2. TINJAUAN PUSTAKA
2.1 Temilok
Temilok merupakan kerang pengebor kayu dari genus Bactronophorus
yang memanfaatkan fragmen kayu sebagai sumber makanan pokoknya (Allan 1962). Sebagian besar jenis kerang ini hidup di dalam kayu mati (Olsson 1961).
Identifikasi kerang famili Teredinidae berdasarkan bentuk cangkang ekor yang disebut pallet. Pallet adalah sepasang tulang pipih yang tersusun dari kapur dan khitin dan senantiasa melindungi bagian siphon, serta digunakan untuk menutup terowongan kayu yang dibuatnya (Olsson 1961).
Klasifikasi temilok menurut Allan (1962) adalah sebagai berikut: phylum : Molluska
class : Bivalvia
ordo : Myoida
family : Teredinidae genus : Bactronophorus
species : Bactronophorus thoracites Gould.
Anggota famili teredinidae yang termasuk genus Bactronophorus apabila memiliki pallet berbentuk engrang atau jangkungan (stilt), siphon yang pendek, panjang cangkang kepala kurang lebih 1,85 cm, dan panjang total pallet dapat mencapai kurang lebih 5 cm (Allan 1962). Bentuk fisik temilok dapat dilihat pada Gambar 2.
Gambar 2 Bentuk fisik Bactronophorus thoracites (Leiwakabessy 2011). Bagian mantel kerang ini mensekresikan sejenis kapur yang berguna untuk melapisi dinding bagian dalam terowongan kayu buatannya (Olsson 1961).
Lapisan kapur tersebut tetap menempel pada dinding kayu meskipun kerang ini ditarik keluar. Kerang ini dapat berputar dan melakukan kontraksi di dalam terowongan buatannya serta dapat menutup mulut terowongan buatannya menggunakan kedua palletnya dengan terlebih dulu menarik kedua siphonnya ke dalam sehingga dapat bertahan hidup di dalam dinding kapal kayu selama berminggu-minggu di dalam air tawar (Kaestner 1967). Terowongan kayu akan bertambah panjang seiring dengan pertumbuhan kerang yang hidup di dalamnya. Kerang ini mampu menggali terowongan sepanjang 18 cm hingga 2 m (Morton 1978). Usia harapan hidupnya antara satu hingga beberapa tahun bergantung pada jenisnya (Waterbury et al. 1983).
Kerang-kerangan dari famili teredinidae merupakan kerang spesialis pelobang kayu. Palvast dan Velde (2011) melaporkan bahwa dalam tahun pertama kehidupannya di dermaga yang rentang salinitas perairannya cukup lebar, kerang pengebor kayu (teredinidae) mengkonsumsi 12,4% panel kayu. Rata-rata
pertambahan panjang per hari kerang ini dapat mencapai 1,5 mm. Cragg et al. (2009) melaporkan bahwa kerang ini menjadikan kayu sebagai
makanan sekaligus terowongan tempat tinggal dengan mengebor. Apabila perairan hanya menyediakan sedikit plankton maka siphon kerang ini akan membuka lebih lebar. Larva kerang ini juga memperoleh makanan lewat mekanisme matrotrophy dimana induk menyediakan partikel kayu yang sudah dihancurkan sebagai bahan makanan bagi larva.
Kerang-kerangan umumnya menyimpan hasil pencernaannya dalam bentuk glikogen dan lemak. Kerang-kerangan famili teredinidae mencerna kayu di dalam lambung dengan suatu kelenjar pencernaan. Enzim pencernaannya atau enzim dari simbion bakteri dapat menghancurkan 80% selulosa dan 15-56% hemiselulosa menjadi gula, serta hanya sekitar 21% selulosa dan 55% lignin yang dapat diperoleh kembali dari kotorannya. Asam-asam amino hampir tidak ditemukan di dalam kayu, oleh karena itu keluarga kerang ini memanfaatkan plankton sebagai sumber protein (Purchon 1968). Syaputra (2007) melaporkan bahwa temilok hidup dalam kayu di hutan bakau di sebelah utara Kabupaten Bangka yang ditumbuhi oleh Rhizophora apiculata.
