PERTUMBUHAN UDANG VANAME PADA TAHAP
PEMBENIHAN DAN PEMBESARAN PASCARENDAM
HORMON PERTUMBUHAN REKOMBINAN IKAN
KERAPU KERTANG
AULIA SAPUTRA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul: Pertumbuhan Udang Vaname pada Tahap Pembenihan dan Pembesaran Pascarendam Hormon Pertumbuhan Rekombinan Ikan Kerapu Kertang adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dan tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2016
RINGKASAN
AULIA SAPUTRA. Pertumbuhan udang vaname pada tahap pembenihan dan pembesaran pascarendam hormon pertumbuhan rekombinasi ikan kerapu kertang. Dibimbing oleh ODANG CARMAN dan ALIMUDDIN.
Peningkatan produksi dapat dicapai dengan meningkatkan laju pertumbuhan. Pertumbuhan udang vaname dapat dipacu melalui perendaman hormon pertumbuhan rekombinan ikan kerapu kertang (rElGH). Pada proses perendaman, umur atau stadia udang vaname merupakan salah satu aspek yang perlu diketahui. Secara teknis, proses perendaman rElGH dapat dilakukan pada stadia naupli, pascalarva muda (PL2-PL5) atau pascalarva yang akan ditebar di tambak (PL10-PL14). Penggunaan stadia yang lebih muda lebih efisien karena jumlah udang yang dapat direndam lebih banyak. Namun stadia yang lebih tua diduga lebih efektif karena memiliki insang yang lebih berkembang dan insang merupakan organ penting yang menyerap rElGH pada proses perendaman. Penelitian ini bertujuan untuk mengevaluasi efek perendaman rElGH pada stadia naupli, PL4 dan PL11 terhadap pertumbuhan udang vaname Litopenaeus vannamei pada tahap pembenihan dan pembesaran.
Dosis perendaman yang digunakan pada penelitian ini adalah 15 mg/L. Jumlah benih udang yang direndam diseragamkan berdasarkan biomassa sekitar 5 mg/L air laut, sehingga jumlah benih udang dalam tiap liter pada setiap stadia adalah adalah 60 ribu ekor naupli, 2250 ekor PL4 dan 400 ekor PL11. Durasi perendaman dilakukan selama dua jam.
Penelitian ini dilakukan tiga tahap, yaitu: tahap pembenihan 1, tahap pembenihan 2 dan tahap pembesaran. Pada tahap pembenihan 1 dimulai dengan melakukan perendaman rElGH pada stadia naupli dan perendaman tanpa rElGH sebagai kontrol. Setelah proses perendaman, naupli ditebar ke dalam bak fiber berisi 600 L air laut dengan kepadatan 200 ekor/L kemudian diperlihara selama 11 hari. Pada tahap pembenihan 2, sebanyak 30 ribu ekor benih yang direndam rElGH pada stadia naupli, 30 ribu ekor benih yang direndam rElGH pada stadia PL4 dan 30 ribu ekor benih kontrol masing-masing diperlihara di dalam bak fiber berisi 750 L air laut selama 8 hari. Pada tahap pembesaran, 100 ekor benih yang direndam rElGH pada stadia naupli, 100 ekor benih yang direndam rElGH pada stadia PL4, 100 ekor benih yang direndam rElGH pada stadia PL11 dan kontrol masing-masing dipelihara di dalam bak fiber berisi 750 L air laut selama 55 hari.
mempengaruhi tingkat kelangsungan hidup, rasio konversi pakan dan koefisien variasi bobot (P≥0,05).
Pada tahap pembenihan, benur yang direndam rElGH pada stadia PL4 dapat mencapai ukuran jual (>1 cm) dua hari lebih cepat dibandingkan benur yang tidak diberi rElGH, sehingga mengurangi biaya produksi. Pemberian rElGH juga menghasilkan benur dengan keseragaman ukuran yang lebih baik, walaupun kualitas daya tahan benur tidak berbeda dengan benur yang tidak diberi rElGH. Perbaikan kualitas ukuran tersebut akan mempengaruhi daya saing benur ketika dipasarkan.
Pada tahap pembesaran, perendaman pada stadia PL11 menghasilkan peningkatan bobot dan biomassa yang lebih tinggi daripada perendaman pada stadia naupli dan PL4. Hal ini menunjukkan bahwa respons udang vaname stadia PL11 terhadap perendaman rElGH lebih baik daripada stadia sebelumnya. Perbedaan respons tersebut diduga karena perbedaan kapasitas insang dan lapisan epidermis benur dalam menyerap rElGH.
Pemberian rElGH dapat meningkatkan pertumbuhan udang vaname. Untuk meningkatkan dan menyeragamkan ukuran benur pada tahap pembenihan dapat menerapkan perendaman rElGH pada stadia PL4. Untuk meningkatkan rataan bobot tubuh dan biomassa udang pada tahap pembesaran dapat menerapkan perendaman rElGH pada stadia PL11.
SUMMARY
AULIA SAPUTRA. White shrimp growth at nursery and grow-out phases after immersing in giant grouper recombinant growth hormone. Supervised by ODANG CARMAN and ALIMUDDIN.
Production escalation can be obtained by increasing growth rate. The growth of white shrimp (Litopenaeus vannamei) can be improved by using recombinant growth hormone through immersion administration. In immersion administration, age or shrimp stage is one of the aspect to be determined. Technically, recombinant growth hormone immersion can be applied at the nauplius stage, early postlarvae (PL2-PL5) or late postlarvae (PL10-PL14). Immersion at nauplius stage possibly will more efficient because a large number of shrimp can be treated at once. However, immersion at PL stage is expected to be more effective due to their developed organs, which is important aspect in absorbing the hormone during the immersion process. This research was performed to evaluate the white shrimp growth at nursery and grow-out phases after recombinant giant grouper growth hormone (rElGH) immersion at nauplius, PL4 and PL11 stages.
The immersion was performed at the dose 15 mg of rElGH/L seawater for two hours. The shrimp density at each stages were 60000 nauplii, 2250 PL4, 400 PL11 for one liter seawater, or equivalen 5 mg biomass per liter respectively.
The research was conducted in three steps: nursery 1, nursery 2 and grow-out phases. During nursery 1, the rElGH immersion at the nauplius stage was compared to immersion without rElGH as control. After the immersion process, nauplius were reared in separated fiber tank (600 L seawater) for 11 days at a density of 200 nauplii/L. During nursery 2, 30000 postlarvae of rElGH immersed at nauplius stage, rElGH immersion at PL4 and control were reared in separated fiber tank (750 L seawater) for 8 days. During the grow-out phase, 100 PL11 of rElGH immersed at nauplius stage, rElGH immersed at PL4, rElGH immersed at PL11 and control were separately reared into fiber tanks containing 750 L seawater for 55 days.
