensable, food organism for first feeding of a large variety of cultured marine finfish and
Ž .
crustacean larvae Watanabe et al., 1983; Lubzens et al., 1997 . However, suppressed
) 1692 Houghton Ct North, Dunwoody, GA 30338, USA. Tel.: q1-770-671-9393; E-mail:
philippe [email protected]–
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
growth or unforeseen death of rotifers are frequently observed in mass cultures ŽHirayama, 1987; Ushiro et al., 1990; Maeda and Hino, 1991; Hino, 1993 . Rotifer. cultures harbor very large bacterial populations, which have been estimated to be in the
7 y1 Ž .
order of 10 cells ml Nicolas and Joubert, 1986; Nicolas et al., 1989 . Rapid successional processes in the microbiota have been observed during the culture of
Ž .
rotifers Maeda and Hino, 1991 , and changes in the microbial ecosystem have been
Ž .
postulated as the cause of the collapse of rotifer cultures Hino, 1993 .
The effects of bacteria on rotifer cultures are strain specific, as demonstrated by the
Ž . Ž . Ž .
findings of Yasuda and Taga 1980 , Yu et al. 1988 , Gatesoupe et al. 1989 , Maeda
Ž . Ž .
and Hino 1991 and Hagiwara et al. 1994 . These authors reported strains, from diverse taxonomical groups, that were able to either decrease or increase the growth rate ŽGR of B. plicatilis. However, bacterially mediated changes in rotifer GRs are caused. by diverse mechanisms. A nutritional contribution of bacteria to rotifer diets has been
Ž . Ž
demonstrated by supply of vitamin B12 Yu et al., 1988 or inorganic nutrients Hessen .
and Andersen, 1990 . In contrast, production of bacterial toxins has been found to
Ž .
reduce rotifer survival rates Yu et al., 1990 . Another possible effect of bacteria in rotifer cultures is the biochemical transformation of accumulated waste products. Nitrogen budgets carried out with rotifers fed Nanochloropsis sp. revealed that 82%– 84% of the ingested N was released into the water as metabolic excretion and feces ŽTanaka, 1991; Hino et al., 1997 . Accumulation of metabolic products and excess.
Ž .
uneaten food cause deterioration of water quality Lubzens, 1987 , which may affect
Ž .
rotifer growth and reproduction Tanaka, 1991 . In fact, rotifer densities have been Ž
reported to decrease with increases of either un-ionized ammonia Yu and Hirayama,
. Ž .
1986 or nitrite Lubzens, 1987 . Removal of waste products from rotifer cultures has
Ž .
been reported to extend the harvest period Lubzens, 1987 . A bacterially mediated improvement in water quality might be a very plausible mechanism for increasing rotifer GRs.
In this study, the effects of additions of laboratory-grown microbes and several commercial bacterial additives were evaluated on the GR of B. plicatilis cultured under synxenic conditions, i.e., rotifers were grown in the presence of a known number, one or more, of microbial species. Single strains and commercial products with diverse characteristics that might be beneficial for rotifers were selected so that the evaluation of microbes would cover different plausible bacterial mechanisms for rotifer culture Ž . enhancement. The screening of microbes was carried out under an artificial diet AD and different algae feeding regimes.
2. Materials and methods
2.1. Preparation of rotifers
Ž .
Cysts of the rotifer B. plicatilis Muller formerly called L-type B. plicatilis were
¨
( )
P.A. DouilletrAquaculture 182 2000 249–260 251
Ž1998 . To confirm that the rotifers were axenic, incubation tests of samples of rotifers. in broth and agar under aerobic and anaerobic conditions were continued for 30 days. Axenicity tests were also performed on axenic and starved cultures at the end of the experiments. The experiment was discarded if bacterial contamination was detected.
2.2. Preparation of diets
An AD was developed and tested in preliminary experiments. The diet was prepared Ž
by dissolving 8 g of microfine Spirulina 8–10 by 20 mm; Aurum Aquaculture,
. Ž
Washington and 8 g of Torula dried yeast Lake States Division of Rhinelander Paper, .
Wisconsin in 1 l of seawater at 15 ppt salinity. The dissolved diet was autoclaved. After
Ž .
cooling, filter-sterilized cyanocobalamine vitamin B12 was added at a concentration of 120 mg ly1 to the flask of AD to be used for first feeding only. The adequacy of the diet was ascertained by observing no significant difference between rotifer production in
Ž . cultures fed either this diet or the diet developed by Gatesoupe and Luquet 1981 .
Ž .
