www.elsevier.nlrlocateraqua-online
Influence of daylength on growth, heterogeneity,
gonad development, sexual steroid and thyroid
levels, and N and P budgets in Perca flu
Õ
iatilis
S. Jourdan
a, P. Fontaine
a,), T. Boujard
b, E. Vandeloise
c,
J.N. Gardeur
a, M. Anthouard
d, P. Kestemont
ca
Laboratoire de Sciences Animales, INPL-UHP Nancy 1, MAN, 34 rue Sainte-Catherine, 54000 Nancy, France
b
Laboratoire de Nutrition des Poissons, Unite mixte INRA-IFREMER, Station INRA d’Hydrobiologie,´
64310 Saint-Pee-sur-Ni´ Õelle, France
c
Unite de Recherche en Biologie des Organismes, Facultes Uni´ ´ Õersitaires N.D. de la Paix,
61 rue de Bruxelles, B-5000 Namur, Belgium
d
Laboratoire d’Ethologie et de Neurobiologie, UniÕersite Louis Pasteur, 12, rue Goethe,´
67800 Strasbourg, France
Accepted 7 November 1999
Abstract
A 4-month study was conducted to determine the influence of daylength on growth, hetero-geneity, gonad development, sexual steroid and thyroid levels, and N and P budgets in Eurasian
Ž . Ž .
perch juveniles Perca fluÕiatilis . Fishes mean weight: 5.1 g were reared in tanks in
recirculat-y3 Ž .
ing systems at an initial biomass of 1 kg m under three different daylengths DL , 12, 18, and 24 h, including a simulated dawn and dusk phase. Perch were fed by demand feeders. Growth was
Ž .
checked monthly 100 fish per tank , carcass analyses and sex determinations were made on an average of 20 fish per tank with at least five small, five medium and five large fish. The increase
Ž
of DL, from 12, 18, or 24 h, induced significantly higher specific growth rates 1.78"0.17% dayy1 and 1.77"0.06% dayy1 for the 18- and 24-h groups, respectively, vs. 1.36"0.12%
y1 .
day for the 12-h group . After 112 days, a lower coefficient of variation in weight was found in
Ž . Ž
the 12-h group 40.3"2.6% , compared to the 18- and 24-h groups 50.2"6.6% and 54.1"4.1%,
. Ž
respectively . However, continuous daylight decreased survival in a significant way 76.0"7.4%
.
for the 24-h group, vs. 89.0"3.2% and 88.2"3.3% for the 12- and 18-h groups, respectively . The increase of DL from 12, 18, and 24 h also inhibited gonad development, especially in males.
)Corresponding author. Tel.:q33-3-83-30-28-41; fax:q33-3-83-32-30-16.
Ž .
E-mail address: [email protected] P. Fontaine .
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
However, for females and males, there was no significant DL effect on plasma sexual steroids and thyroid hormones levels. Likewise, no DL effect on N and P retention and loss was found. From this study, it appears that manipulating DL is a promising tool for improving rearing performance of Eurasian perch.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Eurasian perch; Perca fluÕiatilis; Daylength; Growth; Weight variability; Gonad development;
Food efficiency
1. Introduction
Ž .
The Eurasian perch Perca fluÕiatilis L. has been described recently as a new
Ž .
candidate for intensive freshwater aquaculture Kestemont and Dabrowski, 1996 . However, this species shows large variations in growth between fish of the same cohort ŽMelard et al., 1995 and significant mortality rates due to cannibalism Kestemont et
´
. Ž. Ž .
al., 1995 . These led to economic losses in aquaculture Smith and Reay, 1991 . Growth Ž
heterogeneity is considered to be caused by initial size heterogeneity Loadman et al., . 1986; Katavic et al., 1989; Van Damme et al., 1989; Folkvord, 1991; Brabrand, 1995 ,
Ž . Ž
density Baardvik and Jobling, 1990; Melard et al., 1996 , social interactions Abbott et
´
. Ž
al., 1985; McCarthy et al., 1992; Jobling et al., 1993 or genetic factors Bry and Gillet, .
1980; Kindschi and MacConnell, 1989 . It was recently reported for Eurasian perch that
Ž . Ž .
rearing conditions Fontaine et al., 1996a and temperature Melard et al., 1995, 1996
´
can also affect variation in growth.Considering that Eurasian perch show a typical diurnal feeding activity when fed on
Ž .
demand with self-feeders Anthouard and Fontaine, 1998 , one might hypothesise that daylength will influence its growth and heterogeneity, as it has already been shown in
Ž . Ž
other perciform species such as juvenile sunfish Lepomis cyanellus Gross et al., .
1965 .
Other effects of daylength may relate to growth heterogeneity and cannibalism, but whether this latter parameter is a cause or a consequence of the former is not known.
Ž .
Indeed, daylength can also act on gonad development De Vlaming, 1972 , which in turn Ž
may induce changes in body and muscle composition Norton and Macfarlane, 1995; .