The results showed that the rElGH immersion at the nauplius stage were significantly increased 19% metamorphosis index value at the end of metamorphosis process (nine days after treatment) compared to control (P<0.05). At the end of the 2, rElGH immersion at PL4 stage increased 19% of body length, 30% of the body weight and the coefficient of length variation was significantly 35% lower compared to control (P<0.05). However, the rElGH immersion at different stages of white shrimp was not significantly affected on the shrimp hardiness. The activities and the stress test using both aquadest and formalin 200 µL/L of rElGH immersed at the nauplius or PL4 stages shrimps were not significantly different compared to the control (P≥0.05). At the grow-out phase, rElGH immersion at the PL11 stage enhanced 38% of body weight and 32% of biomass compared with control (P<0.05), but there no significantly differences on the survival rate, feed conversion ratio and size variation (P≥0.05).
better shrimp size uniformity, even the response to stress were similar. The improved size quality might increase the competitiveness of the mentioned shrimp on the market.
During the grow-out phase, rElGH immersion at the PL11 stage resulted in increasing body weight and biomass. It showed that the PL11 responded better to the rElGH immersion compared to nauplius and PL4 stages. The difference in term of response was expected to be related to the gill capacity and epidermis in absorbing rElGH.
The immersion of rElGH was shown to increased shrimp growth. To increase the size and shrimp uniformity in nursery, rElGH immersion could be applied at the PL4 stage. To increase shrimp body weight and biomass at the grow-out, rElGH could be applied at the PL11 stage.
viii
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
ix
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
AULIA SAPUTRA
PERTUMBUHAN UDANG VANAME PADA TAHAP
PEMBENIHAN DAN PEMBESARAN PASCARENDAM
HORMON PERTUMBUHAN REKOMBINAN IKAN
KERAPU KERTANG
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
x
Judul Tesis : Pertumbuhan Udang Vaname Pada Tahap Pembenihan dan Pembesaran Pascarendam Hormon Pertumbuhan Rekombinan Ikan Kerapu Kertang
Nama : Aulia Saputra
NIM : C151114091
Disetujui oleh Komisi Pembimbing
Dr Ir Odang Carman, MSc Ketua
Dr Alimuddin, SPi, MSc Anggota
Diketahui oleh
Ketua Program Studi Ilmu Akuakultur
Dr Ir Widanarni, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian: 29 Desember 2015
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala nikmat dan karunia-Nya sehingga karya ilmiah ini dapat diselesaikan. Tema yang dipilih dalam karya ilmiah ini adalah perendaman hormon pertumbuhan rekombinan, dengan judul Pertumbuhan Udang Vaname Pada Tahap Pembenihan dan Pembesaran Pascarendam Hormon Pertumbuhan Rekombinan Ikan Kerapu Kertang.
Penulis menyampaikan terima kasih kepada Bapak Dr Odang Carman dan Dr Alimuddin atas bimbingan dan motivasi yang telah diberikan. Ucapan terima kasih penulis sampaikan kepada Bapak Dr Dedi Jusadi selaku penguji luar komisi dan Bapak Dr Eddy Supriyono selaku wakil program studi Ilmu akuakultur atas kritik dan saran. Penghargaan dan terima kasih juga penulis sampaikan kepada Bapak Dr Agus Somamihardja dari PT Suri Tani Pemuka dan Bapak Asep Yusup, SPi beserta staf unit Pembenihan Udang PT Suri Tani Pemuka Negara, Bali atas segala bantuan kepada penulis selama penelitian. Ungkapan terima kasih juga disampaikan kepada keluarga atas segala do’a dan kasih sayangnya.
Akhir kata, penyusun berharap semoga karya ilmiah ini dapat bermanfaat bagi semua pihak yang membutuhkan.
Bogor, Januari 2016
DAFTAR ISI
DAFTAR TABEL xiv
DAFTAR GAMBAR xiv
DAFTAR LAMPIRAN xiv
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
Hipotesis Penelitian 2
METODE 2
Waktu dan Tempat Penelitian 2
Materi Uji 2
Prosedur Perendaman 2
Prosedur Penelitian Tahap Pembenihan 1 2
Prosedur Penelitian Tahap Pembenihan 2 4
Prosedur Penelitian Tahap Pembesaran 5
Analisis Data 6
HASIL DAN PEMBAHASAN 6
Hasil Tahap Pembenihan 1 6
Hasil Tahap Pembenihan 2 7
Hasil Tahap Pembesaran 10
Pembahasan 11
KESIMPULAN 14
DAFTAR PUSTAKA 14
LAMPIRAN 17
DAFTAR TABEL
1. Total pakan yang diberikan setiap hari pada tahap pembenihan 1 3 2. Kuantifikasi nilai mutu metamorfosis udang vaname 3 3. Total pakan yang diberikan setiap hari pada tahap pembenihan 2 4 4. Nilai mutu metamorfosis pascarendam rElGH pada stadia naupli 6 5. Aktivitas benur dan uji stres benur setelah simulasi transportasi
selama 2 jam 9
6. Pertumbuhan harian, tingkat kelangsungan hidup, tingkat konversi
pakan dan koefisien variasi bobot udang pada tahap pembesaran 11
DAFTAR GAMBAR
1. Aktivitas larva pascarendam pada benur yang direndam rElGH pada
stadia naupli (A1) dan kontrol (K1). 7
2. Aktivitas benur pascarendam PL4 pada benur yang direndam rElGH pada stadia naupli (A2), benur yang direndam rElGH pada stadia PL4
(B2) dan kontrol (K2). 7
3. Pertumbuhan panjang benur yang direndam rElGH pada stadia naupli (A2) benur yang direndam rElGH pada stadia PL4 (B2) dan kontrol (K2) yang diukur pada hari ke-2 (stadia PL5), 4 (PL7), 6 (PL9) dan
akhir tahap pembenihan 2 (PL11). 8
4. Bobot rerata benur PL11 yang direndam rElGH pada stadia naupli (A2), benur PL11 yang direndam rElGH pada stadia PL4 (B2) dan
benur PL11 kontrol (K2). 8
5. Bobot rerata benur PL11 yang direndam rElGH pada stadia naupli (A2), bebenur PL11 yang direndam rElGH pada stadia PL4 (B2) dan
benur PL11 kontrol (K2). 9
6. Pertumbuhan bobot udang yang direndam rElGH pada stadia naupli (A3), udang yang direndam rElGH pada stadia PL4 (B3), udang yang direndam rElGH pada stadi PL11 (C3) dan kontrol (K3) selama tahap
pembesaran. 10
7. Biomassa udang yang direndam rElGH pada stadia naupli (A3), udang yang direndam rElGH pada stadia PL4 (B3), udang yang direndam
rElGH pada stadi PL11 (C3) dan kontrol (K3) pada tahap pembesaran. 11
DAFTAR LAMPIRAN
1. Komposisi dan kadar protein pakan buatan yang digunakan pada tahap
pembenihan 1 18
2. Metamorfosis udang menurut Wyban dan Sweeney (1991) beserta
xv
3. Komposisi dan kadar protein pakan buatan yang digunakan pada tahap
pembenihan 2 20
4. Jumlah dan jenis pakan buatan yang diberikan pada hari ke-1 hingga
hari ke-25 tahap pembesaran 21
5. Jenis pakan dan persentase pemberian pakan hari ke-26 hingga hari
PENDAHULUAN
Latar Belakang
Peningkatan produksi dapat dicapai dengan meningkatkan laju pertumbuhan. Teknologi yang dapat digunakan untuk meningkatkan laju pertumbuhan adalah dengan menggunakan hormon pertumbuhan rekombinan (recombinant growth hormone, rGH). Pemberian rGH telah diuji dan dilaporkan secara signifikan dapat meningkatkan laju pertumbuhan pada ikan yang sama dengan spesies sumber rGH (Funkenstein et al. 2005, Acosta et al. 2007), atau berbeda spesies (Promdonkoy et al. 2004, Liao et al. 2008, Acosta et al. 2009, Alimuddin et al. 2010, Handoyo et al. 2012). Hormon pertumbuhan dari mamalia juga dapat meningkatkan pertumbuhan ikan (Silverstein et al. 2000, Leedom et al. 2002, Haghighi et al. 2010). Bahkan hormon pertumbuhan dari vertebrata juga telah teruji efektivitasnya terhadap pertumbuhan avertebrata (Moriyama dan Kawauchi 2004, Moriyama et al. 2009). Berdasarkan hal tersebut, maka penggunaan rGH dari vertebrata khususnya ikan diduga akan memberikan efek yang positif terhadap pertumbuhan udang vaname. Pada faktanya pertumbuhan udang vaname dapat dipacu dengan pemberian rGH ikan nila (Santiesteban et al. 2010) dan rGH ikan kerapu kertang (Subaidah et al. 2012).