Axenic Isochrysis galbana clone C-ISO, CCMP463 and Chlorella minutissima Žclone 2341 used in Experiments 2 and 4 were obtained from the National Center for.
Ž .
Culture of Marine Phytoplankton Maine and The Culture Collection of Algae at The Ž University of Texas at Austin, respectively. Algae were grown in fr2 media Guillard
.
and Ryther, 1962 at 20–25 ppt salinity. Algal cultures were maintained in an incubator at 258C under constant cool-white fluorescent light at an intensity of 2250–4600 lx. Axenicity of algae was determined as described above for rotifers. Both species of algae were grown in 200 ml Erlenmeyer flasks. The cells were concentrated by centrifugation and resuspended at high concentrations in FASW, so that their daily addition to rotifer
Ž .
cultures approx. 20ml would have little impact on rotifer densities. Rotifer cultures fed AD only were amended daily with FASW to maintain similar volumes to algae-fed rotifer cultures.
2.3. Preparation of bacteria
Commercially available bacterial additives were added directly to rotifer cultures. Bacterial strains kindly provided by other scientists or isolated by the author were cultured on Difco marine agar for 2–3 days, resuspended in FASW, washed by
Ž .
centrifugation 10,000=g for 10 min and resuspended in FASW. Photosynthetic
Ž . Ž
bacteria PH were cultured on Rhodospirillum ATTC Medium 1308 Atlas and Parks, .
1993, p. 774 for 1 week at 258C, under constant cool-white fluorescent light at an intensity of 2250–4600 lx.
All glassware was washed in 10% nitric acid and rinsed seven times with tap water. Heat sterilization was carried out for 15 min at 1218C and a pressure of 1.06 kg cmy2. All manipulations were done under a laminar flow hood.
2.4. Experimental protocol
experiments to corroborate initial rotifer densities. Culture experiments were initiated by the addition of food and the different bacteria.
Ž .
Control cultures consisted of: 1 cultures fed the same diets but maintained bacteria-Ž .
free, 2 cultures fed the same diets and inoculated with bacteria present in 100 ml Ž . Ž . samples of freshly collected seawater filtered through a 1-mm screen SW , 3 starved
Ž .
cultures in Experiment 3, and 4 rotifer cultures fed only axenic I. galbana in Experiment 1.
Rotifers were fed AD andror algae daily. The first day of culture, the AD was added at a final concentration of 0.2 mg mly1; then, the ration was decreased to 0.14 mg mly1 dayy1. The final concentration of cyanocobalamine after the first feeding was 1.5 g
y1 Ž .
ml , as recommended by Hirayama and Funamoto 1983 . This vitamin was added only with the first feeding. Rotifers were fed on AD in Experiments 1 and 3. In Experiment 2, rotifers were fed either AD or axenic I. galbana. Rotifers fed I. galbana received daily additions to maintain a final concentration of 2=106 cells mly1. In Experiment 4, rotifers were fed either AD or a combination of AD and axenic C.
minutissima. Rotifers were fed the same concentrations of AD under both feeding
treatments. Rotifers supplemented with C. minutissima received daily algal additions to maintain a final concentration of 1=107cells mly1.
Commercial bacterial additives and pure bacterial isolates were added only once to rotifer cultures, on day one of the experiments, at a final concentration of 2=107cells mly1. Bacteria concentrations were derived from equations relating spectrophotometric
Ž .
absorbance 600 nm and bacteria numbers; the latter value was determined by direct
Ž .
count using DAPI staining techniques Porter and Feig, 1980 . Such equations were developed and used for each bacterial additive tested. Commercial bacterial products and cultured strains tested in this research are presented in Table 1. In order to determine consistency of beneficial effects, strains that improved rotifer GR over axenic controls were repeatedly tested and discarded if the beneficial effects were not maintained. Bacterial treatments referred by their code name in Table 1 and tested in Experiment 1
Ž
included nine commercial products Acc, A2, A5, A6, A1100, A1200, F9, Mplus and
. Ž
Sy as well as eight laboratory cultured strains Vibrio alginolyticus, Alteromonas sp., .
B1, B2, B3, B4, B5 and PH . In Experiment 2, the strain Enterococcus faecium was Ž
tested along with nine inoculants A1200, A5, Alteromonas sp., B2, B3, B4, B5, PH .
and V.a which were evaluated for a second time. In Experiment 3, the additives Acc Ž
and B1 were evaluated for a second time, and five strains Alteromonas sp., B3, B4, B5 .
()
P.A.