Jørgensen et al., 1997 . In addition, daylength may interfere with feeding activity rhythm
Ž . Ž
and feed intake Barlow et al., 1995 and, in turn, may affect nutrient retention Boujard
. Ž .
and Leatherland, 1992; Boujard and Luquet 1996 and loss Thorpe and Cho, 1995 . Consequently, the purpose of our study was to investigate the influence of three daylengths on growth and growth heterogeneity in Eurasian perch juveniles reared in recirculation systems, with particular attention to physiological variables, such as feed efficiency, plasma thyroid and sexual steroid levels, and N and P balance.
2. Materials and methods
2.1. Fish, facilities and water quality
Ž .
In April 1997, a ribbon of perch eggs was harvested at Lindre pond Moselle, France
Ž .
hatching, larvae were introduced to a 500-l tank and fed Artemia nauplii for the first 30
Ž .
days, then weaned and fed a pelleted diet Biomar, Aquastart 15.50 no. 01 for 2 months. Juvenile perch were then transfered to two 1750-l tanks and reared at 20–228C
Ž . Ž .
under 12 h light 100 lux at the water surface :12 h dark photoperiod LD 12:12 . At the Ž
beginning of the experiment, the population 2660 perch, 150 days old, mean weight:
. Ž y3
5.1 g was divided into nine batches of 280–310 juveniles initial biomass: 1 kg m , .
initial coefficient of variation in weight CVi: 25–30% and then randomly allotted to Ž
nine blue-walls tanks of 1750 l each, functioning as recirculation systems Fontaine et .
al., 1996b . The experiment lasted 16 weeks from September 1997 to January 1998. During the experiment, the physico-chemical parameters of the water were checked three times a week. Water temperature was maintained at 23.0"0.48C, optimum for
Ž .
perch growth according to Melard et al. 1996 . Dissolved oxygen content and pH were
´
y1 Ž y1. Ž . Ž .
always above 7.0 mg l 7.6"0.3 mg l mean"range and 7 7.1"0.1 , respectively. The N–NHqand N–NOy levels were measured with indophenol blue and
4 2
Ž .
sulfanilamide methods, respectively Cary I spectrophotometer, Varian . The values remained between 0.00 and 0.13 mg N–NHq4 ly1, and between 0.00 and 0.15 N–NOy2
mg ly1.
2.2. Lighting conditions
Ž
The lighting of each tank was provided by incandescent white bulbs OSRAM L, 18 .
W, type 12-950 . Each tank was separated from the others by a plastic frame. After switching off, light intensity was nil. Light intensity, measured at the centre of the square tank and at the water surface, was set to 100 lux. This value was in the range of
Ž
optimal values for a similar species, the yellow perch Perca flaÕescens Hinshaw,
. Ž . Ž
1986 . Perch were subjected to three different daylengths DL : a 12-h L group light on
. Ž .
at 0800 h , a 18-h L group light on at 0500 h and a 24-h L group. Each group comprised three replicates. In the 12- and the 18-h groups, DL was controlled by electrical clocks and a switch phase of 30 min, when light intensity was reduced to 10 lux. This light intensity was provided by a 15-W bulb located above the water surface of each tank. This phase simulated dawn and dusk, known as the most adequate period for
Ž .
Eurasian perch feeding in their natural environment Thorpe, 1977; Dabrowski, 1982 .
2.3. Feeding
Ž
Perch juveniles were fed with an extruded diet Biomar, Ecolife 15.50 no. 2, 46% .
proteins, 15% lipids according to the manufacturer . Each tank was equipped with an
Ž .
electronic self-feeder Anthouard and Wolf, 1988 . The following procedure was applied: during the first week, the lever was hit artificially every 15 min from 0800 to 1900 h. Then, the number of artificial hits was reduced by 25% every 2 days. However, this level of artificial hits was kept until day 56 in tanks where fish did not activate the rod themselves, i.e., the 12-h group. During the first week of the learning phase, perch Ž . Ž . y0 .24
period of the experiment and checked every week. The amount of feed remaining in the feed dispenser was checked twice daily at 0800 and 1800 h, and special care was taken to always maintain filled feed dispensers.
2.4. Growth, sex determination and gonado-somatic indices
Mortality was recorded daily and each dead fish was weighed. An assessment of causes of mortality was performed through observation of external symptoms, such as necrosis, injuries, enucleation or regurgitation. A growth control was performed at day Ž .D 56 and 84 by individually weighing 100 fish to the nearest 0.1 g for each replicate,
Ž y1.
after anaesthesia 2-phenoxyethanol C H O ; 0.3 ml l8 10 2 . Mean body weights and
Ž .
coefficients of variation in weight were calculated. At the end of the experiment D112 , all fishes were counted and individually weighed. Survival rate, growth, growth
hetero-Ž .
geneity variation in individual weights among fish of the same replicate and feed efficiency were calculated as follows:
Final coefficient of variation in weight CVf, % s100=SD=Wf
Ž . Ž . y1
Feed efficiency FE s BfqBdyBi =Feed supply
where N and N are the initial and final fish number, W and W are the initial and finali f i f mean wet body weight, SD is the final standard deviation for weight, B and B are thei f
initial and final fish biomass, and N and B are the number and the biomass of deadd d
fish collected during the course of the experiment, respectively.
