PENINGKATAN PERKECAMBAHAN BENIH ANDALIMAN ( Zanthoxylum acanthopodium DC.) MELALUI PENENTUAN
UMUR PANEN, SORTASI, DAN PEMATAHAN DORMANSI
DISERTASI
Oleh
BENEDICTA LAMRIA SIREGAR NIM: 138104004
Program Doktor (S3) Ilmu Pertanian
PROGRAM DOKTOR ILMU PERTANIAN PASCASARJANA FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA MEDAN
PENINGKATAN PERKECAMBAHAN BENIH ANDALIMAN ( Zanthoxylum acanthopodium DC.) MELALUI PENENTUAN
UMUR PANEN, SORTASI, DAN PEMATAHAN DORMANSI
DISERTASI
Oleh
BENEDICTA LAMRIA SIREGAR NIM: 138104004
Program Doktor (S3) Ilmu Pertanian
PROGRAM DOKTOR ILMU PERTANIAN PASCASARJANA FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA MEDAN
PENINGKATAN PERKECAMBAHAN BENIH ANDALIMAN ( Zanthoxylum acanthopodium DC.) MELALUI PENENTUAN
UMUR PANEN, SORTASI, DAN PEMATAHAN DORMANSI
DISERTASI
Oleh
BENEDICTA LAMRIA SIREGAR NIM: 138104004
Program Doktor (S3) Ilmu Pertanian
PROGRAM DOKTOR ILMU PERTANIAN PASCASARJANA FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA MEDAN
PENINGKATAN PERKECAMBAHAN BENIH ANDALIMAN (Zanthoxylum acanthopodium DC.) MELALUI PENENTUAN UMUR PANEN, SORTASI, DAN PEMATAHAN DORMANSI
DISERTASI
Sebagai Salah Satu Syarat untuk Memperoleh Gelar Doktor dalam Program Doktor Ilmu Pertanian pada Program Pascasarjana Fakultas Pertanian Universitas Sumatera Utara di bawah pimpinan Rektor Universitas Sumatera Utara
Prof. Dr. Runtung Sitepu, S.H., M.Hum.
Dipertahankan di hadapan Sidang Terbuka Senat Universitas Sumatera Utara pada tanggal 2 Juni 2020
Oleh
BENEDICTA LAMRIA SIREGAR NIM: 138104004
Program Doktor (S3) Ilmu Pertanian
PROGRAM DOKTOR ILMU PERTANIAN PASCASARJANA FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA MEDAN
Diuji pada Ujian Disertasi Terbuka (Promosi Doktor) Tanggal : 2 Juni 2020
__________________________________________________________________
PANITIA PENGUJI DISERTASI
Pemimpin Sidang :
Prof. Dr. Runtung Sitepu, S.H., M.Hum. (Rektor Universitas Sumatera Utara)
Ketua : Luthfi A.M. Siregar, S.P., M.Sc., Ph.D. Universitas Sumatera Utara Anggota : Prof. Dr. Ir. T. Chairun Nisa, M.Sc. Universitas Sumatera Utara : Dr. Ir. Chairani Hanum, M.P. Universitas Sumatera Utara : Ir. Revandy Damanik, M.Sc., Ph.D. Universitas Sumatera Utara : Dr. Ir. Syahbudin Hasibuan, M.Si. Universitas Medan Area
(Zanthoxylum acanthopodium DC.) Through Determination of Harvest Age, Sorting, and Breaking of Dormancy (under The Supervision of LUTHFI A.
M. SIREGAR, as Promotor, T. CHAIRUN NISA, as Co-Promotor).
Andaliman plant (Zanthoxylum acanthopodium DC.) is a local wild spice plant endemic to North Sumatra. Farmers still use wild seeds in propagating andaliman plants, because the seeds are difficult to germinate. Low germination and long germination may be due to dormancy. The cause of andaliman seed dormancy is unclear, so the effort to release the seed dormancy still showed low results. Low germination might also be caused by many seeds without embryos (empty seeds) and low seed viability.
The research aimed to describe the development of andaliman seeds to physiological maturity and determine the physiological maturity of the andaliman seeds; determine the causes of low andaliman seed germination; determine the causes of andaliman seed dormancy; get a sorting technique and breaking of andaliman seed dormancy which results in high germination. The general aim was to increase the seed germination of andaliman (Zanthoxylum acanthopodium DC.). The research consisted of four stages. The study used the flower, fruit, and seed of Simanuk accession plants in the Dairi Regency at 2o50’32.5" North Latitude, 98o24'45.4" East Longitude, at an altitude of 1300 m above sea level.
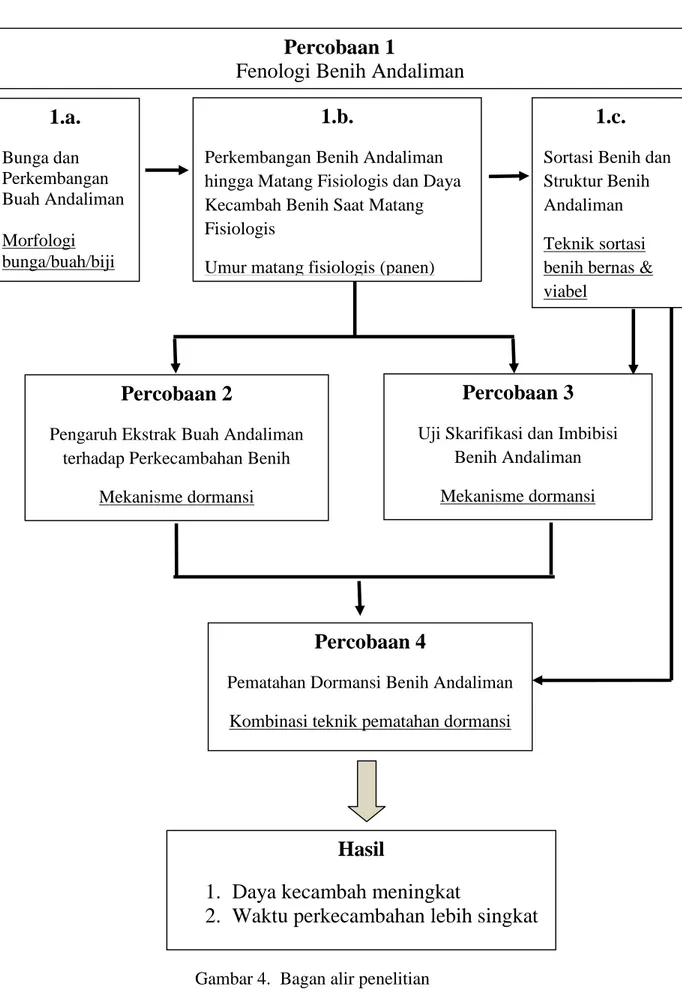
The first stage, Andaliman Seed Phenology, consisting of three sub-parts, namely (a). Flower and Development of Andaliman Fruit; (b) Development of Andaliman Seeds to Physiologically Mature and Andaliman Seed Germination;
(c) Sorting Andaliman Seeds and Andaliman Seed Anatomy. The dry weight of seeds increased up to 24-25 weeks after anthesis and did not increase significantly between the ages of 26-29 weeks after anthesis. The physiological mature age of andaliman seeds was achieved at the age of 28 week after anthesis, that was characterized by red exocarp, shiny black seed coat, seed moisture content of 17.26%, seed dry weight of 0.0068 g, seed dry weight percentage 82.74%, and percentage of seed germination 53.75%. The low of andaliman seed germination was caused that most of the seeds did not have embryo (empty seeds/wrinkled seeds), also the psresence of non-viable embryos due to immature or aging and deterioration. Sorting andaliman seeds with 15% sugar solution and sorting with water increased percentage of germination (60.0% and 57.5% respectively) and increased growth speed (1.153%/etmal and 1.172%/etmal respectively). The possibility to obtain filled and viable seeds was higher in sorting with a 15% sugar solution.
The second stage, Effect of Andaliman Fruit Extract on Seed Germination;
and the third stage, Scarification and Imbibition Test of Andaliman Seeds.
Andaliman seed dormancy was caused by several dormancy mechanisms, namely the hardness and thickness of seed coat, the presence of lignin on the seed coat, and germination inhibitor, terpenoid, in the seed coat and embryo. Germination inhibitor from terpenoid compounds was identified as geraniol and geranyl acetate.
with the highest growth rate, 1.517%/etmal; each difference in germination of 23.33%, time of germination 8.17 days, growth rate of 0.58%/etmal compared to treatment without breaking dormancy. The breaking technique of seed dormancy that can accelerate germination significantly had not been found in this study.
The results show that, the age of seed harvesting 28 weeks after anthesis, sorting with 15% sugar solution or water, and combination of broken dormancy seeds were heated in an oven 46oC for 30 minutes and then the seeds soaked in gibberellin 500 ppm for 10 hours, could improve seed germination.
Keywords: Zanthoxylum acanthopodium, andaliman, seed, viable, physiologically mature, embryo, sorting, inhibitor, terpenoid, geraniol, geranyl acetate, dormancy, imbibition, thick and hard seed coat, breaking dormancy, scarification, gibberellin
Andaliman (Zanthoxylum acanthopodium DC.) Melalui Penentuan Umur Panen, Sortasi, dan Pematahan Dormansi (dibawah bimbingan LUTHFI A. M.
SIREGAR, sebagai Promotor, T. CHAIRUN NISA, sebagai Co-Promotor).
Tanaman andaliman (Zanthoxylum acanthopodium DC.) merupakan tanaman rempah liar lokal endemik di Sumatera Utara. Petani masih menggunakan bibit liar dalam perbanyakan tanaman andaliman, karena bijinya sulit berkecambah. Daya kecambah yang rendah dan lamanya berkecambah diduga disebabkan adanya dormansi. Penyebab dormansi benih andaliman belum jelas, sehingga usaha memecahkan dormansi benihnya masih menunjukkan hasil yang rendah. Daya kecambah yang rendah kemungkinan juga disebabkan banyaknya biji tanpa embrio (biji kosong/biji keriput) dan biji tidak viabel.
Penelitian bertujuan menggambarkan perkembangan benih hingga matang fisiologis dan menentukan umur matang fisiologis benih andaliman; menentukan penyebab rendahnya daya kecambah benih andaliman; menentukan penyebab dormansi benih andaliman; mendapatkan teknik sortasi dan pematahan dormansi benih yang menghasilkan daya kecambah yang besar. Tujuan umum penelitian meningkatkan daya kecambah benih andaliman (Zanthoxylum acanthopodium DC.). Penelitian terdiri atas empat tahap. Penelitian menggunakan bunga, buah, dan benih dari tanaman aksesi Simanuk di Kabupaten Dairi berada pada 2o50'32,5” Lintang Utara, 98o24'45,4” Bujur Timur, pada ketinggian 1300 m di atas permukaan laut.