Douillet
r
Aquaculture
182
2000
249
–
260
253
Table 1
Bacterial additives or bacteria strains tested in four experiments with B. plicatilis
Ž . Ž .
Commercial name code Bacterial composition Origin presentation
Ž . Ž .
AB-1 AB1 Bacillus subtilis Advanced Microbial Systems, MN liquid
Ž . Ž .
Aqua-Bacta-Aid N2 ABA B. subtilis, B. amyloliquefaciens, B. licheniformis, Water Quality Science International, MO liquid
Ž .
Nitrosococcus two strains
Ž . Ž .
Accelobac Acc B. subtilis, B. cereus, B. megaterium, B. licheniformis, American Biosystems, VA dry
B. polymyxa, Rumenococcus albus, Aspergillus oryzae
Ž . Ž .
Alken Clear-Flo 1100 A1100 Nitrosomonas sp., Nitrobacter sp. Alken Murray, NY liquid
Ž . Ž . Ž .
Alken Clear-Flo 1200 A1200 B. subtilis two strains , Pseudomonas aeruginosa, P. stutzeri, Alken Murray, NY liquid
P. fluorescens, Escherichia hermanii, Nitrosomonas sp., Nitrobacter sp.
ŽA2. B. subtilis three strains , B. licheniformis,Ž . Alken Murray, NY dryŽ .
P. aeruginosa, P. stutzeri
ŽA5. P. aeruginosa, P. stutzeri, P. putida Alken Murray, NY dryŽ . ŽA6. B. subtilis, B. licheniformis, B. polymixa Alken Murray, NY dryŽ .
Ž . Ž .
Fritz-Zymea9 F9 Nitrosomonas sp., Nitrobacter sp. Fritz, TX liquid
Ž . Ž . Ž .
Microbials Plus Mplus Streptococcus faecium now E. faecium Medipharm, IA dry
ŽSy. P. aeruginosa 2203 Sybron Chemicals, VA dryŽ .
ŽA.sp.. Alteromonas sp. Seawater isolates cultured at UTMSIŽ .
ŽB1, B2, B3, B4, B5. Five unidentified marine GramŽy.rods Seawater isolates cultured at UTMSIŽ . ŽE.f. E. faecium P. Bogaert, University of Gent, Belgium dryŽ .
ŽPH. Photosynthetic red non-sulfur bacteria Qingdao Oceanogr. University, China
Žcultured at UTMSI.
ŽV.a. V. alginolyticus Dr. Don Lewis, Texas A&M University,
Ž .
2.5. Data collection and analysis
At the end of each experiment, starved and axenic controls were sampled and tested as described above for bacterial contamination. Cultures were then homogenized by
Ž .
shaking and five samples 100–1000 ml were withdrawn for estimation of rotifer densities. Rotifer population GR for each culture tube was calculated as:
GRs ln final density
Ž
.
yln initial densityŽ
.
rculture period in days 4 daysŽ
.
Parametric assumptions were evaluated using Hartley’s test for homogeneity of variances, Tukey’s test for non-additivity and Wilk–Shapiro’s test for normality. Bacterial treatments were tested under two different feeding regimes in Experiments 2 and 4. These data sets were initially analyzed by two-way ANOVA, with feeding regime and bacterial treatment as variables. GR in all experiments was then analyzed using
Ž .
one-way ANOVA, followed by Tukey’s range test T-method, Sokal and Rohlf, 1981 to determine differences between bacterial treatments at the 0.05 level of probability.
Ž U. Ž
Coefficients of variation V between replicates under four treatments axenic, SW, .
B3 and Alteromonas sp. were calculated in each experiment as in Sokal and Rohlf Ž1981 :.
U
V s
Ž
100 sd.
rmeanŽ
1q1r4 n.
where sd is the standard deviation, mean is the average GR and n is the number of
Ž . U
replicates ns4 . Independent V were determined for the different treatments under each one of the diets in Experiments 2 and 4; therefore, six VU values were calculated for each treatment in the four experiments. VU values under the SW treatment were compared with the VU values determined under the axenic, B3 and Alteromonas sp.
Ž .
treatments using the t-test for paired comparisons Sokal and Rohlf, 1981 . All tests
Ž .
were performed with the computer program Statistix 2 NH Analytical Software .
