HIDROLISAT PROTEIN TEMPE KOMAK ((
Lablab purpureus
(L.) Sweet) SEBAGAI PENGHAMBAT ACE (
ANGIOTENSIN
CONVERTING ENZYME)
WIWIK R JULIANA SINAGA
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Hidrolisat Protein Tempe Komak ((Lablab pupureus (L.) Sweet) Sebagai Penghambat ACE (Angiotensin Converting Enzyme) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Penelitian ini merupakan bagian dari kegiatan penelitian Teknologi Pengolahan Kacang-kacangan sebagai Sumber Protein untuk Substitusi Impor Kedelai dengan penanggungjawab Dr. Endang Yuli Purwani M.Si. Penelitian ini didanai oleh DIPA BB Pascapanen 2013 dengan nomor DOK-INT-RE-3.2/032/2013. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2014
Wiwik R Juliana Sinaga
ABSTRAK
WIWIK R JULIANA SINAGA. Hidrolisat Protein Tempe Komak ((Lablab purpureus (L.) Sweet) sebagai Penghambat ACE (Angiotensin Converting Enzyme). Dibimbing oleh WARAS NURCHOLIS dan ENDANG YULI PURWANI.
Ketidakseimbangan antara produksi dan konsumsi kedelai nasional menyebabkan ketergantungan terhadap impor kedelai meningkat. Alternatif untuk mengatasi hal tersebut adalah memanfaatkan sumber protein nabati lain sebagai substitusi kedelai, salah satunya adalah kacang komak (Lablab purpureus (L.) Sweet). Tujuan penelitian ini adalah menentukan aktivitas penghambatan terhadap ACE (Angiotensin Converting Enzyme) dari hidrolisat protein kacang komak sebagai substitusi kedelai untuk bahan baku tempe. Hidrolisat protein dari kacang komak diteliti aktivitas penghambatan terhadap ACE secara in vitro dengan 3 perlakuan, yaitu hidrolisat kacang komak, hidrolisat tempe komak mentah dan hidrolisat tempe komak kukus. Sebagai pembanding dari aktivitas penghambatan terhadap ACE dari kacang komak, digunakan kedelai dengan analisis yang sama dengan kacang komak. Hasil penelitian menunjukkan bahwa hidrolisat protein tempe komak mentah memiliki aktivitas penghambatan ACE yang lebih besar dibandingkan dengan kedelai. Nilai IC50 tempe komak mentah dan tempe kedelai secara berurutan adalah 0.64 mg/g dan 1.69 mg/g.
Kata Kunci : Angiotensin converting enzyme, kacang komak, tempe.
ABSTRACT
WIWIK R JULIANA SINAGA. Protein Hydrolyzate of Komak Tempeh ((Lablab purpureus (L.) Sweet) as ACE (Angiotensin Converting Enzyme) Inhibitor. Supervised by WARAS NURCHOLIS and ENDANG YULI PURWANI.
Unbalanced condition of local soybean production and consumption causes the increasing dependence through import soybean. The alternative to overcome this problem is to utilize another vegetable protein source as soybean substitution is needed. One of those protein is the komak nut toward ACE. The obstruction activity of komak nut hydrolyzate was analyzed in vitro involving three treatments, they were hydrolyzates of whole grain of komak tempeh, hydrolyzates of unpride komak tempeh, and hydrolyzates of steamed komak tempeh. Soybean analysis was also done due to compare the obstruction activity toward ACE with the same analysis done for komak nut. The result showed that unpride komak tempeh was more potential as ACE inhibitor compared to soybean. The IC50 value of unpride komak tempeh and soybean were 0.64 mg/g and 1.69 mg/g.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biokimia
HIDROLISAT PROTEIN TEMPE KOMAK ((
Lablab purpureus
(L.) Sweet) SEBAGAI PENGHAMBAT ACE (
ANGIOTENSIN
CONVERTING ENZYME)
WIWIK R JULIANA SINAGA
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Hidrolisat Protein Tempe Komak ((Lablab purpureus (L.) Sweet) sebagai Penghambat ACE (Angiotensin Converting Enzyme)
Nama : Wiwik R Juliana Sinaga NIM : G84090069
Disetujui oleh
Waras Nurcholis, SSi, MSi Pembimbing I
Dr Endang Yuli P, MSi Pembimbing II
Diketahui oleh
Dr Ir I Made Artika, MAppSc Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan April 2013 sampai September 2013 ini ialah diversifikasi pangan, dengan judul Hidrolisat Protein Tempe Komak ((Lablab purpureus (L.) Sweet) Sebagai Penghambat ACE (Angiotensin Converting Enzyme).
Terima kasih penulis ucapkan kepada BB Pasca Panen yang telah mendanai penelitian ini serta Waras Nurcholis S.Si, M.Si dan Dr. Endang Yuli P M.Si selaku pembimbing yang telah banyak memberi saran. Disamping itu, penghargaan penulis sampaikan kepada Bapak Tri, Ibu Ika, Ibu Mely, Bapak Yudi, Irman beserta staf dan teknisi lain BB Pasca Panen yang telah membantu selama pengumpulan data. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga, atas doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2014
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
METODE 2
Waktu dan Tempat Penelitian 2
Bahan 2
Alat 2
Prosedur Analisis Penelitian 3
HASIL 7
Tempe Komak 7
Kadar Proksimat 7
Derajat Hidrolisis 10
Kadar Protein Terlarut 10
Aktivitas Penghambatan ACE 11
PEMBAHASAN 14
Tempe Komak 14
Persentase Proksimat 14
Persentase Derajat Hidrolisis 15
Kadar Protein Terlarut 16
Aktivitas Penghambatan ACE 17
SIMPULAN DAN SARAN 18
Simpulan 18
Saran 19
DAFTAR PUSTAKA 19
LAMPIRAN 22
DAFTAR TABEL
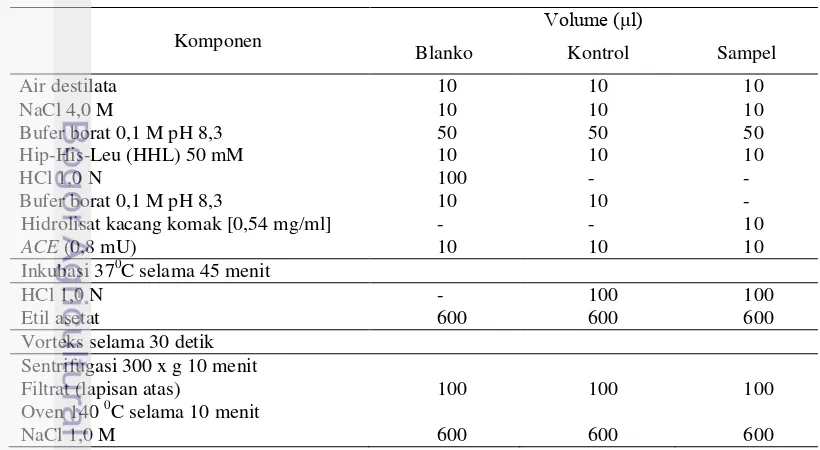
1 Komposisi Bahan Analisis Penghambatan ACE 6
2 Derajat Hidrolisis dari Hidrolisat Protein Komak dan Kedelai 10 3 Kadar Protein Terlarut dari Hidrolisat Protein Komak dan Kedelai 11 4 Nilai IC50 dari Hidrolisat Protein Komak dan Kedelai 13
DAFTAR GAMBAR
1 Kacang Komak dan Tempe Komak 7
2 Hasil Analisis Kadar Air 8
3 Hasil Analisis kadar Abu 8
4 Hasil Analisis Kadar Lemak 9
5 Hasil Analisis Kadar Protein 9
6 Hasil Analisis Kadar Karbohidrat 10
7 Aktivitas Penghambatan ACE dari Hidrolisat Protein Kacang Komak
dan Kedelai 11
8 Aktivitas Penghambatan ACE dari Hidrolisat Protein Tempe Komak
Mentah dan Tempe Kedelai Mentah 12
9 Aktivitas Penghambatan ACE dari Hidrolisat Protein Tempe Komak
Kukus dan Tempe Kedelai Kukus 12
10 Aktivitas Penghambatan ACE dari Kaptropil 13
DAFTAR LAMPIRAN
1 Diagram alir penelitian 22
2 Tabel dan gambar hasil pengamatan absorbansi pada konsentrasi
standar BSA yang berbeda 23
3 Tabel dan gambar hasil pengamatan absorbansi pada konsentrasi
standar L-Leusin yang berbeda 24
4 Analisis kadar air 25
5 Analisis kadar abu 26
6 Analisis kadar lemak 27
7 Analisis kadar protein 28
8 Analisis kadar karbohidrat 29
9 Derajat Hidrolisis 30
10 Analisis ProteinTerlarut 32
11 Korelasi antara kosentrasi hidrolisat protein sampel dengan aktivitas
penghambatan ACE 33
PENDAHULUAN
Indonesia merupakan negara yang kaya sumber hayati, salah satunya adalah beragam jenis kacang-kacangan. Kacang-kacangan merupakan sumber protein kompleks, dapat diolah menjadi tempe, sumber protein hidrolisat. Di Indonesia, bahan baku utama tempe ialah kedelai (Syarief et al. 1999). Berdasarkan data dari Departemen Pertanian RI pada tahun 2005, tingkat konsumsi kedelai per tahun 2004 adalah 4.186.157 ton, sedangkan kedelai yang tersedia adalah 1.311.196 ton, sehingga menyebabkan impor sebesar 2.874.961 ton. Sampai dengan tahun 2011 produksi kedelai nasional masih mengalami penurunan. Produksi kedelai tahun 2011 sebesar 851.29 ribu ton biji kering turun sebesar 55.74 ribu ton (sekitar 6.15%) dibandingkan produksi tahun 2010 (BPS 2012).