Penelitian tahap pertama, yakni Fenologi Benih Andaliman, terdiri atas tiga sub bagian, yaitu (a). Bunga dan Perkembangan Buah Andaliman; (b) Perkembangan Benih Andaliman Hingga Matang Fisiologis dan Daya Kecambah Benih Andaliman Saat Matang Fisiologi; (c) Sortasi Benih Andaliman dan Struktur Benih Andaliman. Hasil penelitian menunjukkan bobot kering benih terus meningkat hingga 24-25 minggu setelah antesis dan tidak meningkat lagi secara signifikan pada kisaran umur 26-29 minggu setelah antesis. Umur matang fisiologis benih andaliman dicapai pada umur benih 28 minggu setelah antesis, dengan ciri: wama eksokarp merah, kulit biji hitam mengkilap, kadar air benih 17,26%, bobot kering benih 0,0068 g, persentase bobot kering benih mencapai 82,74%, dan daya kecambah sebesar 53,75%. Daya kecambah benih andaliman rendah disebabkan sebagian besar biji tidak memiliki embrio (biji kosong/biji keriput), juga adanya biji tidak viabel akibat belum matang ataupun mengalami penuaan dan kerusakan (deteriorasi). Sortasi benih andaliman dengan larutan gula 15% dan sortasi dengan air meningkatkan daya kecambah (masing- masing 60,0% dan 57,5%) dan meningkatkan kecepatan tumbuh (masing-masing 1,153 %/etmal dan 1,172 %/etmal). Kepastian untuk mendapatkan benih yang bernas dan viabel lebih besar pada sortasi dengan larutan gula.
Penelitian tahap kedua, yakni Pengaruh Ekstrak Buah Andaliman terhadap Perkecambahan Benih; dan penelitian tahap ketiga, yakni Uji Skarifikasi dan Imbibisi Benih Andaliman. Benih andaliman dorman disebabkan oleh beberapa mekanisme dormansi, yaitu kekerasan dan ketebalan kulit biji, adanya lignin pada
dormansi benih andaliman dipanaskan dalam oven 46oC selama 30 menit dan selanjutnya benih direndam dalam giberelin 500 ppm selama 10 jam memberi daya kecambah tertinggi, yakni sebesar 82,22 %, dengan umur berkecambah tercepat 58,20 hari, dengan kecepatan tumbuh tertinggi, yakni 1,517 %/etmal;
masing-masing selisih daya kecambah 23,33%, umur berkecambah 8,17 hari, kecepatan tumbuh 0,58 %/etmal dibanding dengan perlakuan tanpa pematahan dormansi. Teknik pematahan dormansi yang dapat mempercepat perkecambahan secara nyata belum ditemukan dalam penelitian ini.
Hasil penelitian menunjukkan bahwa umur panen benih 28 minggu setelah antesis, sortasi dengan larutan gula 15% atau air, dan kombinasi pematahan dormansi benih dipanaskan dalam oven 46oC selama 30 menit dan selanjutnya benih direndam dalam giberelin 500 ppm selama 10 jam, dapat meningkatkan daya kecambah benih andaliman.
Kata kunci: Zanthoxylum acanthopodium, andaliman, benih, viabel, matang fisiologis, embrio, sortasi, senyawa penghambat, terpenoid, geraniol, geranil asetat, dormansi, imbibisi, kulit benih tebal dan keras, pematahan dormansi, skarifikasi, giberelin.
atas bimbingan, lindungan, serta berkatNya bagi penulis sehingga penulis dapat menyelesaikan penulisan disertasi ini.
Selama melakukan penelitian dan penulisan disertasi ini ,penulis banyak memperoleh bantuan moril dan materil dari berbagai pihak. sehingga penulis dapat menyelesaikan Oleh karena itu, pada kesempatan ini penulis menyampaikan rasa hormat dan ucapan terima kasih kepada:
1. Bapak Prof. Dr. Runtung Sitepu, S.H., M.Hum. selaku Rektor Universitas Sumatera Utara.
2. Bapak Prof. Dr. Robert Sibarani, M.S. selaku Direktur Sekolah Pascasarjana Universitas Sumatera Utara.
3. Bapak Dr. Ir. Hasanuddin, M.S. selaku Dekan Fakultas Pertanian Universitas Sumatera Utara.
4. Bapak Prof Ir. Edison Purba, Ph.D., selaku Ketua PS Doktor Ilmu Pertanian, Universitas Sumatera Utara dan Ibu Dr. Lisnawita, S.P., M.Si. selaku Sekretaris PS Doktor Ilmu Pertanian, Universitas Sumatera Utara.
5. Bapak Luthfi A.M. Siregar, S.P., M.Sc., Ph.D sebagai promotor atas waktu, pemikiran, masukan, dan dorongan yang diberikan selama pembuatan proposal, pelaksanaan penelitian, dan penulisan disertasi.
6. Ibu Prof. Dr. Ir. T. Chairun Nisa, M.Sc. sebagai co promotor yang dengan tulus memberi waktu, pemikiran, masukan, dan motivasi yang diberikan selama pembuatan proposal, pelaksanaan penelitian, dan penulisan disertasi.
dan pelaksanaan penelitian.
8. Ibu Dr. Ir. Chairani Hanum, M.P. dan almarhum Bapak Prof. Dr. Ir. J. A.
Napitupulu, M.Sc. yang bersedia dan memberi waktu untuk menyampaikan masukan dan pertanyaan bagi pengembangan proposal dan persiapan penelitian sewaktu seminar proposal. Ibu Dr. Ir. Chairani Hanum, M.P., Bapak Dr. Ir. Syahbudin Hasibuan, M.Si., dan Bapak Ir. Revandy Damanik, M.Sc., Ph.D. sebagai penguji luar komisi yang bersedia dan memberi waktu untuk menyampaikan masukan dan pertanyaan bagi pendalaman dan pengembangan pemahaman penulis dan penyempurnaan disertasi ini.
9. Bapak Prof Dr. Ir. Abdul Rauf, M.P., sebagai Ketua PS Doktor Ilmu Pertanian, Universitas Sumatera Utara dan Dr. Ir. Hamidah Hanum, M.P.
sebagai Sekretaris PS Doktor Ilmu Pertanian, Universitas Sumatera Utara, di awal masa pendidikan, dan ketika pembuatan proposal dan penelitian ini dimulai.
10. Bapak Prof. Dr. Ir. Abdul Rauf, M.P., almarhum Bapak Prof. Dr. Ir. J. A.
Napitupulu, M.Sc., Ibu Prof. Dr. Ir. T. Chairun Nisa M.Sc., dan almarhumah Ibu Dr. Ir. Lollie Agustina P. Putri, M.Si., yang telah berbagi pemikiran dan memberi masukan untuk penyempurnaan proposal dalam prakualifikasi.
11. Kementerian Riset Teknologi dan Perguruan Tinggi, atas pemberian Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN) kepada penulis.
bapak/ibu pegawai yang telah membantu admisnistrasi pengurusan izin.
13. Yayasan Universitas HKBP Nommensen yang telah memberi bantuan biaya kepada penulis dalam pendidikan Doktor.
14. Rektor Universitas HKBP Nommensen, Dekan Fakultas Pertanian Universitas HKBP Nommensen, dan Ketua Program Studi Agroteknologi Universitas HKBP Nommensen atas pemberian izin tugas belajar kepada penulis untuk melanjutkan pendidikan Doktor. Juga bapak/ibu pegawai yang telah membantu admisnistrasi pengurusan izin dan pengurusan bantuan biaya pendidikan.
15. Bapak dan ibu dosen yang telah membekali penulis selama mengikuti perkuliahan di Program Studi Doktor Ilmu Pertanian, Universitas Sumatera Utara. Almarhumah Ibu Dr. Ir. Lollie Agustina P. Putri, M.Si. yang telah mendorong penulis untuk mendalami andaliman melalui penelitian disertasi.
Ibu Dr. Ir. Chairani Hanum, M.P. yang telah berbagi pendapat, sehingga penulis memutuskan untuk meneliti andaliman dalam penelitian disertasi.
16. Ibu Wiwik Kusdiningsih dan pegawai lain Sekolah Pascasarjana Universitas Sumatera Utara yang telah membantu adiministrasi pengurusan pendaftaran dan beasiswa BPPDN.
17. Ibu Runi Siahaan dan Ibu Sri Wahyuni, sebagai pegawai PS Doktor Ilmu Pertanian, Universitas Sumatera Utara, yang telah membantu penulis dalam pengurusan administrasi terkait perkuliahan dan penelitian disertasi.
19. Pimpinan/kepala laboatorium/laboran Unit Pelaksana Teknis Sertifikasi Benih Tanaman Pangan dan Hortikultura Dinas Tanaman Pangan dan Hortikultura Propinsi Sumatera Utara.
20. Pimpinan/pegawai/laboran Unit Pelaksana Teknis Benih Induk Hortikultura Gedung Johor Dinas Tanaman Pangan dan Hortikultura Propinsi Sumatera Utara
21. Pimpinan/dosen/asisten Laboratorium Fitokimia Fakultas Farmasi Universitas Sumatera Utara.
22. Pimpinan/dosen/pegawai/laboran Laboratorium Penelitian Fakultas Farmasi Universitas Sumatera Utara.
23. Pimpinan dan laboran Laboratorium Pusat Penelitian Biomaterial, LIPI, di Cibinong.
24. Pimpinan dan laboran Laboratorium Morfologi, Anatomi dan Sitologi, Pusat Penelitian Biologi, LIPI, di Cibinong.
25. Pimpinan dan asisten Laboratorium Teknologi Pangan Fakultas Pertanian Universitas Sumatera Utara.
26. Dekan Fakultas Pertanian Universitas Katolik St. Thomas, pimpinan dan laboran Laboratorium Fakultas Pertanian, Universitas Katolik St. Thomas.
27. Pimpinan dan laboran Laboratorium Fakultas Kedokteran dan Fakultas Pertanian, Universitas HKBP Nommensen
28. Bapak Simon Petrus Hutauruk dan bapak Ir. Rapen Nainggolan yang telah membantu dalam pengenalan awal tanaman andaliman dan lokasi penelitian.
Tamba, Parningotan Siregar, S.P., Sara Anggelina, S.P., Febrianto, Ir. Conos Tamba, alumni/mahasiswa/mahasiswi Fakultas Pertanian Universitas Katolik St. Thomas yang telah membantu di lapangan. Saudara Hendra Gunawan, S.