3. Results
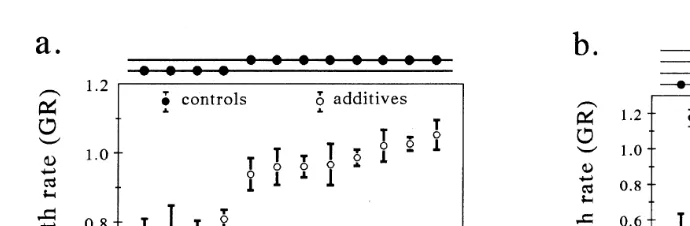
No evidence of bacterial contamination was found in either starved or axenic control cultures at the end of the culture period. Axenic rotifers populations grew with all diets tested. GRs in rotifer populations inoculated with SW bacteria did not differ from those
Ž .
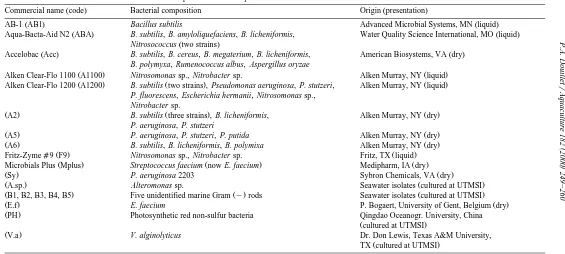
obtained with axenic controls Figs. 1 and 2 . Significant differences in GRs between treatments were found in all experiments.
Ž
In Experiment 1, additions of eight cultured bacterial additives B5, Alteromonas sp.,
. Ž
PH, B3, B2, B4, V. alginolyticus and B1 and five commercial products A1200, A2, .
A6, Acc and A5 resulted in larger GRs than those determined in axenic control cultures ŽTukey’s, p-0.05 . The second highest GR was obtained in control cultures fed only.
Ž .
on axenic I. galbana Fig. 1a .
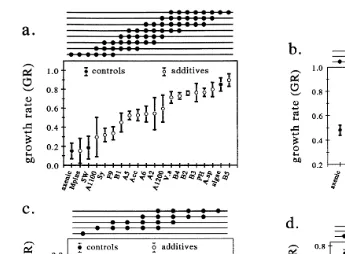
In Experiment 2, significant differences in GR were determined between feeding
Ž .
regimes and bacterial treatments two-way ANOVA, p-0.05 . Therefore, data sets for each feeding regime were analyzed by one-way ANOVA. Under the AD feeding regime,
Ž .
( )
P.A. DouilletrAquaculture 182 2000 249–260 255
Fig. 1. GRs of B. plicatilis cultured under synxenic conditions with different bacterial additives and fed axenic
Ž . Ž . Ž . Ž .
ADs in Experiment 1 a , Experiment 2 b , Experiment 3 c and Experiment 4 d . Results of Tukey’s range tests are displayed above the histogram. Circles that occur together on any one of the horizontal lines indicate mean values that are not different at the 0.05% level of significance. See Table 1 for details of diets.
Ž . Ž
inoculated with one commercial product A5 and five cultured bacterial additives B5, . Ž .
Alteromonas sp., B3, B4 and PH Fig. 1b . Under the I. galbana feeding regime, GRs Ž
were significantly improved over axenic controls with all bacterial treatments Tukey’s,
. Ž .
p-0.05 except for A1200 and A5 Fig. 2a .
In Experiment 3, a negative GR was observed in starved cultures due to mortality of
Ž . Ž
initial rotifer populations Fig. 1c . Addition of four strains Alteromonas sp., B3, B5
. Ž
and B4 significantly enhanced GRs of rotifers over axenic control cultures Tukey’s, .
p-0.05 .
As in Experiment 2, significant differences in GRs were determined between feeding
Ž .
regimes and bacterial treatments in Experiment 4 two-way ANOVA, p-0.05 . There-fore, data sets for each feeding regime were analyzed by one-way ANOVA. Under the AD feeding regime, addition of all commercial additives and strain B4 resulted in lower GRs than those determined in axenic controls; however, the differences were not
Ž . Ž .
significant Fig. 1d . Under the mixed feeding regime C. minutissimaqAD , addition of the commercial products AB1 and ABA resulted in lower GRs than in axenic
Ž .
Fig. 2. GRs of B. plicatilis cultured under synxenic conditions with different bacterial additives and fed either
Ž . Ž .
axenic I. galbana in Experiment 2 a or a combination of axenic AD and C. minutissima in Experiment 4 b . Results of Tukey’s range tests are displayed above the histogram. Circles that occur together on any one of the horizontal lines indicate mean values that are not different at the 0.05% level of significance. See Table 1 for details of diets.
ŽFig. 2b . Under both feeding regimes, addition of strains B3 and Alteromonas sp. did.
Ž .
improve rotifer GR over axenic controls Tukey’s, p-0.05 , while addition of strain B5 resulted in GRs that did not differ from those of the axenic controls.
