L
Journal of Experimental Marine Biology and Ecology 246 (2000) 31–52
www.elsevier.nl / locate / jembe
The effects of symbiotic crabs on the pumping activity and
growth rates of Chaetopterus variopedatus
1
*
Michael W. Grove , Christopher M. Finelli , David S. Wethey, Sarah A. Woodin
Department of Biological Sciences, University of South Carolina, Columbia, SC 29208, USA Received 23 June 1999; received in revised form 18 November 1999; accepted 25 November 1999
Abstract
This study investigates how the presence of symbiotic crabs (Pinnixa chaetopterana or
Polyonyx gibbesi ) in the tubes of the polychaete Chaetopterus variopedatus affects the worms’
pumping activity and growth rates under laboratory and field conditions. In the field, worms whose tubes are inhabited by Pinnixa beat their fan segments significantly more frequently than do worms hosting Polyonyx, but other measures of pumping activity do not differ according to symbiont species. In the lab, worms tend to move water through their tubes at higher rates when crabs are present. In 7-month laboratory experiments, growth rates of worms hosting either species of crab did not differ from growth rates of worms without crab symbionts. Although worms hosting Polyonyx are, on average, significantly larger than worms hosting Pinnixa, this appears to be due to competition between the crab species for hosts and not due to differential effects on host growth. Unlike the crabs in this study, pea crab species inhabiting bivalves are known to have strong deleterious effects on host growth and reproduction, suggesting that the evolution of virulence in symbiotic interactions is dependent upon specific ecological context. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Benthic; Pinnotherid; Polychaete; Symbiosis
1. Introduction
Symbiotic interactions are ubiquitous in marine habitats, and at least two of these
*Corresponding author. Academy of Natural Sciences Estuarine Research Center, 10545 Mackall Road St. Leonard, MD 20685, USA.
E-mail address: [email protected] (M.W. Grove)
1
Louisiana Universities Marine Consortium, Chauvin, LA 70344, USA.
interactions, coral–algal and fish–anemone symbioses, have been the focus of intensive research programs. The usefulness of such systems in studying evolutionary patterns in symbiosis is somewhat limited by the fact that the symbiotes only interact with a narrow range of host life histories. Conversely, species in the decapod family Pinnotheridae form symbiotic relationships with a vast range of marine organisms that have very divergent life history patterns. These pea crabs are found ectosymbiotically on sea urchins and sand dollars (Bell, 1984) and in the tubes of polychaete worms (Gray, 1961; McDermott, 1962), echiurans (Boesch, 1971), and burrowing shrimp (Pearse et al., 1942), as well as endosymbiotically in the gill chambers of bivalves (Stauber, 1945; Pearce, 1966), branchial chambers of tunicates (Wass, 1955), and anal cavities of sea cucumbers (Wells and Wells, 1961; Takeda et al., 1997). Given the range of host morphologies and life histories displayed in these relationships, and the relative ease with which host and symbiont growth and reproductive patterns may be studied, this family is well suited for the study of evolutionary patterns in symbiosis. By correlating the pattern of radiation among hosts with changes in the ecological effects of the crabs on host growth, reproduction, and population dynamics, a fuller understanding of the nature of evolution of symbiotic relationships can be gained.
Such an analysis is, however, not possible at this point in time due to a profound lack of knowledge regarding the basic ecology and genetics of pea crab species. The most widely studied species have been those which inhabit the gill chambers of bivalve molluscs. Presence of the crabs significantly reduces growth and reproductive output in several species of host bivalves (Haven, 1959; Pearce, 1966; Kruczynski, 1972; Anderson, 1975; Lalitha Devi, 1986; Bierbaum and Ferson, 1986; O’Beirn and Walker, 1999) and also results in distortion of shell shape (Bierbaum and Ferson, 1986). The crabs obtain nutrition by scraping mucous food strings off the gills of the host (Orton, 1920; Stauber, 1945; Kruczynski, 1972), resulting in the formation of lesions on the gills (Christensen and McDermott, 1958; Haven, 1958; McDermott, 1962). Fibrous masses may also be formed on other soft body tissues due to the activity of the crabs (Stauber, 1945; Dix, 1973; Jones, 1977). This damage results in lowered water filtration rates (Prezenger, 1979; Bierbaum and Shumway, 1988) and lowered oxygen consumption (Bierbaum and Shumway, 1988) by the host, putting it at a metabolic disadvantage when compared to uninfested hosts.
The effects of pea crabs on host growth, reproduction, or behavior have been little studied for non-bivalve hosts. Telford (1982) presents evidence that crabs in the genus
Dissodactylus may break off and ingest parts of the spines or tube feet of host sand
dollars, while Bell (1984) has observed the crabs moving their chelipeds from host food grooves to their mouths, apparently ingesting mucous food strands. The quantitative effects of these activities on host growth, however, remain unexplored. Similarly, Wells and Wells (1961) described the relationship of Pinnaxodes species with holothurians as being commensal based on the normal location of the crabs in the cloaca or respiratory tree of the hosts, but Takeda et al. (1997) suggest that Pinnixa tumida reduce the growth of their host holothurian Paracaudina chilensis as a product of their collection of mucous secreted by the host.
between these crabs and their hosts is quite different in nature from that which occurs between bivalves and their pinnotherid species. Chaetopterus constructs a U-shaped tube made of fibrous parchment-like material with the two ends of the tube exposed for several centimeters above the sediment surface. The ends of the tube taper to constricted openings which have diameters only 19% the diameter of the main body of the tube (Brown, 1977). The worm forces water through the tube by movement of three highly-modified fan-shaped body segments, and feeding is accomplished by filtering the water through a mucous bag which is anchored anteriorly by the two wing-shaped aliform notopodia and posteriorly by a dorsal cupule. At the conclusion of the feeding period, the mucous bag is rolled into a ball by the cupule, passed forward to the mouth by ciliary action, and ingested (Macgintie, 1939).
Previous observations of the pumping and feeding activity of Chaetopterus have been made under laboratory conditions on worms housed in artificial glass tubes without crab
˚
symbiotes present (Brown, 1977; Riisgard, 1989), but in intertidal populations in South Carolina, up to 95% of Chaetopterus tubes may hold a pair of crabs of Pinnixa
chaetopterana or of the anomuran crab Polyonyx gibbesi Haig (Grove and Woodin,
1996). Members of the two crab species are rarely found occupying the same host tube simultaneously (only 1.6% of 184 tubes examined: Grove and Woodin, 1996). Similar results have been reported for Chaetopterus populations in Massachusetts, North Carolina, and Florida (Enders, 1905; Gray, 1961).
The presence of these crabs may have several effects on the pumping activities of the host worm. Passively, the crabs will present obstructions to flow through the tubes and will create an additional pressure drop which must be overcome by the worm. On a more active basis, instead of ‘stealing’ food strands directly from the host, the crabs filter the water pumped by the worm for their food using seteose mouthparts (Caine, 1975). By reducing the food available to the worm, or by presenting an increased oxygen demand, the crabs may directly influence the metabolism and growth of the host. Since Polyonyx is a significantly larger crab than Pinnixa, at least in South Carolina populations, (Grove and Woodin, 1996), it might be expected to negatively affect the host more in both the active and passive senses. We have previously shown (Grove and Woodin, 1996), however, that worms inhabited by Polyonyx are significantly greater in weight than are worms inhabited by Pinnixa. It is uncertain whether this difference results from differential effects of the crabs on growth of the host or if Pinnixa recruits into small
Chaetopterus and is subsequently competitively replaced by Polyonyx.
