TINJAUAN PUSTAKA
Kendala Budidaya Padi Gogo Pada Tanah Masam
Pertumbuhan dan produksi tanaman pada tanah masam tidak optimum akibat pengaruh langsung atau tidak langsung dari keracunan Al. Pengaruh keracunan Al secara langsung menyebabkan terhambatnya pemanjangan akar. Pengaruh tidak langsung terjadi karena Al mengganggu pengambilan, transpor, pengunaan hara dan air (Polle dan Konzak 1990; Ma 2000; Rout et al. 2001) yang menginduksi kekahatan hara dan cekaman kekeringan bagi tanaman (Samac dan Tesfaye 2003; Kochian et al. 2004).
Keracunan Al umumnya terjadi pada tanah masam dengan pH kurang dari 5.0, tetapi dapat juga terjadi pada pH 5.5 (Rout et al. 2001; Vitorello et al. 2005). Tingginya kelarutan Al pada pH tersebut menyebabkan Aldd meningkat sampai pada konsentrasi beracun bagi tanaman (Matsumoto 2000; Kochian et al 2004; Vitorello et al. 2005). Induksi kekahatan hara pada tanah masam terjadi karena sebagian daerah jerapan pada mineral liat dikuasai oleh Al3+ dengan menggantikan Mg2+ dan Ca2+. Al yang ada pada daerah jerapan tersebut juga dapat menjerap fosfat (P) dan molibdat (Mo) dengan kuat (Marschner 1995; Rout et al. 2001) sehingga P dan Mo tidak tersedia bagi tanaman.
Pada akar tanaman yang mendapat perlakuan Al dengan konsentrasi rendah, hanya apoplas atau dinding sel saja yang dipengaruhi oleh Al (Ma 2000). Sebagian besar Al di akar berada dalam dinding sel bagian luar sel kortek. Pemelaran dan kelenturan dinding sel merupakan prasyarat yang diperlukan untuk pembesaran dan pemanjangan sel. Interaksi Al dengan gugus karboksil dari senyawa pektat dinding sel menggantikan kation penstabil dinding sel seperti Ca (Tabuchi dan Matsumoto 2001) menyebabkan kekakuan dinding sel (Blamey dan Dowling 1995; Horst et al. 1999). Hal ini mengakibatkan pemelaran (Tabuchi dan Matsumoto 2001) dan kelenturan dinding sel menurun (Ma et al. 2004) sehingga pembelahan sel terhambat dan mengakibatkan pertumbuhan akar juga terhambat (Matsumoto 2000; Kochian et al. 2005).
Aluminium tidak hanya mempengaruhi dinding sel tetapi juga mengakibatkan kerusakan struktur membran plasma akar (Matsumoto et al. 1992). Interaksi Al dengan senyawa penyusun membran sel seperti lipid dan protein membran dapat memicu peroksidasi lipid yang dapat menyebabkan kehilangan integritas membran plasma (Yamamoto et al. 2003) dan membran plasma sel akar menjadi bocor (Ishikawa dan Wagatsuma 1998). Kerusakan fungsional membran plasma terjadi karena saluran masuk
Ca2+ (Huang et al. 1992), K+ (Liu dan Luan 2001) dan sisi tempat pengikatan (binding site) pada protein transpor Mg2+ plasma dihalangi oleh Al3+ (Rengel dan Robinson 1989) sehingga pengambilan Ca, K dan Mg oleh akar terhambat.
Proses transpor pada membran plasma dihambat oleh Al3+ dengan menggantikan Ca2+ dari asam pektat dinding sel yang dapat mengurangi pergerakan air dan hara melalui ruang antar sel (Blamey dan Dowling 1995). Interaksi Al dengan membran plasma mengurangi muatan negatif pada permukaan membran plasma yang me ngakibatkan gangguan H+ homeostasis dalam sitosol. Induksi deposisi kalus pada saluran plasmodesmata oleh Al menyebabkan transpor simplastik antar sel terhambat (Sivaguru et al. 2000).
Sebagian kecil Al dapat masuk ke simplas akibat kebocoran membran (La zof et al. 1996) dan terakumulasi dalam inti sel ujung akar (Silva et al. 2000; Taylor et al. 2000). Interaksi Al dengan isi simplastik mengganggu Ca homeostasis dan menghambat aktifitas enzim fosfolipase (Jones dan Kochian 1995). Pengikatan Al ke fosfat yang terdapat pada kedua utas DNA dapat menghambat replikasi DNA karena rigiditas helik ganda DNA meningkat (Ma 2000). Aluminium dapat menginduksi produksi reactive oxygen species (ROS) dalam mitokondria dan menyebabkan penurunan fungsi mitokondria (Yamamoto et al. 2003).
Secara morfologi, akar sekunder kelihatan pendek gemuk dan percabangannya berkurang, akar-akar adventif lebih banyak tumbuh pada leher akar (Rout et al. 2001) serta rapuh (Tan et al 1993; Rusdiansyah et al 2001). Akibatnya akar mengalami kesukaran untuk menembus lapisan tanah dan menghambat pengambilan hara dan air.
Keracunan Al pada padi menyebabkan terjadinya penghambatan pemanjangan akar (Khatiwada et al. 1996; Rusdiansyah et al 2001; Watanabe dan Okada 2005b). Hambatan pertumbuhan tajuk (Fageria et al. 1988) merupakan pengaruh sekunder akibat induksi kekahatan hara terutama Mg, Ca dan P serta induksi cekaman kekeringan sebagai akibat dari gangguan pertumbuhan dan aktivitas perakaran sehingga pertumbuhan padi menjadi kerdil (Coronel et al. 1990). Pada tahap selanjutnya keracunan Al dapat mengurangi pembentukan anakan (Hai et al. 1993), menghambat pertumbuhan batang (Howeler dan Cadavid 1976), menurunkan kesuburan, jumlah gabah per malai dan bobot 1000 butir (Alluri 1986).
Adaptasi Tana man pada Tanah Masam
Genotipe tanaman tertentu dapat bertahan hidup pada tanam masam karena mempunyai mekanisme untuk menenggang kelebihan Al yang ada dalam media tanam. Kochian (1995) mengusulkan dua mekanisme tenggang tanaman terhadap cekaman aluminium, yaitu mekanisme eksternal dan mekanisme internal. Perbedaan utama antara kedua mekanisme tersebut adalah tempat detoksifikasi Al, di apoplas untuk eksternal dan di simplas untuk internal.
Pada mekanisme eksternal, Al dicegah agar tidak menyeberangi membran plasma dan masuk ke simplas dan tempat yang sensitif terhadap Al di sitoplasma ujung akar. Kemungkinan mekanisme eksternal adalah permeabilitas membran plasma yang selektif untuk mengurangi pengikatan Al ke apoplas dan mengurangi pengambilan Al ke dalam sitosol (Okada et al. 2003; Archambault et al. 1996), peningkatan pH rhizosfer (Degenhardt et al. 1998), pelepasan asam organik sebagai ligan pengkelat Al (Ma 2000; Kochian et al. 2004; Mariano et al. 2005) dan immobilisasi Al di akar (Zheng et al. 2005). Bagian akar pertama yang dimasuki dan dilewati oleh Al agar dapat mencapai bagian meristematik yang sensitif adalah musilagel akar. Archambault et al. (1996) mendapatkan jika musilagel akar dibuang maka Al yang dijerap akar 25-35% lebih rendah dibandingkan akar yang masih bermusilagel. Hal ini menunjukkan bahwa lapisan musilagel akar menyebabkan Al tidak mobil di akar sehingga meristem akar akan terlindungi dari kerusakan akibat Al.
Kemampuan dinding sel akar menjerap Al yang rendah dan permeabilitas membran sel yang selektif terhadap Al diduga terlibat dalam mekanisme ketenggangan Al secara eksternal. Beberapa peneliti menunjukkan bahwa genotipe tenggang cenderung menyerap Al dalam jumlah kecil dibandingkan genotipe peka (Matsumoto 2000; Silva et al. 2000). Jumlah Al yang dapat dipertukarkan pada dinding sel akar padi varietas peka Al lebih tinggi dibandingkan varietas tenggang Al (Okada et al. 2003), sehingga kandungan Al pada akar padi tenggang Al lebih rendah dibandingkan padi peka Al (Watanabe dan Okada 2005b).
Tanaman yang mampu menaikkan pH di sekitar daerah perakaran akan terhindar dari keracunan Al. Peningkatan pH larutan pada Arabidopsis dapat meningkatkan ketahanan Al secara nyata (Degenhardt et al. 1998). Padi tenggang Al dapat me ngubah pH media tanam dari 4 menjadi 6.2 setelah 10 hari (Ganessan et al. 1993) dan dari 4 ± 0.1 menjadi 7.5 ±
0.1 setelah 30 hari mendapat cekaman Al (Sivaguru dan Paliwal 1993). Peningkatan pH di sekitar perakaran terjadi akibat peningkatan influx H+ pada ujung akar (Degenhardt et al. 1998). Peningkatan pH mengakibatkan pengendapan Al dan penurunan aktivitas Al3+ sehingga menjadi bentuk yang kurang beracun bagi tanaman (Samac dan Tesfaye 2003). Dengan demikian aktivitas Al menurun sebelum kontak dengan tempat metabolisme yang sensitif dalam akar.
Tanaman yang beradaptasi pada tanah masam, selain tenggang terhadap kelebihan Al dan juga tenggang terhadap NH4+ tinggi. Sorgum tenggang Al mempunyai nisbah penyerapan NO3-/NH4+ lebih tinggi dibandingkan sorgum peka (Kaltjens dan Ulden 1987). Penyerapan NO3- dalam jumlah besar menyebabkan terjadinya pelepasan ion hidroksil (OH-) atau ion bikarbonat (HCO3-) ke rhizosfer sehingga meningkatkan pH dan menekan kalarutan Al (Haynes 1990; Rout et al. 2001) sehingga tanaman akan terhindar dari keracunan Al.
Sejumlah tanaman dapat menghambat Al melalui pelepasan asam organik terutama malat, sitrat dan oksalat dari akar dalam merespon keracunan Al (Ma 2000; Mariano et al. 2005; Kochian et al. 2005). Over ekspresi enzim yang terlibat dalam metabolisme asam organik seperti sitrat sintase (Koyama et al. 2000) dan malat dehidrogenase (Tasfaye et al. 2001) pada tanaman transgenik dapat meningkatkan kandungan asam organik dan ketenggangan Al. Asam organik dapat mengkelat Al dengan membentuk komplek asam organik-Al yang tidak beracun bagi tanaman (Ma et al. 2001; Kochian et al. 2004). Namun demikian sekresi asam organik dari akar bukan mekanisme utama untuk ketenggangan Al pada padi (Ishikawa et al. 2000). Hasil penelitian Ma et al. (2002) menunjukkan bahwa tanaman padi tenggang dan peka Al yang ditanam pada kultur hara yang mengandung Al3+ melepaskan asam sitrat dalam jumlah yang hampir sama. Hal ini menunjukkan bahwa ketenggangan Al pada tanaman padi tidak berhubungan dengan pelepasan asam organik.
Bentuk adaptasi lain dari tanaman terhadap kondisi tanah masam berkaitan dengan penyerapan unsur-unsur hara ke dalam tanaman (Clark et al. 1990). Tanaman yang beradaptasi baik pada tanah masam mempunyai efisiensi penyerapan atau penggunaan hara atau keduanya yang sangat tinggi, terutama P, Ca dan Mg, agar dapat menghindari kahat hara yang diinduksi oleh cekaman Al (Fageria et al. 1988; Masrchner 1995; Baligar et al. 1997). Padi tenggang Al dapat menyerap dan menggunakan Ca (Watanabe dan Okada 2005b) dan P secara efisien (Sivaguru dan Paliwal 1993).