VU
values in cultures seeded with SW bacteria were found to be significantly larger
Ž .
than either those calculated in axenic cultures t-test, ps0.018 , cultures inoculated
Ž . Ž .
with B3 ps0.018 or cultures inoculated with Alteromonas sp. ps0.025 .
4. Discussion
Axenic rotifers were used to evaluate the effects of additions of several bacterial additives on the GRs of rotifer populations fed diverse diets. By using axenic rotifers, the effects determined under each treatment can be ascribed to the microbes added to the culture system without interference from bacterial contaminants that could carry out any of the different mechanisms that could affect rotifer GR. Rotifer populations were able to grow on all diets tested under axenic conditions. Lower GRs resulted in cultures of
Ž .
axenic rotifers fed bacteria-free AD 0.28"0.15, mean"sd, ns16 , than bacteria-free
Ž .
I. galbana 0.81"0.07, ns8 or a mixed bacteria-free diet of AD and C. minutissima Ž0.84"0.03, ns4 . The GRs of rotifers in cultures inoculated with several laboratory. isolates were significantly higher than those recorded in cultures fed the same diet, but kept axenic. The largest GR improvement was determined with AD in Experiment 1, where the addition of strain B5 resulted in a GR 5.9 times larger than in axenic controls fed the same diet. The importance of a bacterial component in rotifer cultures was illustrated in this first experiment, whereas the addition of five bacteria strains to cultures of rotifers fed AD resulted in GR levels that did not differ from those observed in cultures of rotifers fed axenic algae.
signifi-( )
P.A. DouilletrAquaculture 182 2000 249–260 257
cantly larger VU values than in axenic cultures. Except for Experiment 4, lower GRs occurred in cultures seeded with SW bacteria compared with cultures seeded with any other bacterial amendment.
Most marine fish and crustacean hatcheries use seawater for the culture of rotifers. However, filtration and chlorination, which are the most widely used methods of water
Ž
disinfection in hatcheries, may not eliminate all seawater microbes Douillet and .
Pickering, 1999 . So bacteria present in seawater may constitute one of the mayor sources of contamination in rotifer cultures. Most studies on the effects of bacteria on rotifer cultures have been carried out under xenic culture conditions; therefore, in the following discussion, the effects of bacterial additives were compared with control cultures inoculated with SW bacteria.
Commercial bacterial products tested in this study were either beneficial or slightly adverse to rotifer production. The largest improvement caused by the addition of a
Ž .
commercial product Alken 1200, Experiment 1 was an increase in rotifer GR of 4.5 times the value determined in SW controls. Unfortunately, none of the commercial products that showed beneficial characteristics for rotifer culture was consistent in repeated trials. Furthermore, addition of isolate B5 in Experiment 1 resulted in rotifer GR 5.7 times larger than in SW controls. Therefore, the magnitudes of improvement of rotifer growth caused by commercial products were smaller than those obtained with laboratory grown microbes.
Addition to rotifer cultures of the commercial products F9 and A1100, which contain
Ž .
nitrifying bacteria Nitrosomonas sp. and Nitrobacter sp. , did not result in any GR
improvement. Several strains of Bacillus and Pseudomonas are well known for their
Ž .
ability to produce exoenzymes that break down organic matter Pollock, 1962 , and thus they are frequently included in commercial products for waste water treatment. The
Ž .
commercial product that most enhanced the GR of rotifers Alken 1200 is a blend of
Bacillus strains and nitrifying bacteria. Unfortunately, no water quality analyses were
carried out to determine a bacterially mediated water quality improvement mechanism. Photosynthetic bacteria are frequently used for wastewater treatment. The photosynthetic
Ž .
strains tested in this study PH were isolated from the bottom of shrimp ponds, and are currently used as probiotics in Chinese hatcheries. PH bacteria were beneficial for rotifer growth in two out of three experiments.
Bacteria strains that improve production of aquatic organisms can be either consistent
Ž .
or not in their enhancement properties Douillet and Langdon, 1993 . Alteromonas sp. Ž .
and an unidentified Gram negative strain B3 were found in this study to be consis-tently beneficial to rotifers in all experiments and under all feeding regimes.
Al-teromonas sp. tested in this study was evaluated in previous studies with larvae of the
Pacific oyster Crassostrea gigas. Consistent enhancement of oyster larvae production Ž
was achieved with the addition of Alteromonas sp. under synxenic Douillet and
. Ž .