Kerang-kerangan anggota famili teredinidae adalah contoh yang paling umum dari hewan hermaprodit protandri. Kondisi saat perkembangan testis dipercepat dan perkembangan ovarium melambat atau menjadi terbelakang merupakan ciri hewan hermaprodit yang bersifat protandri. Fase muda dari organisme menjalankan fungsi sebagai organisme jantan, dan ketika menuju tahap akhir kedewasaan menjadi betina. Pada kondisi seperti ini, semua individu yang selamat hingga dewasa pertama kali harus melalui fase sebagai individu jantan dan kemudian harus memasuki fase sebagai individu betina (Purchon 1968). 2.2 Glikogen
Sintesis glikogen pada hewan sebenarnya terjadi pada semua jaringan, terutama pada otot hati dan kerangka. Tahap awal sintesis glikogen dari glukosa bebas adalah reaksi heksokinase, yang melakukan fosforilasi glukosa menjadi glukosa 6-fosfat. Reaksi kunci di dalam biosintesis glikogen yaitu pembentukan uridin difosfat glukosa (UDP-glukosa) oleh kerja glukosa 1-fosfat uridiltransferase. Reaksi ini dialihkan menuju ke kanan oleh kerja pirofosfatase, yang menghidrolisis pirofosfat inorganik (PPi) menjadi ortofosfat inorganik (Pi). Uridin difosfat glukosa adalah senyawa antara di dalam pengubahan D-galaktosa menjadi D-glukosa. Uridin difosfat glukosa merupakan donor langsung residu glukosa di dalam pembentukan glikogen enzimatik oleh kerja glikogen sintetase, yang menggiatkan pemindahan residu glukosil dari UDP-glukosa ke ujung nonreduksi molekul glikogen bercabang. Pada reaksi ini, pautan baru α-1,4 dibentuk di antara atom karbon 1 molekul glukosa yang datang dan atom karbon 4 residu glukosa ujung pada cabang glikogen. Keseimbangan keseluruhan rangkaian reaksi ini sangat mendorong sintesis glikogen. Glikogen sintetase memerlukan suatu rantai atau cabang α-1,4 poliglukosa primer dengan sedikitnya 4 residu glukosa yang menambahkan gugus glukosil secara berurutan pada ujung nonreduksi molekul primer (Lehninger 1982).
2.2.1 Glikogen alami
Karbohidrat di dalam tubuh hewan dan manusia yang merupakan sumber energi adalah glikogen (Pedjiadi dan Supriyanti 2006). Glikogen merupakan sumber polisakarida utama pada sel hewan, sedangkan pati adalah sumber
polisakarida pada sel tanaman. Glikogen merupakan polisakarida bercabang dari D-glukosa dalam ikatan α-1,4 seperti amilopektin, tetapi pada glikogen terdapat lebih banyak percabangan dan strukturnya lebih kompak. Ikatan pada percabangannya adalah α-1,6. Glikogen banyak terdapat di dalam hati (mencapai 7% berat basah), di samping itu juga terdapat pada otot kerangka. Glikogen ditemukan sebagai granula besar-besar di dalam sel hati, yang merupakan kumpulan dari granula yang lebih kecil, bercabang rapat dengan berat molekul rata-rata beberapa juta. Granula glikogen tersebut juga mengandung enzim-enzim
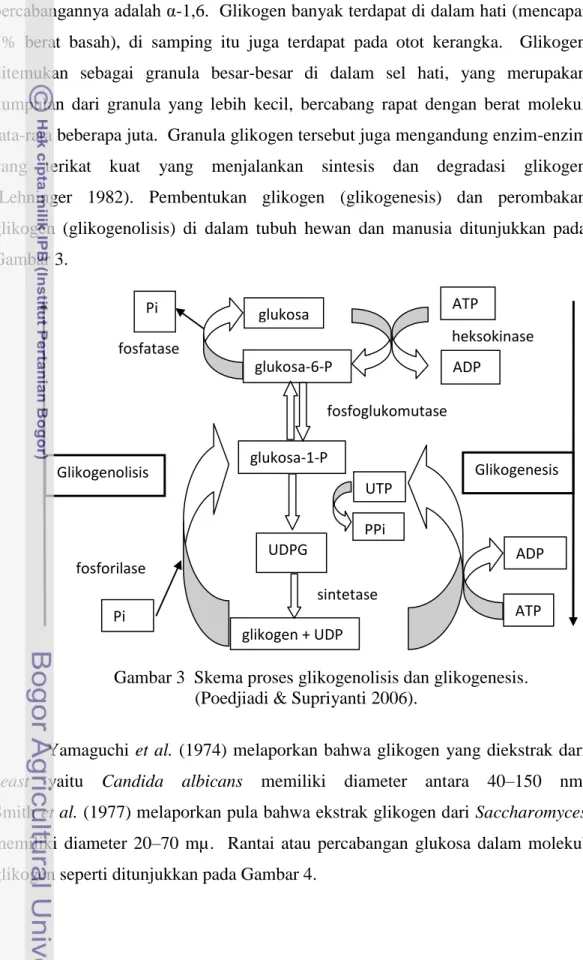
yang terikat kuat yang menjalankan sintesis dan degradasi glikogen (Lehninger 1982). Pembentukan glikogen (glikogenesis) dan perombakan
glikogen (glikogenolisis) di dalam tubuh hewan dan manusia ditunjukkan pada Gambar 3.