Farm., Ibu Herawaty Pane S.P., Ir. Nuriman Tambunan, Bapak Negara Simbolon, saudara Dominggo A.T. Nainggolan, S.P. yang telah membantu di laboratorium. Kepada keluarga bapak Elias Benjamin Nainggolan dan ibu Romiannna Sinabutar, serta penduduk di Desa Linggaraja II atas bantuan yang diberikan. Kepada semua pihak yang tak dapat disebutkan satu persatu, termasuk semua orang yang tanpa sepengetahuan penulis juga berkontribusi.
29. Ibu Ir. Donna Sinambela, M.P., (Kepala Laboratorium UPT Sertifikasi Benih Tanaman Pangan dan Hortikultura Dinas Tanaman Pangan dan Hortikultura Propinsi Sumatera Utara), Bapak Dr. rer. nat. Sunaryo (Kepala Laboratorium Morfologi, Anatomi dan Sitologi, Pusat Penelitian Biologi, LIPI, di Cibinong.), dan Ibu Dr. Ir. Hariati, M.P. (Kepala Laboratorium Benih Fakultas Pertanian Universitas Sumatera Utara ) yang telah banyak memberi masukan.
30. Teman-teman mahasiswa seangkatan 2013, angkatan 2014, dan teman-teman angkatan lainnya di Program Studi Doktor Ilmu Pertanian Universitas Sumatera Utara.
Ucapan terima kasih dan rasa hormat kepada keluarga bapak Daniel Situmeang dan ibu Relli Sinabutar serta keluarga bapak Febrianto dan Ibu Marsaulina Friska Sinaga, keluarga almarhumah Ibu Kosti boru Nainggolan, yang telah memberi tanaman andaliman sebagai bahan penelitian. Kepada
Penulis menyampaikan rasa hormat dan terima kasih kepada ibunda tercinta Ibu S. M. Sitorus yang senantiasa berdoa buat penulis, memotivasi, dan menemani keluarga selama penelitian berlangsung; kepada suami terkasih Sictus Hutauruk yang telah mengizinkan dan mendukung penulis, serta atas pengertiannya pada penulis selama mengikuti pendidikan Doktor; kepada anak- anak Laurentia Hutauruk, Doli Benedictus Hutauruk, Sabam Aloysius Hutauruk atas doa, dukungan, pengorbanan, dan pengertiannya selama penulis mengikuti pendidikan. Penulis juga berterima kasih kepada keluarga besar Siregar dan Hutauruk, pastor, dan para sahabat yang mendoakan penulis selama mengikuti program pendidikan Doktor.
Penulis tidak akan berhasil dalam program pendidikan doktor, tanpa semua orang yang telah berkontribusi kepada penulis. Penulis mempersembahkan tulisan ini bagi petani andaliman, ibunda tercinta, dan semua orang yang telah menjadi guru bagi penulis.
Penulis menyadari disertasi ini masih memiliki kekurangan dan belum sempurna. Penulis mengharapkan kiranya tulisan ini dapat berguna bagi petani andaliman, dosen, mahasiswa, peneliti, dan pihak lainnya yang memerlukan informasi tentang tanaman andaliman dan hendak menggunakan sebagai acuan penelitian dan pengembangan tanaman andaliman. Semoga Tuhan Yang Maha Pengasih memberkati kita semua. Amin.
Medan, Juni 2020 Penulis,
ibu S. M. Sitorus. Pada tahun 1994 penulis menikah dengan Sictus Hutauruk, dan dikaruniai tiga orang anak, Laurentia Hutauruk, S.TP., Doli Benedictus Hutauruk, dan Sabam Aloysius Hutauruk.
Penulis mulai mengenyam pendidikan sejak tahun 1972 di TK RK Antonius Medan, melanjutkan di SD RK Antonius V Medan (lulus tahun 1980), melanjutkan di SMP RK Tri Sakti Medan (lulus tahun 1983), dan melanjutkan di SMA Negeri 5 Medan (lulus tahun 1986). Pada tahun 1986 penulis diterima di Institut Pertanian Bogor melalui jalur Penelusuran Minat dan Kemampuan (PMDK) dan mengikuti pendidikan strata 1 di Program Studi Agronomi dan menyelesaikannya pada tahun 1991. Pada tahun 1995, penulis melanjutkan pendidikan strata 2 di Program Studi Agronomi Program Pascasarjana Universitas Sumatera Utara dengan biaya Beasiswa Pendidikan Pascasarjana (BPPS) dan menyelesaikannya pada tahun 1999. Pada tahun 2013 penulis mendapat kesempatan untuk melanjutkan pendidikan strata 3 di Program Studi Doktor Ilmu Pertanian Universitas Sumatera Utara dengan biaya Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN).
Penulis mulai bekerja di bidang pendidikan sejak tahun 1992 di Program Studi Agronomi Fakultas Pertanian Universitas Katolik St. Thomas di Medan hingga tahun 2005. Sejak tahun 2005 hingga sekarang penulis sebagai dosen Lembaga Layanan Pendidikan Tinggi (LLDIKTI) Wilayah I dpk Program Studi Agroteknologi Fakultas Pertanian Universitas HKBP Nommensen di Medan.
Halaman
SUMMARY... i
RINGKASAN ... iii
KATA PENGANTAR ... v
RIWAYAT HIDUP ... xi
DAFTAR ISI ... xii
DAFTAR TABEL ... xiv
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xx
BAB I PENDAHULUAN ... 1
1.1. Latar Belakang ... 1
1.2. Perumusan Masalah ... 5
1.3. Tujuan Penelitian ... 5
1.4. Manfaat Penelitian ... 6
BAB II TINJAUAN PUSTAKA ... 7
2.1. Tanaman Andaliman dan Potensi Pemanfaatannya ... 7
2.2. Pembentukan Biji dan Perkecambahan ... 11
2.3. Dormansi Benih ... 16
2.4. Perkecambahan Benih Andaliman ... 24
2.5. Kerangka Konseptual Penelitian ... 29
2.6. Hipotesis Penelitian ... 32
BAB III. FENOLOGI BENIH ANDALIMAN ... 33
Abstrak ... 33
3.1. Pendahuluan ... 35
3.2. Bahan dan Metode ... 40
3.3. Hasil dan Pembahasan ... 50
3.4. Kesimpulan dan Saran ... 94
BAB IV. PENGARUH EKSTRAK BUAH ANDALIMAN TERHADAP PERKECAMBAHAN BENIH ... 97
Abstrak ... 97
4.1. Pendahuluan ... 98
4.2. Bahan dan Metode ... 101
4.3. Hasil dan Pembahasan ... 107
4.4. Kesimpulan dan Saran ... 118
5.2. Bahan dan Metode ... 124
5.3. Hasil dan Pembahasan ... 128
5.4. Kesimpulan dan Saran ... 139
BAB VI. PEMATAHAN DORMANSI BENIH ANDALIMAN ... 141
Abstrak ... 141
6.1. Pendahuluan ... 142
6.2. Bahan dan Metode ... 146
6.3. Hasil dan Pembahasan ... 151
6.4. Kesimpulan dan Saran ... 166
BAB VII. PEMBAHASAN UMUM ... 167
BAB VIII. KESIMPULAN DAN SARAN ... 178
8.1. Kesimpulan ... 178
8.2. Saran ... 179
DAFTAR PUSTAKA ... 180
LAMPIRAN ... 192
No. Judul Halaman 1. Daya kecambah (%) saat 8 MSP, 10 MSP, 12 MSP, 14 MSP
pada 4 umur panen benih andaliman ... 73 2. Potensi tumbuh maksimum (PTM) dan daya kecambah (DB)
saat penghitungan akhir, umur berkecambah dan kecepatan
tumbuh pada 4 umur panen benih andaliman... 74 3. Daya kecambah dan potensi tumbuh maksimum benih
andaliman pada beberapa perlakuan sortasi benih... 82 4. Umur berkecambah dan kecepatan tumbuh benih andaliman pada
beberapa perlakuan sortasi benih... 83 5. Jumlah dan persentase benih yang tenggelam dan terapung pada
dua metode sortasi... 84 6. Jumlah dan persentase benih tenggelam dan terapung pada
sortasi bertahap... 85 7. Hasil uji tetrazolium benih andaliman pada dua metode sortasi.... 87 8. Hasil pengecambahan benih yang mengapung hasil sortasi... 91 9. Potensi tumbuh maksimum (PTM) benih padi saat 5 HSP, 7
HSP, dan 12 HSP akibat perlakuan perendaman benih... 108 10. Daya kecambah benih padi 5 HSP, 7 HSP, dan 12 HSP, umur
berkecambah dan kecepatan tumbuh akibat perlakuan
perendaman benih... 108 11. Panjang plumula, panjang akar, dan jumlah akar seminal
kecambah padi akibat perlakuan perendaman benih... 110 12. Bobot kering kecambah, bobot segar dan bobot kering plumula,
bobot segar dan bobot kering akar kecambah padi akibat
perlakuan perendaman benih... 111 13. Hasil pengecambahan dan hasil uji Tetrazolium benih padi
akibat perlakuan direndam ektrak buah andaliman... 113 14. Hasil skrining fitokimia ekstrak heksan buah andaliman... 114
teknik pematahan dormansi benih ... 153 18. Daya kecambah benih andaliman pada beberapa teknik
pematahan dormansi benih ... 155 19. Umur berkecambah dan kecepatan tumbuh benih andaliman
pada beberapa teknik pematahan dormansi benih ... 157
1. Tahap perkembangan fisiologis biji (Hartmann, et al., 2011)... 12
2. Reaksi kimia yang terlibat dalam perubahan tetrazolium chloride yang tidak berwarna menjadi formazan berwarna merah (Elias et al., 2012) ... 14
3. Kerangka konseptual penelitian ... 30
4. Bagan alir penelitian ... 31
5. Bunga majemuk andaliman, aksilar (a) dan berbatas (b)... 51