At the end of the experiment, the fishes were killed with an excess of 2-pheno-xyethanol. Sexes were determined for an average of 30 fish per tank. For each batch and
Ž .
both sexes, the average final value of gonado-somatic index GSI % was calculated:
GSIs100=GW=Wy1
Ž . Ž .
where GW is gonad weight g and W is body weight g .
2.5. Assays of sexual steroid and thyroid hormones
Ž .
On D112, a 1-ml blood sample was taken from the hearts of 10 fish per tank . Using
Ž .
a heparinized syringe, the plasma was separated 4000 rpm, 25 min then stored at
Ž . Ž .
y258C until assayed. Plasma concentrations of testosterone T , 17b-estradiol E2 and
Ž .
11-keto-testosterone 11 KT were assayed by means of the RIA analysis after two
Ž .
extractions with cyclohexanerethyl acetate according to Fostier and Jalabert 1986 . The
Ž .
inter-assay coefficients of variation were 10.8%, 7.9% and 7.7% for T ns5 , E2
Žns5 , and 11 KT n. Ž s5 , respectively. The intra-assay coefficients of variation were.
Ž . Ž . Ž .
14.2%, 12.1% and 8.9% for T ns5 , E2 ns5 , and 11 KT ns5 , respectively.
Ž . Ž .
Ž . RIA using commercial kits T RIA kit, Monobind, USA and T RIA kit, Pantex, USA .3 4
Ž .
The inter-assay coefficients of variation were 10.9 and 11.3% for T3 ns10 and T4
Žns14 , respectively. The intra-assay coefficients of variation were 9.9 and 10.6% for.
Ž . Ž .
T3 ns10 and T4 ns14 , respectively.
2.6. N and P balance study
An initial pooled sample of 100 g of fish, and a final sample of approximately 20 fish from each replicate, were crushed, homogenized and freeze-dried before composition analyses. Analyses of diet and whole-body samples were made following the usual
Ž .
procedures: dry matter after drying at 1058C for 24 h; nitrogen proteinsN=6.25 by the Kjeldahl method after acid digestion, total phosphorus by spectrophotometric analysis of the phosphovanadomolybdate complex after mineralization and acid
diges-Ž .
tion ISOrDIS 6491 method .
N and P retention and loss were deducted from N and P supply, carcass and feed N and P content:
Ž . ŽŽŽ . Ž . Ž
Retention % s100= BfqBd = final carcass nutrient content y Bi=initial
.. y1
carcass nutrient content =Nutrient supply ;
Ž y1. Ž
Total loss g=kg fish gain s1000=Nutrient supplyy Bf=final carcass
nutri-. Ž .. y1
ent content y Bi=initial carcass nutrient content =Gain
Ž y1. Ž ŽŽ
Faecal and metabolic loss g=kg fish gain s1000= Nutrient supplyy Bfq
. . Ž . Ž y1
Bd =final carcass nutrient content y Bi=initial carcass nutrient content = Gain
Ž y1.
Loss due to mortality g=kg fish gain sTotal lossyFaecal and metabolic loss Ž .
with Nutrient supply g sFeed supply=Feed nutrient content, and GainsBfyB .i
2.7. Data analysis
Ž Each parameter was analysed by a one-way ANOVA with daylength effect DL:
.
dfs2; error term: dfs6 . For parameters which had values fluctuating with time, a one-way ANOVA was performed at each time to verify the hypothesis of the normality of the variance; then, the model used to analyse data was a hierarchical mixed model Žprocedure MIXED, SAS Software; Little et al., 1996 :.
Yi jk lsm qDLi qTimej qTime)DLi j qTankkŽi. qTime)TankjkŽi.
df :35s 2 3 6 6 18
Ž DL and time effect were considered as fixed effects and the hierarchical effect tank wDLx. as the reference means square in testing the DL effect. The data were analysed with a split plot ANOVA. Finally, influence of daylength and sex on mean weights and hormonal levels were tested through a hierarchical model with a split plot ANOVA:
Yi jk lsm qDLi qSexj qDL)Sexi j qTankkŽi. qSex)TankjkŽi.
Ž
DL and sex are fixed effects sex is considered as being naturally randomised between .
fish within each tank . Consequently, the analysed variables corresponded to Ls means Ž"SD of measures carried out per experimental unit, adjusted by effects of other.
Ž factors. For one-way ANOVAs, the means were tested by a Newman–Keuls test fit for
.
balanced design . For the split-plot ANOVAs, the Ls means were tested by a Fisher test with the contrast method. In all tests, a significance level of P-0.05 was used.