Evidence from the life-histories of different crab species suggests that Pinnixa and
Polyonyx do not cause direct harm to the host. Pea crab species infesting bivalves have
greatly increased fecundities as compared to free-living crab species (Hines, 1992); such increases are often characteristic of parasitic species (Poulin, 1995a,b). Pea crabs occupying bivalve molluscs also are often reported to have lower host occupation rates than the species examined in this study (as low as 5% of hosts occupied: Stauber, 1945; Haven, 1959; O’Beirn and Walker, 1999). Conversely, Pinnixa chaetopterana and
Polyonyx gibbesi put only about 10% of their dry body weight into each clutch (Grove
tubes may not be parasitic. These pea crabs thus may represent the other end of the evolutionary continuum from those in bivalves.
A major focus of this study was therefore the effect of Pinnixa and Polyonyx on the performance of the worm hosts. Specifically, we address three aspects of the relationship between Chaetopterus and its crab symbionts. Firstly, do the two crab species cause differential pumping activity patterns (as measured by activity duration, volume flow-rates, or work done) in host worms under field conditions? Secondly, does the presence of either crab species cause individual worms to alter their activity pattern from what would be the case without crabs present? This question may be addressed in a laboratory setting by monitoring the initial activity of worms with or without crabs in their tubes and then measuring subsequent activity after adding or removing crabs to the same individuals. Field observations of worm activity in the absence of crab symbionts is made impractical by the high occupation rates (95%) of worm tubes by crabs in natural populations. Finally, does the presence of crabs result in long-term differences in the growth rates of Chaetopterus hosts? Answers to these questions will both address the ecological patterns observed in the sizes of Chaetopterus hosts and provide a test of patterns of evolution in symbiotic interactions.
2. Materials and methods
2.1. Background
Activity patterns of Chaetopterus have been studied in detail in the laboratory (Wells and Dales, 1951; Brown, 1977; Brown and Rosen, 1978). In the absence of chemical or mechanical stimulants to feeding, the worms show stereotyped ‘feeding’ activity periods with lengths of approximately 17 or 18 min (Macgintie, 1939; Wells and Dales, 1951), but when feeding stimulants such as phytoplankton are present, the length of these periods is significantly reduced (Sumida and Case, 1983). Estimates of volume
flow-21
rates during active pumping range from 0.13 to 0.54 ml s (Wells and Dales, 1951;
˚
Dales, 1969; Brown, 1977; Riisgard, 1989).
2.2. Growth rate experiments
Experimental animals were housed in fiberglass containers (25 cm high335 cm wide356 cm long) in an unfiltered flow-through seawater system at the Baruch Marine Laboratory in Georgetown, SC. In order to keep the worm tubes upright, the tubes were sewn with nylon thread to fiberglass window screening which had been attached to the walls of the container with hot-melt glue. The window screening was then cut to match the shape of the attached tube so as not to impede water flow through the container.
21
Flow rates through the containers were approximately 15 l min although this fluctuated as the supply tubes became partly filled with sediment. The supply tubes were cleaned weekly to prevent flow-rate from dropping too low. At these times sediment was also removed from the containers housing the experimental animals if it had noticeably accumulated. Salinity of the seawater was generally 34‰ but dropped to lower levels following storms and subsequent freshwater runoff events. Water temperature at this site in 1995 ranged from an average daily high of 29.78C in June to an average daily low of 10.48C in December. Water temperatures in 1994 followed a similar pattern.
In 1994, 21 worms were collected and randomly assigned to three containers (with seven animals each) supplied from the same seawater line. This experiment was initiated on May 26th and ended on December 30th at which time the worms and crabs were removed from their tubes and final weights were obtained. In the first few days following the initiation of the experiment, crabs could be observed actively moving about the container housing the worms, but this activity appeared to cease following this time. Because of this movement, worms were assigned to treatments for analysis based on the crab species present at the end of the experimental period.
This experiment was repeated in 1995 using a total of 36 worms housed in four containers. This experiment began on April 18th and concluded on November 21st.
Although the initial wet weights of worms in these experiments were not significantly different between treatments (F50.96, MSE520.858, df52,36, p50.392) or across tanks (F51.53, MSE520.858, df56,36, p50.198), average growth varied widely across tanks. To reduce the influence of uncontrolled changes in water supply rates to the tanks on the results of the experiments, data from tanks in which control worms showed net weight losses were discarded before statistical analysis. This resulted in the removal of data from only one tank. Across the remaining tanks, ten out of the original 48 worms died during the course of the experiments.
The experiments were analyzed statistically using a two-way ANCOVA with holding tank (or block) and crab treatment as the main effects and initial worm weight as the covariate. All interaction terms were nonsignificant and were dropped from the final model. All statistical analyses were done using SAS, version 6.12 for PC (SAS Institute, Cary, NC).
2.3. Field activity measurements
not rise by more than 18C, during any 30-min recording period. In general, the activity periods of two worms were monitored during the falling tide and two during the rising tide of a given tidal cycle. Worms chosen for monitoring during falling tides were not exposed for more than 5 min before being enclosed, while worms monitored during rising tides had been exposed for greater than 30 min.
Pumping activity of the worms was monitored using an acoustic Doppler flow probe (probe: Iowa Doppler Products, Iowa City, IA; circuitry by Hopkins Marine Station, using a design by Iowa State University, Department of Biophysics). The face of the
2
probe, with an area of 1 mm , was oriented directly into the flow at the centerline of one of the tube openings of the worm before filling the enclosure with water. The face of the probe and part of the lead wire were enclosed in a 1-ml pipette tip in an attempt to streamline flow around the probe. The Doppler probe emits a 20-MHz signal in six cycle bursts with a pulse repetition frequency of 62.5 kHz. The Doppler shift frequency of pulses echoing off particles in the water column and returning to the probe is then subtracted from the initial frequency and sent from the circuitry as audio output. Doppler shift frequency is related to water flow velocity by the equation:
*
Velocity5(Df *c) / 2f coso a)
whereDf is the output Doppler shift frequency, c is the speed of sound in the medium
21
((1500 m s in seawater), f is the emitted frequency of 20 MHz, ando a is the angle
(here 08) between the direction of the sound beam and the velocity vector (Baker, 1970). The frequency of the output signal thus lies within the audible range of 40–10 000 Hz
21
for water flows of 0.15–36.5 cm s . Output was recorded onto Hi-8-mm videotape using a Sony CCD-TR8 video camera.
In the laboratory, these recordings were played back over audio speakers and the timings of worm pumping activities noted. Ten- to 15-s intervals from at least ten different activity periods for each worm were then digitized at 22.05 kHz onto computer disc using a Turtle Beach ‘Tahiti’ sound card for IBM PC and WAVE for Windows software (Turtle Beach Systems, York, PA). The resultant files were fast Fourier transformed in 4096 cycle (i.e. 0.186 s) segments using a program written inPERL. The
output provided a measure of the Doppler frequency distribution for each 0.186-s time interval. The five highest frequency peaks in each segment were then weighted by height and a mean flow velocity for the segment calculated using SAS. The speed of sound in the Doppler velocity calculations for each sample was assumed to be:
21 2 3
c (m s )51449.214.6T20.055T 10.00029T 1(1.3420.01T )(S235)
where T is water temperature (8C) and S is salinity (‰) (Jumars, 1993). The effect of water depth on sound speed was assumed to be negligible. Mean velocities were calculated for both short-duration activity periods (,30 s) (tube irrigation or cleaning events), and longer-duration periods (.60 s), which are feeding activity (Sumida and Case, 1983).