Gambar 3 Skema proses glikogenolisis dan glikogenesis. (Poedjiadi & Supriyanti 2006).
Yamaguchi et al. (1974) melaporkan bahwa glikogen yang diekstrak dari
yeast yaitu Candida albicans memiliki diameter antara 40–150 nm.
Smith et al. (1977) melaporkan pula bahwa ekstrak glikogen dari Saccharomyces memiliki diameter 20–70 mµ. Rantai atau percabangan glukosa dalam molekul glikogen seperti ditunjukkan pada Gambar 4.
glukosa glukosa-6-P heksokinase ADP ATP glukosa-1-P UDPG fosfoglukomutase UTP PPi glikogen + UDP fosforilase fosfatase ADP ATP Glikogenolisis sintetase Pi Pi Glikogenesis
(a)
Gambar 4 (a) Model molekuler glikogen tiram (Matsui et al. 1996).
(b) Monomer glukosa dan struktur dasar glikogen (McCormick 2006). Nicoletti dan Baiocchi (1994) melaporkan bahwa kandungan glikogen di dalam jaringan tubuh yang berbeda pada individu yang sama juga berbeda, bahkan di dalam jaringan tubuh yang sama pada spesies yang sama karena dipengaruhi oleh beberapa faktor seperti status gizi dan musim. Beberapa glikogen yang berasal dari jenis-jenis hewan berbeda, sejauh ini menunjukkan perbedaan pada derajat percabangan polisakaridanya. Percabangan polisakarida pada glikogen yang diekstrak dari mammalia berbeda nyata dengan invertebrata.
Alonso et al. (1995) melaporkan bahwa jumlah glikogenin sebagai protein yang mengawali sintesis glikogen mempengaruhi banyaknya glikogen yang dapat disimpan oleh suatu sel, atau sebagai pembatas tingkat pembentukan glikogen dan merupakan bagian di dalam molekul glikogen. Meezan et al. (1995) melaporkan bahwa glikogenin merupakan proteoglikan, suatu ikatan dengan rantai tepi asam amino serina, bertindak sebagai primer bagi pembentukan polisakarida dimana gula ditambahkan oleh kinerja enzim glikosil transferase. Proteoglikan yang mampu melakukan glukosilasi secara mandiri dan mengkatalis permulaan biosintesis glikogen. Proteoglikan ini juga tahan terhadap perlakuan alkali.
2.2.2 Ekstraksi glikogen
Ekstraksi adalah proses penarikan komponen yang diinginkan dari suatu bahan dengan memisahkan satu atau lebih komponen dari bahan sumber komponennya (Khopkar 2003). Cara ekstraksi yang paling sering digunakan adalah maserasi. Prinsip dasar ekstraksi dengan maserasi adalah menarik
komponen terlarut pada bahan padat menggunakan suatu pelarut yang disebut
solid-liquid extraction (Handa 2008).
Bueding dan Orrell (1964) telah membandingkan hasil ekstraksi glikogen dengan alkali panas dan air dingin. Bobot molekul glikogen Ascaris lumbricoides hasil isolasi air dingin 50 kali lebih tinggi daripada dengan alkali panas, residu protein ikutan pada ekstraksi alkali panas 5-10 kali lebih tinggi daripada ekstraksi air dingin. Sentrifugasi dengan kecepatan tinggi dan waktu yang lama dapat meningkatkan hasil atau perolehan glikogen karena komponen-komponen glikogen berbobot molekul rendah terambil atau terpisah dari supernatannya, sedangkan sentrifugasi yang lambat dan lama akan menyebabkan kehilangan komponen berbobot molekul lebih tinggi. Kerly (1930) melaporkan bahwa ekstraksi glikogen dengan metode alkali panas dapat menurunkan kadar pengotor glikogen yaitu residu nitrogen, namun menurut laporan Orrel dan Buedding (1964) dan Yamaguchi et al. (1974) juga dapat mendegradasi glikogen sehingga dapat menurunkan rendemen ekstrak.