3. Results
3.1. SurÕiÕal and growth
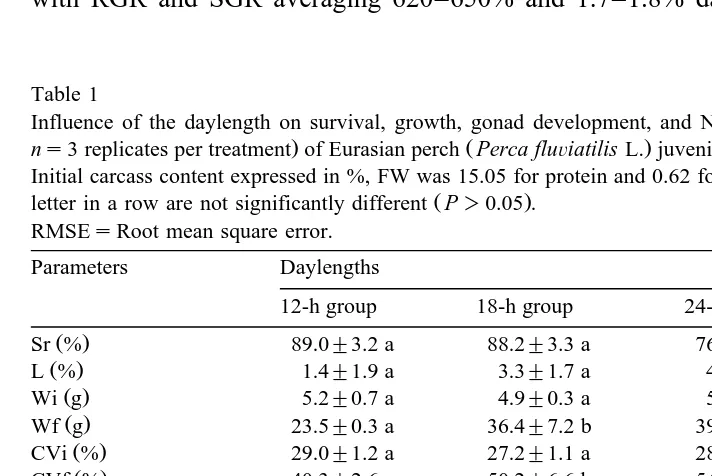
The 12- and 18-h groups have significantly higher survival rates than the 24-h group ŽP-0.05. ŽTable 1 . In all tanks, mortality was caused mostly by social interactions.
Ž
leading to high disappearance, regurgitation and enucleation rates 61"16% of total .
mortality .
Ž . Ž .
Relative RGR and specific growth rates SGR were significantly affected by DL, with RGR and SGR averaging 620–650% and 1.7–1.8% dayy1 for both the 18- and
Table 1
Ž
Influence of the daylength on survival, growth, gonad development, and N and P budgets Ls means"SD;
. Ž .
ns3 replicates per treatment of Eurasian perch Perca fluÕiatilis L. juveniles reared in recirculation systems
Initial carcass content expressed in %, FW was 15.05 for protein and 0.62 for P. Values followed by the same
Ž .
letter in a row are not significantly different P)0.05 . RMSEsRoot mean square error.
Parameters Daylengths RMSE
12-h group 18-h group 24-h group
Ž .
24-h groups, respectively, and significantly higher P-0.05; Table 1 than those of the
Ž .
12-h group. From D56 onwards, a highly significant difference P-0.01 was also recorded between the mean body weights of the 12-h group and the 18- and 24-h groups ŽFig. 1 . No growth difference was observed between the 18- and 24-h groups. This. enhancement of growth with DL increase did not significantly affect the feed efficiency Ž0.80"0.05, 0.74"0.03, and 0.74"0.06, respectively, for the 12-, 18- and 24-h
. groups .
3.2. Weight heterogeneity, sexual growth dimorphism and gonad deÕelopment
Ž .
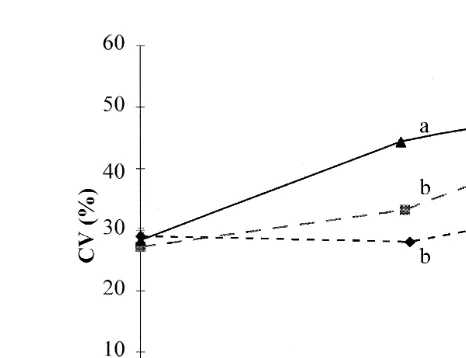
The analysis of the coefficient of variation in weight Fig. 2 showed a significantly
Ž .
higher CV for the 24-h group at D56 P-0.05 compared to the 12- and 18-h groups.
Ž .
At the end of the experiment Table 1 , there was a significant difference between the 12-h group and the 18- and 24-h groups. This lower weight heterogeneity in the 12-h group, compared to that of the 18- and 24-h groups, developed from D84 onwards ŽP-0.05; Fig. 2 ..
The necropsy made at the end of the experiment indicated that the sex ratio was Ž similar in all groups with a percentage of females slightly higher than males 55.0"
. 6.1%, 54.7"11.0%, and 52.5"10.7% for the 12-, 18-, and 24-h groups, respectively .
Ž .
A highly significant effect P-0.01 of DL on the mean weight of males and females was recorded. The mean weights of males and females from the 12-h group were
Ž .
significantly lower than those of the 18- and 24-h groups Table 1 . However, for a Ž given DL, mean weights of males and females showed no statistical difference P
-.
0.05 , regardless of treatment. At the end of the experiment, males of the 12-h group had
Ž .
a significantly higher GSI P-0.05 compared to the males of the 18- and 24-h groups
Ž .
Fig. 1. Growth of juvenile Eurasian perch mean weightqSD; ns3 replicates per treatment reared in
Ž .
recirculation systems and submitted to three different DL 12, 18, and 24 h over 112 days. Values followed by
Ž .
Ž .
Fig. 2. Coefficient of variation in individual weight Ls meansqSD; ns3 replicates per treatment of juvenile
Ž .
Eurasian perch reared in recirculation systems and submitted to three different DL 12, 18, and 24 h over 112
Ž .
days. Values followed by different letters at a same day are significantly different P-0.05 .
ŽTable 1 . There was also an influence of DL on the female GSI with an average 24-h.
Ž .
group female GSI significantly lower P-0.05 than the 12- and 18-h ones. Males of
Ž .
the 12-h group had a higher GSI than the females of the same group P-0.05 , whereas it was the opposite in the 18- and 24-h groups.