6. Jumlah bunga dalam satu rangkaian dan ibu tangkai bervariasi .. 51
7. Perkembangan jumlah bunga mekar pada 10 rangkaian rakis ... 52
8. Perkembangan buah (a,b,c,d) dan pematangan buah (e,f) beragam... ... 52
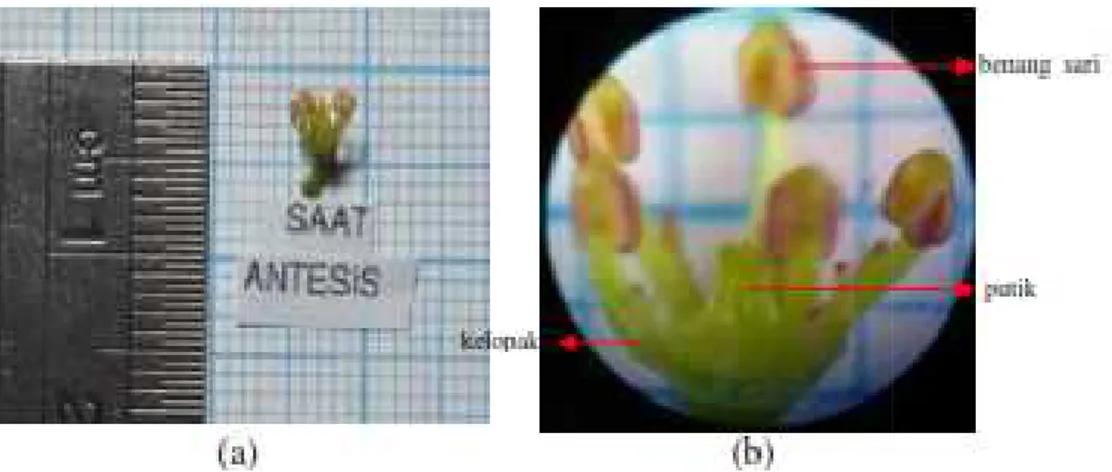
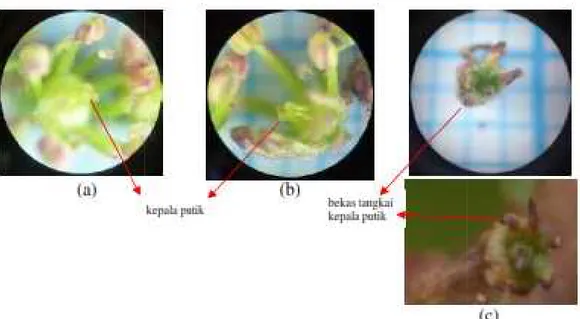
9. Bunga andaliman berukuran kecil (a), bunga andaliman dengan bagian-bagiannya (kelopak, benang sari, dan putik) perbesaran 10x4 (b) ... 53
10. Benang sari bunga andaliman dan bagian-bagiannya, bunga dengan lima benang sari a); bunga dengan enam benang sari (b), bunga, perbesaran 10x4 (c), kepala sari, perbesaran 10x10 (d), kepala sari, perbesaran 10x40 (e)... 54
11. Bunga andaliman dengan kepala putik berjumlah tiga (a), empat (b); buah muda dengan tangkai kepala putik berjumlah lima (c); perbesaran 10x4 ... 55
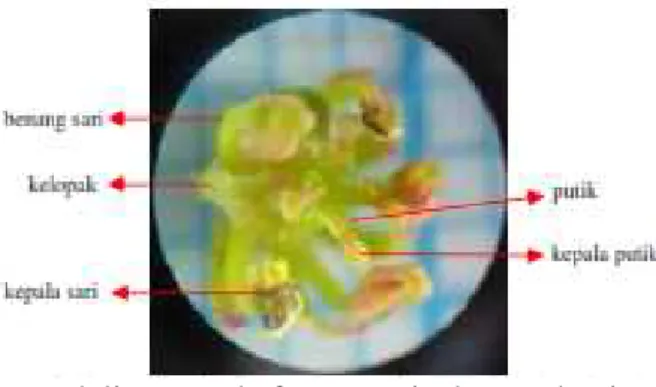
12. Bunga andaliman pada fase antesis dengan bagian-bagiannya (kelopak, benang sari, dan putik); perbesaran 10x4... 56
13. Perkembangan buah saat 2 minggu setelah antesis (MSA) di tanaman (a), di atas kertas milimeter (b), dan mikroskop perbesaran 10x4 ... 57
14. Perkembangan buah saat 4 minggu setelah antesis (MSA) di tanaman (a), di atas kertas milimeter (b), dan mikroskop perbesaran 10x4 ... 57
16. Perkembangan buah saat 8 minggu setelah antesis (MSA) di tanaman (a), di atas kertas milimeter (b), dan mikroskop
perbesaran 10x4 (c,d) ... 58 17. Perkembangan buah saat 10 minggu setelah antesis di tanaman
(a), di atas kertas milimeter (b), dan mikroskop perbesaran 10x4
(c,d) ... 58 18. Perkembangan buah saat 12 minggu setelah antesis di tanaman
(a), di atas kertas milimeter (b), dan mikroskop perbesaran 10x4
(c,d) ... 59 19. Perkembangan buah saat 14 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah dan biji di atas kertas
milimeter (c), buah di mikroskop perbesaran 10x4 (d, e, f) ... 59 20. Perkembangan buah saat 16 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah dan biji di atas kertas milimeter (c), buah (d), sayatan buah (e), biji (f); mikroskop
perbesaran 10x4 (d, e, f) ... 60 21. Perkembangan buah saat 17 minggu setelah antesis di tanaman
(a), sayatan buah (b), biji (c); mikroskop perbesaran 10x4 (b,c) .. 60 22. Perkembangan buah saat 18 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah (c), sayatan buah (d,
e), biji (f); mikroskop perbesaran 10x4 (c, d, e, f) ... 61 23. Perkembangan buah saat 20 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah (c), sayatan buah (d),
biji (e); mikroskop perbesaran 10x4 (c, d, e) ... 62 24. Perkembangan buah saat 22 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah (c), sayatan buah (d),
biji (e); mikroskop perbesaran 10x4 (c, d, e) ... 62 25. Perkembangan buah saat 24 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah (c), sayatan buah (d),
biji (e); mikroskop perbesaran 10x4 (c, d, e) ... 63 26. Perkembangan buah saat 25 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b) ... 63 27. Perkembangan buah saat 26 minggu setelah antesis di tanaman
28. Perkembangan buah saat 27 minggu setelah antesis di tanaman (a), buah dan biji di atas kertas milimeter (b), buah (c), sayatan buah (d), sayatan biji (e), biji (f); mikroskop perbesaran 10x4 (c,
d, e, f) ... 64 29. Perkembangan buah saat 28 minggu setelah antesis di tanaman
(a), buah dan biji di atas kertas milimeter (b), buah (c), sayatan buah (d), sayatan biji (e), biji (f); mikroskop perbesaran 10x4 (c,
d, e, f) ... 65 30. Perkembangan buah saat 29 minggu setelah antesis di tanaman
(a), buah di atas kertas milimeter (b), buah (c,d); mikroskop
perbesaran 10x4 (c, d) ... 65 31. Proses keluarnya biji dari buah, mikroskop perbesaran 10x4
(a,b,c,d); biji yang sudah keluar di tanaman (e,f) ... 66 32. Buah apokarp, yang berasal dari satu bunga, terdiri atas 1-5 buah
(a), 5 buah (b), 7 buah (c), 9 buah (d) ... 67 33. Dalam 1 buah terdapat 2 biji (a); biji berasal dari buah dengan 2
biji (b, c; perbesaran 10x4) ... 67 34. Perbedaan warna kulit buah pada satu tangkai buah (a,b,c);
perbedaan warna kulit biji pada satu buah (d) ... 68 35. Laju perkembangan bobot kering (g) biji andaliman ... 69 36. Laju perkembangan bobot segar (g), bobot kering (g), kadar air
(x100%), dan persentase bobot kering (x100%) biji andaliman ... 70 37. Keluarnya radikula pada proses perkecambahan (a); perkecam-
bahan andaliman tipe epigea (b) ... 71 38. Perkembangan kecambah berturut-turut: saat berkecambah, 1, 2,
5, 7, 9, 10, 14, 21, 28 hari setelah berkecambah ... 71 39. Perkembangan daya kecambah (%) benih andaliman umur 29-
117 hari pada umur panen yang berbeda ... 72 40. Saat pasca antesis yang serempak (a) dan tidak serempak (b,c)
antar kepala putik (perbesaran 10x4) ... 79 41. Perkembangan daya kecambah (%) benih andaliman umur 39-90
43. Sayatan benih andaliman hasil uji tetrazolium yang terapung
pada metode sortasi dengan air (perbesaran 10x4) ... 88 44. Sayatan benih andaliman hasil uji tetrazolium yang terapung
pada metode sortasi dengan larutan gula (perbesaran 10x4) ... 89 45. Sayatan benih andaliman hasil uji tetrazolium yang tenggelam
pada metode sortasi dengan air (perbesaran 10x4) ... 90 46. Sayatan benih andaliman hasil uji tetrazolium yang tenggelam
pada metode sortasi dengan larutan gula (perbesaran 10x4) ... 91 47. Kecambah padi akibat perlakuan tanpa perendaman (S0), benih
direndam dengan aquadest (S1), benih direndam dengan ekstrak heksan buah andaliman (S2) (a); dan hasil uji Tetrazolium untuk
perlakuan benih padi direndam ektrak buah andaliman (b,c) ... 113 48. Kromatogram ekstrak heksan andaliman ... 116 49. Jalur biosintesis geraniol dalam serai (Cymbopogon flexuosus)
mutan cv. GRL-1 (Ganjewala dan Luthra, 2009) ... 118 50. Perkembangan penambahan jumlah air imbibisi (g) benih
andaliman tanpa skarifikasi selama 72 jam ... 129 51. Perkembangan penambahan jumlah air imbibisi (g) benih
andaliman selama, 24 jam, 48 jam, dan 72 jam di atas kertas
saring pada berbagai perlakuan skarifikasi ... 130 52. Jumlah air imbibisi (g) benih andaliman selama 72 jam pada
berbagai perlakuan skarifikasi ... 131 53. Kekerasan benih andaliman (kg/cm2) setelah berbagai perlakuan
skarifikasi ... 133 54. Perkembangan daya kecambah benih andaliman saat 16-115 hari
pada berbagai perlakuan pematahan dormansi (P0:tanpa pematahan dormansi; P1:air panas 60oC 24 jam; P2:oven 46oC 30 menit; P3:KNO3 0,6 g/l 24 jam; P4:air panas 60oC 24 jam, GA 500 ppm 5 jam; P5:air panas 60oC 24 jam, GA 500 ppm 10 jam; P6:oven 46oC 30 menit, GA 500 ppm 5 jam; P7:oven 46oC 30 menit, GA 500 ppm 10 jam; P8:KNO30,6 g/l 24 jam, GA 500
ppm 5 jam; P9:KNO 0,6 g/l 24 jam, GA 500 ppm 10 jam) ... 152
No. Judul Halaman 1. Perkembangan jumlah bunga mekar pada 10 rangkaian rakis... 192 2. Perkembangan bobot segar, bobot kering, kadar air, dan
persentase bobot kering biji andaliman ... 193 3. Perkembangan daya kecambah benih andaliman umur 29-117
hari pada umur panen benih yang berbeda ... 194 4. Rataan berbagai peubah perkecambahan benih andaliman akibat
pengaruh umur panen benih yang berbeda... 195 5. Sidik ragam berbagai peubah perkecambahan benih andaliman
akibat pengaruh umur panen benih yang berbeda ... 196 6. Uji beda rataan daya kecambah (DB) saat 10 MSP, 12 MSP, 14
MSP dan pada penghitungan akhir, potensi tumbuh maksimum (PTM) pada penghitungan akhir, serta kecepatan tumbuh (KCT)
pada 4 umur panen benih andaliman ... 197 7. Rataan kekerasan benih (kg/cm2) pada umur panen benih yang
berbeda... 198 8. Sidik ragam kekerasan benih (kg/cm2) pada umur panen benih
yang berbeda ... 198 9. Perkembangan daya kecambah benih andaliman umur 39-90 hari
pada tiga perlakuan sortasi benih ... 199 10. Rataan berbagai peubah perkecambahan benih andaliman akibat