Perbedaan metode pemberian, dosis dan umur atau stadia benih udang berpotensi memberikan hasil yang berbeda. Pemberian rGH dapat dilakukan melalui penyuntikan, pakan dan perendaman. Metode penyuntikan, rGH dapat dipastikan masuk ke tubuh melalui peredaran darah. Namun, secara teknis pemberian rGH melalui perendaman dan pakan lebih mudah dan lebih praktis dilakukan daripada penyuntikan, terutama jika jumlah organisme target sangat banyak. Menurut Subaidah (2013) pemberian rElGH melalui perendaman lebih efektif dibandingkan melalui pakan untuk meningkat pertumbuhan udang.
2
Tujuan Penelitian
Penelitian ini bertujuan untuk mengevaluasi efek perendaman hormon pertumbuhan rekombinan ikan kerapu kertang (rElGH) pada stadia naupli, PL4 dan PL11 terhadap pertumbuhan udang vaname Litopenaeus vannamei pada tahap pembenihan dan pembesaran.
Manfaat Penelitian
Manfaat penelitian ini diharapkan dapat memberikan pengetahuan yang lebih rinci tentang prosedur perendaman rElGH pada benih udang vaname.
Hipotesis Penelitian
Perendaman rElGH pada benih udang vaname pada stadia yang berbeda akan mengakibatkan laju pertumbuhan yang berbeda-beda.
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan di unit pembenihan udang PT. Suri Tani Pemuka, Negara, Bali pada bulan November 2014 sampai dengan Februari 2015.
Materi Uji
Benih udang vaname specific pathogen free (SPF) stadia naupli diperoleh dari unit pembenihan udang PT. Suri Tani Pemuka, Banyuwangi, Jawa Timur. Hormon pertumbuhan rekombinan yang digunakan adalah hormon pertumbuhan rekombinan ikan kerapu kertang (rElGH) yang dikembangkan oleh Alimuddin et al. (2010) dan diproduksi oleh Balai Besar Perikanan Budidaya Air Tawar Sukabumi, Jawa Barat.
Prosedur Perendaman
Dosis perendaman yang digunakan pada tahap ini adalah 15 mg/L (Subaidah et al. 2012). Jumlah benih udang yang direndam diseragamkan berdasarkan biomassa sekitar 5 mg/L air laut, sehingga jumlah benih udang dalam tiap liter pada setiap stadia adalah adalah 60 ribu ekor naupli, 2250 ekor PL4 dan 400 ekor PL11. Perendaman dilakukan dengan memasukkan hewan uji ke dalam plastik kemas dan diberi oksigen empat kali volume air. Durasi perendaman dilakukan selama dua jam. Suhu air perendaman dijaga stabil pada suhu 25 °C.
Prosedur Penelitian Tahap Pembenihan 1
3
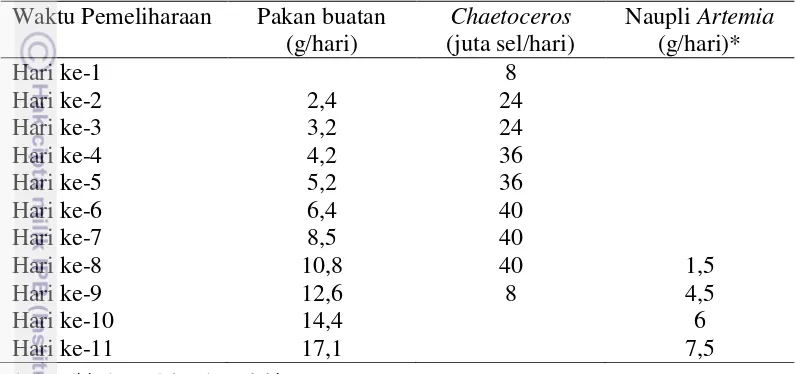
kepadatan 200 ekor/L kemudian diperlihara selama 11 hari. Selama pemeliharaan, benih diberi campuran dari beberapa jenis pakan buatan dengan frekuensi pemberian tujuh kali sehari (pukul 6.00, 10.00, 12.00, 16.00, 18.00, 22.00 dan 24.00), fitoplankton Chaetoceros amami tiga kali sehari (pukul 08.00, 14.00 dan 20.00) dan naupli Artemia sp. tiga kali sehari (pukul 08.00, 14.00 dan 20.00). Jumlah pakan yang diberikan dijabarkan pada Tabel 1. Komposisi dan kadar protein campuran pakan buatan dijabarkan dalam Lampiran 1.
Tabel 1. Total pakan yang diberikan setiap hari pada tahap pembenihan 1 Waktu Pemeliharaan Pakan buatan Chaetoceros Naupli Artemia
(g/hari) (juta sel/hari) (g/hari)*
Hari ke-1 8
* bobot ditimbang dalam bentuk kista
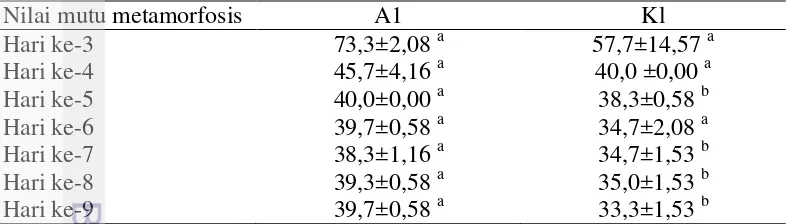
Pengamatan aktivitas benur dilakukan setelah proses perendaman. Aktivitas benur dihitung berdasarkan fototaksis positif dari 240 naupli. Pada hari ke-3 hingga hari ke-9, stadia larva diamati dari 40 ekor larva untuk menghitung nilai mutu metamorfosis. Kuantifikasi nilai mutu metamorfosis dilakukan berdasarkan Tabel 2 dengan stadia baku menurut Wyban dan Sweeney (1991) dan dijelaskan pada Lampiran 2. Nilai mutu metamorfosis menggambarkan percepatan proses metamorfosis pascarendam rElGH. Semakin tinggi nilai mutu metamorfosis (maksimal 80) menunjukkan bahwa proses metamorfosis berlangsung semakin cepat.