3.3. Sexual steroids and thyroid hormones
The results were summed up in Table 2. For females, there was no significant DL
Ž .
effect on E , T, T and T levels P2 3 4 )0.05 . The E levels remained low whatever the2
Table 2
Ž y1
Influence of the daylength on steroid and thyroid levels ng ml , Ls means"SD, ns3 replicates per
. Ž .
treatment of Eurasian perch P. fluÕiatilis L. juveniles reared in recirculation systems
Ž .
In a same line, values followed by a same letter are not significantly different P)0.05 . RMSEsRoot mean square error.
Parameters 12-h group 18-h group 24-h group RMSE
Females
E2 0.64"0.14 a 0.54"0.13 a 0.49"0.15 a 0.14 T 3.57"2.07 a 2.04"1.14 a 2.90"1.95 a 1.77 T3 11.75"2.57 a 12.03"1.72 a 10.84"1.99 a 2.12 T4 14.43"2.03 a 14.29"0.87 a 14.35"0.95 a 1.39
Males
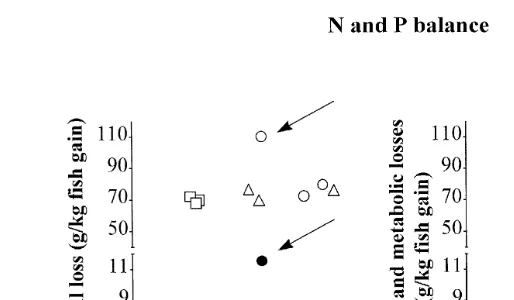
Ž .
Fig. 3. Relationship between individual gain and N and P loss mean, ns15 fish per batch in batches of
Ž .
juvenile Eurasian perch submitted to three different DL 12, 18, and 24 h . Left graph, total loss; right graph, faecal and metabolic loss. In left graph, arrows indicate the values obtained for one of the replicate submitted to a daylength of 24 h and in which mortality rate was high.
Ž y1.
daylength between 0.01 and 1.47 ng ml . Average female T levels were also low, around 3 ng mly1, although they reached 8 ng mly1 in some sampled females. For
Ž .
males, there was no treatment effect on T, 11-KT, T and T levels P3 4 )0.05 .
3.4. N and P balances
Ž .
Neither the protein nor the P content of the carcasses was affected by DL Table 1 . No effect of DL on N and P retention was significant. In addition, there was no
Ž .
treatment effect on N nor P loss Table 1 . An increase in standard deviation was observed for N and P total loss when DL is increased, but this increase was not observed when considering only the faecal and metabolic losses. In fact, it can be seen in Fig. 3 that this difference in standard deviation between total loss and faecal and metabolic losses should be attributed to one 24-h replicate, in which mortality was high. From Fig. 3, it can be seen also that there is no relationship between loss and individual gain. This statement is particularly obvious when considering only the faecal and metabolic losses.
4. Discussion
This study demonstrated that daylength significantly influenced the production vari-ables in Eurasian perch. Survival was also affected by continuous light compared to the
Ž .
12- and 18-h groups with aggressiveness cannibalism; enucleation as the main cause of death. Growth rates recorded in the 18- and 24-h groups were very close to those
Ž . Ž .
obtained by Melard et al. 1995 and Fontaine et al. 1997 with SGR of 1.7–1.8%
´
y1 Ž . Ž
day ; while shorter daylength 12 h reduced the growth performances of perch SGR:
y1. Ž .
Ž .
Lepomis cyanellus juveniles, by Huh et al. 1976 with yellow perch Perca flaÕescens
Ž .
juveniles and by Boehlert 1981 with Sebastes diploproa juveniles. These authors reported an increase of growth with daylength. However, there are data in the literature
Ž
for other species where no difference was found Fuchs, 1978 in Solea solea, Barlow et .
al., 1995 in Lates calcarifer . In our experiment, the difference in growth performance may be due to the initial delay in learning to use the triggering mechanism for demand feeding under 12-h DL. However, when we compared Eurasian perch growth in a
Ž .
similar situation same initial mean weight, good use of self-feeders , a lower SGR was still observed with a 12-h DL, indicating that the DL effect on growth was not only due to this initial delay.
In our experiment, there was no significant difference in growth rate between males
Ž .
and females. Fontaine et al. 1997 showed a sexual growth dimorphism in Eurasian
Ž . Ž
perch juveniles when the feeding level was increased from 1% or 2% to 3% 4 meals .
per day . In fact, in self-feeding conditions, perch juveniles regulate their feed intake and do not show any overconsumption of food with a feeding rate close to the optimum level
Ž . Ž y1.
suggested by Melard et al. 1996
´
2–2.5% day . However, weight heterogeneity increased with a significant effect of DL. The maximal heterogeneity in individualŽ .
weight CV: 54.1% was found under continuous light. In fact, final coefficients of
Ž .
variation for weight were very similar in 18- and 24-h groups 50–55% . Thus, as for growth, the DL effect on weight heterogeneity may be due to the initial delay in the learning of the triggering mechanism for demand-feeding.