Kacang komak direkomendasikan sebagai sumber protein fungsional alternatif pengganti kedelai karena memiliki kandungan protein cukup tinggi 18-25% (Subagio 2006). Sifat fungsional isolat protein kacang komak dan kacang kedelai memiliki banyak kesamaan yaitu daya serap air, daya serap minyak, dan daya emulsi isolat yang tidak berbeda nyata (Suwarno 2003). Kacang komak juga memiliki susunan asam amino yang mendekati pola protein kedelai, yaitu kekurangan asam amino bersulfur (metionin dan sistein) dan kaya akan asam amino lisin (Kay 1979). Pola elektroforesis fraksi protein globulin 7S dan 11S kacang komak hampir sama dengan pola elektroforesis kacang kedelai (Khodijah 2003).
Tempe merupakan pangan tradisional Indonesia yang dibuat dengan proses fermentasi menggunakan kultur murni Rhizopus sp. (Rhizopus oryzae atau
Rhizopus oligosporus) atau kultur campuran dalam bentuk ragi tempe sebagai inokulan (Handajani 2001). Selain mudah ditemui dan harganya murah, tempe memiliki kandungan gizi yang baik untuk tubuh. Penggunaan tempe kacang komak sebagai bahan baku pembuatan hidrolisat protein dilakukan untuk meningkatkan daya cerna proteinnya. Hal ini dimungkinkan dengan adanya aktivitas degradasi protein oleh kapang tempe selama proses fermentasi berlangsung (Alvina 2009). Selanjutnya, hidrolisat protein kacang komak ini dibandingkan dengan karakteristik hidrolisat protein kacang kedelai sehingga dapat diketahui secara pasti peranan dan fungsinya dalam pengolahan pangan sebagai alternatif substitusi hidrolisat protein kedelai.
2
Angiotensin Converting Enzyme (ACE) sudah lama dikenal sebagai bagian kunci pada sistem renin angiotensin yang penting pada pengaturan tekanan darah. Jenis peptida ini pada dasarnya mengatalisis reaksi angiotensin I menjadi angiotensin II dengan memecah dipeptida histidil-leusina dari ujung-C angiotensin I menghasilkan angiotensin II dan asam hipurat (Actis et al. 2003). Jika hidrolisis angiotensin I berlebihan, maka tekanan darah akan meningkat. Salah satu obat yang digunakan untuk mengendalikan tekanan darah adalah ACE-inhibitor. ACE-inhibitor bekerja dengan menghambat efek angiotensin II yang bersifat vasokonstriktor. Dengan adanya peptida yang bersifat sebagai ACE inhibitor maka kerja ACE dalam mengubah angiotensin I menjadi angiotensin II akan terganggu sehingga tekanan darah dapat diturunkan (Astawan et al. 2003). Sampai saat ini belum ada yang melaporkan kemampuan kacang komak sebagai ACE inhibitor seperti kedelai. Oleh sebab itu, penelitian ini dilakukan untuk menentukan aktivitas penghambatan ACE (Angiotensin Converting Enzyme) dari hidrolisat protein kacang komak yang digunakan sebagai substitusi kedelai untuk bahan baku tempe.
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan selama 6 bulan mulai April sampai September 2013. Tempat pelaksanaannya di Laboratorium Kimia dan Laboratorium Proksimat, BB Pasca Panen; Laboratorium Balai Tanah dan Laboratorium Biologi Molekuler, BB Biogen.
Bahan
Bahan yang digunakan dalam penelitian adalah kacang komak varietas Bali, sebagai pembanding dari komak digunakan kacang kedelai varietas Grobogan, dan laru tempe RAPRIMA produksi PT. Aneka Fermentasi Industri Bandung. Enzim yang digunakan ialah α-Amilase (A 6380), pepsin (P 6887), tripsin (T 0303), α-kemotripsin (C 4129), ACE (A 6778), dan Hippuril-His-Leu (HHL) (H 1635). Bahan kimia lain yang digunakan adalah HCl 1 N, etil asetat, NaCl 4.0 M, buffer borat 0.1 M pH 8.3, buffer glisin 0.01 M pH 7.0,
2.4.6-trinitrobenzenesulphonic acid (TNBS) (P 2297), L-Leusin, buffer fosfat 0.2 M pH 8.2, sodium sulfit 0.1 M, CBB (coomassie brilliant blue) G-250, kaptropil, etanol 95%, asam fosfat 85%, BSA (bovine serum albumin).
Alat
Alat yang digunakan dalam penelitian ini adalah proofer untuk fermentasi tempe, sentrifuse Beckman Coulter Avanti J-E, spektrofotometer UV 6500 Kruss Germany, spektrofotometer UV/VIS spectroquant Pharo 300, sentrifuse Tomy
3 tabung reaksi, mikropipet, waterbath, gelas piala, tabung eppendorf, pipet ukur, tabung vial, botol plastik, styrofoam, blender dan stirer.
Prosedur Analisis Penelitian
Pembuatan Tempe Kacang Komak
Proses pembuatan tempe komak dilakukan dengan modifikasi dari metode van den Hil dan Nout (2011). Proses diawali dengan perendaman komak dalam air selama 16 jam dan pengeringan dengan oven pengering bersuhu 70oC selama 24 jam. Komak kering dikupas dengan pengupas mekanis untuk memisahkan biji dan kulit. Komak kering tanpa kulit direbus dalam air mendidih selama 15 menit dan direndam selama 50 jam dalam air dengan perbandingan komak kering tanpa kulit dan air 1:4. Komak selanjutnya dicuci, dikukus selama 10 menit, ditiriskan, dan didinginkan pada suhu ruang (29 sampai 31oC). Komak diinokulasikan dengan 0.1 % laru (0.1 gram laru untuk 100 gram komak). Komak dikemas dalam plastik PP yang telah dilubangi dengan jarak 2 cm dan diinkubasi dalam fermentor bersuhu 35oC ±1oC selama 24 sampai 36 jam.