Fig. 1. Plot of the stroke periodicity (i.e. the time between the initiation of two successive strokes for any given fan segment) versus spectral density for a ten second activity period of Chaetopterus variopedatus in the field. This worm was host to Pinnixa. Stroke periodicity was estimated to be 2.148 s based upon the peak with the highest energy.
analysis (Fig. 1) displays peaks with periodicities of less than 1 s, which are assumed to be random noise components of the signal, and a single peak with periodicity between 1 and 7 s. When this periodicity is regressed against values obtained from direct observation of the activity of animals housed in glass tubing, a relationship of:
Estimated stroke frequency50.941*Observed stroke frequency
2
is obtained, with a r value of 0.93 ( p50.0001 n586).
Volume flow-rates were estimated for individual worms by multiplying the mean velocity for a given type of activity (i.e. long or short period) times the cross sectional area of the tube opening where recording occurred. A time-averaged rate was also calculated for each worm by multiplying the flow-rate for each activity type by the percentage of total activity comprised by that activity type. These products were then summed and multiplied by the percent of recording time that the worm was engaged in active pumping to yield a flow-rate averaged across both active and inactive periods.
˚
Flow through the tubes is assumed to be laminar (Riisgard, 1989), yielding an average flow velocity equal to one-half the maximum velocity occurring at the center of the tube opening (Vogel, 1981).
was used to examine the interiors of worm tubes in 1995 in order to determine the species of crab present. Wet weights of worms were estimated using the relationship:
Wet weight (g)51.037*(Average tube height in cm)
10.415*(Distance between tube openings in cm)
2
(r 50.43, p50.001, n562)
Worms monitored in 1996 were dug up after recordings were made and crab symbionts present and worm wet weights were observed directly.
Results for these experiments were analyzed using three-way ANCOVA with recording site, tidal exposure, and crab species present as the main effects. Recording site had no significant effect on any measure of worm activity, and therefore, no data are presented for this main effect. All interaction terms were nonsignificant and were dropped from the final models. Worm wet weight and water temperature (for all activity measures) and tube opening diameter (for flow velocity only) were tested as covariates. Water temperature was found to be a nonsignificant covariate for all activity measures and was dropped from the final models. Worm weight was marginally significant for several measures and was retained in all final models, while tube opening diameter was highly significant for long-period flow velocities and was retained for all water velocity models.
2.4. Laboratory activity measurements
Chaetopterus were collected and brought into the laboratory in February 1997 where
they were maintained in an 800-l recirculating seawater system at 34‰ salinity and 178C and fed ad libitum a mixture of Isochrysis galbana and Phaeodactylum tricornutum. After a 2-day acclimation period, the worms were removed from their tubes and placed in artificial tubes along with crabs appropriate for their assigned experimental treatment. These glass tubes were 30 cm in length, with interior diameters ranging from 1.55 to 2.1 cm, and tapered at one end to openings of approximately 4.5 mm diameter. After the worms had been weighed and placed into tubes which approximated the diameter of their natural tubes, the non-tapered ends of the artificial tubes were closed with corks which had 4.5-mm diameter holes drilled in their centers.
Worms were randomly assigned to one of four treatments: (1) worms with a mixed-sex pair of Pinnixa chaetopterana in their tubes during an initial round of activity recordings, which were subsequently removed before a second round of recordings were made; (2) worms with a mixed-sex pair of Polyonyx gibbesi in their tubes during the initial recording, which were removed before the second recordings; (3) worms which had no crabs in their tubes during the initial activity recordings, but subsequently received a mixed-sex pair of Pinnixa before the second round of recordings; and (4) worms which had no crabs in their tubes during the initial recordings, but subsequently received a mixed-sex pair of Polyonyx. After being transferred to glass tubes, worms were maintained in the seawater system for 18 days before the first round of activity recordings were made.
thin layer of secreted material, worms were placed singly into a 120-l aquarium filled with water drawn from the recirculating system, and the acoustic Doppler probe was positioned at the center of the opening at the tapered end of the tube. Worms were given a 15-min acclimation period after transfer (during which many individuals extended their bodies and began pumping activity) and the activity patterns of the worms were then videotaped for 30 min in indirect red light while the output of the Doppler probe was recorded onto the audio track of the tapes. After all worms had been videotaped, crabs were transferred between appropriate individuals and the worms were given another 18 day acclimation period before a second round of recordings were made.
The video portions of the tapes were subsequently examined in order to note the position of the worms in their tubes and the positions of the crabs relative to the body of the worms. The Doppler output of the recordings were analyzed in the same manner as the field recordings with two exceptions: (1) equal numbers of intervals were analyzed in which worms were pumping towards and away from the probe face; (2) Doppler output from a section of recording in which the worm was inactive was subtracted from active signal segments during Fourier transformation in order to remove the signature of background electrical noise (predominantly 60 Hz hum and in some recordings a constant 850-Hz signal of unknown origin). Stroke frequency of the fan segments during these activity periods was measured by direct visual observation of the video recordings. Activity changes within worms when hosting crabs versus when tubes were unoccupied were analyzed using paired t-tests. Potential activity differences caused by tube occupation by the two crab species were analyzed using two-way ANOVA with crab species and time of crab occupancy (i.e. whether crabs were present during the first or second round of recordings in a given worm tube) as the main effects. No worm was thus used more than once in this analysis. Since the data for this analysis were, however, used previously in the tests for within-worm activity changes, the alpha value for significance in this analysis was set to p50.025. Internal tube diameter and worm wet weights were not significant covariates and were dropped from the final model. All interaction terms were nonsignificant and were also dropped from the final model.
3. Results
3.1. Growth rate experiments
Analysis of the effects of the presence or absence of crabs on growth is complicated to some degree by the movement of crabs between host worms during the course of the experiment. Thirteen worms which had a pair of crabs in the tubes at the initiation of the experiments subsequently lost both members of the pair (four worms with Pinnixa, nine with Polyonyx). Six of these pairs recruited into worm tubes which had no crabs initially present (four Pinnixa, two Polyonyx).
Table 1
Average final wet weights of Chaetopterus variopedatus in laboratory growth rate experiments conducted in
a
1994 and 1995
Method of assignment Assigned treatment
to treatment No crabs Pinnixa Polyonyx
Use worms which lose crabs but
not worms which gain crabs (N533) 16.3 (1.0) 16.4 (0.8) 15.9 (0.8) Use worms which gain crabs but
not worms which lose crabs (N525) 16.3 (1.0) 17.0 (0.9) 17.2 (1.0) Use crab species present
at end of experiment (N531) 16.3 (0.7) 17.0 (0.8) 17.1 (0.9) Use only worms which have the
same crab species at initiation
and end of experiment (N517) 17.1 (1.1) 18.5 (1.2) 17.0 (1.1)
a
Means are least-squares means using initial wet weight as a covariate. Values in parentheses are one standard error. Means within any given row are not significantly different at p50.05.
top line assume that most losses of crabs are due to active movement of the crabs out of tubes early in the experiment with subsequent recruitment into new tubes occurring at unspecified but delayed times. Conversely, crab movement may be both immediate and direct in which case losses of crabs from the other tubes would be due to crab mortality at undetermined times; thus the use of worms which gain crabs but not worms which lose crabs in the analysis in the second line. This analysis requires dropping the results from one additional tank in order to maintain a complete block design.