Langdon, 1993 and xenic culture conditions Douillet and Langdon, 1994 . The magnitude of the enhancement of rotifer GRs caused by the addition of Alteromonas sp. and B3 with respect to SW control cultures changed between experiments when using
Ž .
AD 1.7–4.3 times increase in GR for Alteromonas sp. and 1.6–4.1 times for B3 . It was of less magnitude and more constant for cultures fed either algae or a combination
Ž .
Ž .
al. 1988 found the magnitude of bacterial improvement in GRs of rotifers higher when using AD than when using algae. Using data from Figs. 4 and 5 in Yu’s paper, the addition of a Pseudomonas producer of vitamin B12 resulted in a GR of rotifers 2.9 times higher than in controls when fed AD and 2.6 times higher than controls when fed algae.
Nutritional supplementation leading to enhanced rotifer GR has been reported for
Ž . Ž
algae with blue-green algae Snell et al., 1983 , algae with yeast Hirayama and
. Ž .
Watanabe, 1973 , dried algae with yeast Hirayama and Nakamura, 1976 , and
photo-Ž .
synthetic bacteria with either algae or yeast Sakamoto and Hirayama, 1983 . The bacterial enhancement of GR determined in rotifers fed AD, and to a lesser extend on algae, might be due to the fact that bacteria provide essential compounds such as
Ž . Ž .
vitamins Yu et al., 1988 or inorganic nutrients Hessen and Andersen, 1990 deficient in the diets. Soluble compounds excreted by bacteria into the culture medium are strain
Ž .
specific Yu et al., 1989; Hagiwara et al., 1994 , which could explain variation in rotifer production under different bacterial treatments. A nutritional contribution of Al-teromonas sp. to oyster larvae was demonstrated by14C-feeding techniques whereby this strain provided in some cases over 180% of the carbon metabolic requirements of the
Ž .
larvae Douillet, 1993 .
This study, carried out under synxenic conditions, showed a very significant variation in rotifer population growth resulting from changes in the species composition of the microbiota. Addition of a few commercial products significantly improved rotifer GRs; unfortunately, the beneficial effects were not consistent in repeated trials with any of these products. In contrast, selected bacterial strains were consistent in their enhance-ment of rotifer GRs under all feeding regimes. Furthermore, variations in GR between replicate cultures seeded with these selected strains were significantly lower than in cultures seeded with SW bacteria. GR is highly dependent on the previous life history, i.e., accumulated nutrients, reproductive condition, of the rotifers used to start
popula-Ž .
tions Scott and Baynes, 1978; Snell et al., 1983 ; therefore, it is difficult to compare GRs from different studies. However, GRs determined in batch cultures of rotifers fed
Ž
AD oscillate between 0.15 to 0.62 Hirayama and Nakamura, 1976; Yufera and Pascual,
´
.
1980; Komis et al., 1991; Shiri Harzevili et al., 1995 , while those determined with Ž
rotifers fed algae fall between the wider range of 0.09–1.09 Theilacker and McMaster, 1971; Scott and Baynes, 1978; Sakamoto and Hirayama, 1983; Snell et al., 1983; Maeda
. Ž .
and Hino, 1991; Lubzens et al., 1993 . Two findings from this research, a poor GRs occurred in cultures inoculated with SW bacteria, which are the microbes likely to
Ž .
colonize commercial rotifer production systems, and b consistent improvements in GR resulted by adding selected microbes, indicate the potential for microbial management to improve rotifer production in culture facilities.
Acknowledgements
This work was supported by GrantaNA56RG0388 from the National Oceanic and
( )
P.A. DouilletrAquaculture 182 2000 249–260 259
Ž
of NOAA or any of its sub-agencies. I am indebted to Dr. Ron Benner The University .
of Texas at Austin, Marine Science Institute for the use of his epifluorescence
Ž .
microscope. I am grateful to P. Bogaert University of Gent, Belgium and Dr. Don
Ž .
Lewis Texas A & M University, USA for supplying bacteria samples. Photosynthetic
Ž .
microbes were kindly provided by Dr. Anja Robinson Oregon State University , who obtained this product from researchers at Qingdao Oceanographic University, China. I am indebted to the following companies for supplying samples of their products: Advanced Microbial Systems, Alken Murray, American Biosystems, Medipharm, Sybron Chemicals and Water Quality Science International.
References
Atlas, R.M., Parks, L.C., 1993. Handbook of Microbiological Media. CRC, Boca Raton, 1079 pp.
Douillet, P., 1993. Carbon contribution through bacterivory in larvae of the Pacific oyster Crassostrea gigas. Mar. Ecol. Prog. Ser. 102, 303–314.