Pembuatan Tempe Kedelai
Proses pembuatan tempe kedelai dilakukan dengan modifikasi dari metode van den Hil dan Nout (2011). Proses diawali dengan perebusan kedelai dalam air mendidih selama 40 menit dan direndam selama 30 jam dalam air dengan perbandingan keledai utuh dan air 1:4. Kedelai dikupas dan dicuci bersih. Kedelai tanpa kulit yang sudah bersih dikukus selama 30 menit, ditiriskan, dan didinginkan pada suhu ruang (29 sampai 31oC). Kedelai diinokulasikan dengan 0.1 % laru (0.1 gram laru untuk 100 gram kedelai). Kedelai dikemas dalam plastik PP yang telah dilubangi dengan jarak 2 cm dan diinkubasi dalam fermentor bersuhu 35oC ±1oC selama 24 sampai 36 jam.
Analisis Proksimat
Analisis proksimat dilakukan melalui beberapa tahapan, meliputi preparasi sampel, penentuan kadar air, penentuan kadar abu, penentuan kadar lemak, penentuan kadar protein, dan penentuan kadar karbohidrat. Sampel terdiri dari 3 perlakuan yaitu kacang komak, tempe komak mentah dan tempe komak kukus. Sebagai pembanding dari kacang komak digunakan kedelai dengan perlakuan yang sama dengan komak, yaitu kacang kedelai, tempe kedelai mentah, dan tempe kedelai kukus. Sampel diblender hingga halus dan rata.
Penentuan kadar air dilakukan dengan modifikasi dari metode AOAC (2005). Cawan yang akan digunakan untuk mengukur bobot sampel dioven terlebih dahulu selama 30 menit pada suhu 100-105oC, kemudian didinginkan dalam desikator untuk menghilangkan uap air dan ditimbang. Sampel ditimbang sebanyak 2 gram dalam cawan yang sudah dikeringkan kemudian dioven pada suhu 100-105oC selama 6 jam lalu didinginkan dalam desikator selama 30 menit dan ditimbang. Tahap ini diulangi hingga dicapai bobot yang konstan. Kadar air dihitung dengan rumus:
4 Cawan porselin yang akan digunakan untuk mengukur bobot sampel, dikeringkan menggunakan oven selama 30 menit pada suhu 100-105 oC. Cawan yang sudah dikeringkan tersebut kemudian didinginkan dalam desikator untuk menghilangkan uap air dan ditimbang. Sampel ditimbang sebanyak 2 gram dalam cawan yang sudah dikeringkan kemudian dibakar di atas nyala pembakar sampai tidak berasap dan dilanjutkan dengan pengabuan di dalam tanur pada suhu 550-600oC sampai pengabuan sempurna. Dinginkan sampel beserta cawan porselin dalam desikator dan timbang bobotnya. Tahap pembakaran dalam tanur diulangi sampai didapat bobot yang konstan. Kadar abu dihitung dengan rumus:
00
Penentuan kadar lemak dilakukan sesuai dengan metode AOAC (2005). Labu lemak yang akan digunakan dioven selama 30 menit pada suhu 100-105oC, kemudian didinginkan dalam desikator untuk menghilangkan uap air dan ditimbang. Sampel yang akan diukur kadar lemaknya ditimbang sebanyak 2 gram lalu dibungkus dengan kertas saring, ditutup dengan kapas bebas lemak dan dimasukkan ke dalam alat ekstraksi sokhlet yang telah dihubungkan dengan labu lemak yang telah dioven dan diketahui bobotnya. Pelarut heksan dituangkan sampai sampel terendam dan dilakukan refluks atau ekstraksi lemak selama 5-6 jam atau sampai pelarut lemak yang turun ke labu lemak berwarna jernih. Pelarut lemak yang telah digunakan, disuling dan ditampung setelah itu ekstrak lemak yang ada dalam labu lemak dikeringkan dalam oven bersuhu 100-105oC selama 1 jam, lalu labu lemak didinginkan dalam desikator dan ditimbang. Tahap pengeringan labu lemak diulangi sampai diperoleh bobot yang konstan. Kadar lemak dihitung dengan menggunakan persamaan berikut:
00
Keterangan:
5 Penentuan kadar protein total dilakukan sesuai dengan metode AOAC (2005). Penentuan kadar protein total dilakukan dengan metode Kjeldahl. Sampel yang akan diuji ditimbang sebanyak 0.1-0.5 gram dimasukkan ke dalam labu kjeldahl 100 ml, ditambahkan dengan ¼ buah tablet kjedahl, kemudian didekstruksi (pemanasan dalam keadaan mendidih) sampai larutan menjadi hijau jernih dan SO2 hilang. Larutan dibiarkan dingin dan dipindahkan ke labu 50 ml dan diencerkan dengan akuades sampai tanda tera, dimasukkan ke dalam alat destilasi, ditambahkan dengan 5-10 ml NaOH 30-33% dan dilakukan destilasi. Destilat ditampung dalam larutan 10 ml asam borat 3% dan beberapa tetes indikator (larutan bromcresol green 0.1% dan larutan metil merah 0.1% dalam alkohol 95% secara terpisah dan dicampurkan antara 10 ml bromcresol green
dengan 2 ml metil merah) kemudian dititrasi dengan larutan HCl 0.02 N sampai larutan berubah warnanya menjadi merah muda. Kadar protein dihitung dengan menggunakan persamaan sebagai berikut:
V V N 4 007 F 00 )
Keterangan :
FK = Faktor koreksi, 5.75 untuk kedelai dan 6.25 untuk kacang komak
Penentuan kadar karbohidrat dilakukan dengan by difference artinya kadar karbohidrat yang diukur sebagai sisa dari nilai dari kadar air, kadar abu, kadar protein dan kadar lemak dari sampel.
Kadar karbohidrat (% bb) = 100 % - (kadar air (%bb) + kadar abu (%bb) + kadar protein (%bb) + kadar lemak (%bb))
Hidrolisis Protein
Hidrolisis protein dilakukan secara in vitro dengan menggunakan
gastrointestinal simulation (GIS) digestions mengikuti metode yang diuraikan oleh Rui (2012). Substrat dari tempe kacang komak dihomogenkan dengan buffer glisin (0.01 M, pH 7.0) dengan konsentrasi 2.5% b/v. Substrat kemudian dihidrolisis dengan enzim α-amilase solution (1 mg/mL, 0.01 M buffer glisin, pH 7.0) pada rasio di 1:12.5 (v/w) pada suhu 37 °C selama 3 menit. Hidrolisis dilanjutkan secara berturut-turut oleh pepsin, tripsin dan α-kemotripsin dengan E/S: 1/250 (b/b, berdasarkan kandungan protein). Hidrolisis berlangsung selama 10 menit dan selanjutnya enzim diinaktivasi dengan pemanasan dalam air mendidih selama 10 menit. Campuran disentrifus (12.000 g, 4°C selama 20 menit). Supernatan dikumpulkan untuk uji lebih lanjut.
Pengukuran Derajat Hidrolisis
Derajat hidrolisis diukur berdasarkan reaksi dari jumlah kadar protein total dengan 2.4.6-trinitrobenzenesulphonic acid (TNBS) mengikuti metode yang diuraikan oleh Rui (2012). Berbagai konsentrasi dari L-Leusin mulai dari 0 – 2000 ppm digunakan untuk memperoleh kurva standar. Hidrolisat protein sampel (125 µl) ditambahkan dengan 2.0 ml buffer fosfat 0.2 M pH 8.2 dan 1.0 ml larutan TNBS 0.01 %. Larutan dicampur seluruhnya dan ditempatkan pada water bath
6
Analisis Protein Terlarut
Analisis protein terlarut dilakukan dengan menggunakan metode Bradford (1976). Larutan Bradford dibuat dengan cara sebagai berikut: sebanyak 100 mg CBB (coomassie brilliant blue) G-250 dilarutkan dalam 50 ml etanol 95%. Setelah itu 100 ml asam fosfat 85% ditambahkan. Terakhir larutan diencerkan hingga 1 liter. Larutan disaring dengan kertas saring. Larutan tersebut diencerkan 5 kali dengan akuades ketika akan digunakan. Larutan standar segar dibuat menggunakan protein BSA (bovine serum albumin) fraction V. Sebanyak 100 mg BSA ditimbang dan ditambahkan 25 ml akuades. Larutan kemudian dikocok pelan-pelan, setelah larut, diencerkan sampai 50 ml.