If both crab movement and mortality take place early in the experiment, then the crab condition at the end of the experiment represents the condition experienced by the worms during most of the growth period; this is the analysis on the third line in Table 1. One additional incomplete block has been dropped from this analysis to maintain a complete design. The most conservative test is to use only worms which are in the same crab treatment at both the start and the end of the experiment. This necessitates dropping three tanks from the analysis to maintain a complete block design, resulting in a greatly reduced sample size. These tests all demonstrate that crab treatment has no significant effect on final worm weight (line 1: F50.10, MSE57.275, df52,24, p50.901; line 2:
F50.23, MSE57.275, df52,17, p50.799; line 3: F50.32, MSE56.214, df52,23,
p50.729; line 4: F50.47, MSE56.526, df52,10, p50.640, respectively).
These analyses all utilize initial worm weight as a covariate, which is highly significant in all cases. Analysis based on the percentage change in weight over the course of the experiment is unsatisfactory because this change was related to the initial weight of the worms by the regression equation:
2
% change in weight582.724.8*Initial weight (r 50.49, p50.001, n545)
initial weight was less than approximately 17.2 g. Thirty of the 45 surviving worms had initial weights of less than 17.2 g.
3.2. Field activity measurements
When compared using paired t-tests for individual worm means, mean water flow velocity (t52.37, p50.022, n557) and fan stroke rate (t52.73, p50.011, n549) were both significantly greater for short-duration activity periods than for long-duration periods. Therefore, short and long activities are analyzed separately.
Increasing time of tidal exposure resulted in a significant decrease in flow velocity of long activity periods (F53.34, MSE52.777, df52,44, p50.045) although volume flow-rates did not change (F50.82, MSE50.062, df52,45, p50.446) (Table 2). This decrease was accompanied by a similar, but not significant, decrease in the stroke rate for these periods (F51.00, MSE526.02, df52,36, p50.499). Short duration activity, however, showed no such trends and varied with no clear pattern. The percent of time spent actively pumping (F50.23, MSE50.046, df52,51, p50.796) percentage of activity comprised of feeding periods (F51.47, MSE50.059, df52,49, p50.239), and time-averaged volume flow-rates (F50.47, MSE50.024, df52,41, p50.627) also did not change significantly with increasing exposure times.
Flow velocities tended to be higher, although not significantly so, for worms which housed Polyonyx for both feeding activity (F50.14, MSE52.78, df51,44, p50.708) and irrigation activity (F50.43, MSE56.03, df51,44, p50.515) (Table 3). Active volume flow-rates, however, showed no trends. Polyonyx hosts had significantly lower
Table 2
Average activity measurements of field populations of Chaetopterus variopedatus exposed for different periods
a
Water flow velocity (cm s ) 5.95 (0.40) 4.73 (0.78) 4.55 (0.39)
21 21 A A A
Fan strokes min segment 18.8 (1.4) 18.2 (2.2) 16.1 (1.3)
21 A A A
Volume flow-rate (ml s ) 0.49 (0.06) 0.36 (0.12) 0.39 (0.06)
Short-duration activity periods
21 A A A
Water flow velocity (cm s ) 6.22 (0.59) 4.54 (1.07) 6.39 (0.58)
21 21 A A A
Fan strokes min segment 18.8 (2.3) 17.7 (3.1) 22.5 (2.1)
21 A A A
Volume flow-rate (ml s ) 0.51 (0.07) 0.37 (0.12) 0.50 (0.06)
Total activity measures
A A A
% Time pumping 58.3 (4.8) 64.3 (9.3) 57.3 (4.5)
A A A
% Feeding activity 60.7 (5.8) 70.4 (10.5) 52.0 (5.2)
Time-averaged volume
21 A A A
flow-rate (ml s ) 0.29 (0.04) 0.23 (0.07) 0.24 (0.04)
a
Table 3
Average activity measurements of field populations of Chaetopterus variopedatus inhabited by a pair of crabs
a
of Pinnixa chaetopterana or Polyonyx gibbesi
Crab species p-value
Pinnixa Polyonyx Long-duration activity periods
21
Water flow velocity (cm s ) 4.97 (0.45) 5.19 (0.40) 0.708
21 21
Fan strokes min segment 19.4 (1.5) 15.3 (1.2) 0.028
21
Volume flow-rate (ml s ) 0.42 (0.07) 0.41 (0.06) 0.911
Short-duration activity periods
21
Water flow velocity (cm s ) 5.42 (0.65) 6.02 (0.59) 0.515
21 21
Fan strokes min segment 21.6 (2.5) 17.1 (1.6) 0.178
21
Volume flow-rate (ml s ) 0.45 (0.07) 0.47 (0.06) 0.838
Total activity measures
% Time pumping 57.1 (5.5) 62.9 (4.8) 0.436
% Feeding activity 63.7 (6.2) 58.3 (5.6) 0.528
Time-averaged volume
21
flow-rate (ml s ) 0.26 (0.04) 0.25 (0.04) 0.932
a
Values in parentheses are one standard error. Means are least-squares means using tube opening diameter as the covariate for velocity estimates. Total sample sizes are 22 worms for Pinnixa and 35 worms for
Polyonyx. p-values are results of Tukey HSD tests.
fan stroke rates than did worms whose tubes housed Pinnixa for feeding activity (F54.90; MSE526.02, df51,36, p50.028) and a similar trend for short-duration activity periods (F51.57, MSE539.28, df51,28, p50.178). Total activity rates and proportional feeding activities were not different (Table 3). Time-averaged volume flow-rates were not significantly affected by crab treatment.
Average volume flow-rates were not related to average stroke frequency across worms
2
for either Pinnixa hosts (r 50.099, p50.219, n517) or for worms hosting Polyonyx
2
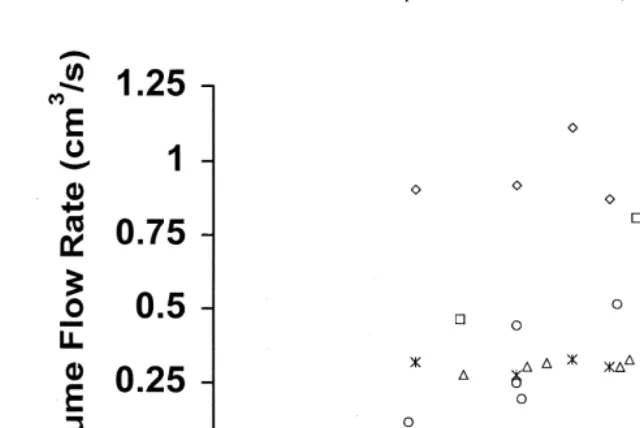
(r 50.001, p50.911, n532). Examination of the volume flow-rate / stroke rate relation-ship for Doppler records from individual worms (Fig. 2), however, suggests that these characters are somewhat related within some worms but are independent within others.
3.3. Laboratory activity measurements
The presence of Pinnixa in a host tube does not significantly alter water flow velocity or volume flow-rates (paired t-test: t51.54, p50.150, n514), the percent of time spent actively pumping (t50.52, p50.615, n514), fan stroke rates (t50.45, p50.663,
n514), or volume flow-rates averaged across active and inactive periods (t51.01,
p50.332, n514). Similarly, the presence or absence of Polyonyx has no effect on flow velocity or volume flow-rates (t50.49, p50.630, n513), percent activity (t50.69,
p50.505, n513), stroke rates (t50.34, p50.739, n513), or volume flow-rates (t5
1.12, p50.284, n513). Means for these variables are presented in Table 4.
Fig. 2. Plot of estimated fan stroke rate versus measured volume flow-rates for activity periods of five different Chaetopterus variopedatus in the field. All five worms were hosts to Pinnixa. Each point represents the values obtained for a distinctly separate activity period.
MSE5426.585, df51,23, p50.250), fan stroke rates (F50.03, MSE531.25, df51,23,
p50.861), or time-averaged volume flow-rates (F51.40, MSE50.00311, df51,23,
p50.249). Order effects (i.e. whether a given worm was tested with a crab during the first or second recording period) were tested for all activity variables and found to be nonsignificant as were crab treatment–order interactions.