If the DL had few effects on final weight between the sexes, it acted on gonad development. A 12-h DL is close to the natural daylength observed in September, when
Ž . Ž .
gametogenesis begins Sulistyo et al., 1998 . Thus, a decrease of DL from 24 or 18 h to 12 h favoured the gonad development in males, with the appearance of higher GSI. DL seemed to play a significant role in the onset of the testis development. These results confirm that males can develop their gonads as soon as they are 150–260 days old Ž5–25 g and that increasing DL prevents an early maturation of males Fontaine et al.,. Ž
.
1997 . In spite of the lack of DL effect on T and 11-KT plasma levels, the highest values were all measured in the 12-h group males. These values were similar to those observed under natural conditions in November–December and higher than those
Ž .
recorded during the sexual resting period in summer Sulistyo, 1998 . Our results showed that the ovarian development in female Eurasian perch was only slightly influenced by DL and continuous light significantly inhibited ovarian development. The final E2 and T levels were not significantly affected by DL and steroid values were
Ž .
strongly below the levels reported by Sulistyo et al. 1998 in female perch in September
Ž y1 y1 .
3–4 ng ml for E and 15–20 ng ml2 for T, respectively . These results confirms that the gonad development of perch females occurs later than in males. In this experiment, females were probably immature, explaining the low effect of DL on ovarian development. However, as observed in other species like turbot Psetta maxima ŽImsland et al., 1997 , an extended DL could delay the age at the first maturity in. Eurasian perch.
N and P retention were about 27–30% and 50%, respectively. N and P retention efficiencies vary according to fish species and weight, temperature, amount of feed
Ž . Ž .
Ž .
P retention by yellow perch Perca flaÕescens varies seasonally, with greater values in early summer. Therefore, it is difficult to compare the various retention values obtained. Nevertheless, our values are well within the range of available data in the literature. Among other examples, N retention was found to be slightly higher in salmonids Ž30–35%, Kim et al., 1998 ; in turbot Psetta maxima and cod Gadus morhua, with.
Ž
values in the range of 34–39% Houlihan et al., 1989; Burel et al., 1996; Mallekh et al.,
. Ž .
1999 ; and slightly lower in sea bass Dicentrarchus labrax 25–28%, Diaz et al., 1998
Ž .
and in sea bream Sparus aurata 20–25%, Lupatsch and Kissil, 1998 . Data on P retention are also very similar to our results, with values between 37% and 48% in
Ž .
turbot Mallekh et al., 1999 , 46% and 65% in rainbow trout Oncorhynchus mykiss ŽHardy et al., 1993 , and 26–45% in sea bream Lupatsch and Kissil, 1998 . The values. Ž . of N and P loss estimated in the present trial are also well within the range of already
Ž
published data with around 70–80 g of N and 7–8 g of P per kg fish gain Cho et al., .
1994; Kaushik, 1998 .
To the best of our knowledge, these data are the first available on Eurasian perch, indicating that this fish species is obviously not different from other cultivated species in terms of N and P retention and waste. Moreover, the fact that DL affects growth performance and sexual maturation but not N and P retention and loss, leads us to the conclusion that manipulating DL is a promising tool for improving rearing performances of Eurasian perch using demand feeders.
Acknowledgements
The study has been carried out with financial support from the Commission of the
Ž .
European Communities, Agriculture and Fisheries FAIR specific RTD programme, CT96-1572, ‘‘An integrated study of interindividual competition and its relationships with feeding physiology and behaviour in cultured predatory fish’’.
References
Abbott, J.C., Dunbrack, R.L., Orr, C.D., 1985. The interaction of size and experience in dominance relationships of juveniles Steelhead trout Salmo gairdneri. Behaviour 92, 241–253.
Ž .
Anthouard, M., Fontaine, P., 1998. L’auto-alimentation chez la perche Perca fluÕiatilis : adaptation a cette`
modalite de nourrissage, mise en evidence du rythme nycthemeral de la prise alimentaire. Ichtyophysiol.´ ´ ´ ´
Acta 21, 1–13.
Anthouard, M., Wolf, V., 1988. A computerized surveillance method based on self-feeding measures in fish populations. Aquaculture 71, 151–158.
Baardvik, B.M., Jobling, M., 1990. Effect of size-sorting on biomass gain and individual growth rates in Arctic charr SalÕelinus alpinus L. Aquaculture 90, 11–16.
Barlow, C.G., Pearce, M.G., Rodgers, L.J., Clayton, P., 1995. Effects of photoperiod on growth survival and
Ž .
feeding periodicity of larval and juveniles barramundi Lates calcarifer Bloch . Aquaculture 138, 159–168.
Boehlert, G.H., 1981. The effects of photoperiod and temperatures on laboratory growth of juveniles Sebastes
diploproa and a comparison with growth in the field. Fish. Bull. 79, 789–794.