Setelah semua pereaksi siap, langkah selanjutnya adalah memipet masing-masing larutan dalam tiap tabung sebanyak 0.1 ml dan dimasukkan ke dalam tabung reaksi lain yang bersih. Sebanyak 5 ml pereaksi Bradford ditambahkan ke dalam masing-masing tabung reaksi. Blanko dibuat dengan cara mencampurkan 0.1 ml aquades dan direaksikan dengan 5 ml pereaksi Bradford. Setelah sekitar 5 menit, masing- c y 595 nm. Penentuan Aktivitas Penghambatan ACE
Aktivitas penghambatan ACE (Angiotensin I-Converting Enzyme) dari hidrolisat protein sampel diukur dengan spektrofotometer UV berdasarkan laju pembentukan asam hippurat dari hippuryl-L-histidyl-L-leusine (HHL) dengan modifikasi dari metode Cushman DW dan Cheung HW (1971). Untuk setiap , 0 h protein sampel dengan konsentrasi protein yang
h L 50 M 0 E Sigma)
0,8 mU pada suhu 370C selama 45 menit. Reaksi dihentikan dengan penambahan 00 N, h 600 v 30 detik, lalu disentrifugasi pada kecepatan 2000 rpm selama 10 menit. Sebanyak 00 filtrat diambil dan diuapkan pada suhu 100 – 120 0C selama 10 menit. R y h 600 N M v dan diukur absorbansinya pada panjang gelombang 228 nm.
Tabel 1 Komposisi bahan analisis penghambat ACE
Komponen V )
Blanko Kontrol Sampel
Air destilata 10 10 10
Inkubasi 370C selama 45 menit
HCl 1,0 N - 100 100
Etil asetat 600 600 600
Vorteks selama 30 detik Sentrifugasi 300 x g 10 menit
Filtrat (lapisan atas) 100 100 100
Oven 140 0C selama 10 menit
7
Aktivitas penghambatan ACE dari hidrolisat sampel dinyatakan sebagai persen (%) penghambatan yang dihitung dengan rumus:
Aktivitas Penghambatan ACE (%) = (Ac-As) x 100% (Ac-Ab)
Keterangan : Ac = absorbansi kontrol As = absorbansi sampel Ab = absorbansi blanko
Aktivitas ACE = 100 - % penghambatan
Penentuan Nilai IC50
Nilai IC50 ditentukan berdasarkan hubungan kurva linear antara nilai aktivitas penghambatan ACE (%) dengan konsentrasi hidrolisat protein dari sampel.
HASIL DAN PEMBAHASAN
Hasil
Tempe Komak
Tempe komak memiliki tampilan fisik menyerupai tempe kedelai, permukaannya ditutupi oleh miselium kapang secara merata, kompak, dan berwarna putih. Antar butiran kacang dipenuhi oleh miselium dengan ikatan yang kuat dan merata sehingga bila diiris tidak hancur. Tempe komak memiliki aroma tempe normal.
Gambar 1 Kacang komak (A) dan tempe komak (B) Kadar Proksimat
Kadar Air
Hasil analisis kadar air dapat dilihat pada Gambar 2. Penentuan kadar air dilakukan untuk mengetahui jumlah air yang terkandung di dalam sampel. Berdasarkan Gambar 2, kadar air terbesar ditunjukkan oleh tempe komak kukus sebesar 66.64%, diikuti oleh tempe komak mentah 65%, dan kacang komak 10.62%. Hal ini tidak berbeda jauh dengan kedelai, kadar air terbesar ditunjukkan
8
oleh tempe kedelai kukus sebesar 64.72%, diikuti oleh tempe kedelai mentah 63.69% dan kacang kedelai 12.30%.
Gambar 2 Kadar air kacang, tempe mentah dan tempe kukus dari komak dan kedelai
Kadar Abu
Hasil analisis kadar abu dapat dilihat pada Gambar 3. Kadar abu dari suatu bahan pangan menunjukkan total mineral yang terkandung dalam bahan tersebut. Berdasarkan Gambar 3, kadar abu terbesar ditunjukkan oleh kacang kedelai sebesar 4.77%, diikuti oleh tempe kedelai mentah 0.75% dan tempe kedelai kukus 0.74%. Hal ini tidak berbeda jauh dengan komak, kadar abu terbesar ditunjukkan oleh kacang komak sebesar 4.49%, diikuti oleh tempe komak mentah 0.51% dan tempe komak kukus 0.44%.
9 Kadar Lemak
Hasil analisis kadar lemak dapat dilihat pada Gambar 4. Berdasarkan Gambar 4, kadar lemak terbesar ditunjukkan oleh kacang kedelai sebesar 15.55%, diikuti oleh tempe kedelai mentah 1.90% dan tempe kedelai kukus 1.86%. Hal ini agak berbeda dengan komak, nilai kadar abu terbesar hanya sebesar 1.05% yang ditunjukkan oleh kacang komak, diikuti oleh tempe komak mentah 0.17% dan tempe komak kukus 0.15%.
Gambar 4 Kadar lemak kacang, tempe mentah dan tempe kukus dari komak dan kedelai
Kadar Protein Total
Hasil analisis kadar protein dapat dilihat pada Gambar 5. Berdasarkan Gambar 5, kadar protein total terbesar ditunjukkan oleh kacang kedelai sebesar 32.40%, diikuti oleh tempe kedelai mentah 21.07% dan tempe kedelai kukus 19.65%. Hal ini tidak berbeda jauh dengan komak, kadar protein terbesar ditunjukkan oleh kacang komak sebesar 28.76%, diikuti oleh tempe komak mentah 13.98% dan tempe komak kukus 13.07%.
10
Kadar Karbohidrat
Hasil analisis kadar karbohidrat dapat dilihat pada Gambar 6. Berdasarkan Gambar 6, kadar karbohidrat terbesar ditunjukkan oleh kacang komak sebesar 55.08%, diikuti oleh tempe komak mentah 20.34% dan tempe komak kukus 19.70%. Hal ini agak berbeda dengan kedelai, kadar karbohidrat dari kedelai lebih rendah dibandingkan dengan komak. Kadar karbohidrat terbesar ditunjukkan oleh kacang kedelai sebesar 34.97%, diikuti oleh tempe kedelai kukus 13.04% dan tempe kedelai mentah 12.59%.
Gambar 6 Kadar karbohidrat kacang, tempe mentah dan tempe kukus dari komak dan kedelai
Derajat Hidrolisis
Pada penelitian ini derajat hidrolisis diukur berdasarkan perbandingan total L-Leusin dengan N-total yang terdapat pada hidrolisat protein sampel. Berdasarkan tabel 2, derajat hidrolisis yang paling besar ditunjukkan oleh kacang komak sebesar 18.03%, diikuti oleh tempe komak kukus 14.20% dan tempe komak mentah 10.01%. Hal ini tidak berbeda jauh dengan kedelai, derajat hidrolisis yang paling besar ditunjukkan oleh kacang kedelai sebesar 17.05%, diikuti oleh tempe kedelai kukus 10.05% dan tempe kedelai mentah 7.22%.