Silikat (Si) merupakan salah satu unsur hara yang bermanfaat bagi beberapa spesies tanaman termasuk padi (Epstein 1999). Pertumbuhan dan produksi biji tanaman pearl millet yang ditanam pada tanah masam sangat baik, didukung oleh konsentrasi Si di daun tinggi (Clark et al. 1990). Kandungan Si dalam daun sorgum tenggang Al yang ditanam pada tanah masam juga tinggi (Clark dan Gourley 1988). Silikat dapat mengurangi keracunan Al dan Mn pada sorgum (Galvez et al. 1987; Ma 2004).
Pada mekanisme internal, tanaman membiarkan Al masuk ke simplas dan tidak memperlihatkan gejala keracunan. Mekanisme ketenggangan Al internal umumnya dimiliki oleh spesies tanaman pengakumulasi Al seperti tanaman teh (Nagata et al. 1992), hydrangea (Ma et al. 1997a), buckwheat (Ma et al. 1998) dan melastoma (Watanabe et al. 1998). Selanjutnya Watanabe et al. (2006) melaporkan bahwa mekanisme ketenggangan internal juga dijumpai pada tanaman bukan pengakumulasi Al seperti tanaman pakan ternak Brachiaria hybrid.
Analisis 27Al NMR (Nuclear Magnetic Resonance Spectroscopy) menunjukkan Al membuat komplek dengan asam organik dalam simplas akar (Watanabe et al. 2006). Mereka melaporkan juga bahwa asam organik dalam jaringan akar membantu detoksifikasi Al pada Brachiaria hybrid. Ma dan Hiradate (2000) mendapatkan pada tanaman buckwheat, ion Al3+ melewati plasma membran melalui protein transpor, Al dikelat oleh asam oksalat yang ada dalam sitosol, selanjutnya ditranspor ke daun dalam bentuk komplek Al-sitrat melalui pembuluh xylem, selanjutnya Al dikompartementasi ke vakuola. Aktivitas transpor H+ tonoplas berperan penting dalam pertukaran proton dengan Al yang disimpan di vakuola. Taylor et al. (2000) mengamati bahwa Al dalam bentuk hidroksida Al dapat menembus tonoplas untuk masuk ke vakuola. Mereka juga mengamati pergerakan Al melewati membran plasma kemudian menuju tonoplas ke vakuola.
Penelitian terbaru mendapatkan sebanyak 17 protein responsif Al telah diidentifikasi, 12 diantaranya meningkat dan 5 lainnya menurun. Diantara protein yang meningkat akibat cekaman Al adalah copper/zinc superoksida dismutase (Cu-Zn SOD), dan S-adenosilmetionin sintase 2, sistein sintase (CS), 1-aminosiklopropana-1-karboksilat oksidase, G protein beta subunit-like protein, protein yang diinduksi ABA dan diinduksi cekaman, protein putative Avr9/Cf-9 rapidly elicited 141 kDa dan protein secretory 33 kDa merupakan protein terinduksi Al. Sebagian besar protein tersebut merupakan protein fungsional yang berkaitan dengan transduksi sinyal, antioksidan, dan deto ksifikasi. CS
konsisten terdeteksi dengan Western blot and asai aktivitas CS pada cekaman Al. Hasil metabolisme yang dikatalisis oleh CS seperti total pool glutathion dan glutathion tereduksi juga meningkat secara nyata dalam merespon cekaman Al. Mereka juga melaporkan bahwa antioksidan dan detoksifikasi berhubungan dengan metabolisme sulfur, terutama CS (Yang et al. 2007).
Penyakit Blas pada Tanaman Padi
Penyakit blas disebabkan oleh cendawan Pyricularia grisea (Cooke) Sacc, sinonimnya Pyricularia oryzae Cavara (Rossman et al. 1990). Penyakit blas merupakan penyakit utama pada padi gogo karena periode embun pada daun padi gogo lama dan kelembaban udara mencapai 90% sangat mendukung perkembangan penyakit blas (Amir dan Nasution 1995). P. grisea dapat menginfeksi daun pada stadia perkembangan vegetatif dan malai pada stadia generatif (Ou 1985).
Tingkat kerentanan padi gogo meningkat bila daun padi dalam keadaan lembab dalam waktu lama dan penyerapan hara yang tidak seimbang akibat kekeringan atau kemasaman tanah (Bonman 1992b). Kebanyakan areal pertanaman padi gogo miskin unsur hara makro dan kadang-kadang jenuh hara mikro tertentu, bereaksi masam dan kering seperti di Sitiung Sumatera Barat dan Tamanbogo Lampung (Amir dan Nasution 1995). Penyerapan unsur hara yang tidak seimbang akan mendukung perkembangan penyakit blas, terutama kelebihan unsur N (Ahn dan Amir 1986).
Infeksi pada daun dapat mengurangi luas daun hijau oleh bercak blas. Serangan blas daun pada varietas rentan dapat menyebabkan seluruh tanaman mati sebelum berbunga. Serangan blas leher dapat menurunkan hasil secara langsung karena leher malai busuk dan patah sehingga pengisian biji terganggu (Ahn dan Amir 1986) dan bulir padi me njadi hampa (Ou 1985). Kehilangan hasil pada varietas rentan dapat mencapai 50-90% (Amir dan Kardin 1991).
Cendawan P. grisea masuk ke dalam tanaman dengan cara penetrasi langsung melalui sel penjaga stomata (Oku 1994). Infeksi dimulai ketika konidia menempel pada permukaan padi karena ada perekat atau getah di ujungnya. Sebelum mempenetrasi jaringan tanaman padi, konidia berkecambah dengan membentuk tabung kecambah yang berupa apresorium. Apresorium dapat menghasilkan tekanan hidrostasis sebesar 8 MPa untuk me nembus dinding sel epidermis padi (Howard dan Valent 1996). Kemampuan
untuk menghasilkan tekanan hidrostatis tersebut berkaitan dengan terbentuknya melanin pada dinding sel apresorium (Chumley dan Valent 1990).
Apresorium tidak mampu mempenetrasi sebagian varietas tahan akibat adanya deposisi silikat pada dinding sel epidermis (Oku 1994). Hasil pengamatan dengan menggunakan mikroskop elektron menunjukkan bahwa lapisan silikat banyak dijumpai di bawah kutikula pada dinding sel epidermis daun padi (Kim et al. 2002). Lapisan silikat tersebut menjadi hambatan fisik bagi penetrasi hifa blas (Takahashi 1997).
Menurut Ou (1985), setelah penetrasi, lapisan silikat pada sel spidermis tidak dapat menghambat pertumbuhan miselia lebih lanjut. Namun demikian, varietas tahan dapat menghambat pertumbuhan cendawan dengan melokalisasi infeksi. Tanaman segera merespon infeksi blas pada tahap awal infeksi, sel yang terinfeksi dan beberapa sel di sekelilingnya segera menjadi coklat dan mati yang disertai matinya hifa (reaksi hipersensitif). Sebaliknya, respon terhadap infeksi pada varietas rentan lebih lambat yang ditandai dengan lambatnya timbul bercak coklat dan pertumbuhan hifa terus berlanjut.
Pencoklatan jaringan yang diinfeksi oleh P. grisea merupakan akibat dari oksidasi polifenol menjadi quinon yang beracun bagi cendawan (Ou 1985). Konsentrasi polifenol selalu lebih tinggi pada varietas tahan dibandingkan pada varietas rentan. Beberapa fitoaleksin diterpenoid seperti momilakton (Cartwright et al. 1981), oryzalesin (Akatsuka et al. 1985), fitokasan (Koga et al. 1995) telah diisolasi dari daun padi yang terinfeksi P. grisea. Akumulasi fitoaleksin tersebut berhubungan dengan peningkatan ketahanan tanaman padi terhadap penyakit blas (Dillon at al. 1997) karena dapat menghambat perkecambahan dan pertumbuhan tabung kecambah P. grisea (Huang 2001).
Fenomena kematian sel dengan cepat akibat adanya infeksi oleh patogen dikenal dengan reaksi hipersensitif (Oku 1994). Kematian hifa pada bercak hipersensitif disebabkan oleh jaringan tanaman terinfeksi yang mengeluarkan fitoaleksin sebagai akibat interaksi antara penyakit blas dan tanaman (Rodrigues et al. 2004). Fitoaleksin dapat mendetoksifikasi piricularin, suatu toksin yang dikeluarkan oleh P. grisea (Ou 1985). Dengan demikian reaksi hipersensitif dan kemampuan mensintesis fitoaleksin berhubungan dengan ketahanan terhadap blas.
Menurut Correa-Victoria dan Zeigler (1995) reaksi tanaman pada padi terhadap penyakit blas dibagi menjadi tiga yaitu tahan (completely resistant), moderat (partially resistant) dan rentan (susceptible). Cendawan tidak dapat menimbulkan bercak sama sekali
pada varietas tahan. Varietas yang mempunyai ketahanan parsial membiarkan cendawan untuk bereproduksi atau berspora tetapi tidak separah pada varietas peka.
Varietas tahan sempurna ditandai dengan respon hipersensitif terhadap infeksi dan cenderung tidak bertahan lama akibat peubahan populasi ras P. grisea (Seebold et al 2001; Marchetti 1983; Bonman 1992b) dan ketahanan dikendalikan oleh gen mayor tunggal (Kiyosawa 1982). Varietas tahan parsial mempunyai ketahanan lebih lama (Marchetti 1983) dibandingkan varietas tahan sempurna tetapi seleksi ketahanan parsial lebih sulit dilakukan karena dikendalikan oleh gen-gen minor yang belum dapat ditentukan jumlahnya (Yeh dan Bonman 1986) dan heritabilitasnya rendah (Wang et al. 1989).
Pengujian ketahanan padi terhadap blas bisa dilakukan di lapangan pada daerah endemik dengan inokulum alami atau di rumah kasa dengan inokulasi buatan (Ou 1985). Kelebihan pengujian di rumah kasa adalah benih yang diperlukan lebih sedikit dibandingkan dengan pengujian di lapangan, sehingga lebih sesuai untuk pengujian generasi awal pada siklus pemuliaan (Roumen 1993). Pengujian lapangan dapat dilakukan pada tahap lebih lanjut untuk mendapatkan varietas tahan di lapangan terhadap berbagai komposisi ras karena varietas yang dilepas ke petani akan menghadapi kondisi lingkungan yang multi ras (Amir 2002).
Hasil pemetaan ras P. grisea di daerah endemik blas di Jawa Barat dapat diidentififasi 30 ras. Ras 001 dominan di Kuningan, Ciamis, Garut, dan Tasikmalaya, ras ras 033 dominan di Sukabumi tetapi dijumpai juga di Kalijati, Ciamis, Garut, dan Tasikmalaya, sedangkan ras 173 dijumpai di Sukabumi (Amir et al. 2003). Hasil identifikasi pada musim hujan 2004/2005 pada daerah endemik blas di Lampung diperoleh ras 001, 033 dan 173 selalu dijumpai baik pada stadia anakan dan berbunga sebagai blas daun maupun pada stadia menjelang panen sebagai blas leher malai (Nasution et al. 2004).
Analisis dengan penanda RFLP dengan menggunakan pelacak MGR586 pada 13 ras dan 11 haplotipe isolat blas dominan di lapangan menunjukkan terdapat tiga kelompok utama. Kelompok I mewakili ras avirulen, menyebar luas dan mampu bertahan lama di lapangan yang diwakili oleh ras 001. Kelompok II mewakili ras yang sangat bervariasi virulensinya dan diduga mampu beradaptasi terhadap tekanan seleksi yang diwakili oleh ras 033. Kelompok III mewakili ras sangat virulen, tetapi tidak bertahan lama di lapangan yang diwakili oleh ras 173 (Utami et al. 2000).
Pemanfaatan Tanaman Haploid Hasil Kultur Antera dalam Pemuliaan Padi
Pada pemuliaan tanaman konvensional, varietas tanaman menyerbuk sendiri seperti padi terdiri atas genotipe-genotipe homogen dan homozigos. Hasil rekombinasi sifat-sifat yang diinginkan dari genom tetua yang disilangkan kemudian diseleksi pada generasi bersegregasi selanjutnya di-selfing 6 - 10 generasi untuk fiksasi gen agar diperoleh galur murni homozigos (Khush dan Virmani 1996; Dewi dan Purwoko 2001). Hal ini mengakibatkan pembentukan varietas memerlukan waktu lama.