Boujard, T., Leatherland, J.F., 1992. Circadian rhythms and feeding time in fishes. Environ. Biol. Fishes 35
Boujard, T., Luquet, P., 1996. Rythmes alimentaires et alimentation chez les siluroidei. Aquat. Living Resour.
Ž .
9, 113–120 Hors serie .´
Ž .
Brabrand, A., 1995. Intra-cohort cannibalism among larval stages of perch Perca fluÕiatilis . Ecol.
Freshwa-ter Fish 4, 70–76.
Bry, C., Gillet, C., 1980. Reproduction of cannibalism in pike Esox lucius fry by isolation of full-sib families. Reprod. Nutr. Dev. 20, 173–182.
Burel, C., Person-Le Ruyet, J., Gaumet, F., Le Roux, A., Severe, A., Boeuf, G., 1996. Effects of temperature´ `
on growth and metabolisme in juvenile turbot. J. Fish Biol. 49, 678–692.
Cho, C.Y., Hynes, J.D., Wood, K.R., Yoshida, H.K., 1994. Development of high-nutrient-dense, low-pollution diets and prediction of aquaculture wastes using biological approaches. Aquaculture 124, 293–305. Dabrowski, K.R., 1982. The influence of light intensity on feeding of fish larvae and fry: II. Rutilus rutilus
Ž .L. and Perca fluÕiatilis L. . Zool. Jb. Physiol. 86, 353–360.Ž .
De Vlaming, V.L., 1972. Environmental control of teleost reproductive cycles: a brief review. J. Fish Biol. 4, 131–140.
Diaz, J., Alvarez, M.J., Diez, A., Arzel, J., Corraze, G., Bautista, J.M., Kaushik, S.J., 1998. Regulation of
Ž .
hepatic lipogenesis by dietary proteinrenergy in juvenile European seabass Dicentrarchus labrax .
Aquaculture 161, 169–186.
Ž .
Folkvord, A., 1991. Growth, survival and cannibalism of cod juveniles Gadus morhua : effects of feed type, starvation and fish size. Aquaculture 97, 41–59.
Fontaine, P., Gardeur, J.N., Kestemont, P., Georges, A., 1997. Influence of feeding level on growth,
Ž .
intraspecific weight variability and sexual growth dimorphism of Eurasian perch Perca fluÕiatilis L.
reared in a recirculation system. Aquaculture 157, 1–9.
Ž .
Fontaine, P., Tamazouzt, L., Capdeville, B., 1996a. Growth of the Eurasian perch Perca fluÕiatilis L. reared
in floating cages and in water recirculated system: first results. J. Appl. Icthyol. 12, 181–184.
Fontaine, P., Terver, D., Georges, A., 1996b. Application of aquariological techniques to an intensive fish-rearing process using recycled, warmed water for the production of rainbow trout fry, Oncorhynchus
mykiss. Aquacult. Eng. 15, 485–498.
Ž .
Fostier, A., Jalabert, B., 1986. Steroidogenesis in rainbow trout Salmo gairdneri at various preovulatory stages: changes in plasma hormone levels and in vivo and in vitro responses of the ovary to salmon gonadotropin. Fish Physiol. Biochem. 2, 87–99.
Fuchs, J., 1978. Influence de la photoperiode sur la croissance et la survie de la larve et du juvenile de Sole´ ´
ŽSolea solea en elevage. Aquaculture 15, 63–74.. ´
Gross, W.L., Roelofs, E.W., Fromm, P.O., 1965. Influence of photoperiod on growth of green sunfish
Lepomis cyanellus. J. Fish. Res. Board Can. 22, 79–86.
Hardy, R.W., Fairgrieve, W.T., Scott, T.M., 1993. Periodic feeding of low-phosphorus diet and phosphorus
Ž . Ž .
retention in rainbow trout Oncorhynchus mykiss . In: Kaushik, S.J., Luquet, P. Eds. , Fish nutrition in practice. IVth Symposium in fish nutrition and feeding. INRA, Paris, pp. 403–411.
Hinshaw, J.M., 1986. Factors affecting feeding, survival and growth of larval and early juvenile yellow perch
ŽPerca flaÕescens, Mitchill . PhD Thesis, North Carolina State University, Raleigh, USA, 80 pp..
Houlihan, D.F., Hall, S.J., Gray, C., 1989. Effects of ration on protein turnover in cod. Aquaculture 79, 103–110.
Huh, H.T., Calber, H.E., Stuiber, D.A., 1976. Effects of temperature and light on growth of yellow perch and walleye using formulated feed. Trans. Am. Fish. Soc. 2, 254–258.
Imsland, A.K., Folkvord, A., Jonsdottir, O.D.B., Stefansson, S.O., 1997. Effects of exposure to extended
Ž
photoperiods during the first winter on long term growth and age at first maturity in turbot Scophtalmus
.
maximus . Aquaculture 159, 125–141.
Jobling, M., Jørgensen, E.H., Arnesen, A.M., Ringø, E., 1993. Feeding, growth and environmental require-ments of arctic charr: a review of aquaculture potential. Aquacult. Int. 1, 20–46.