Tabel 2. Kuantifikasi nilai mutu metamorfosis udang vaname
Waktu pengamatan Stadia baku* Kriteria penilaian
Hari ke-3 Zoea 1 Naupli = 0; Zoea 1 = 1; Zoea 2 = 2
4
Prosedur Penelitian Tahap Pembenihan 2
Tahap pembenihan 2 merupakan lanjutan dari tahap pembenihan 1. Tahap ini terdiri dari tiga perlakuan dan tiga ulangan. Perlakuan tersebut adalah :
A2 : Perendaman rElGH pada stadia naupli B2 : Perendaman rElGH pada stadia PL4 K2 : Tanpa pemberian rElGH
Pada tahap pembenihan 2, sebanyak 30 ribu ekor benih dari unit perlakuan K1 direndam dengan rElGH sebagai perlakuan B2, 30 ribu ekor benih K1 yang lain direndam tanpa rElGH sebagai perlakuan K2. Pada saat yang sama, 30 ribu ekor benih dari perlakuan A1 direndam tanpa rElGH sebagai perlakuan A2. Setelah perendaman, benih ditebar ke dalam wadah berisi 750 L air laut (salinitas 30 g/L) dan dipelihara selama delapan hari. Selama pemeliharaan, benih udang diberikan campuran pakan buatan tujuh kali sehari (pukul 6.00, 10.00, 12.00, 16.00, 18.00, 22.00 dan 24.00) dan naupli Artemia sp. tiga kali sehari (pukul 08.00, 14.00 dan 20.00). Jumlah pakan yang diberikan dijabarkan pada Tabel 3. Komposisi dan kadar protein campuran pakan buatan dijabarkan pada Lampiran 3. Pergantian air pemeliharaan sebanyak 25% dilakukan setiap pagi hari mulai hari ke-2 hingga hari ke-7. Suhu air selama pemeliharaan berkisar antara 28-30 °C. Tabel 3. Total pakan yang diberikan setiap hari pada tahap pembenihan 2
Waktu pemeliharaan Pakan buatan (g/hari) Naupli Artemia (g/hari)*
Hari ke-1 9,9 9
* bobot ditimbang dalam bentuk kista
Aktivitas benur diamati setelah proses perendaman dan dihitung berdasarkan persentase PL yang aktif bergerak berenang. Panjang benur diukur dari 20 ekor benur udang pada hari ke-2, 4, 6 dan 8. Pada akhir tahap pembenihan, benur dari setiap perlakuan dimasukkan ke dalam plastik kemas selama 2 jam, setelah itu, aktivitas benur diukur dari 100 ekor PL11 serta kelangsungan hidup benur pada uji stres dalam 1 L akuades selama satu jam dan larutan formalin 200 µL/L selama dua jam. Nilai rerata bobot benur diperoleh dari 100 ekor benur udang PL11. Koefisien variasi panjang benur dihitung dari 20 ekor PL11 dengan menggunakan rumus :
Keterangan :
CVL = Koefisien variasi panjang benur (%)
SDL = Simpangan baku panjang 20 ekor PL11 (mm)
�̅L = Rataan panjang 20 ekor PL11 (mm)
� = �̅�
5
Prosedur Penelitian Tahap Pembesaran
Penelitian kemudian dilanjutkan ke tahap pembesaran. Pada tahap ini, terdapat empat perlakuan masing-masing terdiri dari tiga ulangan. Perlakuan tersebut adalah :
A3 : Perendaman rElGH pada stadia naupli B3 : Perendaman rElGH pada stadia PL4 C3 : Perendaman rElGH pada stadia PL11 K3 : Tanpa pemberian rElGH
Pada tahap ini, 100 ekor PL11 dari setiap unit perlakuan ditebar kedalam air laut (salinitas 30 g/L) berkapasitas 750 L dan kemudian dipelihara selama 55 hari. Suhu air selama pemeliharaan berkisar antara 28-30 °C. Pergantian air pemeliharaan sebanyak 50% dilakukan setiap dua hari. Selama pemeliharaan, pemberian pakan dibedakan menjadi dua jenis berdasarkan umur pemeliharaan. Pada awal tahap pembesaran hingga hari ke-25 benur diberikan pakan dengan jumlah yang sama untuk setiap perlakuan (Lampiran 4). Pada hari ke-26 hingga akhir tahap pembesaran, jumlah pakan yang diberikan berdasarkan persentase perkiraan biomassa pada tiap unit perlakuan (Lampiran 5).
Rerata bobot udang dihitung dari 25 ekor tiap unit perlakuan yang ditimbang pada hari ke-0 (PL11), 15, 25, 35, 45 dan 55 tahap pembesaran. Dari data bobot tersebut, laju pertumbuhan mutlak (LPM) dari setiap segmen waktu dihitung berdasarkan Goddard (1996).
Keterangan :
LPM = Laju pertumbuhan mutlak (g/hari) W0 = Bobot udang pada awal (g)
Wt = Bobot udang pada akhir (g) t = Lama pemeliharaan (hari)
Perhitungan koefisien variasi bobot udang, biomassa, tingkat kelangsungan hidup dan rasio konversi pakan dilakukan pada akhir tahap pembesaran. Koefisien variasi bobot udang dihitung dari 25 ekor udang dengan menggunakan rumus :
Keterangan :
CVW = Koefisien variasi bobot udang (%)
SDW = Simpangan baku bobot 25 ekor udang (g)
�̅W = Rataan bobot 25 ekor udang (g)
� = �− � �
� = �̅�
6
Rasio konversi pakan dihitung dengan menggunakan rumus :
Keterangan :
RKP = Rasio konversi pakan
F = Pakan yang diberikan selama tahap pembesaran (g) W0 = Bobot udang pada awal tahap pembesaran (g) Wt = Bobot udang pada akhir tahap pembesaran (g)
Analisis Data
Seluruh data pada tahap pembenihan 1 dianalisis menggunakan uji t, sedangkan data pada tahap pembenihan 2 dan pembesaran dianalisis menggunakan analisis sidik ragam dan jika terdapat perbedaan nyata (P<0,05) maka dilanjutkan dengan uji Tukey dengan bantuan piranti lunak SPSS versi 21.