Waktu untuk pelepasan varietas baru terbukti dapat dipersingkat dari 12 menjadi 5 tahun dengan memanfaatkan tanama n haploid ganda pada barley (Hu 1988). Genotipe homozigos tesebut dapat diperoleh secara langsung dalam satu generasi. Pada tanaman haploid ganda, hasil dan sifat-sifat kuantitatif lainnya yang memiliki heritabilitas rendah lebih cepat dapat dievaluasi dibandingkan yang diturunkan melalui metode pedigree (Khush dan Virma ni 1996; Bouchez dan Gallais 2000).
Efisiensi seleksi sifat kuantitatif dengan ragam aditif rendah pada tanaman haploid ganda juga lebih tinggi karena ragam dominan tidak muncul dan ragam aditif mengganda (Snape 1989). Hal ini dapat diterapkan pada pemuliaan padi gogo untuk ketenggangan terhadap cekaman aluminium dan ketahanan terhadap penyakit blas. Ketenggangan Al merupakan sifat kuantitatif yang melibatkan banyak gen (Ma et al. 2002; Nguyen et al. 2001; Wu et al. 2000). Ketahanan blas parsial dikendalikan oleh gen minor yang belum dapat ditentukan jumlahnya (Yeh dan Bonman 1986) dan heritabilitasnya rendah (Wang et al. 1989). Panjang akar relatif sebagai kriteria ketenggangan Al pada padi dikendalikan secara aditif dan pengaruh dominan (Khatiwada et al. 1996). Ketenggangan terhadap Al pada tahap awal pembibitan lebih dikendalikan oleh pengaruh aditif, sedangkan pengaruh epistasis lebih berperan pada tahap pembibitan lebih dewasa (Wu et al. 2000).
Secara konvensional, seleksi sifat yang dikendalikan oleh gen major akan efektif pada populasi F2 jika alelnya dominan, tetapi jika dikendalikan oleh alel resesif maka proporsi kombinasi alel yang akan muncul pada F2 sangat kecil yaitu (¼)n, dimana n adalah jumlah lokus. Akibatnya peluang fiksasi alel-alel yang diinginkan pada populasi yang dihasilkan rendah. Sebaliknya, dengan menggunakan populasi haploid ganda, genotipe tersebut akan muncul dengan frekuensi (½)n. Frekuensi fiksasi sifat pada populasi haploid ganda yang berasal dari F1 sama dengan akar kuadrat dari frekuensi pada populasi F2 (Snape 1989) sehingga populasi yang diperlukan untuk seleksi lebih sedikit. Metode
haploid ganda akan memungkinkan fiksasi alel-alel resesif yang diinginkan walaupun sifat tersebut dikendalikan oleh gen major.
Sifat-sifat tanaman haploid ganda stabil dari generasi ke generasi karena tanaman haploid ganda secara genetik identik dari satu generasi ke generasi berikutnya (Hu 1988; Zhang 1989; Sasmita 2006), sehingga seleksi hasil dan kualitas biji dapat dilakukan mulai dari generasi awal (DH0). Evaluasi sifat-sifat agronomi dapat dilakukan pada generasi pertama (DH1) dan DH2 (Dewi dan Purwoko 2001). Sifat-sifat yang dikendalikan oleh gen-gen minor seperti sifat ketenggangan terhadap cekaman botik atau abiotik dapat segera dievaluasi DH1 atau DH2 (Fehr 1987;Chung 1992).
Tanaman haploid ganda dapat diperoleh melalui produksi tanaman haploid yang dilanjutkan dengan penggandaan kromosom. Tanaman haploid merupakan tanaman yang memiliki jumlah kromosom sama dengan jumlah kromosom gametik karena berasal dari satu sel gametik di dalam kantong embrio atau tepung sari (Poehlman dan Sleper 1996).
Tanaman haploid dapat dimanfaatkan dalam pemuliaan tanaman karena beberapa kelebihan. Tanaman diploid homozigos dapat diperoleh dalam satu generasi dengan pengandaan kromosom, sedangkan pada pemuliaan konvensional diperlukan 7 sampai 8 generasi untuk mendapatkannya. Mutasi lebih mudah dapat terdeteksi karena sifat resesif juga diekspresikan. Genotipe gametik diekspresikan sepenuhnya pada tingkat tanaman, sehingga memungkinkan dilakukan seleksi (Roy 2000; Mukherjee 1999). Seleksi antar tanaman haploid untuk sifat yang dikendalikan oleh alel dominan tidak dikaburkan oleh sulitnya membedakan individu diploid homozigos dominan dan heterozigos (Fehr 1987).
Kultur antera merupakan metode yang banyak digunakan untuk memproduksi tanaman haploid (Jensen 1986). Kultur antera lebih sederhana dan efisien diantara metode produksi tanaman haploid yang ada (Hu 1988), sehingga dapat meningkatkan efisiensi program pemuliaan tanaman. Kelemahan penggunaan kultur antera dalam pemuliaan tanaman padi diantaranya adalah genotipe tetua memiliki daya kultur yang berbeda, diperoleh regenerasi tanaman albino. Antera tanaman F1 dari persilangan subspesies japonica memiliki daya kultur lebih tinggi dibandingkan persilangan indica/japonica dan indica (Dewi 2003; Chung 1992). Banyaknya regenerasi tanaman albino melalui kultur antera (Hanarida et al. 2002; Chung 1992) dapat diatasi dengan penggunaan poliamin pada tahap induksi dan regenerasi untuk meningkatkan regenerasi tanaman hijau (Dewi 2003; Purwoko et al. 2001; Purwoko et al. 2000). Galur yang dikembangkan melalui metode
haploid ganda memiliki kesempatan pengulangan rekombinasi lebih rendah dibandingkan galur yang dikembangkan melalui metode pedigree. (Chahal dan Gosal 2002; Roy 2000; Fehr 1987).
Sumber antera yang digunakan berasal dari tanaman F1 atau F2 agar diperoleh keragaman genetik maksimum antara tanaman haploid yang dihasilkan (Poehlman dan Sleper 1996). Apabila individu tanaman haploid tersebut digandakan kromosomnya maka akan diperoleh individu tanaman diploid homozigos untuk semua lokusnya. Galur haploid ganda hasil kultur antera tersebut memungkinkan untuk diseleksi terhadap sifat-sifat yang diinginkan.
Keberhasilan pemanfaatan tanaman haploid ganda asal kultur antera dalam pengembangan varietas unggul baru telah banyak dilaporkan berbagai spesies (Datta 2005) termasuk pada padi (Li 1992). Galur harapan dari persilangan antara tetua intermediet dan tetua yang membawa sifat tahan blas telah diperoleh melalui kultur antera (Liu et al. 1983). Tanaman haploid ganda asal kultur antera juga telah digunakan untuk memasukkan gen-gen pemulih kesuburan dari padi indica ke padi hibrida japonica (Shen et al. 1983). Berbagai varietas padi unggul yang dikembangkan melalui kultur antera juga telah dilepas di Cina, salah satu diantaranya adalah varietas Huan Yu No 1 yang dapat menghasilkan 7.5 ton/ha dan masih digunakan petani di Cina bagian Selatan (Li 1992).
Menurut Dewi et al. (1996), pemuliaan padi melalui kultur antera di Indonesia dimulai sejak tahun 1990-an. Namun demikian sampai saat ini (tahun 2007) belum ada satu varietas padi asal kultur antera yang telah dilepas. Purwoko et al. (2000) telah mendapatkan beberapa galur haploid ganda padi gogo asal kultur antera. Galur tersebut diduga membawa gen tenggang aluminium dan tahan blas, yang sangat strategis untuk dikembangkan untuk menjadi varietas unggul baru.
Metode Penapisan untuk Ketenggangan Aluminium
Berbagai metode penapisan ketenggangan Al telah diperkenalkan. Metode tersebut diantaranya adalah seleksi di lapangan pada lahan bercekaman Al (Sutaryo et al. 2005; Spehar dan Sauza 2006), pada kultur hara yang mengandung Al (Ma et al. 1997b; Khatiwada et al. 1996; Howeler dan Cadavid 1976), dalam pot dengan mengunakan media tanah masam (Giaveno dan Filho 2002; Toda et al 1999) dan media pasir dengan mengaplikasikan larutan hara yang mengandung Al setiap hari (Vilagarcia et al. 2001), dan
metode pewarnaan (Narasimhamoorthy et al. 2007), seleksi in vitro (Jan et al. 1997) serta dengan menggunakan penanda molekuler (Wu et al. 1997; Wu et al. 2000).
Pemilihan metode penapisan tergantung pada kemampuan laboratorium dan bahan tanaman yang akan diseleksi seperti koleksi plasma nutfah untuk identifikasi tetua yang sesuai, populasi yang bersegregasi, atau galur harapan yang akan dilepas (Carver dan Ownby 1995). Metode yang dipilih harus murah, pelaksanaannya cepat dan handal untuk menapis genotipe dalam jumlah banyak sekaligus (Samac dan Tesfaye 2003; Gupta 1997) dan dapat memisahkan antara genotipe tenggang dan peka Al (Ma et al. 1997b; Lewis dan Christiansen 1981).
Seleksi untuk mendapatkan genotipe tenggang Al dapat dilakukan secara langsung dan tidak langsung. Seleksi langsung terhadap hasil dilakukan di lapangan pada lingkungan bercekaman sesuai dengan lingkungan target. Tekanan seleksi yang homogen pada tanah masam di lapangan sangat sulit dicapai karena variasi tingkat keracunan Al sangat besar antar ruang pada satu lokasi (Giaveno dan Filho 2002; Lewis dan Christiansen 1981). Hal ini akan memungkinkan terpilihnya genotipe peka Al dan terbuangnya genotipe tenggang (Ma et al. 1997b).
Pada tanah masam di lapangan, dijumpai lebih dari satu cekaman dan faktor-faktor lain yang tidak dapat dikendalikan dan mempengaruhi pertumbuhan dan perkembangan tanaman (Spehar dan Sauza 2006). Cekaman pada tanah masam di lapangan tidak hanya keracunan Al, tetapi dijumpai juga keracunan Mn, kekahatan fosfat atau fosfat diikat oleh Al (Kochian et al. 2004; Samac dan Tesfaye 2003; Rout et al. 2001), kekeringan dan hama penyakit yang saling berinteraksi (Howeler dan Cadavid 1976).
Adanya berbagai interaksi antara genotipe dan lingkungan pada tanah masam di lapangan, akan menyulitkan untuk memisahkan antara pengaruh lingkungan dan genotipe (Blum 1988). Beberapa laporan menyebutkan bahwa heritabilitas pada lingkungan bercekaman lebih rendah dibandingkan pada lingkungan optimum (Calhoun et al. 1994; Banzinger at al 1997; Brancourt-Hulmel et al. 2005). Rendahnya heritabilitas pada lingkungan bercekaman akibat menurunnya keragaman genetik dan meningkatnya ragam galat (Ud-Din et al. 1992; Calhoun et al. 1994; Banzinger at al. 1997; Brancourt-Hulmel et al. 2005). Hal ini dapat diperbaiki dengan menyeleksi pada lingkungan yang lebih terkendali di rumah kaca.