Contrary to what was found in field recordings, worms in the laboratory did show positive correlations between average volume flow-rates and stroke rates for worms
2 2
hosting both Pinnixa (r 50.626, p50.016, n514) and Polyonyx (r 50.495, p50.092,
Table 4
Average laboratory activity measurements for Chaetopterus variopedatus housed in glass tubing with and
a
without crab symbionts
Pinnixa Polyonyx
With Without With Without
21
Water flow velocity (cm s ) 3.96 (0.32) 3.49 (0.29) 4.02 (0.49) 3.73 (0.39)
% Time pumping 19.2 (4.4) 16.4 (3.1) 28.0 (7.0) 22.5 (5.3)
21 21
Fan strokes min segment 20.2 (1.5) 22.3 (1.7) 20.0 (2.1) 20.4 (1.6)
21
Volume flow-rate (ml s ) 0.30 (0.02) 0.27 (0.02) 0.31 (0.04) 0.28 (0.03) Time-averaged
21
volume flow-rate (ml s ) 0.06 (0.01) 0.04 (0.01) 0.08 (0.02) 0.06 (0.02)
a
Table 5
Mean percentage of time spent by crab symbionts in different positions relative to the body position of a host
a
Chaetopterus
Position Crab species
Pinnixa Polyonyx
a a
In front of host head 47.4 42.1
a,b a
Behind host tail 36.5 41.6
In contact with head
b b
or fan body regions 16.1 16.1
a
Means are back-transformed from arcsine-square root transformed data. Sample sizes are ten and 13 crabs, respectively, for Pinnixa and Polyonyx. Means sharing the same letter within a column are not significantly different at p50.05 (Tukey HSD test).
n513) when worm weight and internal tube diameter are included in the regression models.
The position of crabs relative to the body of the worm during recording was observed, and neither Pinnixa (F54.32, MSE50.058, df52,27, p50.024) nor Polyonyx (F5
5.57, MSE50.051, df52,36, p50.001) were found to spend significantly different amounts of time in front of the head or behind the tail of the host worm, although both spent less time in contact with the region of the worm body near the fans where mechanical interference with pumping activity might take place (Table 5). Pinnixa adjusted their position relative to the worm’s body after only one of the 24 times that worms reversed positions in the tube, while Polyonyx changed position after five of 32 observed worm turns. Worms reversed body orientation on average four times per hour, but this rate was not different in worms when their tubes were unoccupied as compared to when hosting Pinnixa (t50.953, p50.358, n514) or Polyonyx (t51.00, p50.337,
n513).
4. Discussion
Symbiotic interactions between crustaceans and marine host species vary in character from examples of true mutualism (Glynn, 1980; Stimson, 1990) to commensalism (Wirtz and Diesel, 1983) to direct parasitism (Kuris, 1974). The exact nature of any specific relationship is dependent upon the ecology and behavior of both the host and the symbiont. Pinnotherid crabs in the genus Pinnotheres reduce filtration rates (Bierbaum and Shumway, 1988) and growth (Bierbaum and Ferson, 1986) in host bivalves because of the tissue damage that the crabs’ method of feeding inflicts on the host. Although
Pinnixa chaetopterana and Polyonyx gibbesi do not directly take food from Chaetop-terus hosts, their presence in the tube could influence host food uptake and growth either
by active filtering of the water pumped by the host or by passively presenting an obstruction to water flow through the worm tube.
Chaetopterus hosts which house Pinnixa in their tubes do have significantly higher
volume flow. Polyonyx is a significantly larger crab than Pinnixa (Grove and Woodin, 1996) and should remove food and oxygen from the water column at higher rates than
Pinnixa as well as presenting a greater passive obstruction to water flow through the
tube. Although this may explain differences in total activity times of worms hosting the two crabs, estimates of these specific factors suggest that they should in reality have minimal impact upon the host (see below). There appears to be no clear explanation why fan stroke rates should vary between crab treatments.
These activity differences, when integrated over a 6-month growth period in the laboratory, do not result in significantly different growth in worms with or without either crab species (Table 1). Although analysis of growth patterns in the lab experiments is complicated by the movement of crabs between hosts at undetermined times, no method of assignment of worms to crab treatment resulted in significant differences between treatments, suggesting that the presence of crabs has no real long-term effect on host worms. Experiments with Pinnixa rathbunae inhabiting Chaetopterus tubes have shown that movement of crabs between hosts occurs very quickly (within 1–2 weeks) after set-up of a laboratory population and remains relatively unchanged after that period (Britaev and Smurov, 1988). If this same behavior is found in Pinnixa chaetopterana, the second or third methods of assigning worms to growth rate treatments in Table 1 are more likely to give an accurate picture of conditions in our experiments than the first method would. In both of these methods, the trend is actually for increased growth with either species of crab present in the tube.
Given that individuals of Chaetopterus in intertidal populations in North Inlet survive for less than 2 years on average (Grove, unpublished data), a 6-month growth period represents a significant fraction of the lifespan of these animals. These experimental results suggest that the weight difference seen in field populations of worms hosting pairs of Pinnixa and Polyonyx are not due to differential crab effects on worm growth, but rather, may be due to recruitment of Pinnixa into newly settled Chaetopterus and subsequent replacement of these crabs by competitively dominant Polyonyx (Gray, 1961). Active movement of small Pinnixa out of sympatric populations of the abundant alternate polychaete host Amphitrite ornata may provide a pool of recruits which encounter Chaetopterus hosts at higher rates than Polyonyx larvae that lack an alternate host.
The lack of difference in activity patterns of individual worms when measured with and without crabs or between worms with the two species of crabs in the lab (Table 4) supports the general lack of difference found in the field samples. Sumida and Case (1983) have found that the activity patterns of Chaetopterus housed in natural and artificial glass tubes do not differ significantly in laboratory settings. Although all activity measures were lower in the laboratory than in the field, much of this difference is probably due to the difference in water temperature between the laboratory (178C) and the field (.268C) experiments. Activity rates of many polychaete worms, including the facultative filter feeder Nereis diversicolor, are known to be dependent upon temperature (Mangum and Sassaman, 1969; Veder et al., 1994).
21 21
ml O g2 h at 158C (Dales, 1969; Brown, 1977; Riisgard, 1989); so, total
consump-21
tion by a 15-g worm would be between 1.2 and 3.4 ml O h2 . The uptake rates of
21 21
Pinnixa and Polyonyx have been measured to be 60 and 40ml g h , respectively, at 158C (Craig, 1974). Thus, the additional oxygen demand placed upon a host worm by a pair of symbiotic crabs, each with a wet weight of 0.5 g (Grove and Woodin, 1996), would be less than 5% of the worm’s own requirements.
The passive pressure loss caused by the presence of crabs in a worm tube during active pumping can be estimated to be equally small. Pressure losses between the incurrent and excurrent openings of an actively pumping Chaetopterus housed in straight glass tubing have been measured at approximately 30 Pascals (Brown, 1977;
˚
Riisgard, 1989), with much of this loss caused by resistance to flow through the mucous
˚
filter (Riisgard, 1989). The additional pressure loss imposed by curvature of a natural tube should be on the order of 0.3 Pascal and should be independent of crab species present (Appendix). Losses through the constrictions in internal tube diameter caused by the presence of a crab can similarly be estimated to be approximately 0.075 Pascal (Appendix), or less than 0.25% of the normal pressure drop experienced by an active worm.