HASIL DAN PEMBAHASAN
Hasil Tahap Pembenihan 1
Perendaman rElGH pada stadia naupli mempengaruhi nilai mutu metamorfosis (Tabel 4). Semakin tinggi nilai mutu (maksimum 80) menunjukkan bahwa waktu ganti kulit larva (proses metamorfosis) semakin cepat. Pada hari ke-3,4 dan 6, nilai mutu metamorfosis antar perlakuan tidak berbeda yang nyata
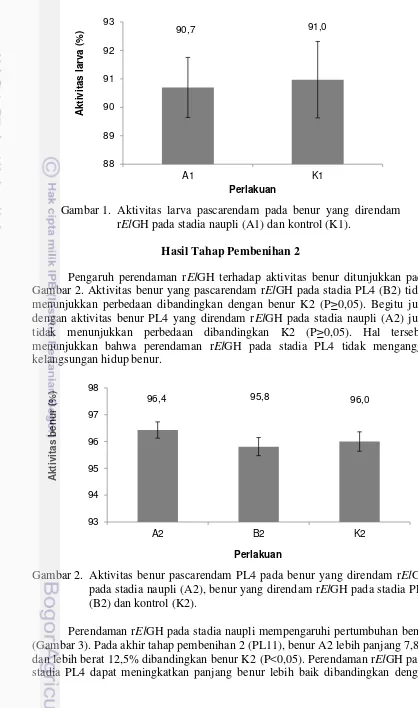
(P≥0,05). Pada hari pengamatan ke-5, 7, 8, dan 9, nilai mutu metamorfosis A1 lebih besar dibandingkan dengan K1 (P<0,05), dengan perbedaan terbesar pada hari ke-9 sebesar 19%. Pada Gambar 1 menunjukkan perendaman rElGH pada stadia naupli tidak mempengaruhi aktivitas larva (P≥0,05), maka perendaman rElGH selama dua jam tidak menggangu kelangsungan hidup naupli.
Tabel 4. Nilai mutu metamorfosis pascarendam rElGH pada stadia naupli
Nilai mutu metamorfosis A1 Kl
Hari ke-3 73,3±2,08 a 57,7±14,57 a
Nilai ditampilkan dalam bentuk rerata±simpangan baku. Huruf superskrip yang berbeda pada baris yang sama menunjukkan nilai berbeda yang nyata (P<0,05). A1: Perendaman rElGH pada stadia naupli. K1: Tanpa pemberian rElGH.
� = �
7
Hasil Tahap Pembenihan 2
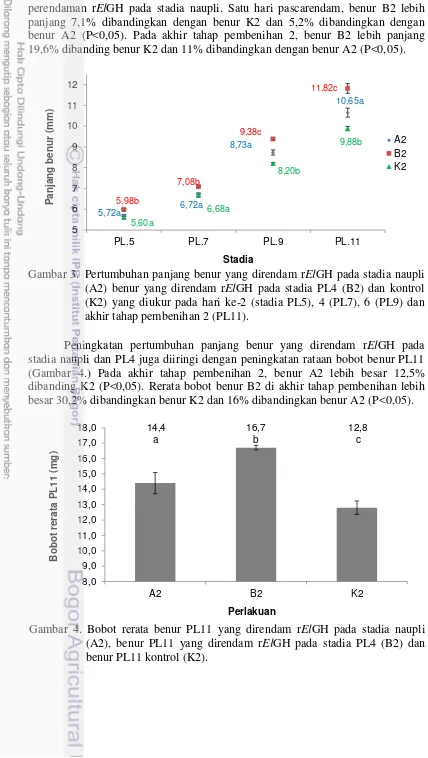
Pengaruh perendaman rElGH terhadap aktivitas benur ditunjukkan pada Gambar 2. Aktivitas benur yang pascarendam rElGH pada stadia PL4 (B2) tidak menunjukkan perbedaan dibandingkan dengan benur K2 (P≥0,05). Begitu juga dengan aktivitas benur PL4 yang direndam rElGH pada stadia naupli (A2) juga tidak menunjukkan perbedaan dibandingkan K2 (P≥0,05). Hal tersebut menunjukkan bahwa perendaman rElGH pada stadia PL4 tidak menganggu kelangsungan hidup benur.
Perendaman rElGH pada stadia naupli mempengaruhi pertumbuhan benur (Gambar 3). Pada akhir tahap pembenihan 2 (PL11), benur A2 lebih panjang 7,8% dan lebih berat 12,5% dibandingkan benur K2 (P<0,05). Perendaman rElGH pada stadia PL4 dapat meningkatkan panjang benur lebih baik dibandingkan dengan
Gambar 1. Aktivitas larva pascarendam pada benur yang direndam rElGH pada stadia naupli (A1) dan kontrol (K1).
8
perendaman rElGH pada stadia naupli. Satu hari pascarendam, benur B2 lebih panjang 7,1% dibandingkan dengan benur K2 dan 5,2% dibandingkan dengan benur A2 (P<0,05). Pada akhir tahap pembenihan 2, benur B2 lebih panjang 19,6% dibanding benur K2 dan 11% dibandingkan dengan benur A2 (P<0,05).
Peningkatan pertumbuhan panjang benur yang direndam rElGH pada stadia naupli dan PL4 juga diiringi dengan peningkatan rataan bobot benur PL11 (Gambar 4.) Pada akhir tahap pembenihan 2, benur A2 lebih besar 12,5% dibanding K2 (P<0,05). Rerata bobot benur B2 di akhir tahap pembenihan lebih besar 30,2% dibandingkan benur K2 dan 16% dibandingkan benur A2 (P<0,05). Gambar 3. Pertumbuhan panjang benur yang direndam rElGH pada stadia naupli
(A2) benur yang direndam rElGH pada stadia PL4 (B2) dan kontrol (K2) yang diukur pada hari ke-2 (stadia PL5), 4 (PL7), 6 (PL9) dan akhir tahap pembenihan 2 (PL11).
9
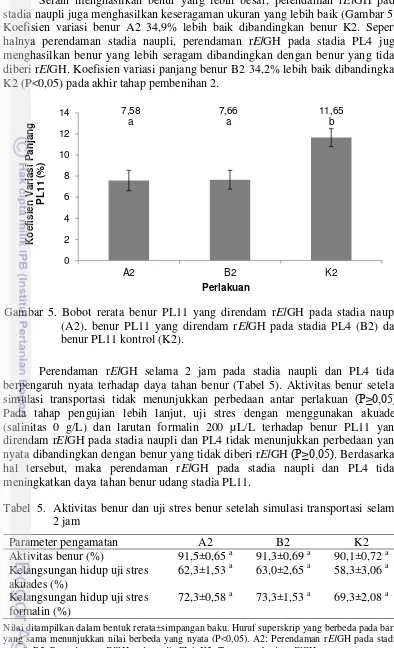
Selain menghasilkan benur yang lebih besar, perendaman rElGH pada stadia naupli juga menghasilkan keseragaman ukuran yang lebih baik (Gambar 5). Koefisien variasi benur A2 34,9% lebih baik dibandingkan benur K2. Seperti halnya perendaman stadia naupli, perendaman rElGH pada stadia PL4 juga menghasilkan benur yang lebih seragam dibandingkan dengan benur yang tidak diberi rElGH. Koefisien variasi panjang benur B2 34,2% lebih baik dibandingkan K2 (P<0,05) pada akhir tahap pembenihan 2.