Penapisan dengan menggunakan media tanah masam di rumah kaca akan memberikan tekanan seleksi yang hampir mendekati keadaan pada tanah masam di lapangan (Narasimhamoothy et al. 2007; Giaveno dan Filho 2002). Media tanam yang digunakan dapat diambil dari daerah target produksi dan memiliki hampir semua cekaman yang dijumpai di lapangan, kecuali cekaman kekeringan dan hama penyakit masih dapat dikendalikan. Pada percobaan pot dijumpai juga adanya hambatan mekanis untuk penetrasi akar ke dalam tanah. Kondisi lingkungan percobaan relatif terkontrol dibandingkan di lapangan (Spehar dan Sauza 2006; Makmur 2001). Tanaman dapat dipelihara sampai panen, bobot biji per rumpun dapat dijadikan sebagai kriteria seleksi langsung (Rusdiansyah 2002), dan dapat dijadikan sebagai tahapan penapisan diantara penapisan pendahuluan pada kultur hara dan penapisan di lapangan pada kondisi alami yang mahal (Carver dan Ownby 1995).
Kekurangan metode pot adalah media tanam yang akan digunakan harus tanah dengan kejenuhan Al tinggi yang didatangkan dari lapangan yang sangat bervariasi antar tempat dan waktu pengambilan tanah. Tanah yang digunakan sebagai media tanam harus dianalisis terlebih dahulu untuk menentukan unsur hara yang tidak cukup dan unsur yang berlebih yang pengaruhnya dapat mengaburkan pengaruh cekaman Al. Tanah masam dengan pH yang sama memiliki kejenuhan Al dan kandungan Mn berbeda (Narasimhamoothy et al. 2007). Hasil penapisan dengan media tanah kurang konsisten walaupun diulang pada tanah yang sama (Carver dan Ownby 1995). Di samping itu, pengaruh utama keracunan Al terhadap hambatan pemanjangan akar sukar diamati (Ma et al 1997b; Giaveno dan Filho 2002).
Seleksi ketenggangan Al pada generasi awal dalam program pemuliaan akan lebih efisien dilakukan pada lingkungan yang terkontrol. Pada generasi awal benih yang tersedia masih sedikit, tetapi genotipe yang akan diseleksi sangat banyak, sehingga diperlukan metode yang cepat, mudah dan murah seperti metode kultur hara yang mengandung Al atau dikombinasikan dengan metode pewarnaan akar (Narasimhamoothy et al. 2007).
Pada metode kultur hara, cekaman yang diberikan ke tanaman lebih mudah diatur untuk cekaman Al saja dan jumlah benih yang diperlukan lebih sedikit. Metode ini dapat digunakan untuk menapis genotipe dalam jumlah banyak, relatif mudah, menghemat waktu dan tenaga (Ma et al 1997b; Villagarcia et al. 2001; Samac dan Tesfaye 2003). Genotipe
tenggang dapat dipindahtanam ke tanah untuk dievaluasi kembali hingga panen (Howeler dan Cadavid 1976; Coronel et al. 1990).
Sifat yang dijadikan sebagai kriteria seleksi tidak langsung harus memiliki keragaman genetik yang tinggi, heritabilitasnya lebih tinggi dibandingkan hasil, berkorelasi dengan hasil pada kondisi tercekam, sifat tersebut mempunyai pengaruh terhadap hasil, mudah dan lebih cepat diamati dibandingkan hasil (Acevedo dan Fereres 1993). PAR, yang merupakan nisbah panjang akar maksimum pada perlakuan Al terhadap panjang akar maksimum pada perlakuan tanpa Al (kontrol), telah digunakan sebagai kriteria seleksi (Khatiwada et al. 1996; Wu et al. 1997; Wu et al. 2000; Rusdiansyah et al, 2001).
Heritabilitas dalam arti luas dan dalam arti sempit PAR pada tanaman padi tergolong tinggi dan pengaruh lingkungan rendah (Khatiwada et al. 1996; Wu et al. 1997). Dengan demikian penggunaan PAR akan memberi peluang untuk mendapatkan genotipe padi tenggang Al lebih cepat. Selain peubah PAR juga terdapat beberapa peubah lain yang dapat digunakan sebagai indeks ketenggangan Al seperti bobot kering akar atau tajuk (Fageria et al. 1988). Beberapa metode pewarnaan akar telah digunakan sebagai alternatif untuk pengukuran ketenggangan Al pada kultur hara. Diantaranya adalah hematoxylin pada gandum (Polle et al. 1978), eriochrome cyanin R pada gandum (Aniol 1995) dan barley (Ma et al. 1997b), lumogallion pada kedelai (Kataoka dan Nakanishi 2001) dan pyrocatechol violet (PVP) pada padi (Watanabe dan Okada 2005a).
Korelasi antara ketenggangan Al pada kondisi kultur hara dan ketenggangan Al pada kultur tanah sangat tinggi (Toda et al. 1999; Giaveno dan Filho 2002). Pengelompokan ketenggangan genotipe padi gogo terhadap Al pada metode kultur hara berkorelasi erat dengan tanggapan pertumbuhan dan hasil biji pada tanah masam di lapangan (Howeler dan Cadavid, 1976). Hal ini menunjukkan bahwa metode kultur hara dapat dimanfaatkan untuk penapisan pada tahap awal program dalam pemuliaan tanaman.
Kelemahan metode kultur hara adalah tidak dapat mewakili keseluruhan cekaman yang dijumpai pada tanah masam. Pemanjangan akar pada tanah diperlukan untuk mendapatkan hara yang letaknya jauh dari ujung akar, tetapi pada kultur hara, unsur hara sudah dalam bentuk terlarut dan tersedia bagi tanaman dan tidak adanya hambatan fisik untuk penetrasi akar seperti pada tanah. Genotipe yang peka terhadap genangan juga akan memperlihatkan reaksi peka Al, padahal belum tentu peka Al. Metode kultur hara tidak
cocok untuk penapisan tanaman yang diperbanyak secara vegetatif (Narasimhamoothy et al. 2007).
Hasil seleksi padi gogo dengan cara menanam langsung pada tanah masam (Asfaruddin 1997; Syakhril 1997) tidak selalu konsisten dengan hasil evaluasi pada larutan hara (Narasimhamoothy et al. 2007; Farid et al. 1997). Hal ini disebabkan sifat ketenggangan Al dikendalikan oleh banyak gen (Ma et al. 2002; Nguyen et al. 2001; Wu et al. 2000), sehingga tidak semua gen terekpresi pada satu metode penapisan. Evaluasi daya hasil pada keadaan tercekam Al di lapangan diperlukan karena varietas yang dilepas haruslah yang tenggang di lapangan. Penapisan genotipe tenggang Al akan lebih efisien jika dilakukan dengan mengkombinasikan metode penapisan kultur hara, media tanah masam dalam pot dan di lapangan (Narasimhamoothy et al. 2007; Spehar dan Souza 2006).
PENAPISAN GALUR HAPLOID GANDA PADI GOGO (Oryza sativa L.) UNTUK KETENGGANGAN ALUMINIUM
ABSTRAK
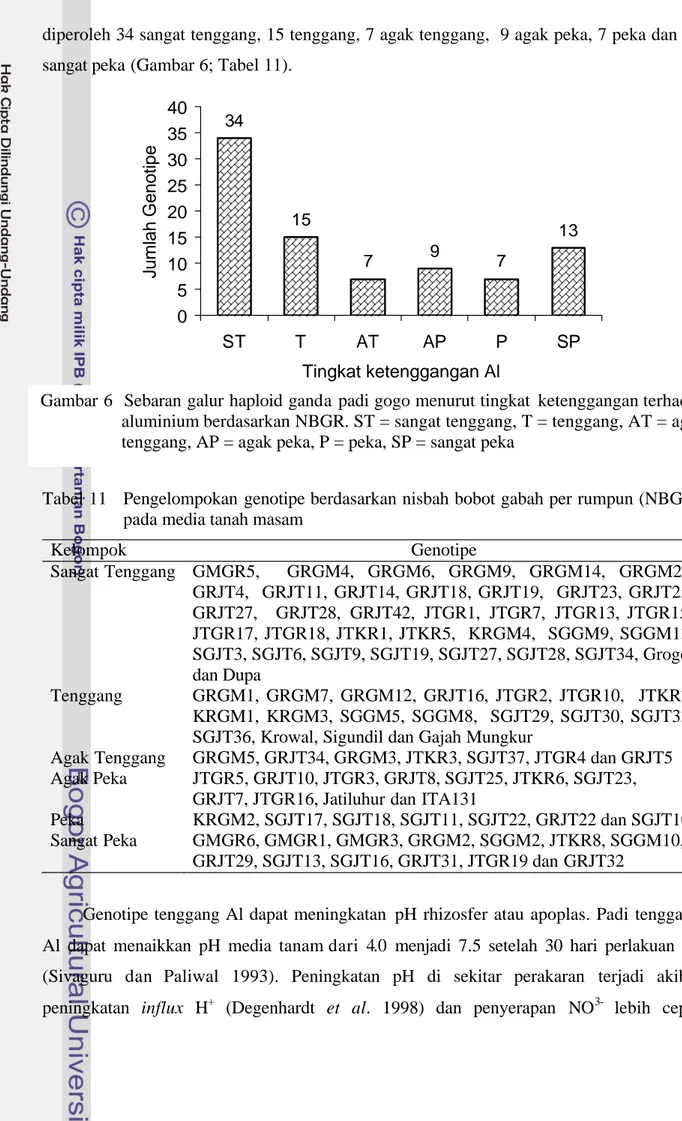
Keracunan Aluminium (Al) merupakan salah satu faktor penting yang menghambat produksi padi gogo pada tanah masam. Penggunaan genotipe tenggang Al merupakan pendekatan yang dapat mengatasi masalah tersebut. Galur haploid ganda hasil kultur antera telah diseleksi pada kultur hara yang mengandung Al berdasarkan panjang akar relatif (PAR) dan pada media tanah masam berdasarkan nisbah bobot gabah per rumpun (NBGR). Berdasarkan penapisan pada kultur hara, dari 113 galur haploid ganda yang diseleksi diperoleh 15 tenggang, 73 moderat dan 25 peka Al. Hasil penapisan pada media tanah masam, dari 85 galur diperoleh 34 sangat tenggang, 15 tenggang, 7 agak tenggang, 9 agak peka, 7 peka dan 13 sangat peka. Galur KRGM4, JTGR13, JTGR17, JTGR18, JTKR1, JTKR5, GRGM4, GRGM6, GRGM9, GRGM14, GRGM25, GRJT11 dan SGJT27 tenggang pada kedua metode penapisan. Korelasi antara tingkat ketenggangan Al pada kultur hara dan pada media tanah masam positif dan nyata. Jumlah anakan produktif dan jumlah biji per malai memiliki heritabilitas yang tinggi dan berpengaruh langsung terhadap hasil gabah serta berpengaruh tidak langsung terhadap hubungan antara hasil dan karakter lain. Seleksi hasil secara langsung pada kondisi tercekam Al lebih efisien dibandingkan seleksi tidak langsung melalui jumlah anakan produktif dan jumlah biji per rumpun.
ABSTRACT
Aluminum toxicity is an important growth limiting factor for upland rice production in acid soils. Al toxicity is not always economically corrected with soil liming. An alternative approach to solve Al toxicity would be to select or breed plant genotype tolerant to Al. The doubled haploid (DH) rice lines were screened under nutrient solution containing either 0 or 45 ppm Al and acid soils containing either low or high Al saturation. The relative root length (RRL) was determined at 14-day-old stage to characterize Al-tolerant genotypes in nutrient solution. The relative grain weight (RGW) was determined to characterize Al-tolerant genotypes in soil conditions. The differential tolerance for Al toxicity among genotypes was found to be highly significant for RRL in nutrient solution and RGW in acid soils. Among the 113 DH lines tested, 15, 73 and 25 lines were found to be Al-tolerant, moderate and susceptible, respectively based on RRL. Result of evaluation of DH lines for Al-tolerant in acid soils, showed that 34, 15, 7, 9, 7 and 13 DH lines were found to be highly Al-tolerant, tolerant, moderately tolerant, moderately susceptible, susceptible and highly susceptible in term RGW, respectively. KRGM4, JTGR13, JTGR17, JTGR18, JTKR1, JTKR5, GRGM4, GRGM6, GRGM9, GRGM14, GRGM25, GRJT11 and SGJT27 lines showed that they were consistently Al-tolerant under both nutrient solution and acid soils. The RLL of doubled haploid upland rice lines in nutrient solutions were strongly correlated with RGW in acid soils. Number of tillers and filled grain per panicle were inherited with high heritability and directly affected grain weight and contributed indirectly to the correlation between grain weight and other characters. However, indirect selection through number of tillers and filled grain per panicle was less efficient than direct selection through grain weight.