Jørgensen, E.H., Johansen, S.J.S., Jobling, M., 1997. Seasonal pattern of growth, lipid deposition and lipid depletion in anadromous Arctic charr. J. Fish Biol. 51, 312–326.
Katavic, I., Jug-Dujakovic, J., Glamuzina, B., 1989. Cannibalism as a factor affecting the survival of
Ž .
intensively cultured sea-bass Dicentrarchus labrax fingerlings. Aquaculture 77, 135–143.
Kestemont, P., Dabrowski, K., 1996. Recent advances in the aquaculture of Percid fish. J. Appl. Ichthyol. 12, 137–200.
Kestemont, P., Fiogbe, E.D., Parfait, O., Micha, J.C., Melard, C., 1995. Relationship between weaning size,´ ´
growth, survival and cannibalism in the common perch larvae Perca fluÕiatilis: preliminary results. In:
Ž .
Larvi’95. Lavens, P., Jaspers, E., Roelants, I. Eds. , EAS Spe. Pub. 24pp. 285–288, Ostende, Belgium.
Ž
Kim, J.D., Kaushik, S.J., Breque, J., 1998. Nitrogen and phosphorus utilisation in rainbow trout
Oncor-.
hynchus mykiss fed diets with or without fish meal. Aquat. Living Resour. 11, 261–264.
Kindschi, G.A., Mac Connell, E., 1989. Factors influencing early mortality of walleye fry reared intensively. Prog. Fish Cult. 51, 220–226.
Kraft, C.E., 1992. Estimates of phosphorus and nitrogen cycling by fish using a bioenergetics approach. Can. J. Fish. Aquat. Sci. 49, 2596–2604.
Little, R.C., Milliken, G.A., Stroup, W.W., Wolfinger, R.D., 1996. SASw
System for mixed models. SAS Institute, Cary, NC, 633 pp.
Ž
Loadman, N.L., Moodie, G.E.E., Mathias, J.A., 1986. Significance of cannibalism in larval walleye
Stizosted-.
ionÕitreum . Can. J. Fish. Aquat. Sci. 43, 613–618.
Ž .
Lupatsch, I., Kissil, G.W., 1998. Predicting aquaculture waste from gilthead seabream Sparus aurata culture using a nutritional approach. Aquat. Living Resour. 11, 265–268.
Mallekh, R., Boujard, T., Lagardere, J.P., 1999. Evaluation of retention and environmental discharge of`
Ž .
nitrogen and phosphorus by farmed turbot Scophthalmus maximus . North Am. J. Aquacult. 61, 141–145. McCarthy, I.D., Carter, C.G., Houlihan, D.F., 1992. The effect of feeding hierarchy on individual variability in
Ž .
daily feeding of rainbow trout, Oncorhynchus mykiss Walbaum . J. Fish Biol. 41, 257–263.
Melard, C., Baras, E., Kestemont, P., 1995. Premiers resultats de l’elevage intensif de la perche europeenne´ ´ ´ ´
ŽPerca fluÕiatilis en bassin : effet de la temperature et du tri sur la croissance. Bull. Fr. Piscic. 336,. ´
19–27.
Ž
Melard, C., Kestemont, P., Grignard, J.C., 1996. Intensive culture of juvenile and adult Eurasian perch Perca´
.
fluÕiatilis : effect of major biotic and abiotic factors on growth. J. Appl. Ichtyol. 12, 175–180.
Norton, E.C., MacFarlane, R.B., 1995. Nutritional dynamics of reproduction in viviparous yellowtail rockfish,
Sebastes flaÕidus. Fish. Bull. 93, 299–307.
Porter, C.B., Krom, M.D., Robbins, M.G., Brickel, L., Davidson, A., 1987. Ammonia excretion and total N
Ž .
budget for gilthead seabream Sparus aurata and its effect on water quality conditions. Aquaculture 66, 287–297.
Smith, C., Reay, P., 1991. Cannibalism in teleost fish. Rev. Fish Biol. Fish. 1, 41–64.
Sulistyo, I., 1998. Contribution a l’etude et a la maıtrise du cycle de reproduction de la perche eurasienne` ´ ` ˆ
Perca fluÕiatilis L. These, Universite Henri Poincare, Nancy 1, 145 pp.` ´ ´
Sulistyo, I., Rinchard, J., Fontaine, P., Gardeur, J.N., Capdeville, B., Kestemont, P., 1998. Reproductive cycle and plasma levels of sex steroids in female Eurasian perch Perca fluÕiatilis. Aquat. Living Resour. 11,
101–110.
Thorpe, J., 1977. Daily ration of adult perch, Perca fluÕiatilis L., during summer in Loch Leven, Scotland. J.
Fish Biol. 11, 55–68.
Thorpe, J.A., Cho, C.Y., 1995. Minimising waste through bioenergetically and behaviourally based feeding strategies. Water Sci. Technol. 31, 29–40.
Van Damme, P., Appelbaum, S., Hecht, T., 1989. Sibling cannibalism in Koı carp, Cyprinus carpio L., larvae¨