Tabel 2 Derajat hidrolisis dari hidrolisat protein komak dan kedelai
Hidrolisat Sampel % Derajat Hidrolisis
Kacang Komak 18.03 ± 0.18
Tempe Komak Mentah 10.01 ± 0.13
Tempe Komak Kukus 14.20 ± 0.16
Kacang Kedelai 17.05 ± 0.05
Tempe Kedelai Mentah 7.22 ± 0.33
Tempe Kedelai Kukus 10.05 ± 0.27
Kadar Protein Terlarut
11 terlarut dari komak lebih rendah dibandingkan dengan kedelai. Kadar protein terlarut terbesar ditunjukkan oleh kacang komak sebesar 39.81%, diikuti oleh tempe komak kukus 9.39% dan tempe komak mentah 1.71%.
Tabel 3 Kadar protein terlarut dari hidrolisat protein komak dan kedelai Hidrolisat Sampel Kadar Protein Terlarut (mg/g berat kering)
Kacang Komak 39.81 ± 0.74
Tempe Komak Mentah 1.71 ± 0.35
Tempe Komak Kukus 9.39 ± 0.70
Kacang Kedelai 165.30 ± 8.85
Tempe Kedelai Mentah 16.46 ± 0.34
Tempe Kedelai Kukus 26.22 ± 1.20
Aktivitas Penghambatan ACE
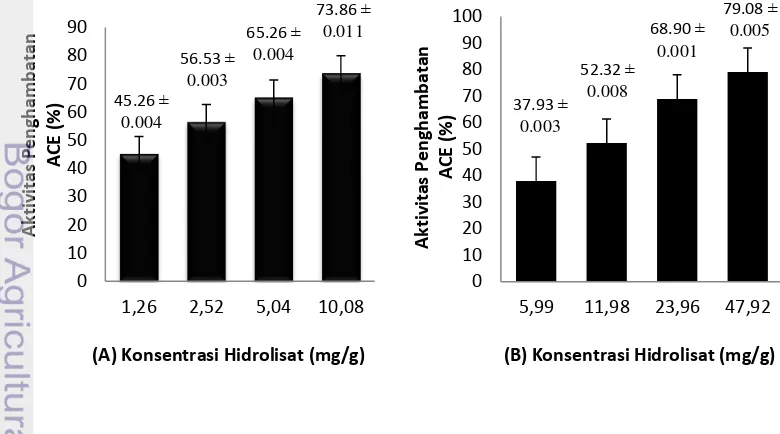
Hasil uji aktivitas penghambatan ACE menunjukkan bahwa hidrolisat protein dari masing - masing sampel memiliki kemampuan yang berbeda – beda. Pengujian dilakukan dengan blanko (tanpa penambahan hidrolisat) dan kontrol positif (kaptropil). Konsentrasi hidrolisat protein sampel divariasikan dengan 4 kali pengenceran. Pengujian pada konsentrasi bervariasi ini dimaksudkan untuk melihat pengaruh penambahan konsentrasi hidrolisat protein sampel terhadap peningkatan aktivitas penghambatan ACE.
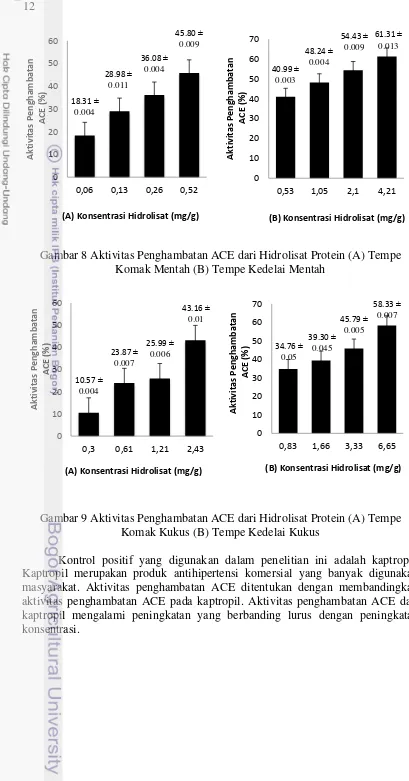
Aktivitas penghambatan ACE dari hidrolisat protein komak mengalami peningkatan yang berbanding lurus dengan peningkatan konsentrasi. Sebagai pembanding aktivitas penghambatan ACE pada hidrolisat protein komak, digunakan kedelai dengan perlakuan yang sama dengan komak yaitu dalam bentuk hidrolisat kacang utuh, hidrolisat tempe mentah dan hidrolisat tempe kukus. Hasil dari aktivitas penghambatan ACE pada kedelai tidak berbeda jauh dengan komak. Aktivitas penghambatan ACE dari hidrolisat protein kedelai mengalami peningkatan yang berbanding lurus dengan peningkatan konsentrasi.
12
Gambar 8 Aktivitas Penghambatan ACE dari Hidrolisat Protein (A) Tempe Komak Mentah (B) Tempe Kedelai Mentah
Gambar 9 Aktivitas Penghambatan ACE dari Hidrolisat Protein (A) Tempe Komak Kukus (B) Tempe Kedelai Kukus
13
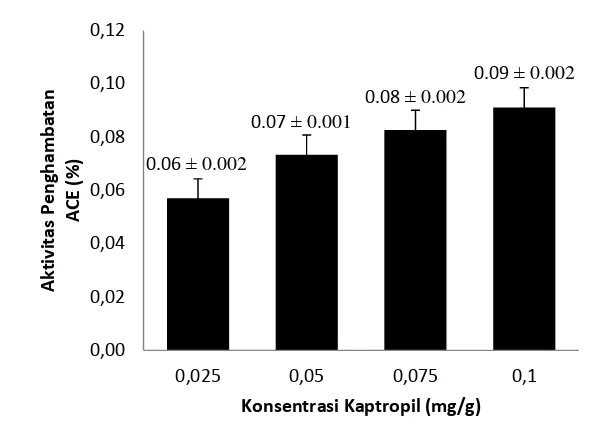
Gambar 10 Aktivitas Penghambatan ACE dari Kaptropil
Banyaknya hidrolisat protein yang diperlukan untuk mereduksi 50% aktivitas ACE, didefenisikan sebagai nilai IC50. Nilai IC50 ditentukan berdasarkan hubungan kurva linear antara nilai aktivitas penghambatan ACE (%) dengan konsentrasi hidrolisat protein dari sampel. Selanjutnya dicari nilai IC50 yang terendah (mempunyai aktivitas penghambatan maksimum) dari hidrolisat sampel.
Berdasarkan Tabel 4, hidrolisat protein tempe komak mentah memiliki nilai IC50 terendah dibandingkan dengan hidrolisat protein sampel lainnya yakni 0.64 mg/g. Hal ini berarti tempe komak mentah memiliki aktivitas penghambatan ACE yang maksimum dibandingkan dengan yang lainnya. Nilai ini tidak terlalu jauh apabila dibandingkan dengan kaptropil sebagai kontrol positif yang memiliki nilai IC50 sebesar 0.00436 mg/g.
Tabel 4 Nilai IC50 dari hidrolisat protein komak dan kedelai
Hidrolisat Sampel IC50 (mg/g)
Kacang Komak 1.24
Tempe Komak Mentah 0.64
Tempe Komak Kukus 3.51
Kacang Kedelai 10.36
Tempe Kedelai Mentah 1.69
Tempe Kedelai Kukus 17.83
Kaptropil (Kontrol Positif) 0.00436
14
Pembahasan
Tempe Komak
Tempe adalah makanan hasil fermentasi yang sangat terkenal di Indonesia. Proses pembuatan tempe melibatkan tiga faktor pendukung, yaitu bahan baku yang dipakai, mikroorganisme (kapang tempe), dan keadaan lingkungan tumbuh (suhu, pH, dan kelembaban) (Ferlina 2009). Fermentasi merupakan proses perombakan makromolekul (karbohidrat dan protein) tanpa memerlukan oksigen. Teknologi fermentasi merupakan suatu cara yang dapat memperbaiki nilai gizi bahan pangan menjadi bahan yang berkualitas baik karena rasa, aroma, tekstur, daya cerna, dan daya simpannya lebih baik dari bahan sebelumnya (Ferlina 2009). Proses pembuatan tempe dimulai dengan pengupasan kulit kacang komak dan kacang kedelai utuh kemudian ditentukan waktu perebusan untuk menghasilkan tekstur kacang yang sesuai dengan proses perendaman. Tekstur tersebut akan mempermudah proses perendaman karena laru tempe Rhizopus lebih mudah menggunakan nutrisi dari kacang (Syarief et al.1999). Perendaman bertujuan menghidrasi biji dan membiarkan fermentasi asam laktat terjadi secara alami sampai tercapai derajat keasaman yang sesuai untuk pertumbuhan kapang tempe, yaitu pH 3.5 sampai 5.2 (Hermana 2001).