PENDAHULUAN
Kebutuhan beras nasional terus meningkat dengan cepat seiring dengan peningkatan jumlah penduduk. Selama ini, produksi beras sangat tergantung pada padi sawah beririgasi. Mengingat banyak lahan sawah telah dikonversi untuk keperluan non pertanian, lahan yang sesuai untuk persawahan baru terbatas dan biaya pembangunan sistem irigasi serta pencetakan sawah baru sangat mahal, maka alternatif peningkatka n produksi padi dapat dilakukan dengan perluasan areal pertanaman padi gogo pada lahan kering.
Lahan kering yang tersedia cukup luas dan tersebar di berbagai provinsi. Lahan kering yang sesuai untuk tanaman pangan diperkirakan mencapai 11 juta ha lebih, yang telah diusahakan mencapai 1.14 juta ha atau 10.5% (Puslitbangtan 2006). Lahan kering tersebut sebagian besar merupakan tanah jenis Podsolik Merah Kuning (PMK).
Keracunan Aluminium (Al) merupakan kendala utama untuk budidaya padi gogo pada tanah PMK. Kelarutan Al pada tanah PMK dengan pH kurang dari 5.5 sangat tinggi sehingga meningkatkan Al dapat dipertukarkan (Aldd) dan dapat beracun bagi tanaman (Kochian et al 2004; Vitorello et al. 2005).
Pendekatan budidaya untuk mengatasi keracunan Al pada tanah masam dapat dilakukan dengan pengapuran. Namun demikian, pendekatan tersebut tidak ekonomis terutama untuk daerah yang jauh dari sumber kapur dan sarana transportasi tidak baik. Alternatif lain yang berjangka panjang adalah perakitan varietas tenggang Al melalui program pemuliaan tanaman. Namun demikian diperlukan waktu yang lama untuk mendapatkan varietas tersebut.
Kultur antera dapat mempercepat perolehan tanaman homozigos untuk pengembangan varietas baru. Purwoko et al. (2000) telah membentuk 113 galur haploid ganda hasil persilangan antara beberapa varietas unggul dan aksesi plasma nutfah tenggang aluminium. Penapisan terhadap bahan genetik tersebut diperlukan untuk mendapatkan galur yang dapat beradaptasi pada lahan masam.
Metode penapisan dengan menggunakan kultur hara pada tahap pembibitan dapat memisahkan genotipe tenggang dan peka Al dalam waktu 14 hari (Jagau 2000). Metode tersebut relatif cepat dan sederhana untuk mengidentifikasi galur tenggang Al (Ma et al. 1997c; Khatiwada et al. 1996; Howeler dan Cadavid 1976) pada tahap awal program pemuliaan karena benih yang tersedia masih sedikit dan jumlah genotipe yang diseleksi sangat banyak (Samac dan Tesfeye 2003). Namun demikian penapisan pada kultur hara
tidak dapat mewakili cekaman yang dijumpai di lapangan seperti hambatan fisik untuk penetrasi akar. Untuk itu diperlukan penapisan secara langsung di lapangan pada lahan tanah masam yang dapat dilakukan pada tahap lebih lanjut apabila benih yang tersedia sudah cukup memadai.
Penapisan dengan menggunakan media tanah masam di rumah kaca akan memberikan tekanan seleksi yang hampir mendekati keadaan pada tanah masam di lapangan. Pada penapisan dengan media tanah masam akan ada hambatan mekanis untuk penetrasi akar ke dalam tanah. Kondisi lingkungan penapisan relatif terkendali dibandingkan dengan di lapangan (Spehar dan Sauza 2006; Makmur 2001). Tanaman dapat dipelihara sampai panen dan bobot biji per rumpun dapat dijadikan sebagai kriteria seleksi langsung.
Penelitian ini bertujuan untuk mendapatka n galur haploid ganda padi gogo hasil kultur antera yang tenggang Al pada kultur hara dan media tanah masam, serta mendapatkan informasi tentang karakter yang dapat digunakan sebagai penciri dalam penapisan galur haploid ganda pada keadaan tercekam Al.
BAHAN DAN METODE
Percobaan terdiri atas dua tahap penapisan, yaitu penapisan pada kultur hara dan pada media tanam tanah masam. Penelitian ini dilaksanakan di rumah kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetika Pertanian (BB-BIOGEN), Bogor. Penelitian tahap pertama dilakukan bulan Agustus 2003 sampai dengan Desember 2003. Penelitian tahap ke dua dilakukan bulan Januari 2004 sampai dengan Juli 2004.
Penapisan dengan Kultur Hara
Bahan tanaman yang digunakan pada kultur hara adalah 120 genotipe, terdiri atas 113 galur haploid ganda hasil kultur antera, 5 tetua asal persilangan dan dua pembanding yaitu Dupa dan ITA131 masing-masing sebagai pembanding tenggang dan peka Al. Varietas tetua digunakan adalah Krowal, Sigundil, Grogol, Jatiluhur dan Gajah Mungkur (Tabel Lampiran 1).
Media tanam yang digunakan pada perlakuan tanpa cekaman Al adalah larutan hara untuk tanaman padi yang disarankan Yoshida et al. (1976). Perlakuan cekaman Al dilakukan dengan penambahan 45 ppm AlCl3 ke larutan hara (Jagau 2000). pH larutan
diatur pada 4.0 ± 0.1 dengan penambahan NaOH 1 N atau HCl 1 N. Setiap pot diisi 2 liter larutan hara dan diberi aerasi supaya Al dan hara tidak mengendap. Air yang hilang akibat transpirasi diganti dengan menambahkan aquades setiap dua hari sekali dengan pH tetap dipertahankan sekitar 4.0 ± 0.1 dengan penambahan NaOH 1 N atau HCl 1 N.
Kecambah normal yang berumur satu minggu dengan panjang akar yang seragam dipindahkan ke media percobaan. Batang kecambah tersebut dibalut dengan gabus busa lunak kemudian dimasukkan ke lubang-lubang styrofoam yang telah disiapkan dan diapungkan pada larutan hara dalam pot. Setiap pot ditanami 5 kecambah selanjutnya dipelihara selama 14 hari di rumah kaca. Galur haploid ganda, tetua dan pembanding ditanam pada media tanam bercekaman Al dan tanpa cekaman Al. Rancangan percobaan yang digunakan adalah rancangan acak kelompok (RAK) dengan empat ulangan.
Pengamatan dilakukan terhadap panjang akar, tinggi tanaman, bobot kering akar, bobot kering tajuk dan dihitung relatif dari setiap peubah tersebut serta nisbah akar tajuk (NAT) pada umur 14 hari setetelah tanam (HST). Analisis ragam dilakukan terhadap semua genotipe yang diuji dan jika terdapat perbedaan yang nyata, setiap genotipe dibandingka n dengan varietas pembanding tenggang dengan uji BNT pada taraf 5%.
Penapisan dengan Media Tanah Masam
Bahan tanam yang digunakan sebanyak 95 genotipe terdiri atas 88 galur haploid ganda, 5 tetua dan 2 pembanding. Semua genotipe ditanam pada media tana m bercekaman Al dan tanpa cekaman Al. Rancangan percobaan yang digunakan adalah rancangan acak kelompok (RAK) dengan tiga ulangan.
Media tanam adalah tanah Podsolik Merah Kuning (PMK) yang diambil dari Jasinga dengan Aldd 17.14 cmol(+) kg-1 dan kejenuhan Al 79 % untuk perlakuan cekaman Al (Tabel Lampiran 2). Tanah tersebut dicampur dan diaduk rata kemudian diayak untuk menjamin keseragaman tanah. Selanjutnya tanah dimasukkan ke dalam polybag sebanyak 8 kg tanah per polybag. Perlakuan tanpa cekaman menggunakan tanah yang sama tetapi empat minggu sebelum tanam ditambahkan kapur sebanyak 1.5 x Aldd (setara 25.71 ton CaCO3 ha-1) untuk mengurangi kejenuhan Al. Penggunakan kapur sebanyak 1.5 x Al Aldd diperkirakan dapat menetralkan 85-90% Aldd (Hardjowigeno 2003).
Benih bernas dari setiap genotipe ditanam secara langsung ke media tanam yang telah disiapkan sebanyak 4 tanaman per polybag. Pemupukan diberikan dengan dosis 3 g Urea, 2 g SP36, 2 g KCl per polybag. Seluruh dosis pupuk SP36, dan KCl diberikan sehari
sebelum tanam sebagai pupuk dasar. Pupuk Urea diberikan 1/3 dosis pada umur 14 hari setelah tanam (HST), 1/3 dosis pada umur 42 HST dan sisanya pada saat inisiasi primordia bunga yaitu 55 HST. Penyiangan dan pengendalian hama dan penyakit dilakukan jika diperlukan. Tanaman dipelihara sampai panen.
Pengamatan dilakukan terhadap jumlah anakan total, jumlah anakan produktif (batang/rumpun), umur berbunga (hari) dihitung sejak tanam sampai keluar malai, umur panen (hari) dihitung sejak tanam sampai 80% malai siap dipanen, tinggi tanaman (cm) pada saat panen yang diukur mulai dari leher akar sampai leher malai, panjang malai (cm) diukur dari leher malai sampai ujung malai, jumlah gabah isi per malai (butir), persentase gabah hampa per malai (%), bobot 100 butir (g) dan bobot gabah per rumpun (g). Data tersebut digunakan untuk menghitung nisbah setiap peubah yaitu pertumbuhan pada Al tinggi terhadap pertumbuhan pada Al rendah.
Sidik ragam dilakukan terhadap setiap nilai nisbah untuk melihat perbedaan antar genotipe. Jika terdapat perbedaan yang nyata, setiap genotipe dibandingkan dengan varietas Dupa sebagai pembanding tenggang dengan uji BNT pada taraf 5%. Untuk memilih karakter agronomi yang sangat berpengaruh terhadap keragaman genotipe dilakukan analisis komponen utama. Pengelompokan genotipe tenggang Al dilakukan berdasarkan nisbah bobot gabah relatif (NBGR) menurut Sarkarung (1986).
Ragam genotipe (Vg2) dan ragam fenotipe (Vp2) dan heritabilitas dalam arti luas (Hbs) diduga berdasarkan sidik ragam untuk percobaan dalam rancangan acak kelompok (Singh et al. 1993). Koefisien korelasi genotipik (rg) dan koefisien korelasi fenotipik (rp) antar sifat diduga berdasarkan sidik kovarian (Singh dan Chaudhary 1979). Untuk memisahkan pengaruh langsung dan tidak langsung karakter yang diamati terhadap bobot gabah per rumpun dilakukan analisis lintas (Li 1975).