Kerly (1930) melaporkan bahwa ekstrak glikogen remis yang diperoleh dari perlakuan tanpa perebusan dalam larutan alkali dengan kadar nitrogen 0,5% dapat melarut sempurna di dalam air pada suhu 20 oC setelah 3-4 hari hingga konsentrasi glikogen sekitar 16%. Sahyun dan Alsberg (1930) melaporkan bahwa glikogen memiliki sifat adesif yang sangat kuat bahkan terhadap permukaan yang licin seperti kaca, serta memiliki warna sedikit coklat (opalescence) dalam bentuk larutan.
Pengukuran residu nitrogen di dalam glikogen terekstrak dapat dilakukan dengan metode Kjeldahl. Nicoletti dan Baiocchi (1994) melaporkan karakteristik ekstrak glikogen yang dihasilkan dari Mytilus gallus Provincialis memiliki residu nitrogen 0% diukur dengan metode Kjeldahl (dengan sensitivitas hingga 60 ppm) dan kandungan atom karbon sebesar 44.44 %.
Vies (1954) melaporkan bahwa kemurnian glikogen ditentukan setelah dikonversi menjadi glukosa dengan kadar sekitar 90-95%. Bennet et al. (2007) melaporkan bahwa fenol-sulfat sangat efektif dalam menghidrolisis glikogen menjadi glukosa tanpa dipengaruhi oleh ukuran molekul, percabangan, dan
sumber bahan baku glikogen sehingga fenol-sulfat digunakan untuk pengujian kadar glukosa di dalam glikogen secara kuantitatif.
2.2.3 Sifat resin kationik (Davis 2010).
Pertukaran ion merupakan reaksi yang dapat berbalik (reversible reaction) dimana ion bermuatan dari suatu larutan ditukar oleh ion lain yang sama muatannya yang menempel pada suatu partikel padat yang tidak bergerak (immobile solid particle).
1) Kapasitas pertukaran
Kapasitas pertukaran adalah banyaknya ion-ion pembalas (counter ions) yang dapat ditukar pada resin, dinyatakan sebagai miliequivalen per gram (meq/g) resin kering (3,6-5,5 meq/g) atau resin basah (1,8-2,0 meq CaCO3/mL).
2) Selektivitas
Resin penukar ion memiliki afinitas atau variabel kecenderungan terhadap ion-ion tertentu di dalam larutan. Kecenderungan ini disebut juga selektivitas yang dinyatakan secara kuantitatif dengan koefisien selektivitas bagi kation atau konstanta keseimbangan nyata. Jika nilai koefisien selektivitas (K) semakin besar maka semakin besar pula kecenderungan ion tersebut dengan resin. Faktor keterpisahan (separation factor) atau αi lebih sering digunakan daripada koefisien selektivitas di dalam evaluasi rancangan proses, seperti ditunjukkan pada Tabel 1. Matriks resin adalah polistirena divinilbenzena (DVB) dengan gugus fungsional yaitu sulfonat diaplikasikan di dalam 0,005-0,010 N larutan CaCO3 dengan nilai total dissolved solid (TDS) sebesar 250-500 mg/L.
3) Ukuran partikel
Ukuran partikel memiliki 2 pengaruh dalam proses tukar ion. Tingkat atau laju tukar ion akan menurun dengan meningkatnya ukuran partikel. Kebalikannya, penurunan ukuran resin meningkatkan hilangnya resin di dasar wadah karena tekanan yang merusak struktur bangun (fisik) resin, sehingga besarnya tekanan hidrolik yang dikenakan di dalam proses yang melibatkan resin sangat menentukan pilihan ukuran partikel resin. Diameter resin yang umumnya berbentuk bulat berkisar antara 0,04 hingga 1,00 mm.