Perendaman rElGH selama 2 jam pada stadia naupli dan PL4 tidak berpengaruh nyata terhadap daya tahan benur (Tabel 5). Aktivitas benur setelah simulasi transportasi tidak menunjukkan perbedaan antar perlakuan (P≥0,05). Pada tahap pengujian lebih lanjut, uji stres dengan menggunakan akuades (salinitas 0 g/L) dan larutan formalin 200 µL/L terhadap benur PL11 yang direndam rElGH pada stadia naupli dan PL4 tidak menunjukkan perbedaan yang nyata dibandingkan dengan benur yang tidak diberi rElGH (P≥0,05). Berdasarkan hal tersebut, maka perendaman rElGH pada stadia naupli dan PL4 tidak meningkatkan daya tahan benur udang stadia PL11.
Tabel 5. Aktivitas benur dan uji stres benur setelah simulasi transportasi selama 2 jam
Nilai ditampilkan dalam bentuk rerata±simpangan baku. Huruf superskrip yang berbeda pada baris yang sama menunjukkan nilai berbeda yang nyata (P<0,05). A2: Perendaman rElGH pada stadia naupli. B2: Perendaman rElGH pada stadia PL4. K2: Tanpa pemberian rElGH.
10
Hasil Tahap Pembesaran
Hasil pada Tabel 5 menunjukkan bahwa pemberian rElGH pada stadia berbeda akan menghasilkan tingkat pertumbuhan udang yang berbeda. Perlakuan C3 menghasilkan udang dengan bobot 38,2% lebih besar daripada perlakuan K3 (P<0,05) setelah 55 hari masa pembesaran. Perendaman rElGH pada stadia PL4 memang menghasilkan benur PL11 yang paling besar, Akan tetapi pada hari ke-25 hingga hari ke-35, pertumbuhan bobot harian udang B3 tidak berbeda nyata dibandingkan dengan pertumbuhan harian udang K3 (P≥0,05). Bobot udang B3 20,1% lebih besar daripada bobot udang K3 pada hari ke-35 tahap pembesaran.
Tingkat kelangsungan hidup udang pada tahap pembesaran dijabarkan pada Tabel 6. Perendaman rElGH pada stadia naupli, PL4 dan PL11 tidak mempengaruhi tingkat kelangsungan hidup udang di tahap pembesaran (P≥0,05). Kombinasi antara tingkat kelangsungan hidup dan bobot rerata akan menghasilkan biomassa (Gambar 7). Perendaman rElGH pada stadia naupli (A3) dan PL4 (B3) juga tidak mempengaruhi biomassa (P≥0,05). Namun, biomassa C3 32,9% lebih berat daripada biomassa K3 (P<0,05). Perendaman rElGH tidak berpengaruh
nyata (P≥0,05) terhadap rasio konversi pakan dan koefisien variasi bobot (Tabel 6).
11
Tabel 6. Pertumbuhan harian, tingkat kelangsungan hidup, tingkat konversi pakan dan koefisien variasi bobot udang pada tahap pembesaran
Parameter A3 B3 C3 K3
Nilai ditampilkan dalam bentuk rerata±simpangan baku. Huruf superskrip yang berbeda pada baris yang sama menunjukkan nilai berbeda yang nyata (P<0,05). A3: Perendaman rElGH pada stadia naupli. B3: Perendaman rElGH pada stadia PL4. C3: Perendaman rElGH pada stadia PL11. K3: Tanpa pemberian rElGH. LPM x-y: Laju pertumbuhan mutlak dari hari ke-x hingga hari ke-y.
Pembahasan
Pemberian rElGH pada stadia naupli dapat mempercepat waktu ganti kulit. Pertumbuhan krustase khususnya udang tidak kontinu karena kulit luar yang keras, sehingga ganti kulit secara periodik berhubungan dengan pertumbuhan (Keller 1992). Ganti kulit dikontrol oleh hormon ecdysteroid dari organ-Y dalam cephalothorax dan dihambat oleh molt inhibiting hormon (MIH) dari kelenjar organ-X (Chen et al. 2007, Tangprasititipap et al. 2010). Pelepasan MIH terbatas Gambar 7. Biomassa udang yang direndam rElGH pada stadia naupli (A3), udang yang direndam rElGH pada stadia PL4 (B3), udang yang direndam rElGH pada stadi PL11 (C3) dan kontrol (K3) pada tahap pembesaran.
12
pada tahap intermolt. Pada tahap premolt, pelepasan MIH menurun sehingga menyebabkan peningkatan sintesis ecdysteroid dan akhirnya berlanjut ke tahap molting (Chung dan Webster 2005). Menurut Subaidah et al. (2012), ekspresi gen MIH menurun pascarendam dengan rElGH, yang berarti penghambatan terhadap ganti kulit dapat dicegah, atau ganti kulit berjalan efektif dan pertumbuhan meningkat.
Laju pertumbuhan panjang pada tahap pembenihan meningkat akibat pemberian rElGH. Benur yang direndam rElGH pada stadia naupli dan PL4 lebih panjang dibanding kontrol. Benur yang direndam rElGH pada stadia PL4 bahkan berkembang pesat 24 jam pascarendam. Moriyama dan Kawauchi (1990) pada penelitian sebelumnya menunjukkan bahwa bobot benih ikan salmon segera meningkat setelah perendaman dengan larutan rGH, dan perbedaan bobot tubuh mulai terlihat nyata 14 jam pascarendam. Bobot benur yang direndam rElGH pada stadia PL4 dan bobot benur yang direndam rElGH pada stadia naupli dan bobot benur yang direndam rElGH pada stadia naupli dan bobot benur yang direndam rElGH pada stadia naupli lebih besar dibandingkan kontrol.
Pada unit pembenihan, benur udang akan dijual jika sudah mencapai panjang 1 cm. Berdasarkan hasil pada penelitian ini, benur yang direndam rElGH pada stadia PL4 akan mencapai ukuran jual dua hari lebih cepat dibandingkan benur yang tidak diberi rElGH, sehingga mengurangi biaya produksi. Pemberian rElGH juga menghasilkan benur dengan keseragaman ukuran yang lebih baik, walaupun kualitas daya tahan benur tidak berbeda dengan benur yang tidak diberi rElGH. Perbaikan kualitas ukuran tersebut akan mempengaruhi daya saing benur ketika dipasarkan.
13
dipengaruhi oleh GH. Selanjutnya, ekspresi SIBD terbukti meningkat pada udang vaname yang diberi perlakuan rElGH (Subaidah et al. 2012).