HASIL DAN PEMBAHASAN
Penapisan dengan Kultur Hara
Kondisi percobaan ideal digunakan untuk penapisan genotipe tenggang Al karena cekaman Al yang diberikan berpengaruh nyata terhadap semua peubah yang diamati. Adanya pengaruh interaksi antara genotipe dan cekaman Al mengindikasikan penapisan genotipe tenggang Al harus dilakukan pada lingkungan target sesuai dengan tingkat cekaman Al (Tabel 1). Chozin (2006) menyatakan bahwa seleksi pada lingkungan
bercekaman cenderung menghasilkan genotipe yang beradaptasi baik pada lingkungan bercekaman dan didapatkan sumber keragaman untuk pemuliaan lingkungan spesifik. Tabel 1 Kuadrat tengah pengaruh cekaman aluminium terhadap panjang akar, panjang
tajuk, bobot kering akar dan bobot kering tajuk galur haploid ganda Sumber
keragaman
Kuadrat tengah
Panjang akar Panjang tajuk BK akar BK tajuk Aluminium (A) 8.243,438** 19.237,311** 0,535** 40,648**
Genotipe (G) 99,279** 148,395** 0,022** 0,193**
A x G 12,474** 28,370** 0,002tn 0,031**
** berpengaruh sangat nyata, tn tidak nyata, BK = bobot kering
Respon setiap peubah berbeda-beda terhadap cekaman Al. Cekaman Al dapat menurunkan bobot kering tajuk dan panjang akar hampir 50%. Penurunan panjang tajuk dan bobot kering akar hanya mencapai sekitar 20% (Gambar 2). Penurunan bobot kering tajuk disebabkan oleh hara yang diperlukan untuk pertumbuhan tajuk tidak optimum akibat terganggunya penyerapan dan transportasi hara di akar (Polle dan Konzak 1990; Rout et al. 2001; Kochian et al. 2004). Sebaliknya, penurunan bobot kering akar tidak sebesar penurunan bobot kering tajuk karena walaupun panjang akarnya menurun tetapi akar lebih besar dan akar-akar adventif lebih banyak tumbuh pada leher akar. Fageria et al (1988) juga mendapatkan bahwa bobot kering tajuk lebih peka terhadap keracunan Al dibandingkan bobot kering akar.
56.57 68.05 78.54 80.90 0 10 20 30 40 50 60 70 80 90
BK tajuk Panjang akar Panjang tajuk BK akar
Nilai relatif (%)
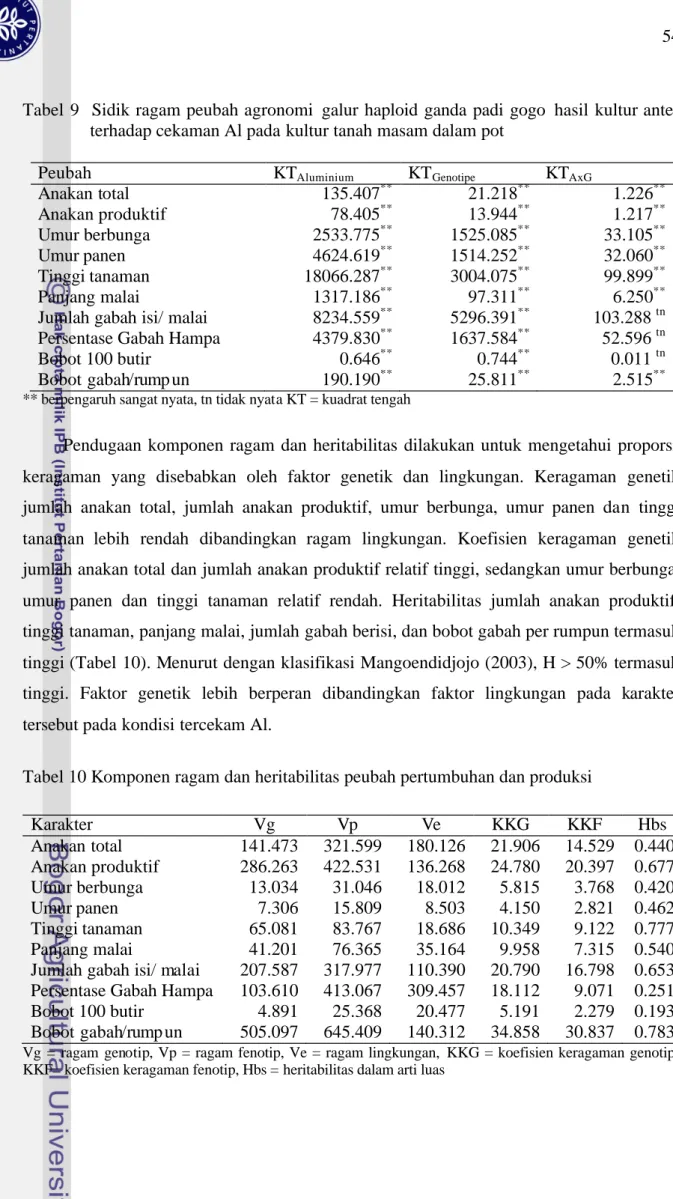
Genotipe yang diuji berbeda nyata pada keempat peubah yang diamati, yang menunjukkan populasi yang diuji sangat beragam. Pendugaan komponen ragam dan heritabilitas dapat dilakukan untuk mengetahui proporsi keragaman yang disebabkan oleh
faktor genetik dan lingkungan. Ragam genetik panjang akar, panjang tajuk, bobot kering akar dan bobot kering tajuk pada kondisi tercekam Al lebih tinggi dibandingkan ragam lingkungan. Ragam genetik nisbah akar tajuk (NAT) lebih rendah dibandingkan ragam lingkungan. Koefisien keragaman genetik panjang tajuk dan NAT lebih rendah dibandingkan karakter lain (Tabel 2).
Tabel 2 Komponen ragam dan heritabilitas dari panjang akar, panjang tajuk, bobot kering akar, bobot kering tajuk dan NAT pada kondisi tercekam Al
Komponen Panjang akar
Panjang tajuk
BK akar BK tajuk NAT
Vg 13.767 20.015 0.003 0.020 0.005 Vp 16.162 29.819 0.004 0.027 0.012 Ve 2.395 9.805 0.001 0.008 0.007 KKG 29.719 13.656 25.801 26.044 19.177 KKF 32.200 16.668 31.595 30.737 28.978 Hbs 0.852 0.671 0.667 0.718 0.438 NAT = nisbah akar/tajuk, Vg = ragam genotip, Vp = ragam fenotip, Ve = ragam lingkungan, KKG = koefisien keragaman genotip, KKF= koefisien keragaman fenotip, Hbs = heritabilitas dalam arti luas
Nilai heritabilitas dalam arti luas panjang akar, panjang tajuk, bobot kering akar, bobot kering tajuk cukup tinggi berkisar 0.67 – 0.85, kecuali NAT. Heritabilitas NAT lebih kecil dibandingkan karakter lain pada kondisi tercekam Al (Tabel 2). Heritabilitas tinggi pada panjang akar, menunjukkan bahwa keragaman panjang akar antar galur disebabkan oleh faktor genetik sehingga seleksi ketenggangan Al dapat dilakukan pada populasi galur haploid ganda padi gogo hasil kultur antera berdasarkan panjang akar.
Analisis korelasi terhadap semua karakter yang diamati menunjukkan adanya korelasi nyata dan searah, kecuali antara bobot kering tajuk dan NAT tidak nyata. Korelasi antara panjang tajuk dan NAT arahnya berlawanan (Tabel 3). Karakter-karakter yang mempunyai korelasi nyata dan searah dapat dipilih salah satu karakter yang mudah diamati sebagai kriteria seleksi karena karakter-karakter lain akan selalu bersama -sama jika diseleksi. Tabel 3 Korelasi panjang akar, panjang tajuk, bobot kering akar, bobot kering tajuk,
nisbah akar/tajuk pada kondisi tercekam Al
Karakter Panjang tajuk BK akar BK tajuk NAT Panjang akar 0.458** 0.430** 0.389** 0.249*
Panjang tajuk 0.624** 0.805** -0.140**
BK akar 0.746** 0.422**
BK tajuk 0.073tn
Panjang akar, bobot kering akar dan bobot kering tajuk dapat dipilih salah satu sebagai kriteria seleksi ketenggangan Al untuk populasi galur haploid ganda padi gogo hasil kultur antera pada kondisi tercekam Al karena memenuhi kriteria yang disarankan oleh Acevedo dan Fereres (1993) dan karakter tersebut memiliki keragaman genetik dan heritabilitas tinggi serta ada korelasi yang erat antar karakter tersebut dengan karakter lain. Diantara keempat karakter tersebut bobot kering tajuk dan panjang akar lebih peka terhadap cekaman Al sehingga dapat membedakan genotipe tenggang dan peka Al. Namun demikian panjang akar lebih mudah dan cepat diamati dibandingkan bobot kering tajuk sehingga dalam penelitian ini digunakan panjang akar relatif sebagai pembeda genotipe tenggang dan peka Al dalam penapisan ketenggangan Al. Beberapa peneliti sebelumnya juga menggunakan panjang akar relatif sebagai kriteria seleksi ketenggangan Al (Khatiwada et al. 1996; Wu et al. 1997; Wu et al. 2000; Rusdiansyah et al, 2001).
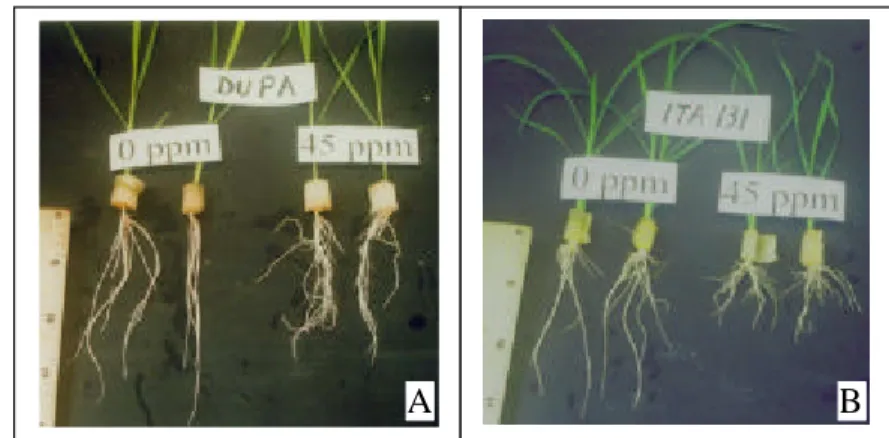
Secara visual dapat diamati dengan mudah perbedaan yang nyata pada pertumbuhan akar akibat cekaman Al pada genotipe peka yaitu adanya hambatan pemanjangan akar, sehingga akar kelihatan lebih pendek dan gemuk, jumlah akarnya lebih banyak yang tumbuh pada leher akar (Gambar 3A) dibandingkan genotipe tenggang (Gambar 3B). Dengan demikian, hambatan panjang akar yang diekspresikan sebagai PAR (panjang akar pada kondisi tercekam Al terhadap panjang akar pada kondisi tidak tercekam Al), dapat digunakan sebagai penciri genotipe peka Al.
Gambar 3 Akar genotipe peka (A) dan tenggang Al (B) pada perlakuan 0 dan 45 ppm Al Hasil pengamatan terhadap panjang tajuk relatif menunjukkan bahwa penurunan panjang tajuk tidak mencapai 50%. Nilai panjang tajuk relatif yang terendah adalah 0,58 pada SGJT31 dan yang tertinggi adalah 1,01 pada SGGM10. Dalam penelitian ini, panjang tajuk tidak digunakan sebagai penciri ketenggangan Al karena tidak mencerminkan tingkat
ketenggangan Al pada populasi galur haploid ganda hasil kultur antera. Koefisien keragaman genetik panjang tajuk juga lebih rendah dibandingkan koefisien keragaman genetik panjang akar (Tabel 2). Rusdiansyah et al (2001) melaporkan bahwa Al kurang berpengaruh terhadap panjang tajuk pada stadia bibit.
Nilai PAR pada varietas Dupa (pembanding tenggang Al) sebesar 0.78 dan PAR varietas ITA131 (pembanding peka) sebesar 0.41 (Tabel 5; Gambar 4). Berdasarkan kriteria yang disarankan Khatiwada et al. (1996) maka Dupa dapat digolongkan sebagai genotipe yang tenggang dan ITA 131 sebagai genotipe peka. Hasil ini konsisten dengan yang dilaporkan sebelumnya oleh Jagau (2000) dan Prasetiyono (2003) bahwa Dupa tergolong tenggang Al. Dengan demikian kedua varietas tersebut dapat digunakan sebagai pembanding dalam penapisan ketenggangan Al untuk menyeleksi populasi galur haploid ganda padi gogo hasil kultur antera.