Proses penirisan dan pendinginan pada suhu ruang dilakukan untuk mengurangi jumlah air dan menurunkan suhu agar kapang pada ragi dapat tumbuh optimal. Kemudian, dilakukan penimbangan sejumlah 100 gram kacang ke dalam kemasan plastik PP yang telah dilubangi sebelumnya, dan inokulasi ragi sejumlah 0.1 % dari berat bahan. Kemasan diperlukan untuk menciptakan suasana sedikit oksigen dan juga untuk mengkondisikan suhu agar selalu sesuai untuk pertumbuhan kapang. Tahapan terakhir dari pembuatan tempe komak adalah fermentasi selama 36 jam pada fermentor bersuhu 35oC.
Syarif et al. (1999) menyatakan bahwa tempe yang baik dicirikan oleh permukaan yang ditutupi oleh miselium kapang (benang-benang halus) secara merata, kompak, dan berwarna putih. Antar butiran kacang dipenuhi oleh miselium dengan ikatan yang kuat dan merata, sehingga bila diiris tempe tersebut tidak hancur. Tempe yang dihasilkan dalam penelitian ini permukaannya ditutupi oleh miselium kapang secara merata dan berwarna putih dan tidak memberi bau ammonia, sehingga tempe ini bisa dikatakan memenuhi syarat sebagai tempe. Persentase Proksimat
15 Hasil analisis kadar abu diperoleh bahwa kandungan mineral tertinggi terdapat pada kacang kedelai sebesar 4.77 % dibandingkan dengan yang lainnya. Kadar abu yang terdapat di dalam suatu bahan menunjukkan jumlah mineral seperti kalsium, besi, tembaga, seng, kalium, dan magnesium. Penelitian sebelumnya oleh Ningsih (2007) menyatakan bahwa kulit kedelai telah digunakan sebagai pakan ternak karena kandungan besi pada kulit kedelai sangat tinggi. Hal ini mengindikasikan bahwa adanya tahap pengupasan kulit pada proses pembuatan tempe mengakibatkan sejumlah mineral yang terdapat dalam kulit kacang terbuang sehingga kadar abu dari tempe lebih rendah dibandingkan dengan kadar abu kacang utuh. Proses pengukusan tidak berpengaruh nyata karena natrium klorida (komponen mineral pembentuk abu) akan hilang dari abu jika suhu pem h 600˚ D λλ7)
Hasil analisis kadar lemak diperoleh bahwa kacang kedelai memiliki kadar lemak yang paling besar yakni sebesar 15.55 % dibandingkan dengan yang lainnya. Hal ini diduga disebabkan karena selama tahapan sebelum fermentasi dan selama fermentasi, kandungan lemak pada bahan mentah ada yang terlarut (selama tahapan pengolahan) dan ada yang terurai menjadi asam-asam lemak (selama fermentasi). Penurunan kadar lemak pada tempe juga menyebabkan asam lemak yang dapat teroksidasi menjadi lebih rendah, sehingga rasa pahit yang ditimbulkan oleh oksidasi asam lemak dan aroma tengik dapat dicegah.
Hasil analisis kadar protein diperoleh bahwa kacang kedelai memiliki kadar protein yang paling besar yakni sebesar 32.40 % dibandingkan dengan kadar protein dalam bentuk tempe. Hal ini disebabkan oleh aktivitas kapang selama fermentasi tempe yang dapat meningkatkan jumlah nitrogen terlarut (Angulo et al. 2007) sehingga tidak ikut mengendap saat pengendapan protein pada pH isoelektriknya. Kapang yang berperan adalah Rhizopus oligosporus
karena dapat menghasilkan enzim protease (Ningsih 2007). Selain itu, proses perendaman, perebusan dan pengupasan kulit ari (kotiledon) biji juga dapat menyebabkan penurunan kadar protein karena dapat meningkatkan jumlah protein terlarut sehingga sebagian protein larut air hilang selama proses perendaman, perebusan, pengukusan dan pengupasan kulit kedelai dan komak.
Hasil analisis kadar karbohidrat ditentukan secara by difference. Kadar karbohidrat kacang komak diperoleh memiliki kadar karbohidrat terbesar yakni sebesar 55.08 % dibandingkan dengan yang lainnya. Data tersebut mengindikasikan bahwa proses fermentasi dapat menurunkan kadar karbohidrat tempe komak. Hal ini disebabkan oleh proses perendaman kacang komak selama dua malam pada pembuatan tempe komak menyebabkan penurunan kadar karbohidrat karena karbohidrat tersebut digunakan oleh bakteri asam laktat, terutama karbohidrat golongan oligosakarida, seperti stakiosa, verbakosa, dan rafinosa (Ningsih 2007). Penurunan kadar karbohidrat selama proses fermentasi yang disebabkan oleh pemecahan gula-gula kompleks seperti pati, stakiosa dan rafinosa yang menyebabkan flatulensi menjadi gula-gula yang mudah dicerna oleh kapang (Hermana 2001).
Persentase Derajat Hidrolisis
16
sifat dari hidrolisat protein berhubungan erat dengan parameter tersebut (Nissen 1979). Pengendalian ini diperlukan karena daya hidrolitik suatu enzim dapat bervariasi berdasarkan sumber dan substrat yang digunakan. Besarnya derajat hidrolisis terkait erat dengan jumlah produk hidrolisat yang dihasilkan, atau dengan kata lain besarnya derajat hidrolisis memiliki kecenderungan yang sama dengan jumlah protein terlarut atau gugus amino bebas (Handajani 2001). Derajat hidrolisis, substrat dan protease yang digunakan akan dapat mempengaruhi karakteristik fisikokimia hidrolisat yang dihasilkan (Kristinsson 2000a). Melalui teknik hidrolisis, protein dari suatu bahan dapat diubah menjadi senyawa asam amino L, nukleotida, dan berbagai ragam peptida.
Pada penelitian ini derajat hidrolisis diukur berdasarkan perbandingan total leusin dengan N total yang terdapat pada hidrolisat protein sampel. Berdasarkan Tabel 2, derajat hidrolisis yang paling besar ditunjukkan oleh kacang komak, diikuti oleh tempe komak kukus dan tempe komak mentah. Hal ini tidak berbeda jauh dengan kedelai. Hal tersebut diduga, pada protein tempe sudah dipecah oleh enzim protease yang dihasilkan kapang tempe saat proses fermentasi. Sehingga pada enzim-enzim yang digunakan saat hidrolisis protein (amilase, pepsin, tripsin, kemotripsin) hanya memecah menguraikan protein yang belum dipecah saat fermentasi tempe. Pada saat fermentasi tempe, sebagian protein telah dipecah menjadi peptida-peptida oleh enzim protease yang dihasilkan oleh kapang. Pemecahan peptida tersebut dipengaruhi oleh konsentrasi enzim dan lamanya waktu fermentasi. Hubungannya dengan hidrolisis protein ini, peptida-peptida tersebut tidak lagi diurai saat proses hidrolisis dengan pepsin, tripsin, dan kemotripsin. Sehingga derajat hidrolisis yang dihasilkan dari tempe lebih kecil daripada kacang mentah.