Douillet, P., 1998. Disinfection of rotifer cysts leading to bacteria-free populations. J. Exp. Mar. Biol. Ecol. 224, 183–192.
Douillet, P., Langdon, C.J., 1993. Effects of marine bacteria on the culture of axenic oyster Crassostrea gigas
ŽThunberg larvae. Biol. Bull. 184, 36–51..
Ž
Douillet, P., Langdon, C.J., 1994. Use of a probiotic for the culture of larvae of the Pacific oyster Crassostrea
.
gigas Thunberg . Aquaculture 119, 25–40.
Douillet, P., Pickering, P., 1999. Seawater treatment for larval culture of the fish Sciaenops ocellatus Linnaeus
Žred drum . Aquaculture 170, 113–126..
Gatesoupe, F.J., Luquet, P., 1981. Practical diet for mass culture of the rotifer Brachionus plicatilis: application to larval rearing of sea bass, Dicentrarchus labrax. Aquaculture 22, 149–163.
Gatesoupe, F.J., Arakawa, T., Watanabe, T., 1989. The effect of bacterial additives on the production rate and dietary value of rotifers as food for Japanese flounder, Paralichthys oliÕaceus. Aquaculture 83, 39–44. Guillard, R.R.L., Ryther, J.H., 1962. Studies on marine planktonic diatoms: I. Cyclotela nana Hustedt and
Detonula conferÕacea Cleve. Can. J. Microbiol. 8, 229–239.
Hagiwara, A., Hamada, K., Hori, S., Hirayama, K., 1994. Increased sexual reproduction in Brachionus
Ž .
plicatilis Rotifera with the addition of bacteria and rotifer extracts. J. Exp. Mar. Biol. Ecol. 181, 1–8.
Hessen, D.O., Andersen, T., 1990. Bacteria as a source of phosphorus for zooplankton. Hydrobiologia 206, 217–223.
Ž .
Hino, A., 1993. Present culture systems of the rotifer Brachionus plicatilis and the function of
micro-Ž .
organisms. In: Lee, C.S., Su, M.S., Liao, I.L. Eds. , Proceedings of Finfish Hatchery in Asia 91, 17–19 December 1991, Tungkang, Taiwan. Tungkang Marine Laboratory Conference Proceedings 3, pp. 51–59. Hino, A., Aoki, S., Ushiro, M., 1997. Nitrogen-flow in the rotifer Brachionus rotundiformis and its
significance in mass cultures. Hydrobiologia 358, 77–82.
Hirayama, K., 1987. A consideration of why mass culture of the rotifer Brachionus plicatilis with baker’s yeast is unstable. Hydrobiologia 147, 269–270.
Hirayama, K., Funamoto, H., 1983. Supplementary effect of several nutrients on nutritive deficiency of baker’s yeast for population growth of the rotifer Brachionus plicatilis. Bull. Jpn. Soc. Sci. Fish. 49, 505–510. Hirayama, K., Nakamura, K., 1976. Fundamental studies on the physiology of rotifers in mass culture: V. Dry
Chlorella powder as a food for rotifers. Aquaculture 8, 301–307.
Hirayama, K., Watanabe, K., 1973. Fundamental studies on physiology of rotifer for its mass culture: IV. Nutritional effect of yeast on population growth of rotifer. Bull. Jpn. Soc. Sci. Fish. 39, 1129–1133. Komis, A., Candreva, P., Franicevic, V., Moreau, V., Van Ballaer, E., Leger, P., Sorgeloos, P., 1991.´
Successful application of a new combined culture and enrichment diet for the mass cultivation of the rotifer Brachionus plicatilis at a commercial hatchery scale in Monaco, Yugoslavia, France and Thailand.
Ž .
Symposium, 27–30 August 1991, Gent, Belgium. European Aquaculture Society, Special Publication 15, pp. 102–103.
Lubzens, E., 1987. Raising rotifers for use in aquaculture. Hydrobiologia 147, 245–255.
Lubzens, E., Wax, Y., Minkoff, G., Adler, F., 1993. A model evaluating the contribution of environmental factors to the production of resting eggs in the rotifer Brachionus plicatilis. Hydrobiologia 255r256, 127–138.
Lubzens, E., Minkoff, G., Barr, Y., Zmora, O., 1997. Mariculture in Israel — past achievements and future directions in raising rotifers as food for marine fish larvae. Hydrobiologia 358, 13–20.
Maeda, M., Hino, A., 1991. Environmental management for mass culture of rotifer, Brachionus plicatilis. In:
Ž .