Gambar 4 Penampilan panjang akar varietas Dupa (A) dan ITA 131 (B) pada keadaan cekaman Al (45 ppm Al) dan tanpa cekaman Al (0 ppm Al).
Pengelompokan tingkat ketenggangan Al pada tanaman padi berdasarkan PAR berbeda-beda antar peneliti. Menurut Khatiwada et al. (1996) pertumbuhan akar pada genotipe peka berkurang lebih dari 50%, sehingga genotipe yang memiliki PAR kurang dari 50% dikelompokkan sebagai peka dan yang memiliki PAR lebih dari 50% sebagai tenggang Al. Kriteria ini lebih sesuai diterapkan untuk mengidentifikasi genotipe tenggang Al dari populasi yang belum diketahui informasi tentang latar belakang genetik karakter ketenggangan Al pada populasi tersebut. Nasution dan Suhartini (1991), mengelompokkan menjadi 3 kelompok berdasarkan nilai PAR yaitu, peka jika PAR = 0.61, moderat antara 0.62 – 0.69 dan tenggang jika = 0.7. Wu et al. (1997) mengelompokkan menjadi tiga kelompok, yaitu peka jika < 0.7, moderat antara 0.7 – 0.9 dan tenggang jika PAR >0.9.
Mengingat perbedaan di atas maka, dalam penelitian ini pengelompokan tingkat ketenggangan tanaman terhadap Al dilakukan berdasarkan penyebaran nilai PAR yang mengikuti pola sebaran normal. Populasi yang dievaluasi sudah diketahui memiliki latar belakang genetik ketenggangan Al yang jelas dari tetua persilanga n yang digunakan untuk menghasilkan tanaman haploid ganda dan populasi yang digunakan merupakan galur homozigot yang tidak bersegregasi lagi pada generasi selanjutnya.
Untuk mendapatkan kurva normal data panjang akat relatif (PAR) ditransformasi menjadi nilai baku Z. Tingkat ketenggangan genotipe dikelompokkan menjadi 3 kelompok yaitu peka jika Z = -1 SD, moderat jika, -1 SD < Z < +1 SD dan tenggang jika Z = +1SD (Aluko dan Oard 2004). Setelah dikembalikan ke data asal maka genotipe dianggap peka jika PAR = 0.54, moderat jika 0.54 < PAR < 0.82 dan toleran, PAR = 0.82 (Gambar 5).
0.54 0.82
PAR
Hasil pengelompokan berdasarkan nilai PAR dengan metode kultur hara yang diberi 45 ppm Al disajikan pada Tabel 3. Dari 113 galur haploid ganda yang diuji diperoleh sebanyak 15 genotipe tenggang, 73 genotipe moderat dan 25 genotipe peka. Sebanyak 11 dari 15 galur haploid ganda tenggang Al merupakan keturunan dari persilangan dengan menggunakan varietas Grogol sebagai salah satu tetua (Tabel 4). Hal ini mengindikasikan bahwa Grogol dapat dipertimbangkan sebagai salah tetua untuk perakitan padi tenggang Al.
Hasil uji BNT pada taraf 5% terhadap galur haploid ganda yang moderat sampai tenggang menunjukkan bahwa ada empat galur haploid ganda yang memiliki nilai PAR nyata lebih tinggi dibandingkan varietas Dupa dan 15 galur nyata lebih rendah dibandingkan varietas Dupa. Galur haploid ganda yang memiliki nilai PAR lebih tinggi dibandingkan varietas Dupa (pembanding tenggang Al) adalah KRGM4, JTGR16, GRGM14 dan GRGM6 keempatnya termasuk tenggang Al. PAR genotipe KRGM4 nyata Gambar 5 Pengelompokan galur haploid ganda padi gogo menurut tingkat ketenggangan
terhadap aluminium berdasarkan PAR. Galur Moderat Galur Tenggang Galur Peka
lebih tinggi dibandingkan PAR kedua tetuanya Krowal atau Gajah Mungkur (Tabel 5). Hal ini diduga akibat adanya segregasi transgresif pada kombinasi antera yang menghasilkan galur tersebut.
Tabel 4 Pengelompokan genotipe berdasarkan PAR pada penapisan dengan metode kultur hara hara Kelompok Genotipe Tenggang KRGM4, JTGR16, GRGM14, GRGM25, GRGM6, GRGM4, GRGM9, SGJT27, JTGR17, JTKR5, JTGR18, JTKR1, GRJT11, JTGR13, JTGR2 dan Grogol Moderat SGGM9, GRJT4, SGJT9, JTGR7, SGGM13, SGJT6, GRGM12, GRJT28, JTGR1, SGJT19, GMGR5, SGJT37, GRGM3, SGJT34, KRGM2, SGGM8, GRJT42, GRJT8, SGJT29, GRJT14, GRJT18, JTGR4, JTKR6, KRGM1, SGGM5, GRJT16, GRJT27, GRGM7, JTKR3, JTGR15, GRJT5, GRJT25, GRJT30, JTGR10, SGJT3, GRJT23, GRGM5, SGJT30, KRGM3, SGJT33, GRGM1, JTGR5, SGJT11, GRJT19, GRJT34, SGJT36, SGJT28, JTGR20, GRJT10, JTKR7, SGJT13, SGJT25, GMGR3, GRJT32, GRGM2, GRJT22, SGJT16, SGJT23, SGJT17, SGJT26, SGJT18, JTGR3, JTGR19, SGGM2, GMGR1, SGGM10, JTKR8, SGJT22, GRJT7, GRJT29, GRJT31, SGJT10, GMGR6, Sigundil, Krowal, Gajah Mungkur dan Dupa
Peka KRJT1, SGJT21, GRJT20, JTKR2, GRJT44, GMGR2, KRJT3, GRJT1, GRJT6, GRGM15, GRJT39, SGJT5, GRJT17, GRJT36, SGJT2, GRJT47, GRJT12, GRGM11, GRJT33, GRGM10, GRJT24, SGJT12, GRJT2, SGJT31, GRJT49, Jatiluhur dan ITA131
Genotipe tenggang diduga memiliki kemampuan untuk mencegah Al agar tidak menyeberangi membran plasma dan masuk ke simplas serta tempat lain yang peka terhadap Al di sitoplasma akar (Kochian 1995; Ma 2000; Rout et al 2001). Dalam keadaan tercekam Al, kandungan Al apoplas padi genotipe tenggang lebih rendah akibat kapasitas tukar kation (KTK) akar rendah dibandingkan genotipe peka. Ca apoplas pada genotipe tenggang lebih rendah dan tidak mudah digantikan oleh Al (Okada et al. 2003). Rendahnya jumlah muatan negatif dari dinding sel genotipe tenggang menyebabkan interaksi Al dengan dinding sel genotipe tenggang lebih rendah dibandingkan genotipe peka (Watanabe dan Okada 2005a; Kochian et al. 2005) sehingga konsentrasi Al di akar genotipe tenggang lebih rendah dibandingkan genotipe peka. Fenomena ini juga telah dilaporkan beberapa peneliti sebelumnya (Ganesan et al. 1993; Sivaguru dan Paliwal 1993; Nursyamsi 2000) bahwa tanaman padi tenggang mempunyai mekanisme ketenggangan Al dengan mengurangi interaksi Al dengan dinding sel akar.
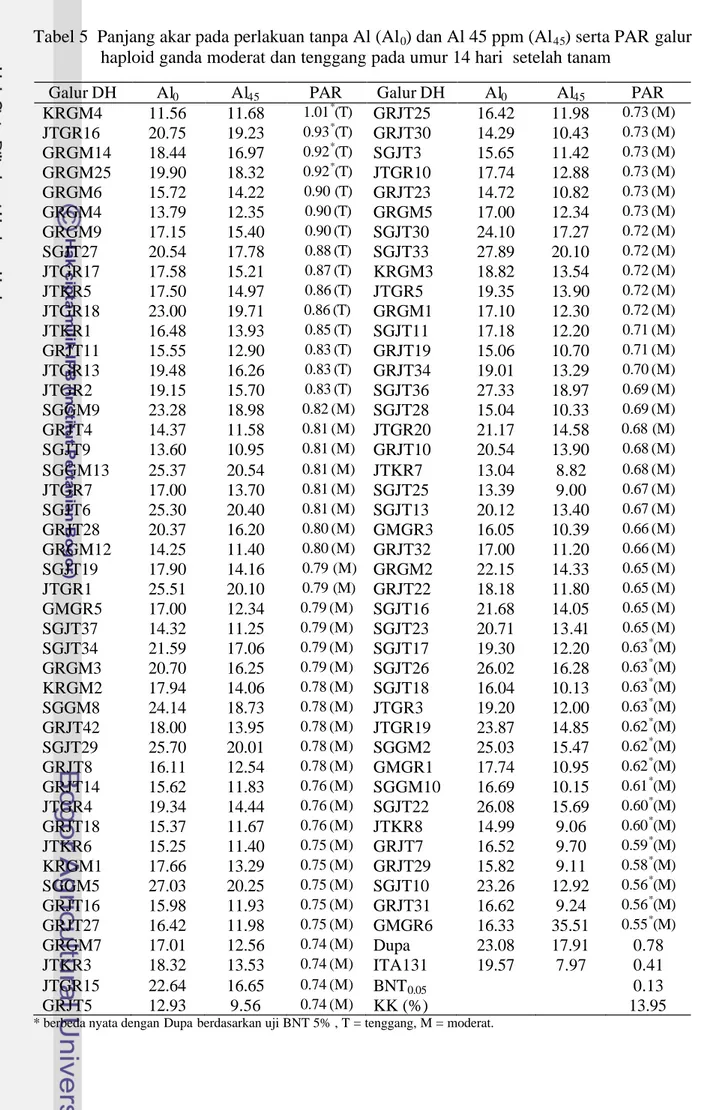
Tabel 5 Panjang akar pada perlakuan tanpa Al (Al0) dan Al 45 ppm (Al45) serta PAR galur haploid ganda moderat dan tenggang pada umur 14 hari setelah tanam
Galur DH Al0 Al45 PAR Galur DH Al0 Al45 PAR
KRGM4 11.56 11.68 1.01*(T) GRJT25 16.42 11.98 0.73(M) JTGR16 20.75 19.23 0.93*(T) GRJT30 14.29 10.43 0.73(M) GRGM14 18.44 16.97 0.92*(T) SGJT3 15.65 11.42 0.73(M) GRGM25 19.90 18.32 0.92*(T) JTGR10 17.74 12.88 0.73(M) GRGM6 15.72 14.22 0.90 (T) GRJT23 14.72 10.82 0.73(M) GRGM4 13.79 12.35 0.90(T) GRGM5 17.00 12.34 0.73(M) GRGM9 17.15 15.40 0.90(T) SGJT30 24.10 17.27 0.72(M) SGJT27 20.54 17.78 0.88(T) SGJT33 27.89 20.10 0.72(M) JTGR17 17.58 15.21 0.87(T) KRGM3 18.82 13.54 0.72(M) JTKR5 17.50 14.97 0.86(T) JTGR5 19.35 13.90 0.72(M) JTGR18 23.00 19.71 0.86(T) GRGM1 17.10 12.30 0.72(M) JTKR1 16.48 13.93 0.85(T) SGJT11 17.18 12.20 0.71(M) GRJT11 15.55 12.90 0.83(T) GRJT19 15.06 10.70 0.71(M) JTGR13 19.48 16.26 0.83(T) GRJT34 19.01 13.29 0.70(M) JTGR2 19.15 15.70 0.83(T) SGJT36 27.33 18.97 0.69(M) SGGM9 23.28 18.98 0.82(M) SGJT28 15.04 10.33 0.69(M) GRJT4 14.37 11.58 0.81(M) JTGR20 21.17 14.58 0.68 (M) SGJT9 13.60 10.95 0.81(M) GRJT10 20.54 13.90 0.68(M) SGGM13 25.37 20.54 0.81(M) JTKR7 13.04 8.82 0.68(M) JTGR7 17.00 13.70 0.81(M) SGJT25 13.39 9.00 0.67(M) SGJT6 25.30 20.40 0.81(M) SGJT13 20.12 13.40 0.67(M) GRJT28 20.37 16.20 0.80(M) GMGR3 16.05 10.39 0.66(M) GRGM12 14.25 11.40 0.80(M) GRJT32 17.00 11.20 0.66(M) SGJT19 17.90 14.16 0.79 (M) GRGM2 22.15 14.33 0.65(M) JTGR1 25.51 20.10 0.79 (M) GRJT22 18.18 11.80 0.65(M) GMGR5 17.00 12.34 0.79(M) SGJT16 21.68 14.05 0.65(M) SGJT37 14.32 11.25 0.79(M) SGJT23 20.71 13.41 0.65(M) SGJT34 21.59 17.06 0.79(M) SGJT17 19.30 12.20 0.63*(M) GRGM3 20.70 16.25 0.79(M) SGJT26 26.02 16.28 0.63*(M) KRGM2 17.94 14.06 0.78(M) SGJT18 16.04 10.13 0.63*(M) SGGM8 24.14 18.73 0.78(M) JTGR3 19.20 12.00 0.63*(M) GRJT42 18.00 13.95 0.78(M) JTGR19 23.87 14.85 0.62*(M) SGJT29 25.70 20.01 0.78(M) SGGM2 25.03 15.47 0.62*(M) GRJT8 16.11 12.54 0.78(M) GMGR1 17.74 10.95 0.62*(M) GRJT14 15.62 11.83 0.76(M) SGGM10 16.69 10.15 0.61*(M) JTGR4 19.34 14.44 0.76(M) SGJT22 26.08 15.69 0.60*(M) GRJT18 15.37 11.67 0.76(M) JTKR8 14.99 9.06 0.60*(M) JTKR6 15.25 11.40 0.75(M) GRJT7 16.52 9.70 0.59*(M) KRGM1 17.66 13.29 0.75(M) GRJT29 15.82 9.11 0.58*(M) SGGM5 27.03 20.25 0.75(M) SGJT10 23.26 12.92 0.56*(M) GRJT16 15.98 11.93 0.75(M) GRJT31 16.62 9.24 0.56*(M) GRJT27 16.42 11.98 0.75(M) GMGR6 16.33 35.51 0.55*(M) GRGM7 17.01 12.56 0.74(M) Dupa 23.08 17.91 0.78 JTKR3 18.32 13.53 0.74(M) ITA131 19.57 7.97 0.41 JTGR15 22.64 16.65 0.74(M) BNT0.05 0.13 GRJT5 12.93 9.56 0.74(M) KK (%) 13.95