When feeding, Chaetopterus clears particles down to 1.5mm from the water column with 100% efficiency (Jørgensen et al., 1984). Although the feeding rates and filtering efficiency of the two crab species have not been studied, the efficiencies of other crustaceans employing setal filter feeding suggests that crabs positioned posteriorly to a feeding host probably cannot engage in energetically advantageous active feeding themselves (Nival and Nival, 1976; Hessen, 1985). It is possible that, when positioned anteriorly to a host, Polyonyx clears food particles from the water column more efficiently than does Pinnixa and causes a resultant increase in the percentage of time
Chaetopterus hosts spend actively pumping (Tables 3 and 4), but given that the crabs do
not actively position themselves to filter incoming water before the worm does so (Table 5) and that worms reposition themselves frequently, competition between the host and crabs for food probably occurs less than 50% of the time.
Activity patterns of Chaetopterus in the laboratory in this study were similar to those recorded by other authors in laboratory settings. Reported fan stroke rates vary between 13 and 64 strokes per fan segment per minute (Macgintie, 1939; Aksyuk and
˚
Sveshnikov, 1971; Brown, 1977; Riisgard, 1989), while previous volume flow-rate
21
estimates range from 0.13 to 0.54 ml s (Wells and Dales, 1951; Dales, 1969; Brown,
˚
1977; Riisgard, 1989). The similarity of responses of worms in this study and others is perhaps somewhat surprising given the sizes of worms in this study, which were at least three to five times larger than those used previously. This study is the first to measure the activity rates of Chaetopterus under field conditions which, given the natural occupation rates of tubes by crabs, provide ecologically more relevant measures of worm performance.
Chaetop-terus tubes after forced closure of circulation has been demonstrated by Wells and Dales
(1951). It is possible that reduced oxygen levels in the tube following sufficiently long tidal exposure periods affect the ability of the worms to feed until this deficit is made up. Given that the resting metabolic rate of Chaetopterus is only about 7% of its active rate, the water contained in a tube during low tide may contain sufficient oxygen to prevent the occupant from experiencing complete anoxia (Dales, 1969).
The tube-building polychaete Diopatra cuprea also shows reduced tube irrigation rates when O levels in its tube water become too low (Dales et al., 1970). Diopatra and2 Chaetopterus (which lacks respiratory pigments) both have low oxygen extraction
efficiencies. Other polychaetes which have higher efficiencies conversely show increased levels of activity in response to low oxygen levels in tube water (Weber, 1978; Woodin and Marinelli, 1991).
The positive relationship found between volume flow-rates and fan stroke rates of
˚
laboratory animals has also been reported by Riisgard (1989), who found relatively constant stroke volumes in undisturbed animals. With increasing disturbance or with contraction of the body axis from other causes, however, the volume displaced by each stroke may be reduced. Thus, the worms do have some control over the shape of this relationship. The large variation in the shapes of natural worm tubes (Brown, 1977; Grove, personal observation) may heavily influence the body orientation of active worms in the field and increase the variability between individuals, while a low variance in stroke rate / pumping rate relations is retained within any given individual (Fig. 2). The uniform shape of artificial glass tubes used in laboratory experiments may act to reduce the potential variation in body positioning available in field samples.
Historically, parasite–host relationships were often considered to evolve in directions that resulted in less harm to the host, i.e. away from true parasitism and towards more commensal or even mutualistic relations (Burnet and White, 1972; Alexander, 1981; Palmieri, 1982). More recent theoretical studies suggest that the evolution of virulence is controlled by tradeoffs between the transmission rates of parasites to new, uninfected hosts and the level of damage done to already infected hosts. Since parasite reproduction and transmission are generally dependent on the use of host resources with the consequent infliction of damage, transmissibility and virulence should be linked (May, 1991). Models based on both population dynamics (Anderson and May, 1981) and game-theory (Renaud and de Meeus, 1991) can predict either increasing or decreasing levels of virulence depending upon the degree of linkage of transmission rates and virulence in a specific symbiotic system. Given that transmission rates of pea crab species between bivalve hosts may be very low, there may have been selection for increased reproductive rates at the expense of direct damage to the host. Conversely, transmission rates of Pinnixa and Polyonyx between hosts may be high enough to obviate the need for increasing reproduction to the point at which hosts are harmed. Empirical studies that determine the success with which pea crab larvae are distributed to new hosts and initiate symbiotic relationships would be useful in addressing this difference in crab life-history patterns.
In cases where parasites compete for host resources (such as could be the case for
Pinnixa and Polyonyx), the selective pressure for increased virulence is especially strong
it is evident from this study that Pinnixa and Polyonyx do not gather resources in ways or at sufficiently high levels that they either actively or passively affect the activities and subsequent growth of Chaetopterus hosts. The effects of the presence of the crabs on reproduction and survival of the host worms is unknown, but given the present results are unlikely to be significant.
The elucidation of the commensal nature of the relationship between these symbiotic crab species is one step towards a useful synthesis of ecological and phylogenetic data in describing evolutionary trends in symbiosis. Fossil- or genetics-based phylogenies are unfortunately largely lacking for pinnotherids except the genus Dissodactylus (Griffith, 1987). When such information becomes available, the ecological data obtained from studies such as this one may be used to describe not just the coevolution of specific host–symbiont relationships, but also macroevolutionary patterns in the radiation of symbionts to new hosts with a broad range of life histories (Brooks, 1988). It is evident from a comparison of the effects of Pinnixa chaetopterana and of the bivalve–symbiont species in Pinnotheres on their hosts that the path to differing levels of virulence can vary widely within closely related species depending on the ecology of the two interacting species.
Acknowledgements
Dr Mark Denny and John Lee of the Hopkins Marine Station kindly provided us with the circuit diagrams for the Doppler probe. Dean Pentcheff assisted in writing thePERL
and SAS computer code for the Fourier transformations. Michelle Johnson and Dr Courtney Richmond provided field help, while Dr Barbara McCraith and Rick Matthews helped keep the water running in the growth rate experiments. The Baruch Institute for Marine Biology and Coastal Research provided access to field sites. This research was supported by an NSF Pre-doctoral Fellowship to MWG, NSF grant IBN-922225 to DSW, and NSF grant OCE-8900212 and EPA grant R821838-01-0 to SAW. Comments by Drs David Lincoln and Mark Luckenbach helped improve the manuscript. This is Belle W. Baruch contribution number 1207. [RW]
Appendix
Pressure drop due to tube curvature
In these calculations, we are assuming a Chaetopterus tube with a maximum internal diameter (D ) of 1.75 cm, a bend angle of 1808, and a bend radius of 10 cm. The worm is assumed to be pumping water at a rate sufficient to produce an exit flow velocity of 5
21
cm s at the centerline of a 4.5-mm diameter tube opening, yielding a flow velocity (U )
21
in the main part of the tube of 0.165 cm s .
The formula for the drop in pressure occurring in a curved pipe (Blevins, 1984) is:
2
21
wherer is the density of seawater (1.024 g ml ), K the dimensionless loss coefficient (here estimated to be 200) for a Reynolds number of 26.25 (Bruins, 1940), L and L the1 2
lengths of straight tube attached to the curved portion of the tube (assumed to be 8 cm each), and f the friction factor for laminar flow in smooth pipe (64 /Re52.438).
21 22
Substituting these values into the equation, we estimate Dp53.09 g cm s , or 0.309 Pascals. This is only 1% of the 30-Pa pressure drop experienced by actively pumping worms due to resistance of the mucous feeding bag and kinetic losses in the
˚
constricted tube openings (Brown, 1977; Riisgard, 1989).