Fulks, W., Main, K.L. Eds. , Rotifer and Microalgae Culture Systems. Proceedings US–Asia Workshop, Honolulu, USA, 28–31 January 1991. The Oceanic Institute, pp. 125–133.
Nicolas, J.L., Joubert, M.N., 1986. Bacteries associees aux productions de Brachionus plicatilis. In: Gerbam,´
Ž .
Ifremer Eds. , 2nd Coll. Int. Bacteriologie Marine, 1–5 October 1984, Brest, France, Vol. 3, pp. 451–458.´ Nicolas, J.L., Robic, E., Ansquer, D., 1989. Bacterial flora associated with a trophic chain consisting of microalgae, rotifers and turbot larvae: influence of bacteria on larval survival. Aquaculture 83, 237–248.
Ž .
Pollock, M.R., 1962. Exoenzymes. In: Gunsalus, I.C., Stanier, R.Y. Eds. , The Bacteria, a Treatise on Structure and Function. Vol. IV, The Physiology of Growth. Academic Press, New York, pp. 121–178. Porter, K.G., Feig, Y.S., 1980. The use of DAPI for identifying and counting aquatic microflora. Limnol.
Oceanogr. 25, 943–948.
Ž .
Sakamoto, H., Hirayama, K., 1983. Dietary effect of Thiocapsa roseopersicina photosynthetic bacteria on the rotifer Brachionus plicatilis. Bull. Fac. Fish., Nagasaki Univ. 54, 13–20.
Scott, A.P., Baynes, S.M., 1978. Effect of algal diet and temperature on the biochemical composition of the rotifer, Brachionus plicatilis. Aquaculture 14, 247–260.
Shiri Harzevili, A.R., Van Duffel, H., Defoort, T., Sorgeloos, P., Swings, P., 1995. The influence of a selected bacterial strain Vibrio anguillarum TR27 on the growth rate of the rotifer, Brachionus plicatilis in two
Ž .
culture conditions. In: Lavens, P., Jaspers, E., Roelants, I. Eds. , Larvi 95 — Fish and Shellfish Larviculture Symposium, 3–7 September 1995, Gent, Belgium. European Aquaculture Society, Special Publication 24, pp. 480–483.
Snell, T.W., Bieberich, C.J., Fuerst, R., 1983. The effects of green and blue-green algal diets on the reproductive rate of the rotifer Brachionus plicatilis. Aquaculture 31, 21–30.
Sokal, R.R., Rohlf, F.J., 1981. Biometry. W.H. Freeman, San Francisco, 859 pp.
Tanaka, Y., 1991. Nitrogen budget for the rotifer Brachionus plicatilis. J. World Aquacult. Soc. 22, 57–64. Theilacker, G.H., McMaster, M.F., 1971. Mass culture of the rotifer Brachionus plicatilis and its evaluation as
a food for larval anchovies. Mar. Biol. 10, 183–188.
Ushiro, M., Yu, J.-P., Hino, A., 1990. Energy feedback in the culture of the rotifer Brachionus plicatilis. In:
Ž .
Hirano, R., Hanyu, I. Eds. , Proceedings of the Second Asian Fisheries Forum, 17–22 April 1989, Tokyo, Japan, pp. 157–160.
Watanabe, T., Kitajima, C., Fujita, S., 1983. Nutritional values of live organisms used in Japan for mass propagation of fish: a review. Aquaculture 34, 115–143.
Yasuda, K., Taga, N., 1980. Culture of Brachionus plicatilis Muller using bacteria as food. Bull. Jpn. Soc.¨ Sci. Fish. 46, 933–939.
Yu, J.-P., Hirayama, K., 1986. The effect of un-ionized ammonia on the population growth of the rotifer in mass culture. Bull. Jpn. Soc. Sci. Fish. 52, 1509–1513.
Yu, J.-P., Hino, A., Hirano, R., Hirayama, K., 1988. Vitamin B -producing bacteria as a nutritive12
complement for a culture of the rotifer Brachionus plicatilis. Nippon Suisan Gakkaishi 54, 1873–1880. Yu, J.-P., Hino, A., Ushiro, M., Maeda, M., 1989. Function of bacteria as vitamin B12 producers during mass
culture of the rotifer Brachionus plicatilis. Nippon Suisan Gakkaishi 55, 1799–1806.
Yu, J.-P., Hino, A., Noguchi, T., Wakabayashi, H., 1990. Toxicity of Vibrio alginolyticus on the survival of the rotifer Brachionus plicatilis. Nippon Suisan Gakkaishi 56, 1455–1460.