Perendaman rElGH pada stadia yang berbeda diduga juga mempengaruhi waktu efektivitas rElGH. Pemberian rElGH 15 mg/L pada stadia naupli hanya efektif meningkatkan laju pertumbuhan pada tahap pembenihan (19 hari) dan tidak efektif meningkatkan laju pertumbuhan udang skala usaha pembesaran. Pemberian rElGH pada stadia PL4 efektif meningkatkan laju pertumbuhan udang hingga hari ke-25 tahap pembesaran (32 hari pascarendam). Perendaman rElGH pada stadia PL11 efektif meningkatkan laju pertumbuhan hingga hari ke-45 pascarendam. Hal tersebut menunjukkan bahwa perendaman rElGH pada udang hanya efektif maksimal 45 hari pascarendam. Pada penelitian Moriyama dan Kawauchi (1990), laju pertumbuhan ikan salmon yang direndam rGH lebih cepat daripada kontrol hingga 85 hari pascarendam. Perbedaan tersebut dapat disebabkan perbedaan ikan resepien. Hal ini didukung dengan hasil penelitian Alimuddin et al. (2014) yang menunjukkan ikan sidat yang direndam rElGH kemudian dilanjutkan dengan pemberian rElGH melalui pakan 60 hari pascarendam tumbuh lebih baik dari pada ikan sidat yang hanya direndam rElGH.
Perendaman rElGH tidak mempengaruhi rasio konversi pakan. Hal yang sama terjadi pada penelitian Subaidah (2013), perendaman rElGH 15 mg/L selama tiga jam tidak mempengaruhi rasio konversi pakan udang yang diperlihara selama 72 hari pascarendam. Namun demikian hasil yang dilaporkan pada udang berbeda dengan yang dilaporkan pada ikan. Sebagai contoh, pemberian rElGH meningkatkan efisiensi pakan atau menurunkan rasio konversi pakan pada ikan sidat (Handoyo et al. 2012).
Pada tahap pembenihan, pengujian aktivitas benur dan uji stres di akhir tahap pembenihan menunjukkan bahwa perendaman rElGH 15 mg/L selama dua jam tidak memberikan pengaruh terhadap daya tahan benur. Pada tahap pembesaran, tingkat kelangsungan hidup pada penelitian ini tidak berbeda nyata dibandingkan kontrol. Pemberian rGH dapat meningkatkan kelangsungan hidup ikan (Acosta et al. 2009). Namun pemberian rGH tidak meningkatkan kelangsungan hidup udang seperti yang dilaporkan oleh Santiesteban et al. (2010) dan Subaidah (2013).
14
KESIMPULAN
Pemberian rElGH dapat meningkatkan pertumbuhan udang vaname. Untuk meningkatkan dan menyeragamkan ukuran benur pada tahap pembenihan dapat menerapkan perendaman rElGH pada stadia PL4. Untuk meningkatkan rataan bobot tubuh dan biomassa udang pada tahap pembesaran dapat menerapkan perendaman rElGH pada stadia PL11.
DAFTAR PUSTAKA
Acosta J, Estrada MP, Carpio Y, Ruiz O, Morales R, Martinez E, Valdes J, Borroto C, Besada V, Sanchez A, Herrera F. 2009. Tilapia somatotropin polypeptides: potent enhacers of fish growth and innate immunity. Biotecnologia Aplicada 26:267-272.
Acosta J, Morales R, Morales A, Alonso M, Estrasa MP. 2007. Pichia pastoris expressing recombinant tilapia growth hormone accelerates the growth of tilapia. Biotechnology Letters 29:1671-1676.
Alimuddin, Handoyo B dan Utomo NBP. 2014. Efektivitas pemberian hormon pertumbuhan rekombinan ikan kerapu kertang Epinephelus lanceolatus melalui perendaman dan oral terhadap pertumbuhan elver ikan sidat Anguilla bicolor bicolor. Jurnal Iktiologi Indonesia 14: 179-189.
Alimuddin, Lesmana I, Sudrajat AO, Carman O, Faisal I. 2010. Production and bioactivity potential of three recombinant growth hormones of farmed fish. Indonesian Aquaculture Journal 5:11-16.
Carpio Y, Leon K, Acosta J, Morales R, Estrada MP. 2007. Recombinant tilapia neuropeptide Y promotes growth and antioxidant defenses in African catfish Clarias gariepinus fry. Aquaculture 272:649-655.
Castellanos M, Jimenez-Vega F, Vargas-Albores F. 2008. Single IB Domain (SIBD) protein from Litopenaeus vannamei, a novel member for the IGFBP family. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics 3:270-274.
Chen HY, Watson RD, Chen JC, Liu HF, Lee CY. 2007. Molecular characterization and gene expression pattern of two putative molt-inhibiting hormones from Litopenaeus vannamei. General and Comparative Endocrinology 151:72-81.
Chung JS, Webster SG. 2005. Dynamics of in vivo release of molt-inhibiting hormone and crustacean hyperglycemic hormone in the shore crab, Carcinus maenas. Endocrinology 146:5545-5551.
[FAO] Food and Agriculture Organization of the United Nations. 2003. Health management and biosecurity maintenance in white shrimp Penaeus vannamei hatcheries in Latin America. FAO Fisheries Technical Paper. Funkenstein B, Dyman A, Lapidot Z, Jesus-Ayson EGD, Gertler A, Ayson FG.
2005. Expression and purification of a biologically active recombinant rabbitfish Siganus guttastus growth hormone. Aquaculture 250:504-515. Goddard S. 1996. Feed management in intensive aquaculture. New York (US):
15
Haghighi M, Rohani MS, Sharifpour I, Sepahdari A, Aghaee GRL. 2011. Oral recombinant bovine somatotropin improves growth performance in rainbow trout Oncorhynchus mykiss. Iranian Journal of Fisheries Sciences 10:415-424.
Handoyo B, Alimuddin, Utomo NBP. 2012. Pertumbuhan, konversi dan retensi pakan, dan proksimat tubuh benih ikan sidat yang diberi hormon pertumbuhan rekombinan ikan kerapu kertang melalui perendaman. Jurnal Akuakultur Indonesia 11:132-140.
Hill JE, Baldwin JD, Graves JS, Leonard R, Powell JFF, Watson CA. 2005. Preliminary observations of topical gill application of reproduction hormones for induced spawning of tropical ornamental fish. North American Journal of Aquaculture 67:7-9.
Keller R. 1992. Crustacea neuropeptides: structures, function and comparative aspects. Experimentia 48:439-448.
Leedom TA, Uchida K, Yada T, Richman H III, Byatt JC, Collier RJ, Hirano T, Grau EG. 2002. Recombinant bovine growth hormone treatment of tilapia: growth response, metabolic clearence, receptor binding and immunoglobulin production. Aquaculture 207:359-380.
Liao WL, Lu HH, Huang SK, Wo JL, Huang JH, Lin EC. 2008. Study of growth and body composition of red snapper Lutjanus erythropterus fed diets containing Escherichia coli expressing recombinant tilapia insulin-like growth factor-1. Fisheries Science 74:354-361.