Secondary (or radial) flow in curved pipes is estimated by the Dean number, which is
0.5
equal to Re(D/d ) , where d is the diameter of the curve in the pipe (here 20 cm). Dean numbers less than 10 indicate small secondary flows and flow profiles similar to those of straight pipes. The estimated Dean number for a Chaetopterus tube with the dimensions above would be 7.76, indicating that the flow through such a tube approximates that which would be found in a straight pipe with similar dimensions.
Pressure drop due to presence of crabs
In these calculations we assume the same tube dimensions and flow characteristics as in the tube curvature calculations above. We further assume that the tube is occupied by a single Pinnixa whose carapace length and height are 5 mm and whose carapace width is 1.1 cm (Grove and Woodin, 1996). Flow area in the unoccupied portion of the tube
2
(A ) is 2.405 cm , while flow area in the tube occupied by the crab (A ) is reduced to1 2 2
2.208 cm . By conservation of mass, the flow velocity (V ) through area A is increased2 21
from 0.165 to 0.180 cm s .
The pressure loss in a sudden contraction in a pipe can be estimated by (Blevins, 1984):
2 2
* *
Dp / 0.5rV 512(A /A )2 1 1K1( f L /D )1 1 1 1( f L /D )2 2 2
where the loss coefficient K is equal to 0.5(12A /A ). f , L , and D are the friction2 1 1 1 1
factor (64 /Re in laminar flow), length (assumed to be 15 cm), and diameter of the unoccupied tube between the tube opening and the crab.
The friction factor f for the section of tube occupied by the crab can be estimated2
from values calculated for eccentric annuli within a circular pipe. Given a pipe radius a of 0.875 cm and a crab body radius b of 0.25 cm, the friction coefficient for an annulus with ratio b /a of 0.286 centered at the centerline of a pipe is 92.10 /Re or 3.509. Since the center of the crab’s body is offset from the pipe centerline (by c50.625 cm), the friction factor is reduced from this value. For values of c /(a2b)51.0 and b /a50.286, the friction factor calculated for c50 is reduced by approximately 11.1% (White, 1974) to yield an estimate for f of 3.119. The length L is the width of the crab’s body (1.12 2
cm) and D is the unoccluded pipe diameter in the section occupied by the crab (1.252
cm). Substituting these values into the equation above yields an estimate forDp of 0.040
Pascal.
2 2
*
Dp / 0.5rV 5(A /A )1 2 211K1( f L /D )1 1 1
2
where the loss coefficient K5(12A /A ) . Thus, the loss due to expansion is 0.0352 1
Pascal.
The sum of losses due to contraction of the tube opening around an individual of
Pinnixa and its subsequent re-expansion is 0.075 Pascal. Similar estimates for Polyonyx
(with carapace length and height of 7 mm and width of 1 cm) yields an estimated loss of 0.089 Pascal. These estimates assume that the crabs are positioned in the widest part of the host tube; positioning in the constricted ends of the tubes would, of course, greatly increase the additional pressure drop that must be overcome by the worm when pumping. Given, however, that the crabs appear to position themselves randomly along the tube, the passive effects of the crabs on worm pumping activity should generally be a trivial component of the worm’s normal work load.
[RW]
References
Aksyuk, T.S., Sveshnikov, V.A., 1971. A plunger mechanism for expulsion of water in Chaetopterus
variopedatus (Polychaeta, Annelida). Dokl. Akad. Nauk SSSR, Biol. Sci. 197, 285–288.
Alexander, M., 1981. Why microbial predators and parasites do not eliminate their prey and hosts. Annu. Rev. Microbiol. 35, 113–133.
Anderson, G.L., 1975. The effects of intertidal height and the parasitic crustacean Fabia subquadrata Dana on the nutrition and reproductive capacity of the California sea mussel Mytilus edulis Conrad. Veliger 17, 299–306.
Anderson, R.M., May, R.M., 1981. The population dynamics of microparasites and their invertebrate hosts. Philos. Trans. R. Soc. Lond., Ser. B 291, 451–524.
Baker, D.W., 1970. Pulsed ultrasonic Doppler bloodflow sensing. IEEE Trans. Sonics Ultrasonics SU–17, 170–185.
Bell, J.L., 1984. Changing residence: dynamics of the symbiotic relationship between Dissodactylus mellitae Rathbun (Pinnotheridae) and Mellita quinquiesperforata (Leske) (Echinodermata). J. Exp. Mar. Biol. Ecol. 82, 101–115.
Bierbaum, R., Ferson, S., 1986. Do symbiotic pea crabs decrease growth rates in mussels? Biol. Bull. 170, 51–61.
Bierbaum, R., Shumway, S.E., 1988. Filtration and oxygen consumption in mussels, Mytilus edulis, with and without pea crabs, Pinnotheres maculatus. Estuaries 11, 264–271.
Blevins, R.D., 1984. Applied Fluid Dynamics Handbook, Van Nostrand Reinhold, New York.
Boesch, D.F., 1971. On the occurrence of Pinnixa lunzi Glassell (Decapoda, Pinnotheridae) off Virginia, USA. Crustaceana 20 (2), 219–220.
Britaev, T.A., Smurov, A.V., 1988. Distribution and redistribution of the commensal crab Pinnixa rathbunae (Pinnotheridae) in its hosts. Dokl. Akad. Nauk SSSR (Engl. Trans.) 300, 370–373.
Brooks, D.R., 1988. Macroevolutionary comparisons of host and parasite phylogenies. Annu. Rev. Ecol. Syst. 19, 235–259.
Brown, S.C., 1977. Biomechanics of water-pumping by Chaetopterus variopedatus Renier: kinetics and hydrodynamics. Biol. Bull. 153, 121–132.
Brown, S.C., Rosen, J.S., 1978. Tube-cleaning behaviour in the polychaete annelid Chaetopterus variopedatus (Renier). Anim. Behav. 26, 160–166.
Bruins, P.F., 1940. Friction of fluid in solder-type fittings. Trans. Am. Inst. Chem. Eng. 36, 721–739. Burnet, M., White, D.M., 1972. Natural History of Infectious Disease, 4th Edition, Cambridge University
Christensen, A.M., McDermott, J.J., 1958. Life-history and biology of the oyster crab, Pinnotheres ostreum Say. Biol. Bull. 114, 146–179.
Caine, E.A., 1975. Feeding and masticatory structures of selected Anomura (Crustacea). J. Exp. Mar. Biol. Ecol. 18, 277–301.
Claessen, D., deRoos, A.M., 1995. Evolution of virulence in a host–pathogen system with local pathogen transmission. Oikos 74, 401–413.
Craig, W.J., 1974. Physiological ecology of the commensal crabs, Polyonyx gibbesi Haig and Pinnixa
chaetopterana Stimpson. Oecologia 15, 235–244.
Dales, R.P., 1969. Respiration and metabolism in annelids. In: Chemical Zoology, Vol. 4, Academic Press, New York, pp. 93–109.
Dales, R.P., Mangum, C.P., Tichy, J.C., 1970. Effects of changes in oxygen and carbon dioxide concentrations on ventilation rhythms in onuphid polychaetes. J. Mar. Biol. Assoc. UK 50, 365–380.
Dix, T.G., 1973. Mantle changes in the pearl oyster Pinctada maxima induced by the pea crab Pinnotheres
villosulus. Veliger 15, 330–331.
Enders, H.E., 1905. Notes on the commensals found in the tubes of Chaetopterus pergamentaceus. Am. Nat. 39, 37–40.
Glynn, P.W., 1980. Defense by symbiotic Crustacea of host corals elicited by chemical cues from predator. Oecologia 47, 287–290.