pengaruh perlakuan sortasi benih... 200 11. Sidik ragam berbagai peubah perkecambahan benih andaliman
akibat pengaruh perlakuan sortasi benih... 200 12. Uji beda rataan daya berkecambah (DB) benih andaliman saat 8
MSP, 12 MSP, dan penghitungan akhir, potensi tumbuh maksimum (PTM) penghitungan akhir serta kecepatan
tumbuh (KCT) pada beberapa perlakuan sortasi benih ... 201 13. Jumlah dan persentase benih yang tenggelam dan terapung pada
dua metode sortasi... 202
kecambah (DB), umur berkecambah, dan kecepatan tumbuh
benih padi akibat perlakuan perendaman benih ... 205 16. Sidik ragam berbagai peubah kecambah padi akibat pengaruh
perlakuan perendaman benih ... 206 17. Uji beda rataan panjang plumula, panjang akar, jumlah akar
seminal, bobot kering kecambah, bobot segar dan bobot kering plumula serta akar kecambah padi akibat perlakuan perendaman
benih ... 206 18. Korelasi antar peubah perkecambahan dan kecambah padi... 207 19. Perkembangan perubahan bobot 20 benih andaliman (g) sejak
dikupas hingga 96 jam pada berbagai perlakuan skarifikasi... 208 20. Perkembangan penambahan jumlah air yang diserap benih
andaliman selama 72 jam di atas kertas saring pada berbagai
perlakuan skarifikasi... 208 21. Sidik ragam jumlah air imbibisi dan kekerasan benih pada
berbagai perlakuan skarifikasi... 208 22. Uji beda rataan jumlah air imbibisi benih andaliman selama 72
jam dan kekerasan benih pada berbagai perlakuan skarifikasi ... 209 23. Komposisi kimia benih andaliman... 209 24. Perkembangan daya kecambah benih andaliman umur 16-115
hari pada berbagai perlakuan pematahan dormansi... 210 25. Rataan potensi tumbuh maksimum (PTM) benih andaliman
akibat pengaruh teknik pematahan dormansi benih yang berbeda 215 26. Sidik ragam potensi tumbuh maksimum (PTM) benih andaliman
akibat pengaruh teknik pematahan dormansi benih yang berbeda 215 27. Uji beda rataan potensi tumbuh maksimum benih andaliman saat
10 MSP, dan 12 MSP pada beberapa teknik pematahan dormansi
benih... 216 28. Rataan daya berkecambah (DB) benih andaliman akibat
pengaruh teknik pematahan dormansi benih yang berbeda... 216
30. Uji beda rataan daya kecambah benih andaliman saat saat 10, 12,
dan 14 MSP pada beberapa teknik pematahan dormansi benih .... 218
BAB I. PENDAHULUAN
1.1. Latar Belakang
Salah satu tanaman rempah liar yang dijumpai di Sumatera Utara adalah andaliman (Zanthoxylum acanthopodium DC.). Tanaman ini masih ditemui pada daerah tertentu saja di Propinsi Sumatera Utara (tanaman endemik). Buahnya dapat meningkatkan nafsu makan dan digunakan pada berbagai masakan tradisional suku Batak.
Buah andaliman mengandung senyawa aromatik, yang memiliki sensasi rasa yang unik dengan rasa pedas dan getir yang khas, serta hangat. Berbagai penelitian dan publikasi telah mengungkap kandungan kimia dan aktivitas biologisnya, sehingga saat ini andaliman diperhitungkan menjadi sumber senyawa aromatik dan minyak esensial. Beberapa penelitian telah membuktikan andaliman mempunyai aktivitas antimikrob (Tarigan, 1999; Siahaan, 2000;
Surbakti, 2002; Parhusip et al., 2003; Parhusip, 2004a; Parhusip, 2004b; Parhusip, et al., 2005); antioksidan (Siahaan, 2000; Tensiska et al., 2003; Parhusip et al., 2003; Suryanto, et al., 2004; Kristanty et al., 2012; Kristanty dan Suriawati, 2014), sitotoksik (Sabri, 2007; Kristanty dan Suriawati, 2014). Hal ini memberi peluang bagi andaliman sebagai bahan baku bagi industri pangan, farmasi, fungisida, dan bakterisida.
Dilain pihak aspek budidayanya masih sangat terbatas diketahui.
Tanamannya juga jarang dikenal, karena tidak dibudidayakan secara luas dan khusus. Di daerah asalnya seperti Kabupaten Dairi, Toba, dan Tapanuli Utara, tanaman andaliman relatif masih liar dan jarang dibudidayakan secara khusus.
Umumnya petani memilikinya dengan memelihara tumbuhan yang tumbuh liar di
ladangnya. Oleh karena itu upaya menggali teknik budidaya tanaman andaliman perlu mendapat perhatian. Salah satu aspek budidaya yang perlu dipelajari adalah perbanyakan bahan tanam. Petani masih menggunakan bibit liar dalam perbanyakannya, karena bijinya sulit berkecambah. Menurut petani perkecambahan biji andaliman tergantung burung (Siregar, 2003). Ada petani yang memperoleh bibit di bekas tempat pembakaran gulma di sekitar tanaman tua (Siregar, 2001). Ini menjadi salah satu hambatan bagi kebanyakan petani untuk memperbanyaknya dan membudidayakannya. Padahal untuk menghasilkan produk, salah satu aspek yang perlu dipertimbangkan dalam industri adalah kuantitas dan kontinuitas penyediaan bahan baku (suistainability supplay of raw material).
Sulitnya benih andaliman berkecambah kemungkinan disebabkan adanya dormansi (Siregar, 2003; Siregar, 2010). Berdasar beberapa hasil penelitian, penyebab dormansi belum jelas, sehingga usaha mematahkannya masih memberi hasil daya kecambah yang rendah (Sirait, 1991; Tampubolon, 1998; Samosir, 2000; Siregar, 2010; Siregar 2013).
Dormansi benih bisa diakibatkan komponen penyusun benih yang bersifat fisik dan atau kimia, yakni kekerasan kulit benih dan adanya senyawa tertentu pada kulit benih (Morris et al., 2000; Briggs et al., 2005; Widyawati et al., 2009;
Okeyo et al., 2011). Pada biji beberapa jenis tumbuhan dapat ditemukan sekaligus dormansi kulit biji dan dormansi embrio (fisiologi) (Bewley dan Black, 1983; Hartmann et al., 2011; Zaman et al., 2011; Mirzaei et al., 2013; Shaik et al., 2014).
Dormansi kombinasi mungkin dialami oleh benih andaliman. Menurut Dianxiang dan Hartley (2008), kebanyakan species Zanthoxylum mempunyai kulit benih yang tebal dengan sklerenkim yang padat. Komponen volatil, berupa senyawa terpenoid pada buah andaliman (Wijaya, 2000; Wijaya et al., 2001;
Parhusip et al., 2003; Kristanty et al., 2012; Moektiwardoyo et al., 2014) dan genus Zanthoxylum lainnya (Uji, 2001), merupakan senyawa penghambat perkecambahan (Hess, 1975; Kolotelo, 2005; Sadia et al., 2015).
Pematahan dormansi secara tunggal, yakni skarifikasi (Sirait, 1991;
Siregar 2013), skarifikasi ataupun zat pengatur tumbuh (Tampubolon, 1998;
Samosir, 2000; Siregar, 2010) masih menghasilkan perkecambahan benih andaliman yang rendah. Kombinasi pematahan dormansi akibat dua atau lebih mekanisma terbukti mematahkan dormansi benih beberapa species, antara lain benih Zanthoxylum rhetsa (Purwaning, 2009), benih Aeluropus lagopoides (Zaman et al., 2011), benih Robinia pseudoacacia (Mirzaei et al., 2013), benih Cassia fistula (Soliman dan Abbas, 2013), benih Hypoxis hemerocallidea (Shaik et al., 2014). Jadi diduga dibutuhkan kombinasi dari metode pematahan dormansi untuk meningkatkan daya berkecambah benih andaliman. Dalam mendapatkan kombinasi pematahan dormansi yang tepat untuk meningkatkan daya berkecambah benih andaliman, perlu dipelajari mekanisme dormansinya.
Menurut Hartmann et al. (2011) salah satu faktor dalam yang memengaruhi perkecambahan adalah tingkat kemasakan biji. Jika buah dipanen terlalu cepat embrio belum cukup berkembang, benih cenderung kecil, ringan, kisut, tidak berkualitas, dan kemampuan hidup singkat. Jika benih dipertahankan pada tanaman induk setelah matang fisiologis, perubahan fisiologis pada biji dapat
menyebabkan pembentukan biji keras, mengalami kerusakan (deteriorasi) atau penuaan (ageing), bahkan mati di tanaman (Khatun et al., 2009). Jika panen terlalu lama, buah akan merekah, benih jatuh ke tanah, atau diambil dan dimakan oleh burung atau binatang lainnya (Hartmann et al., 2011). Pada beberapa penelitian telah terbukti bahwa tingkat kemasakan memengaruhi daya berkecambah benih (Purwaning, 2009; Suwarno, et al., 2014; Tresniawati et al., 2014; Daff-Alla dan Mustafa, 2015; Solikin dan Nurfadilah, 2017).