Peningkatan derajat hidrolisis setelah pengukusan tempe saat ini masih belum diketahui penyebab pastinya. Saat tempe mengalami pengukusan, protein terdenaturasi yang menyebabkan protein menjadi rusak. Hal tersebut seharusnya dapat menurunkan derajat hidrolisis sampel. Namun, kenyataannya derajat hidrolisis pada tempe yang dikukus meningkat dari tempe mentah. Ada kemungkinan, proses pemanasan tersebut mampu mengembalikan kestabilan protein yang terkandung dalam sampel.
Kadar Protein Terlarut
Kadar protein terlarut pada penelitian ini dilakukan sesuai dengan metode Bradford. Prinsip metode Bradford didasarkan pada pengikatan secara langsung zat warna Coomassie Briliant Blue G250 (CBBG) oleh protein yang mengandung rantai samping aromatik (tirosin, triptofan, dan phenylalanin) atau bersifat basa (arginin, histidin, dan leusin). Reagen CBBG bebas berwarna merah kecoklatan, sedangkan dalam suasana asam reagen CBBG akan berada dalam bentuk anion yang mengikat protein membentuk warna biru. Jumlah CBBG yang terikat pada protein proporsional dengan muatan positif yang ditemukan pada protein (Bollag dan Edelstein 1991).
17 struktur sekunder dan tersier yang terdapat dalam struktur protein mengalami kerusakan (Yan Song et al. 2010). Pada proses fermentasi, protein dapat terhidrolisis menjadi asam-asam amino dengan ukuran molekul yang lebih kecil. Hal ini juga mengindikasikan bahwa semakin besar konsentrasi sampel maka kadar protein terlarut hidrolisat sampel semakin tinggi. Hal ini dikarenakan aktivitas enzim-enzim amilase, pepsin, tripsin dan kemotripsin yang terkandung di dalam sampel bersifat memecah protein menjadi peptida pendek dan asam-asam amino yang mudah larut.
Konsentrasi protein terlarut ini sebanding dengan derajat hidrolisis. Saat kacang diolah menjadi tempe, derajat hidrolisisnya menurun begitu pula dengan konsentrasi protein terlarutnya. Dari hasil ini dapat diketahui, peptida yang dihasilkan dari fermentasi tempe sebagian besar protein tak terlarut. Hal ini terbukti dari total protein meningkat setelah proses fermentasi (setelah menjadi tempe) yang ditunjukkan dari hasil proksimat kadar protein. Hal inilah yang menyebabkan nilai derajat hidrolisis dan konsentrasi terlarut tempe menurun. Hasil Analisis Penghambatan ACE
ACE merupakan enzim yang dapat meningkatkan tekanan darah dengan cara mengonversi angiotensin I (suatu dekapeptida) menjadi suatu vasoconstrictor
kuat angiotensin II (suatu oktapeptida). ACE merupakan enzim multifungsi, yang juga dapat mengkatalisis degradasi bradikinin (suatu nonapeptida vasodilator yang dapat melebarkan pembuluh darah) dan enkefalin (suatu pentapeptida). Oleh karena itu, penghambatan aktivitas ACE dapat mengurangi peranan angiotensin II tetapi dapat meningkatkan kadar bradikinin dan enkefalin, sehingga akan menurunkan tekanan darah. Angiotensin II menjadi berbahaya karena dapat mempengaruhi sistem kardiovaskular dengan penyempitan pembuluh darah dan melepaskan hormon yang dapat meningkatkan tekanan darah. Pada proses pengobatan hipertensi, pencegahan terhadap ACE menjadi salah satu teknik pencegahan modern (Hansen et al. 1995).
Aktivitas ACE dapat ditentukan dari jumlah asam hipurat yang terhidrolisis dari substrat HHL setelah penambahan hidrolisat dengan metode spektrofotometri pada panjang gelombang 228 nm. Pengukuran UV dilakukan pada panjang gelombang 228 nm karena senyawa yang akan diukur tidak berwarna dan panjang gelombang maksimum asam hipurat adalah 228 nm. Hidrolisat sampel dilarutkan dalam air destilata, NaCl 4 M, buffer borat pH 8.3 h L E, h 32˚ 45 menit. Reaksi akan dihentikan dengan penambahan HCl dan etil asetat. Asam hipurat yang terbentuk akan terhidrolisis dalam etil asetat. Aktivitas penghambatan ACE ditentukan dengan membandingkan absorbansi dari kontrol dikurang absorbansi sampel dengan absorbansi kontrol dikurang absorbansi blanko.
18
yang dihasilkan, semakin besar aktivitas penghambatan ACE. Absorbansi yang terukur berasal dari sisa asam hipurat hasil reaksi antara substrat dan ACE yang tidak dihambat oleh hidrolisat sampel.
Kontrol positif yang digunakan dalam penelitian ini adalah kaptropil. Kaptropil adalah obat antihipertensi yang mekanismenya adalah menghambat enzim ACE secara langsung. Kaptropil memiliki afinitas yang tinggi terhadap ACE dan berkompetisi secara kompetitif dengan angiotensin I sebagai substrat alami, untuk mencegah terjadinya angiotensin II. Namun, jika dikonsumsi secara terus menerus, kaptropil dapat memberikan efek yang kurang baik bagi tubuh sehingga dengan penelitian ini dapat menjadi solusi dalam memanfaatkan alternatif alamiah yang lebih aman.
Hidrolisat protein sampel sebagai inhibitor ACE bekerja dengan cara mencegah terjadinya reaksi dari angiotensin I menjadi angiotensin II. Proses perubahan angiotensin I menjadi angiotensin II dengan memutuskan terminal C dipeptida yaitu histidil-leusin secara hidrolitik oleh ACE. ACE akan mengubah substrat (HHL sebagai pengganti angiotensin I) menjadi asam hipurat dengan penambahan asam. Semakin kecil asam hipurat yang dihasilkan maka inhibitor yang digunakan akan semakin baik (Stella 2009).
Aktivitas penghambatan ACE ditentukan melalui nilai IC50 dari masing - masing hidrolisat protein sampel. Nilai IC50 berbanding terbalik dengan aktivitas penghambatan ACE. Semakin kecil nilai IC50, maka akan semakin besar aktivitas penghambatan terhadap ACE (Thakam 2012). Berdasarkan Tabel 4, hidrolisat tempe komak mentah memiliki nilai IC50 terkecil yakni 0.64 mg/g dibandingkan dengan yang lainnya. Nilai ini tidak terlalu jauh apabila dibandingkan dengan kaptropil sebagai kontrol positif yang memiliki nilai IC50 sebesar 0.00436 mg/g. Hal ini berarti tempe komak mentah memiliki aktivitas penghambatan ACE yang terbesar dibandingkan dengan yang lainnya yang berarti tempe komak mentah lebih berpotensi sebagai ACE inhibitor dibandingkan dengan kedelai.
Berdasarkan penelitian Karni (1997) bahwa nilai IC50 dari kedelai mentah sekitar 0.85 mg/ ml. Namun setelah dilakukan proses perkecambahan dalam waktu 24 jam, maka nilai IC50 menjadi lebih rendah yakni sebesar 0.46 mg/ml. Hal ini disebabkan oleh adanya proses perkecambahan atau germinasi maka protein kedelai yang bersifat kompleks telah dihidrolisa oleh enzim-enzim proteolitik yang aktivitasnya distimulir oleh air yang membasahi embrio, menghasilkan peptide-peptida yang jumlah dan jenisnya beragam. Hal ini menunjukkan bahwa proses pengolahan kacang utuh memiliki nilai IC50 lebih rendah dibandingkan dalam bentuk kacang utuh, yang berarti aktivitas penghambatan ACE-nya lebih tinggi.