Tabel 1 Nilai faktor keterpisahan (separation factor) [αi ] beberapa kation
Resin kation asam kuat Kation, i αi/Na+ Ra2+ 13,0 Ba2+ 5,8 Pb2+ 5,0 Sr2+ 4,8 Cu2+ 2,6 Ca2+ 1,9 Zn2+ 1,8 Fe2+ 1,7 Mg2+ 1,67 K+ 1,67 Mn2+ 1,6 NH4+ 1,3 Na+ 1,0 H+ 0,67
4) Stabilitas struktural dan umur kerja
Aliran larutan bertekanan tinggi dapat menyebabkan himpitan bahkan kerusakan pada bangun resin sehingga menurunkan umur kerjanya. Penurunan integritas struktural dan unjuk kerja resin dapat pula disebabkan oleh pembengkakan, pecah karena tertusuk, atau abrasi karena pencucian yang berlebihan. Klorinasi terhadap resin DVB tersulfonasi asam kuat menyebabkan resin teroksidasi yang dapat menurunkan umur kerjanya. Konsentrasi besi dan mangan yang berlebihan di dalam air, jika teroksidasi, akan membentuk presipitat yang dapat membuat resin saling menempel dan melekat satu sama lain sehingga kadar besi, mangan, atau kombinasi keduanya tidak boleh lebih dari 0,3 mg/L air. Turbiditas air yang digunakan bagi resin kationik sebaiknya tidak lebih dari 5 NTU (nephelometric turbidity unit). Turbiditas menyatakan banyaknya partikel-partikel bahan tersuspensi seperti tanah, plankton, dan bahan organik. Turbiditas lebih dari 5 NTU dapat dengan mudah diamati dalam segelas air yang biasanya digunakan sebagai alasan estetika.
Resin penukar kation asam kuat bukan mengarah kepada kekuatan fisiknya namun lebih kepada pengertian yang berlandaskan kepada teori Arrhenius tentang kekuatan elektrolit, yaitu gugus fungsional dari resin terdisosiasi secara sempurna dalam bentuk ionnya pada semua kondisi pH. Gugus
fungsional pada resin penukar ion umumnya terdiri dari 4 kategori yaitu strongly
acidic (contohnya –SO3- atau sulfonat), weakly acidic (contohnya –COO- atau
karboksilat), strongly basic (contohnya –N+(CH3)3), dan weakly basic (contohnya
–N(CH3)2). Karakteristik resin kationik Amberlite IR-120 Na disajikan pada
Tabel 2.
Tabel 2 Karakteristik resin kationik Amberlite IR-120 Na
Parameter Keterangan
Bentuk fisik Butiran berbentuk bulat kecil
Matriks Polistirena divinilbenzena
Gugus fungsional Sulfonat
Bentuk ion Na+
Kapasitas pertukaran ion total >2,00 eq/L Kapasitas penahan kelembaban 45-50%
Massa jenis 840 g/L
Diameter partikel 0,6-0,8 mm
Suhu operasi maksimum 135 oC
Kisaran pH operasi 0-14
Sumber : Product Data Sheet www.amberlite.com
2.3 Konsep Spektrophotometri Asam Nukleat
Sifat dan perilaku basa purin dan pirimidin dari suatu asam nukleat yang terdenaturasi digunakan sebagai dasar dalam pelacakan asam nukleat. Protein juga menyerap sinar ultraviolet dengan panjang gelombang 260 nm meskipun pada tingkat yang rendah tergantung pada kandungan asam amino aromatik fenilalanina (Phe), triptofan (Trp) dan tirosina (Tyr). Trp dan Tyr mampu menyerap sinar 280 nm secara maksimum (Adams et al. 1986).
2.3.1 Konsep pengukuran RNA-DNA
Perlakuan suhu atau pH ekstrim terhadap suatu molekul DNA beruntai ganda akan memutus ikatan-ikatan hidrogen di dalam struktur double helix-nya sehingga menjadi suatu kumparan-kumparan yang acak. Suhu ekstrim yang menyebabkan pemutusan ikatan-ikatan hidrogen di dalam molekul DNA disebut suhu transisi atau melting temperature. Hyperchromic effect adalah keadaan dimana komponen basa dari polinukleotida hasil denaturasi asam deoksiribonukleat menyerap sinar secara maksimum pada panjang gelombang 260 nm. Denaturasi asam deoksiribonukleat umumnya mulai terjadi saat pemanasan mencapai suhu 95 oC, yang umumnya memiliki nilai absorbansi lebih
dari 1,2 terhadap sinar ultraviolet pada panjang gelombang 260 nm (Adams et al. 1986). Bartram et al. (2009) melaporkan bahwa penyinaran larutan
glikogen dengan ultraviolet selama 30 menit sebelum digunakan bertujuan menghancurkan kontaminan berupa asam nukleat.