Daya kecambah yang rendah kemungkinan juga disebabkan sedikitnya biji andaliman yang mengandung embrio (Siregar, 2010; Siregar, 2013). Biji keriput atau tanpa embrio (biji kosong) dapat terbentuk disebabkan ketidakmampuan embrio mengumpulkan cadangan makanan, aborsi embrio (embrio mati pada saat perkembangan) dan pertumbuhan tabung sari tidak sempurna atau serbuk sari gagal berkecambah. (Sutopo, 1993). Menurut Bewley dan Black (1983) inhibitor dapat menurunkan laju pertumbuhan biji. Biji yang demikian diharapkan tersortir dalam penyiapan benih. Siregar (2010) membuktikan sortasi dengan air memberi daya berkecambah lebih besar dari tanpa sortasi, namun daya berkecambah yang diperoleh masih rendah. Masih diperlukan teknik sortasi lain untuk memperoleh daya berkecambah andaliman yang lebih besar.
Penelitian diperlukan untuk mempelajari fenologi benih, menentukan umur panen, menggali penyebab rendahnya daya kecambah, menggali penyebab dormansi, mencari teknik sortasi dan pematahan dormansi yang lebih tepat dalam meningkatkan daya kecambah benih andaliman untuk keperluan perbanyakan dan budidaya tanamannya.
1.2. Perumusan Masalah
Aspek fisiologi dan budidaya tanaman andaliman masih sangat terbatas diketahui, termasuk aspek perbanyakannya. Petani masih menggunakan bibit liar dalam perbanyakan tanaman ini, karena bijinya sulit berkecambah. Ini menjadi salah satu hambatan bagi kebanyakan petani untuk memperbanyaknya dan membudidayakannya. Daya kecambah yang rendah dan lamanya berkecambah kemungkinan disebabkan adanya dormansi. Dari beberapa hasil penelitian, penyebab dormansi benih andaliman belum jelas, sehingga usaha mematahkannya masih menunjukkan hasil yang rendah. Penelitian pematahan dormansi secara terpisah, yakni skarifikasi ataupun zat pengatur tumbuh, masih menghasilkan perkecambahan benih andaliman yang rendah dan umur berkecambah yang lama.
Daya kecambah yang rendah kemungkinan juga disebabkan biji keriput atau tanpa embrio (biji kosong) dan viabilitas biji rendah akibat belum matang ataupun mengalami kerusakan (deteriorasi) dan penuaan. Penelitian diperlukan untuk menggali penyebab dormansi dan rendahnya daya kecambah, menentukan umur panen, serta memperoleh teknik sortasi dan pematahan dormansi benih andaliman untuk meningkatkan daya kecambahnya.
1.3. Tujuan Penelitian
Tujuan umum penelitian adalah meningkatkan daya kecambah benih andaliman (Zanthoxylum acanthopodium DC.).
Tujuan khusus penelitian ini adalah sebagai berikut:
1. menggambarkan perkembangan benih andaliman hingga matang fisiologis dan menentukan umur matang fisiologis benih andaliman.
2. menentukan penyebab rendahnya daya kecambah benih andaliman
3. menentukan penyebab dormansi benih andaliman
4. mendapatkan teknik sortasi dan pematahan dormansi benih andaliman yang menghasilkan daya kecambah yang besar
1.4. Manfaat Penelitian
Hasil penelitian ini diharapkan: (1) memberikan informasi tentang pola perkembangan buah/biji andaliman, anatomi benih, perkecambahan benih andaliman, penyebab rendahnya dan lamanya perkecambahan benih andaliman (dormansi benih andaliman), (2) mengisi kesenjangan informasi mengenai aspek fisiologi dan agronomi tanaman andaliman yang sangat terbatas saat ini, (3) menemukan cara penyiapan dan pengecambahan benih andaliman yang berguna sebagai acuan mendapatkan teknik perbanyakan tanaman andaliman melalui benih, (4) menyediakan acuan bagi penelitian terkait andaliman.
BAB II. TINJAUAN PUSTAKA 2.1. Tanaman Andaliman dan Potensi Pemanfaatannya
Andaliman (Zanthoxylum acanthopodium DC.) adalah tanaman rempah liar di Sumatera Utara. Buahnya biasa digunakan dalam makanan tradisional suku Batak sebagai bumbu masakan ikan dan daging, serta berbagai jenis sambal. Buah andaliman dikenal dan digunakan di kalangan terbatas. Tanaman andaliman jarang dikenal. Bahkan kebanyakan orang Batak tidak mengenal tanaman ini, karena tanaman ini tidak dibudidayakan secara luas dan khusus, dan sementara ini hanya ditemui pada daerah tertentu saja di Propinsi Sumatera Utara.
Deskripsi tanaman andaliman seperti uraian berikut ini. Semak atau pohon kecil bercabang rendah, tegak, tinggi mencapai 5 m, menahun. Batang, cabang, dan ranting berduri. Daun tersebar, bertangkai, majemuk menyirip beranak daun gasal, panjang 5-20 cm dan lebar 3-15 cm, terdapat kelenjar minyak. Rakis bersayap, permukaan bagian atas, bagian bawah rakis, dan anak daun berduri, 3- 11 anak daun, berbentuk jorong hingga oblong, ujung meruncing, tepi bergerigi halus, paling ujung terbesar, anak daun berukuran panjang 1-7 cm, lebar 0.5-2.0 cm. Permukaan atas daun hijau berkilat dan permukaan bawah hijau atau pucat, daun muda permukaan atas hijau dan bawah hijau kemerahan. Bunga di ketiak daun, bunga majemuk berbatas, anak payung menggarpu majemuk, kecil-kecil;
dasar bunga rata atau bentuk kerucut; kelopak 5-7 bebas, warna kuning pucat;
berkelamin dua, benang sari 5-6 duduk pada dasar bunga, kepala sari kemerahan, putik 3-4, bakal buah apokarp, bakal buah menumpang. Buah kotak sejati atau kapsul, bulat, diameter 2-3 mm, muda hijau, tua merah; tiap buah satu biji, kulit keras, warna hitam berkilat (Siregar, 2003).
Buah andaliman mengandung senyawa aromatik dengan rasa pedas dan getir yang khas, serta hangat. Jika buahnya dimakan meninggalkan efek menggetarkan alat pengecap, menyebabkan lidah terasa kebal, dan dapat meningkatkan nafsu makan. Kandungan kimia dan/atau aktivitas biologisnya telah dimuat pada beberapa publikasi (Siahaan, 1991; Tarigan, 1999; Siahaan, 2000; Wijaya, 1999; Wijaya, 2000; Uji, 2001; Wijaya et al., 2001; Surbakti, 2002; Parhusip et al., 2003; Tensiska et al., 2003; Parhusip, 2004a; Parhusip, 2004b; Suryanto et al., 2004; Parhusip et al., 2005; Zaridah et al., 2006; Sabri , 2007; Katzer, 2012; Kristanty et al., 2012; Kristanty dan Suriawati, 2014).
Menurut Uji (2001) species Zanthoxylum dikenal dengan minyak esensialnya yang merupakan kelompok senyawa terpenoid. Berdasar penelitian Kristanty et al. (2012) serta Kristanty dan Suriawati (2014) senyawa yang diperoleh dari ekstrak buah andaliman adalah terpenoid. Wijaya et al. (2001) menemukan sebanyak 24 komponen volatil dengan mayoritas terdiri dari monoterpen teroksigenasi, antara lain limonene, citronellal, β-myrcene, 2-β- ocimene, linalool, β-citronellol, neral, geraniol, geranial, geranyl acetate, sesquiterpene. Namun masih ada komponen yang belum teridentifikasi.
Parhusip et al. (2003) menemukan beberapa komponen ekstrak andaliman, yakni limonena, sitronelol, geraniol, dan β-mirsen.
Ekstrak buah andaliman memiliki aktivitas fisiologi sebagai antimikrob yang potensial menghambat pertumbuhan beberapa mikroba perusak pangan (Tarigan, 1999; Siahaan, 2000; Surbakti, 2002; Parhusip et al., 2003; Parhusip, 2004a; Parhusip, 2004b; Parhusip, et al, 2005). Hal ini memberi peluang bagi andaliman sebagai bahan baku senyawa antimikrob bagi industri pangan.
Ekstrak buah andaliman juga mempunyai aktivitas antioksidan (Wijaya, 1999;
Parhusip et al., 2003; Tensiska et al., 2003; Suryanto et al., 2004; Kristanty et al., 2012). Menurut Suryanto et al. (2004), antioksidan dari buah andaliman dapat menangkap oksigen singlet, yang merupakan ancaman bagi keamanan pangan.
Hal ini juga memberi peluang bagi andaliman sebagai bahan baku senyawa antioksidan bagi industri pangan, terutama produk pangan berlemak. Aktivitas antioksidan dan antimikrob memungkinkan andaliman digunakan sebagai bahan pengawet alami.
Selain aktivitas antioksidan, menurut Wijaya (1999) andaliman juga mempunyai efek imunostimulan. Graham et al. (2000) menuliskan bahwa buah andaliman bersifat antitumor. Ekstrak buah andaliman telah terbukti potensial memiliki aktivitas sitotoksik terhadap sel kanker (antikanker) (Kristanty dan Suriawati, 2014). Menurut Uji (2001) species Zanthoxylum lainnya, yakni Z.
nitidium, mempunyai aktivitas antitumor. Menurut Vickery dan Vickery (1981), kelompok monoterpene merupakan senyawa volatil yang dapat digunakan antara lain sebagai bahan industri farmasi.
Aktivitas antimikrob juga memberi peluang bagi andaliman untuk digunakan sebagai bahan baku pestisida nabati. Berdasarkan penelitian Surbakti (2002) disarankan perlunya kajian pemanfaatan andaliman untuk mengatasi jamur atau bakteri dari genus seperti Penicillium, Aspergillus, Fusarium, Pseudomonas, yang cukup banyak menjadi penyebab penyakit-penyakit penting berbagai tanaman. Minyak pati dari batang andaliman bahkan dapat membunuh larva Aedes aegypti (Zaridah et al., 2006). Sabri (2007) telah membuktikan buah andaliman mempunyai efek fetotoksik terhadap mencit (antifertilitas). Anggota
Zanthoxylum lainnya, yakni Z. armatum dan Z. rhetsa, juga dapat membunuh atau menolak serangga, pada stadium larva dan dewasa (Uji, 2001). Menurut Vickery dan Vickery (1981) sejumlah metabolit sekunder merupakan fungsisida atau antibiotik yang bisa melindungi tanaman dari serangan fungi atau bakteri, ataupun toksik terhadap hewan atau serangga.