Griffith, H., 1987. Phylogenetic relationships and evolution in the genus Dissodactylus Smith, 1870 (Crustacea: Brachyura: Pinnotheridae). Can. J. Zool. 65, 2292–2310.
Gray, I.E., 1961. Changes in abundance of the commensal crabs of Chaetopterus. Biol. Bull. 120, 353–359. Grove, M.W., Woodin, S.A., 1996. Host choice and conspecific recognition in a pea crab, Pinnixa
chaetopterana (Brachyura, Pinnotheridae). Biol. Bull. 190, 359–366.
Haven, D.S., 1958. The pea crab Pinnotheres ostreum as a parasite of the oyster. Virginia J. Sci. 8, 301–302. Haven, D.S., 1959. Effects of pea crabs Pinnotheres ostreum on oysters Crassostrea virginica. Proc. Natl.
Shellfish. Assoc. 49, 77–86.
Hessen, D.O., 1985. Filtering structures and particle size selection in coexisting Cladocera. Oecologia 66, 368–372.
Hines, A.H., 1982. Allometric constraints and variables of reproductive effort in brachyuran crabs. Mar. Biol. 69, 309–320.
Hines, A.H., 1992. Constraint on reproductive output in brachyuran crabs: pinnotherids test the rule. Am. Zool. 32, 503–511.
Jones, J.B., 1977. Natural history of the pea crab in Wellington Harbor, New Zealand. NZ J. Mar. Freshwater Res. 11, 667–676.
˚
Jørgensen, C.B., Kiørboe, T., Møhlenberg, F., Riisgard, H.U., 1984. Ciliary and mucus-net filter feeding, with special reference to fluid mechanical characteristics. Mar. Ecol. Prog. Ser. 45, 205–216.
Jumars, P., 1993. Concepts in Biological Oceanography, Oxford University Press, Oxford.
Kruczynski, W.L., 1972. The effect of the pea crab, Pinnotheres maculatus Say, on growth of the bay scallop,
Argopecten irradians concentricus (Say). Chesapeake Sci. 13, 218–220.
Kuris, A.M., 1974. Trophic interactions: similarity of parasitic castrators to parasitoids. Q. Rev. Biol. 49, 129–148.
Lalitha Devi, S., 1986. Some aspects of biology and effect of infestation of Pinnotheres placunae Hornell and Southwell of Kakinnada Bay. J. Mar. Biol. Assoc. India 28, 113–123.
Macgintie, G.E., 1939. The method of feeding of Chaetopterus. Biol. Bull. 77, 115–118.
Mangum, C.P., Sassaman, C., 1969. Temperature sensitivity of active and resting metabolism in a polychaetous annelid. Comp. Biochem. Physiol. 30, 111–116.
May, R.M., 1991. Dynamics and genetics of host–parasite associations. In: Toft, C.A., Aeschlimann, A., Bolis, L. (Eds.), Parasite–Host Associations: Coexistence Or Conflict, Oxford University Press, Oxford, pp. 102–128.
McDermott, J.J., 1962. The incidence and host–parasite relations of pinnotherid crabs (Decapoda, Pin-notheridae). Coastal Shallow Waters Res. Conf. 1, 162–164.
Nowak, M.A., May, R.M., 1994. Superinfection and the evolution of parasite virulence. Proc. R. Soc. Lond., Ser. B 255, 81–89.
O’Beirn, F.X., Walker, R.L., 1999. Pea crab, Pinnotheres ostreum Say, 1817, in the Eastern oyster,
Crassostrea virginica (Gmelin, 1791): prevalence and apparent adverse effects on oyster gonad production.
Veliger 42 (1), 17–20.
Orton, J.H., 1920. The mode of feeding and sex phenomena in the pea crab (Pinnotheres pisum). Nature 106, 533.
Palmieri, J.R., 1982. Be fair to parasites. Nature 298, 220.
Pearse, A.S., Humm, H.J., Wharton, G.W., 1942. Ecology of sand beaches at Beaufort, NC. Ecol. Monogr. 12 (2), 35–190.
Pearce, J.B., 1966. The biology of the mussel crab, Fabia subquadrata, from the waters of the San Juan Archipelago, Washington. Pac. Sci. 20, 3–35.
Poulin, R., 1995a. Clutch size and egg size in free-living and parasitic copepods: a comparative analysis. Evolution 49 (2), 325–336.
Poulin, R., 1995b. Evolutionary influences on body size in free-living and parasitic isopods. Biol. J. Linnean Soc. 54, 231–244.
Prezenger, C., 1979. Effect of Pinnotheres hickmani on neutral red clearance by Mytilus edulis. Aust. J. Mar. Freshwater Res. 30, 547–550.
Renaud, F., de Meeus, T., 1991. A simple model of host–parasite evolutionary relationships. J. Theor. Biol. 152, 319–327.
˚
Riisgard, H.U., 1989. Properties and energy cost of the muscular piston pump in the suspension feeding polychaete Chaetopterus variopedatus. Mar. Ecol. Prog. Ser. 56, 157–168.
Sumida, B.H., Case, J.F., 1983. Food recognition by Chaetopterus variopedatus (Renier): synergy of mechanical and chemical stimulation. Mar. Behav. Physiol. 9, 249–274.
Stauber, L.A., 1945. Pinnotheres ostreum, parasitic on the American oyster, Ostrea(Gryphaea) virginica. Biol. Bull. 88, 269–291.
Stimson, J., 1990. Stimulation of fat-body production in the polyps of the coral Pocillopora damicornis by the presence of mutualistic crabs of the genus Trapezia. Mar. Biol. 106, 211–218.
Takeda, S., Tamura, S., Washio, M., 1997. Relationship between the pea crab Pinnixa tumida and its endobenthic holothurian host Paracaudina chilensis. Mar. Ecol. Prog. Ser. 149, 143–154.
Telford, M., 1982. Echinoderm spine structure, feeding, and host relationships of four species of Dissodactylus (Brachyura: Pinnotheridae). Bull. Mar. Sci. 32, 584–594.
Van Baalen, M., Sabelis, M.W., 1995. The dynamics of multiple infection and the evolution of virulence. Am. Nat. 146, 881–910.
˚
Veder, A., Andersen, B.B., Riisgard, H.U., 1994. Field investigations of pumping activity of the facultatively feeding polychaete Nereis diversicolor using and improved infrared phototransducer system. Mar. Ecol. Prog. Ser. 103, 91–101.
Vogel, S., 1981. Life in Moving Fluids, Princeton University Press, Princeton.
Wass, M.L., 1955. The decapod crustaceans of Alligator Harbor and adjacent inshore areas of northwestern Florida. Q. J. Fla. Acad. Sci. 18 (3), 129–176.
Weber, R.E., 1978. Respiration. In: Mill, P.J. (Ed.), Physiology of Annelids, Academic Press, London, pp. 369–392.
Wells, G.P., Dales, R.P., 1951. Spontaneous activity patterns in animal behaviour: the irrigation of the burrow ¨
in the polychaetes Chaetopterus variopedatus Renier and Nereis diversicolor O.F. Muller. J. Mar. Biol. Assoc. UK 29, 661–680.
Wells, H.W., Wells, M.J., 1961. Observations on Pinnaxodes floridensis, a new species of pinnotherid crustacean commensal in holothurians. Bull. Mar. Sci. 11, 267–279.
White, F.M., 1974. Viscous Fluid Flow, McGraw-Hill, New York.
Wirtz, P., Diesel, R., 1983. The social structure of Inachus phalangium a spider crab associated with the sea anenome Anemonia Sulcal. Z. Tierpsych. 62, 209–234.