Metode seleksi cepat pada stadia bibit dengan menggunakan media kultur hara sangat efisien pada tahapan awal program pemuliaan tanaman untuk menapis genotipe tenggang Al dari populasi bahan seleksi dalam jumlah banyak sebelum diuji di lapangan. Farid et al (1997) mendapatkan korelasi antara nilai PAR pada kultur hara dan hasil biji pada kondisi tanah masam rendah (r = 0.28), sehingga galur tenggang yang diperoleh pada metode kultur hara perlu dievaluasi kembali dengan metode lain, baik pada kondisi lingkungan buatan maupun alami. Hal ini disebabkan ketenggangan Al pada tanaman padi dikendalikan oleh banyak gen (Ma et al. 2002; Nguyen et al. 2001; Wu et al. 2000) sehingga tidak semua genotipe akan memiliki semua gen pengendali ketenggangan Al. Setiap gen atau kombinasinya akan memiliki peran dalam meregulasi mekanisme ketenggangan Al pada tanaman padi, sehingga setiap genotipe akan memiliki mekanisme ketenggangan Al yang berbeda. Ada gen yang terekspresi pada stadia bibit dan ada gen yang terekspresi pada tahap perkembangan tanaman yang lebih dewasa. Hal ini telah dilaporkan Wu et al (2000) bahwa ekspresi gen tenggang Al pada padi tergantung perkembangan tanaman.
Penapisan dengan Media Tanah
Hasil analisis tanah yang digunakan sebagai media tanam menunjukkan bahwa tanah tersebut tergolong sangat masam dengan pH 3.58. Kejenuhan Al sangat tinggi mencapai 79%, KTK tinggi dan kejenuhan basa sangat rendah (Tabel Lampiran 2). Tanah tersebut ideal sebagai lingkungan seleksi untuk penapisan genotipe tenggang Al.
Analisis Komponen Utama
Karakter anakan total, anakan produktif, tinggi tanaman, umur berbunga, umur panen, panjang malai, jumlah gabah isi per malai, persentase gabah hampa, bobot 100 butir dan bobot gabah per rumpun digunakan sebagai peubah untuk mengidentifikasi galur haploid ganda yang memiliki potensi untuk diseleksi sebagai galur tenggang Al melalui analisis peubah ganda.
Untuk menentukan karakter yang memberi pengaruh besar terhadap keragaman genotipe yang diuji maka hubungan antar karakter ditransformasi menjadi peubah baru yang tidak berkorelasi yang dinamakan dengan komponen utama. Peubah baru tersebut merupakan kombinasi linear dari karakter asal dan menurun dengan berdasarkan urutan kepentingan sehingga komponen utama pertama merupakan penyusun keragaman utama
populasi. Dengan membandingkan nilai vektor ciri dari karakter yang disajikan pada Tabel 6 dapat ditentukan karakter asal yang memberikan sumbangan besar terhadap keragaman genotipe yang diuji. Everitt dan Dunn (1991) menyarankan penggunaan batas nilai eigen ≥ 0.7 untuk mendapatkan n komponen utama yang memberikan gambaran yang layak untuk pengelompokan.
Analisis komponen utama terhadap 88 galur haploid ganda berdasarkan sepuluh peubah diperoleh 74.4% keragaman populasi galur haploid ganda yang diuji disumbangkan oleh tiga komponen utama. Bobot gabah per rumpun, jumlah anakan produktif dan jumlah biji berisi per malai dapat dijadikan sebagai penciri untuk seleksi ketenggangan Al pada tanah masam. Karakter tersebut mendominasi komponen utama pertama dan dapat menjelaskan 45.6% keragaman populasi. Bobot gabah per rumpun dan jumlah anakan produktif dapat membedakan tingkat ketenggangan Al pada populasi galur haploid ganda yang dicobakan karena memiliki nilai akar ciri tertinggi pada komponen utama pertama dibandingkan karakter lain, sehingga pada percobaan ini digunakan bobot gabah per rumpun untuk mengitung indek ketenggangan Al (Tabel 6).
Tabel 6 Hasil analisis komponen utama beberapa karakter galur haploid ganda padi gogo hasil kultur antera
Karakter Komponen Utama
1(45.6%) 2 (18.2%) 3 (10.6%) Bobot gabah per rumpun 0.211 -0.21 0.094 Jumlah anakan produktif 0.204 -0.038 -0.025
Jumlah biji berisi 0.193 0.010 0.165
Jumlah anakan total 0.173 -0.042 -0.030
Tinggi tanaman 0.153 0.044 -0.202
Panjang malai 0.141 0.074 -0.330
Persentase gabah hampa -0.150 0.034 -0.163
Umur panen -0.001 0.517 0.091
Umur berbunga 0.014 0.510 0.141
Bobot 100 butir 0.011 -0.098 0.838
Korelasi antar Karakter
Korelasi antar karakter yang diamati diduga berdasarkan koefisien korelasi karena koefisien tersebut merupakan ukuran keeratan hubungan antar karakter yang dianalisis. Korelasi antar karakter, penting dalam pemuliaan tanaman terutama untuk mengetahui respon berkorelasi dalam seleksi tidak langsung. Apabila satu karakter dan karakter lain mempunyai respon berkorelasi maka jika kita ingin memperbaiki karakter yang susah
diamati kita dapat menyeleksi karakter lain yang mudah diamati. Kemajuan seleksi karakter yang sulit diamati akan diperoleh melalui respon berkorelasi dari karakter yang dijadikan sebagai kriteria seleksi. Hal ini dinamakan dengan seleksi tidak langsung, yaitu seleksi dilakukan melalui karakter lain untuk satu karakter yang ingin diperbaiki (Falconer dan Mackay 1996).
Korelasi antar karakter perlu diketahui agar seleksi lebih efektif dilakukan dengan menggunakan satu karakter sebagai kriteria seleksi tidak langsung untuk karakter lain yang berkorelasi. Pada penelitian ini, jumlah anakan total berkorelasi positif dan sangat nyata dengan jumlah anakan produktif, tinggi tanaman, panjang malai dan bobot gabah per rumpun. Tinggi tanaman berkorelasi positif dan sangat nyata dengan panjang malai, jumlah gabah berisi per malai dan bobot gabah per rumpun. Panjang malai berkorelasi dengan jumlah gabah berisi per malai dan bobot gabah per rumpun (Tabel 7).
Persentase gabah hampa berkorelasi negatif dan nyata dengan jumlah anakan total, jumlah anakan produktif, tinggi tanaman, panjang malai, jumlah gabah berisi per malai dan bobot gabah per rumpun (Tabel 7). Hal ini mengindikasikan bahwa penurunan persentase gabah hampa akan meningkatkan bobot gabah per rumpun galur haploid ganda padi gogo. Umur berbunga, umur panen dan bobot 100 butir berkorelasi tidak nyata dengan karakter-karakter lain. Hal ini berbeda dengan yang dilaporkan Sutaryo et al (2005) bahwa bobot 1000 butir berkorelasi positif dan sangat nyata dengan hasil gabah. Perbedaan ini terjadi karena ukuran biji dari galur haploid ganda hasil kultur antera yang digunakan dalam penelitian ini hampir sama.
Korelasi yang tinggi hanya menunjukkan keeratan hubungan antar sifat, tetapi tidak dapat menunjukkan adanya hubungan sebab akibat. Sidik lintas dapat dimanfaatkan untuk mengetahui hubungan sebab akibat dan dapat memilahnya menjadi pengaruh langsung dan tidak langsung melalui karakter lain (Li 1975). Berdasarkan sidik lintas dapat dilihat bahwa jumlah anakan produktif dan jumlah gabah berisi per malai memberikan pengaruh langsung paling besar terhadap bobot gabah per rumpun diantara karakter lain. Jumlah anakan total, tinggi tanaman, panjang malai dan persentase gabah hampa mempunyai pengaruh langsung kecil terhadap bobot gabah per rumpun. Korelasi yang tinggi antara jumlah anakan total, tinggi tanaman, panjang malai dan persentase gabah hampa dengan bobot gabah per rumpun merupakan akibat dari pengaruh tidak langsung melalui jumlah anakan produktif dan jumlah gabah berisi per malai (Tabel 8).
BGPR 0.759** 0.959* 0.062 tn -0.040 tn 0.544** 0.497** 0.904** -0.616** 0.084 tn B obot 100 B utir 0.034 tn -0.024 tn -0.050 tn -0.088 tn 0.015 tn -0.089 tn 0.114 tn -0.068 tn % G abah H ampa -0.340** -0.571** 0.025 tn 0.013 tn -0.369** -0.332** -0.653** Jumlah Gabah Isi 0.558** 0.774** 0.087 tn 0.025 tn 0.473** 0.487** P anjang Malai 0.368** 0.475** 0.077 tn 0.071 tn 0.599** T inggi T anaman 0.512** 0.538** 0.054 tn 0.051 tn U mur P anen -0.062 tn -0.083 tn 0.786** U mur B erbunga 0.009 tn 0.031 tn A nakan P roduktif 0.806** Karakter
Anakan total Anakan
P roduktif U mur B erbunga U mur Panen T inggi T anaman
Jumlah Gabah Isi %
G abah H ampa B obot 100 B utir BGPR Tabel 7 Nilai koefisien k orelasi
antar karakter pada galur haploid ganda padi gogo dalam kondisi tercekam Al
BGPR = bobot gabah per rumpun, tn = tidak nyata,