Saat ini andaliman diperhitungkan menjadi sumber senyawa aromatik dan minyak esensial, yang mempunyai aktivitas antimikrob (antijamur, antibakteri), penolak dan membunuh serangga, antioksidan, dan sitotoksik/antitumor. Siregar (2016) telah menuliskan perlunya penggalian dan pengembangan potensi andaliman sebagai pestisida nabati. Berbagai anggota genus Zanthoxylum juga telah terbukti sebagai genus yang bernilai karena penemuan dan penggunaannya sebagai bahan obat dan produk bahan kimia alami, sehingga berpotensi sebagai sumber berbagai senyawa metabolit sekunder. Potensi pengembangan genus ini terus berkembang, terutama untuk pengembangan antiparasit, antitumor, dan antimikrob yang baru (Patiño et al., 2012). Berdasar kandungan kimia dan aktivitas biologisnya, pemanfaatan andaliman dapat ditingkatkan tidak lagi sekedar bumbu masakan tradisional, namun juga bahan pengawet, bahan obat dan suplemen, serta pestisida nabati. Dengan penggalian dan pengembangan potensinya, andaliman dijual tidak lagi hanya sebagai komoditas primer, namun juga berpeluang sebagai produk yang mendapat nilai tambah (Siregar, 2012).
Berkenaan dengan itu beberapa aspek perlu dikaji lebih lanjut, antara lain:
botani/ekologi, keragaman plasma nutfah, budidaya (termasuk perbanyakan), pasca panen, potensi pemanfaatan dalam industri pangan, farmasi, dan pestisida nabati.
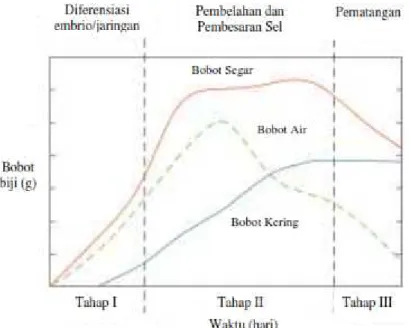
2.2. Pembentukan Biji dan Perkecambahan
Menurut Hartmann, et al. (2011) ada 3 tahap perkembangan fisiologis kebanyakan biji, yakni diferensiasi embrio (histodifferentiation), pembelahan dan pembesaran sel (cell expansion=food reserve deposits), dan pemasakan (maturation drying) (Gambar 1). Tahap I ditandai dengan diferensiasi embrio dan endosperma, yang sebagian besar disebabkan oleh pembelahan sel. Pada tahap ini, embrio mencapai awal tahap perkembangan kotiledon. Ada peningkatan cepat dalam bobot segar dan kering benih. Tahap II adalah periode pembesaran sel cepat, disebut juga pengisian benih, karena terjadi akumulasi cadangan makanan.
Tahap ini, merupakan periode aktif dengan peningkatan sintesis DNA, RNA, dan protein dalam biji yang besar. Cadangan makanan utama termasuk karbohidrat (pati), protein, dan lipid (minyak atau lemak). Pada akhir Tahap II perkembangan, biji telah mencapai matang fisiologis (juga disebut matang panen).
Matang fisiologis adalah waktu sebelum matang pengeringan, ketika benih telah mencapai berat kering maksimum melalui akumulasi cadangan makanan. Biji pada saat matang fisiologis dapat dilepas dari buah dan menunjukkan potensi perkecambahan tinggi yang diukur dengan viabilitas dan vigor benih.
Proses perkecambahan (mulai dari pecahnya kulit biji hingga munculnya radikula) meliputi tahapan berikut: (1) Penyerapan air, (2) Pengambilan O2, (3) Hidrolisis cadangan makanan, (4) Sintesis jaringan baru. Perkecambahan terdiri atas tiga fase, yakni fase I, imbibisi dicirikan dengan peningkatan penyerapan air;
fase II, aktivitas metabolisma yang tinggi, meliputi pematangan mitokondria, sintesis protein, metabolisma cadangan makanan; sintesa enzim spesifik (termasuk untuk melunakkan dinding sel embrio dan sel sekitar embrio); fase III,
keluarnya tonjolan radikula yang diakhiri dengan munculnya kecambah (Bewley, 1997; Hartmann et al., 2011).
Gambar 1. Tahap perkembangan fisiologis biji (Hartmann, et al., 2011)
Menurut Gardner et al. (1991), tahap-tahap proses perkecambahan sebagai berikut:
1. Perkecambahan diawali dengan penyerapan air dari lingkungan sekitar biji, baik tanah, udara, maupun media lainnya (tahap imbibisi). Efek yang terjadi adalah melunaknya kulit biji
2. Selanjutnya terjadi hidrasi jaringan dan absorbsi oksigen.
3. Kehadiran air di dalam sel mengaktifkan sejumlah enzim perkecambahan awal. Fitohormon asam absisat menurun kadarnya, sementara giberelin meningkat. Giberelin menyebabkan terbentuknya enzim.
4. Protein, pati, dan lipid dirombak oleh enzim-enzim hidrolisis (pengaktifan enzim). Molekul yang terhidrolisis ditranspor ke sumbu embrio, keluarnya tonjolan radikula yang diakhiri dengan munculnya kecambah (Bewley, 1997; Hartmann et al., 2011).
Gambar 1. Tahap perkembangan fisiologis biji (Hartmann, et al., 2011)
Menurut Gardner et al. (1991), tahap-tahap proses perkecambahan sebagai berikut:
1. Perkecambahan diawali dengan penyerapan air dari lingkungan sekitar biji, baik tanah, udara, maupun media lainnya (tahap imbibisi). Efek yang terjadi adalah melunaknya kulit biji
2. Selanjutnya terjadi hidrasi jaringan dan absorbsi oksigen.
3. Kehadiran air di dalam sel mengaktifkan sejumlah enzim perkecambahan awal. Fitohormon asam absisat menurun kadarnya, sementara giberelin meningkat. Giberelin menyebabkan terbentuknya enzim.
4. Protein, pati, dan lipid dirombak oleh enzim-enzim hidrolisis (pengaktifan enzim). Molekul yang terhidrolisis ditranspor ke sumbu embrio, keluarnya tonjolan radikula yang diakhiri dengan munculnya kecambah (Bewley, 1997; Hartmann et al., 2011).
Gambar 1. Tahap perkembangan fisiologis biji (Hartmann, et al., 2011)
Menurut Gardner et al. (1991), tahap-tahap proses perkecambahan sebagai berikut:
1. Perkecambahan diawali dengan penyerapan air dari lingkungan sekitar biji, baik tanah, udara, maupun media lainnya (tahap imbibisi). Efek yang terjadi adalah melunaknya kulit biji
2. Selanjutnya terjadi hidrasi jaringan dan absorbsi oksigen.
3. Kehadiran air di dalam sel mengaktifkan sejumlah enzim perkecambahan awal. Fitohormon asam absisat menurun kadarnya, sementara giberelin meningkat. Giberelin menyebabkan terbentuknya enzim.
4. Protein, pati, dan lipid dirombak oleh enzim-enzim hidrolisis (pengaktifan enzim). Molekul yang terhidrolisis ditranspor ke sumbu embrio,
titik tumbuh dan sebagai bahan bakar respirasi. Sitokinin dan auksin terbentuk dalam proses ini.
5. Pembelahan sel di bagian yang aktif (mitosis), seperti di bagian ujung radikula, diikuti oleh pembesaran sel.
6. Ukuran radikula (embrio) makin besar, sehingga kulit biji terdesak dari dalam, yang pada akhirnya pecah.
7. Titik tumbuh pada pucuk menuju ke atas permukaan tanah, kecambah (semai) muncul
Syarat supaya terjadi perkecambahan adalah viabel, lepas dormansi, lingkungan yang sesuai, tidak ada inhibitor. Pengapungan merupakan metode yang umum digunakan untuk memisahkan benih yang viabel dan tidak viabel (Hartmann et al., 2011). Analisis sinar X dapat digunakan sebagai cara cepat menguji kesegaran benih. Sinar X bukan merupakan cara normal untuk mengukur viabilitas benih, namun menyediakan pemeriksaan struktur dalam akibat gangguan mekanis, ketiadaan jaringan vital seperti embrio atau endosperm, infestasi serangga, rusak atau pecahnya kulit benih, dan penyusutan jaringan dalam. Benih dengan ukuran kurang dari 2 mm terlalu kecil untuk melihat detailnya dengan metode sinar X (Hartmann et al., 2011).
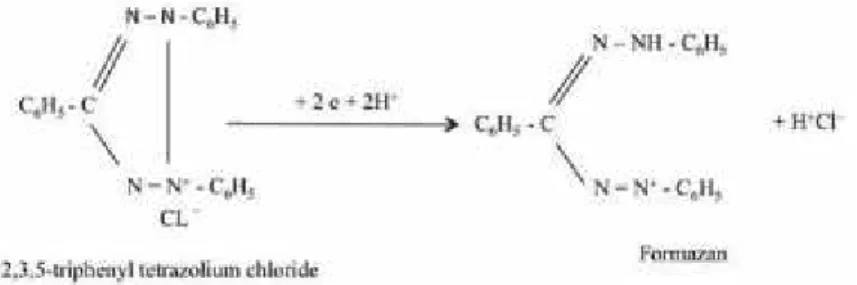
Pengujian viabilitas dapat dilakukan dengan test Tetrazolium (Budiarti et al., 2011; Elias et al., 2012; Kusumawardana et al., 2018). Elias et al. (2012) menuliskan bahwa tes tetrazolium didasarkan pada adanya aktivitas dehidrogenase dalam jaringan benih yang viabel selama proses respirasi. Enzim- enzim ini mengkatalisasi reaksi yang melepaskan ion hidrogen yang bereaksi untuk mereduksi (proses penerimaan ion H+) larutan 2,3,5-triphenyl tetrazolium
chloride yang tidak berwarna dan mengubahnya menjadi berwarna merah yang dikenal sebagai formazan (Gambar 2).
Gambar 2. Reaksi kimia yang terlibat dalam perubahan tetrazolium chloride yang tidak berwarna menjadi formazan berwarna merah (Elias et al., 2012)
Perkecambahan biji dipengaruhi oleh faktor luar dan dalam. Menurut Hartmann et al. (2011) faktor dalam yang memengaruhi perkecambahan biji adalah genotif, tingkat kemasakan biji, ukuran biji, dan ada tidaknya dormansi.
Menurut Hartmann et al. (2011) benih siap untuk dipanen apabila diambil dari tanaman tanpa merusak perkecambahan dan vigor benih, yang disebut matang panen. Pada banyak kasus, keseimbangan antara panen terlambat dan panen dini diperlukan untuk memperoleh jumlah maksimum benih berkualitas tinggi. Biji yang dipanen sebelum tingkat kemasakan fisiologisnya tercapai, viabilitasnya rendah, bahkan beberapa jenis tidak dapat berkecambah. Pada keadaan tersebut biji belum memiliki cadangan makanan yang cukup dan pembentukan embrio belum sempurna. Jika buah dipanen terlalu cepat embrio belum cukup berkembang, benih cenderung kecil, ringan, kisut (berkerut), tidak berkualitas, dan kemampuan hidup singkat. Sebaliknya, jika panen terlalu lama, buah akan merekah, jatuh ke tanah, atau dimakan dan diambil oleh burung atau binatang lainnya. Menurut Khatun et al. (2009) jika benih dipertahankan pada
Tidak berwarna merah
chloride yang tidak berwarna dan mengubahnya menjadi berwarna merah yang dikenal sebagai formazan (Gambar 2).