Warburg dan Christian (1942) menyajikan tiga prinsip dasar untuk pengukuran RNA-DNA, yaitu :
1) Asam nukleat kuat menyerap sinar pada 260 nm, 2) Protein menyerap sinar pada 280 nm, dan
3) Asam nukleat atau protein menyerap sinar pada 320 nm
Rasio spesifik dari nilai absorbansi yang ditentukan dari tiga prinsip di atas dapat digunakan untuk menentukan kemurnian dan jenis asam nukleat dalam sampel. Rumusan yang digunakan untuk mengukur asam nukleat ialah asam nukleat atau pun protein menyerap sinar pada 320 nm, oleh karena itu panjang gelombang ini digunakan untuk memperbaiki pembacaan yang didapat pada 260 nm dan 280 nm. Nilai absorbansi sampel pada panjang gelombang 260 nm dan 280 nm masing-masing dikoreksi dengan nilai absorbansi pada panjang gelombang 320 nm. 2.3.2 Penghitungan kadar asam nukleat
Persamaan yang digunakan untuk menghitung kadar asam nukleat adalah sebagai berikut (Adams et al. 1986) :
Kadar asam nukleat (mg/mL) = (0,064 x A260 nm) - (0,031 x A280 nm)
Warburg dan Christian (1942) melaporkan bahwa nilai absorbansi 1,0 pada 260 nm dengan koreksi latar pada 320 nm, setara dengan 50 µg/mL DNA, 40 µg/mL RNA atau DNA beruntai tunggal, atau 33 µg/mL oligonukleotida sintetik. Jika suatu sampel DNA memiliki absorbansi 0,345 pada 260 nm, maka konsentrasi DNA adalah 17,25 µg/mL, perhitungannya : 0,345 x 50 = 17,25.
2.4 Glikogen sebagai Ko-presipitan Low Copy Number DNA
Presipitasi DNA umumnya dilakukan setelah larutan ekstrak DNA ditambah dengan garam monovalen kationik yaitu sodium asetat, amonium asetat, sodium klorida atau lithium klorida diikuti dengan penambahan sejumlah tertentu etanol atau isopropanol lalu diinkubasi pada suhu rendah hingga asam nukleat terpresipitasi. Menurut McCormick (2009), untuk meningkatkan efisiensi
presipitasi DNA dengan ukuran kurang dari atau sama dengan 1 µg maka perlu ditambahkan suatu molekul ko-presipitan, karena selain dapat meningkatkan laju presipitasi, ko-presipitan yang ditambahkan dapat mempersingkat waktu yang dibutuhkan untuk mendapatkan asam nukleat dari larutannya. Glikogen dan
transfer ribonucleic acid (tRNA) adalah contoh ko-presipitan yang paling sering
digunakan. Glikogen sebagai ko-presipitan dalam presipitasi DNA yang baik karena memiliki sifat kelarutan dan presipitasi yang mirip dengan DNA. Hal ini karena kerangka utama penyusun asam nukleat adalah ribosa atau deoksiribosa yang dihubungkan oleh ikatan phosphodiester sehingga asam nukleat maupun glikogen larut di dalam air dan terpresipitasi ketika konstanta dielektrik
diturunkan dengan penambahan alkohol. Glikogen sangat baik sebagai ko-presipitan karena bermuatan netral dan tidak menghambat reaksi enzimatis
pada asam nukleat, sedangkan tRNA dapat mempengaruhi beberapa reaksi enzimatis seperti dengan kinase.
Gill (2001) melaporkan bahwa pembuatan profil low copy number DNA adalah suatu teknik yang cukup sensitif untuk menganalisis sel yang sangat sedikit. Analisis low copy number DNA dari sampel tulang tidak dilakukan tanpa terlebih dulu membuang lapisan paling luarnya dengan metode fisikawi untuk memperkecil kemungkinan kontaminasi pada DNA dari sumber lain. Sampel helai rambut harus dicuci dengan larutan detergen untuk membuang DNA yang tidak diinginkan. Ekstraksi DNA dan Polymerase Chain Reaction (PCR) dilakukan di laboratorium khusus, yaitu ruang amplifikasi PCR terpisah dari ruangan lainnya di laboratorium untuk memperkecil kemungkinan kontaminasi. Kontaminasi juga dapat diupayakan dengan memastikan setiap personil laboratorium menggunakan jas laboratorium, dan masker serta sarung tangan sekali pakai. Meja kerja dan peralatan juga harus sering dibersihkan dengan alkohol 70% (bleach alcohol) dan disinari sinar ultraviolet. Kontrol negatif digunakan pada setiap uji untuk memastikan tidak ada kontaminasi.