Moriyama S, Furukawa S, Kawauchi H. 2009. Growth stimulation of juvenile abalone Haliotis discus hannai by feeding with salmon growth hormone in sodium alginate gel. Fisheries Science 75:689-695.
Moriyama S, Kawauchi H. 1990. Growth stimulation of juvenile salmonids by immersion in recombinant salmon growth hormone. Nippon Suisan Gakkaishi 56:31-34.
Moriyama S, Kawauchi H. 2004. Somatic growth acceleration of juvenile abalone Haliotis discus hannai, by immersion in and intramuscular injection of recombinant salmon growth hormone. Aquaculture 229:469-478.
Ohlsson C, Mohan S, Sjogren K, Tivesten A, Isgaard J, Isaksson O, Jansson JO, Svensson J. 2009. The role of liver-derived insulin-like growth factor-1 [ulasan]. Endocrine Reviews 30:494-535.
Promdonkoy B, Warit S, Panyim S. 2004. Production of a biologically active growth hormone from giant catfish Pangasianodon gigas in Escherichia coli. Biotechnology Letters 26:649-653.
Santiesteban D, Martin L, Arenal A, Franco R, Sotolongo J. 2010. Tilapia growth hormone binds to a receptor in brush border membrane vesicles from the hepatopancreas of shrimp Litopenaeus vannamei. Aquaculture 306:338-342.
Sherwood NM, Harvey B. 1986. Topical absorption of gonadotropin-releasing hormone (GnRH) in goldfish. General and Comparative Endocrinology 61:13-19.
16
Subaidah S, Carman O, Sumantadinata K, Sukenda, Alimuddin. 2012. Respons pertumbuhan dan ekspresi gen udang vaname, Litopenaeus vannamei setelah direndam dalam larutan hormon pertumbuhan rekombinan ikan kerapu kertang. Jurnal Riset Akuakultur 7:359-369.
Subaidah S. 2013. Respons pertumbuhan dan imunitas udang vaname Litopenaeus vannamei terhadap pemberian hormon pertumbuhan rekombinan ikan kerapu kertang. [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Tangprasitittipap A. Tiensuwan M, Withyachumnarnkul B. 2010. Characterization of candidate genes involved in growth of black tiger shrimp Penaeus monodon. Aquaculture 307:150-156.
17
18
Lampiran 1. Komposisi dan kadar protein pakan buatan yang digunakan pada tahap pembenihan 1
Jenis pakan Waktu pemeliharaan (hari ke-)
2 3 4 5 6 7 8 9 10 11
Pakan buatan (g/hari)
Tepung spirulina
0,6 0,65 0,7 0,75 0,8 0,85 0,9 0,9 CAR 1,8 1,9 2,1 1,45
CD 1,5 2,45 3,4 2,7
MPL 1,8 4,5 5,4 6,3
PL150 0,45
Z1 0,65 0,7 0,75 0,8
Z2 0,4 1,3 1,8 2,25
Z3 2,25 2,7
MPz 0,7 0,75 0,8 0,85
MP1 0,85 2,25 3,6
MP2 5,4 6,3
Flake 0,8 0,85 0,9 0,9 0,9 0,9
Vitamin mix 0,35 0,4 0,45 0,45 0,45 0,45
19
Lampiran 2. Metamorfosis udang menurut Wyban dan Sweeney (1991) beserta foto pengamatan
Naupli 5
Tiap furcal memiliki 7 duri
Zoea 1
Tubuh terbagi 2 bagian besar Mata mulai terlihat
Saluran pencernaan mulai terlihat
Zoea 2
Mata menonjol keluar karapas Rostrum terbentuk
Zoea 3
Uropod terbentuk
Pada somit abdominal terbentuk duri
Mysis 1
Tunas pleopod muncul Telson mulai berkembang Arah gerak renang mundur
Mysis 2
Pleopod terbentuk tapi belum bersegmen
Ujung Telson menyempit
Mysis 3
Pleopod semakin berkembang Ujung Telson semakin menyempit
dari M2
Muncul duri di rostrum Pascalarva 1
20
Lampiran 3. Komposisi dan kadar protein pakan buatan yang digunakan pada tahap pembenihan 2
Jenis pakan Waktu pemeliharaan (hari ke-)
1 2 3 4 5 6 7
Pakan buatan (g/ha
MPL 3,6 4 4,5
PL150 0,4 0,6 0,9 3,6 0,9
PL300 2,45 5,75 7,45 8,65
MP2 3,6 4,1
MP3 4,5 5,4 6,3 7,2 8,1
Z3 1,6 1,8 2 2,2 2,4 2,6 2,8
Flake 0,45 0,45 0,45
Vitamin mix 0,25 0,25 0,25 0,25 0,25 0,25 0,25 Kadar protein MPL : 48%
21
Lampiran 4. Jumlah dan jenis pakan buatan yang diberikan pada hari ke-1 hingga hari ke-25 tahap pembesaran
Usia pemeliharaan Jenis pakan Jumlah pakan
(gram/hari)
Hari ke-1 serbuk 3,0
Hari ke-2 serbuk 3,4
Hari ke-3 serbuk 3,8
Hari ke-4 serbuk 4,2
Hari ke-5 serbuk 4,6
Hari ke-6 serbuk 5,0
Hari ke-7 serbuk 5,5
Hari ke-8 serbuk 6,0
Hari ke-9 serbuk 6,5
Hari ke-10 serbuk 7,0
Hari ke-11 serbuk (50%) & crumble (50%) 7,5 Hari ke-12 serbuk (50%) & crumble (50%) 8,0 Hari ke-13 serbuk (50%) & crumble (50%) 8,6 Hari ke-14 serbuk (50%) & crumble (50%) 9,2
Hari ke-15 crumbel 9,8
Hari ke-16 crumbel 10,4
Hari ke-17 crumbel 11,0
Hari ke-18 crumbel 11,8
Hari ke-19 crumbel 12,6
Hari ke-20 crumbel 13,4
Hari ke-21 crumbel 14,2
Hari ke-22 crumbel 15,0
Hari ke-23 crumbel 15,8
Hari ke-24 crumbel 16,6
Hari ke-25 crumbel 17,4
22
Lampiran 5. Jenis pakan dan persentase pemberian pakan hari ke-26 hingga hari ke-55 pada tahap pembesaran
Usia pemeliharaan Jenis pakan Persentase pemberian pakan (% dari perkiraan biomassa)
23
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 5 September 1982 sebagai anak kedua dari tiga bersaudara oleh pasangan Syafnir Zakir SE dan Dra. Khusdiningsih. Penulis menyelesaikan pendidikan sekolah menengah atas di SMU Negeri 91 Jakarta Timur tahun 2000. Pada tahun 2001 penulis lulus Ujian Masuk Perguruan Tinggi Negeri (UMPTN) pada Program Studi Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Pendidikan sarjana penulis selesaikan tahun 2007. Pada tahun 2012, penulis mendapatkan kesempatan untuk menempuh pendidikan magister ke Sekolah Pascasarjana IPB, program studi Ilmu Akuakultur.