Gambar 2. Reaksi kimia yang terlibat dalam perubahan tetrazolium chloride yang tidak berwarna menjadi formazan berwarna merah (Elias et al., 2012)
Perkecambahan biji dipengaruhi oleh faktor luar dan dalam. Menurut Hartmann et al. (2011) faktor dalam yang memengaruhi perkecambahan biji adalah genotif, tingkat kemasakan biji, ukuran biji, dan ada tidaknya dormansi.
Menurut Hartmann et al. (2011) benih siap untuk dipanen apabila diambil dari tanaman tanpa merusak perkecambahan dan vigor benih, yang disebut matang panen. Pada banyak kasus, keseimbangan antara panen terlambat dan panen dini diperlukan untuk memperoleh jumlah maksimum benih berkualitas tinggi. Biji yang dipanen sebelum tingkat kemasakan fisiologisnya tercapai, viabilitasnya rendah, bahkan beberapa jenis tidak dapat berkecambah. Pada keadaan tersebut biji belum memiliki cadangan makanan yang cukup dan pembentukan embrio belum sempurna. Jika buah dipanen terlalu cepat embrio belum cukup berkembang, benih cenderung kecil, ringan, kisut (berkerut), tidak berkualitas, dan kemampuan hidup singkat. Sebaliknya, jika panen terlalu lama, buah akan merekah, jatuh ke tanah, atau dimakan dan diambil oleh burung atau binatang lainnya. Menurut Khatun et al. (2009) jika benih dipertahankan pada
Tidak berwarna merah
chloride yang tidak berwarna dan mengubahnya menjadi berwarna merah yang dikenal sebagai formazan (Gambar 2).
Gambar 2. Reaksi kimia yang terlibat dalam perubahan tetrazolium chloride yang tidak berwarna menjadi formazan berwarna merah (Elias et al., 2012)
Perkecambahan biji dipengaruhi oleh faktor luar dan dalam. Menurut Hartmann et al. (2011) faktor dalam yang memengaruhi perkecambahan biji adalah genotif, tingkat kemasakan biji, ukuran biji, dan ada tidaknya dormansi.
Menurut Hartmann et al. (2011) benih siap untuk dipanen apabila diambil dari tanaman tanpa merusak perkecambahan dan vigor benih, yang disebut matang panen. Pada banyak kasus, keseimbangan antara panen terlambat dan panen dini diperlukan untuk memperoleh jumlah maksimum benih berkualitas tinggi. Biji yang dipanen sebelum tingkat kemasakan fisiologisnya tercapai, viabilitasnya rendah, bahkan beberapa jenis tidak dapat berkecambah. Pada keadaan tersebut biji belum memiliki cadangan makanan yang cukup dan pembentukan embrio belum sempurna. Jika buah dipanen terlalu cepat embrio belum cukup berkembang, benih cenderung kecil, ringan, kisut (berkerut), tidak berkualitas, dan kemampuan hidup singkat. Sebaliknya, jika panen terlalu lama, buah akan merekah, jatuh ke tanah, atau dimakan dan diambil oleh burung atau binatang lainnya. Menurut Khatun et al. (2009) jika benih dipertahankan pada
Tidak berwarna merah
tanaman induk setelah matang fisiologis, perubahan fisiologis pada biji dapat menyebabkan pembentukan biji keras, mengalami kerusakan (deteriorasi) atau penuaan (ageing), bahkan biji mati di tanaman. Panen benih dini bisa juga diperlukan beberapa benih species kayu-kayuan yang memproduksi kulit biji keras yang dapat menambah dormansi embrio. Jika benih menjadi kering dan kulit biji keras, biji tidak dapat berkecambah hingga waktu yang lama (musim semi kedua). Widyawati et al. (2009) membuktikan bahwa semakin tua benih aren permeabilitasnya terhadap air semakin menurun, sehingga imbibisi berlangsung lebih lama, antara lain disebabkan oleh meningkatnya kandungan lignin dan tanin yang menutupi sel-sel sklereid kulit benih.
Tingkat kemasakan biji akan memengaruhi daya berkecambah benih.
Suwarno et al. (2014) memperoleh benih kemangi mencapai masak fisiologi pada umur 44-49 hari setelah berbunga dengan ciri-ciri buah berwarna cokelat, benih berwarna hitam dengan daya berkecambah 34%. Tresniawati et al. (2014) memperoleh benih kemiri Sunan (Reutalis trisperma) mencapai masak fisiologis 28 minggu setelah berbunga dengan kriteria warna buah hijau kecokelatan, kulit buah lunak, kulit biji berwarna cokelat dengan daya berkecambah 76-80%.
Okeyo et al. (2011) memperoleh daya kecambah benih Zanthoxylum gilletii yang sama antara benih dari buah hijau dewasa (mature green) dan buah merah yang masak (ripe). Sedangkan Purwaning (2009) mendapatkan masak fisiologis benih panggal buaya (Zanthoxylum rhetsa) dicapai pada tingkat kemasakan 4 yang dicirikan dengan wama eksokarp merah merata, ukuran buah dan benih terkecil (diameter buah 7,5 mm, panjang benih 5,4 mm, lebar benih 5,1 mm dan
tebal benih 4,1 mm), kadar air benih turun hingga 20,7%, dengan bobot kering benih mencapai nilai tertinggi (83,9%).
Biji berat (besar) dan biji masak lebih unggul dibanding biji ringan (kecil) dan belum masak. Kelemahan-kelemahan yang terdapat pada biji belum masak juga terdapat pada biji kecil (Justice dan Bass, 1994).
Faktor luar (lingkungan) yang memengaruhi perkecambahan adalah air, O2, temperatur, cahaya, medium.
2.3. Dormansi Benih
Umur biji berkecambah berbeda antar berbagai tumbuhan. Biji tumbuhan tertentu memerlukan masa istirahat (dormansi), baru berkecambah. Biji dorman adalah biji yang gagal berkecambah pada suatu lingkungan yang mendukung perkecambahan biji species lain yang tidak dorman. Biji-biji demikian akan berkecambah dalam waktu yang relatif lebih lama. Jadi dormansi biji adalah suatu keadaaan pertumbuhan yang tertunda atau keadaan istirahat, walaupun berada dalam keadaan yang menguntungkan untuk perkecambahan.
Penyebab dormansi biji adalah dormansi kulit biji dan dormansi embrio.
Hartmann et al. (2011) membagi mekanisma dormansi sebagai berikut:
I. Dormansi primer, kondisi dormansi pada akhir perkembangan benih, terdiri atas:
a. Dormansi eksogen, ditentukan oleh faktor-faktor di luar embrio i. Fisik : Kulit benih yang impermeabel
ii. Kimia : inhibitor pada bagian kulit atau pembungkus benih b. Dormansi endogen, ditentukan oleh faktor dalam embrio
i. Fisiologi, faktor dalam embrio menghambat perkecambahan
1. Nondeep, potensi pertumbuhan embrio tidak cukup untuk keluar dari kulit benih. Dapat sensitif pada cahaya.
2. Intermediate, potensi pertumbuhan embrio tidak cukup kuat untuk keluar dari kulit benih. Embrio dapat berkecambah jika dipisah dari kulit benih.
3. Deep, embrio tidak berkecambah ketika dilepas dari kulit benih atau akan tumbuh kerdil
ii. Morfologi, embrio tidak berkembang penuh pada saat biji dilepas dari tanaman
iii. Morfofisiologi, kombinasi embrio yang tidak berkembang dan dormansi fisiologi embrio
c. Dormansi kombinasi, kombinasi dormansi eksogen dan endogen, misal fisik (kulit benih yang keras) dan dormansi fisiologi
II. Dormansi sekunder
a. Thermodormancy, setelah dormansi primer dilepaskan,temperatur yang tinggi merangsang terjadinya dormansi
b. Conditional dormancy, perubahan kemampuan berkecambah sehubungan dengan waktu dalam setahun
Bewley dan Black (1983) menuliskan bahwa kulit benih adalah struktur penting sebagai suatu pelindung antara embrio dan lingkungan di luar benih.
Kulit benih dapat menyebabkan dormansi karena kulit biji menghalangi penyerapan air dan gas, mengganggu pertukaran gas, mengandung inhibitor, menghalangi keluarnya inhibitor yang berasal dari embrio, memodifikasi cahaya
yang mencapai embrio, dan menghalangi pertumbuhan embrio (pengekangan secara mekanis).
Morris et al. (2000) menguraikan bahwa dormansi yang disebabkan oleh kulit benih dapat terjadi karena adanya komponen penyusun benih baik yang bersifat fisik dan atau kimia. Secara fisik, benih aren termasuk benih keras baik pada bagian kulit maupun endospermanya (Rofik dan Murniati, 2008).
Peningkatan kadar lignin dan tanin sangat berperan menutupi sel-sel sklereid kulit benih yang menurunkan permeabilitas benih aren terhadap air (Widyawati et al., 2009). Briggs et al. (2005) telah mengidentifikasi beberapa mekanisma potensial pada dormansi kulit benih species Grevillea. Embrio secara sempurna dikelilingi oleh barier luar dan dalam bagi difusi senyawa berbobot molekul tinggi. Senyawa fenolik hadir dalam eksotesta yang dapat menghalangi pertukaran gas. Lapisan sklerenkim, secara bersama memperkuat sel-sel dalam endotesta dan eksotesta, berperan sebagai penghalang mekanis. Siddiqui dan Khan (2010) menemukan keberadaan fenolik pada kulit benih Halopyrum mucronatum memengaruhi imbibisi air di awal dan sintesis protein awal selama proses perkecambahan.
Menurut Bewley dan Black (1983) dormansi embrio disebabkan adanya inhibitor pada embrio (termasuk kotiledon), belum matangnya embrio (embrio maturity), dan hambatan metabolis pada embrio. Senyawa fenolik yang terdapat pada benih Palicourea rigida (Rubiaceae) menyebabkan dormansi benih (Inácio et al., 2013). Kolotelo (2005) menuliskan bahwa resin, yang merupakan senyawa terpenoid yang didominasi oleh limonene, pada benih genus dari famili Pinaceae dapat menghambat perkecambahannya. Senyawa limonen terbukti berpotensi