SIMPULAN DAN SARAN
Simpulan
19 berpotensi sebagai ACE inhibitor. Aktivitas inhibitor ACE tertinggi dihasilkan oleh hidrolisat protein tempe komak mentah dengan nilai IC50 0.64 mg/g.
Saran
Hidrolisat protein tempe komak mentah perlu dievaluasi lebih lanjut terkait proses pemurnian untuk melihat komponen peptida yang kemungkinan besar bersifat sebagai ACE inhibitor.
DAFTAR PUSTAKA
Alvina S. 2009. Studi Sifat Fisikokimia, Sifat Fungsional, Nutrisi, dan Kapasitas Antioksidan Konsentrat Protein Tempe Kacang Komak (Lablab purpureus (L.) Sweet.[skripsi].Bogor: Institut Pertanian Bogor.
Angulo Bejarano P, Verdugo Montypya, Cuevas Rodrigues J, Reyes-Moreno. 2007. Tempeh Flour from Chickpea (Cicer arientinum L.) Nutritional and Physicochemical Properties. J Agri Food Chem 106:106-112.
[AOAC] Association of Official Analytical and Chemist. 2005. Official Method of Analysis of The Association Analytical of Chemist. Arlington: The Association of Official Analysis Chemist, Inc.
Astawan M, Mita W. 2003. Teknologi Pengolahan Pangan Nabati Tepat Guna. Jakarta: Akademika Pressindo.
Actis G, Ottaviani, Keen CL, Fraga CG. 2003. Inhibition of Angiotensin Converting Enzyme (ACE) Activity by Flavan 3-ols and Procyanidin.
FEBS 555:597-600.
[BPS] Balai Pusat Statistik. 2012. Produksi Padi, Jagung, dan Kedelai (Angka Tetap 2011 dan Angka Ramalan I 2012). Berita Resmi Statistik : No. 43/07/Th. XV.
Bollag DM, Edelstein SJ. 1991. Protein Methods. Willey-Liss, Inc., New York. Bradford MM. 1976. A rapid and Sensitive Method for Quantitation of
Microgram Quantities of Protein Utilizing the Principle of Protein Dye-binding. Anal Biochem 72:234-254.
Cushman DW, Cheung HW.1971. Spectrophotometric Assay and Properties of the Angiotensin Converting Enzyme of the Rabbit Lung. Biochem Pharmacol 20:1637 – 1648.
DeMan JM. 1989. Kimia Makanan. Bandung: ITB Pr.
Hansen K, Nyman U, Smitt UW, Adsersen A, Gudiksen L, Rajasekharan S, Pushpangadan P. 1995. In Vitro Screening of Traditional Medicines for Antihypertensive Effect Based on Inhibition of Angiotensin Converting Enzyme (ACE). J Ethnopharmacol 48:43-51.
20
of Tempe: The Unique Fermented Soyfood of Indonesia. Jakarta: Indonesian Tempe Foundation. Terjemahan dari: Bunga Rampai Tempe Indonesia.
Ferlina F. 2009. Tempe. [Internet]. [diunduh 2013 September 5]. Tersedia pada : http://www.adln.llb.unair.ac.id/go. php.
Handajani S. 2001. Indigenous Mucuna Tempe As Functional Food. Asia Psific J Clin Nutr 10(3):222-225.
Karni. 1997. Mempelajari Daya Hipotensif Kecambah Kedelai [skripsi]. Bogor: Institut Pertanian Bogor.
Kay EK. 1979. Food Legumes. London: Tropical Products Institute.
Khodijah S. 2003. Pola Elektroforesis Protein Globulin 7S dan 11S dari Kacang Komak (Lablab purpureus (L.) sweet) [skripsi]. Bogor: Institut Pertanian Bogor.
Kristinsson HG, Rasco BA 200a. Kineticts of the Hydrolysis of Atlantic Salmon (Salmo salar) Muscle Proteins by Alkaline Proteases and A Visceral Serine Protease Mixture. Process Biochemistry 36:131-139.
Ningsih W. 2007. Evaluasi Senyawa Fenolik (Asam Ferulat dan Asam p-Kumarat) pada Biji, Kecambah, dan Tempe Kacang Tunggak (vigna ungulculata) [skripsi]. Bogor: Institut Pertanian Bogor.
Nissen J. 1979. Determination of the Degree of Hydrolysis of Food Protein Hydrolysates by Trinitrobenzenesulfonic Acid. J Agri Food Chem
27:1256-1262.
Stella A. 2009. Studi Sifat Fisikokimia, Sifat Fungsional, Nutrisi, dan Kapasitas Antioksidan Konsentrat Protein Tempe Kacang Komak (Lablab purpureus (L.) sweet) [skripsi]. Bogor: Institut Pertanian Bogor.
Subagio A. 2006. Characterization of Hyacinth Bean (Lablab purpureus (L.) Sweet) Seeds from Indonesia and Their Protein Isolate. J Food Chem
95 :65-70.
Suwarno M. 2003. Potensi Kacang Komak (Lablab purpureus (L.) Sweet) Sebagai Bahan Baku Pembuatan Isolat Protein [skripsi]. Bogor: Institut Pertanian Bogor.
Syarief R, Hermanianto J, Hariyadi P, Wiraatmadja S, Suliantari, Dahrulsyah, Suyatna NE, Saragih YP, Arisasmita JH, Kuswardani I, Astuti M. 1999.
Wacana Tempe Indonesia, Yee YB, Basry AA, Puruhita A, Supriyono, editor. Surabaya (ID): Universitas Katolik Widya Mandala.
Thakam A, Saewanm N, Kittigowittang K, Jimtaisong A. 2012. Antioxidant and Antityrosinase Activities of Metal Complexes of Curcuma petiolata
Extract for Cosmetics Applications. 1st mae Fah Luang University International Conference 2012.
21 http://www.intechopen.com/books/soybean-and-health/anti-diarrhoeal-aspects-of-fermented-soya-beans.
Xin Rui et al. 2012. Electrophoretjc Profiles and Angiotensin I- Converting Enzyme Inhibitory Activities of Nine Varieties of Phaseoulus Vulgaris Protein Hydrolysates. Food Res Int 44:2497-2504.
22
Lampiran 1 Diagram alir penelitian
Analisis Proksimat
Hidrolisis Protein
Uji ACE Inhibitor Kacang
komak
Tempe Komak Mentah
Hidrolisat Kacang Komak
Hidrolisat Tempe Komak Kukus
Pengukuran Derajat Hidrolisis
Analisis Protein Metode Bradford
Tempe Komak Kukus
23 Lampiran 2 Tabel dan gambar hasil pengamatan absorbansi pada konsentrasi
standar BSA yang berbeda
Konsentrasi BSA standar (mg/ml) Absorbansi
0.00 0.000
0.20 0.215
0.40 0.340
0.60 0.485
0.80 0.590
1.00 0.690
1.20 0.810
y = 0,6482x + 0,0582 R² = 0,985
0,000 0,200 0,400 0,600 0,800 1,000
0,00 0,20 0,40 0,60 0,80 1,00 1,20 1,40
A
b
sor
b
an
si
24
Lampiran 3 Tabel dan gambar hasil pengamatan absorbansi pada konsentrasi standar L-Leusin yang berbeda
[Leusin] ppm Volume larutan leusin (ml)
Volume akuades
(ml) Absorbansi
0 0 2.0 0
400 0.4 1.6 0.201
800 0.8 1.2 0.415
1200 1.2 0.8 0.546
1600 1.6 0.4 0.721
2000 2.0 0 0.825
y = 0,0004x + 0,0359 R² = 0,9882
0 0,2 0,4 0,6 0,8 1
0 500 1000 1500 2000 2500
A
b
sor
b
an
si
25 Lampiran 4 Analisis kadar air
Sampel Bobot Cawan Kosong (g) Bobot Sampel (g) Bobot Cawan + Sampel Akhir (g) Kadar Air (%) Kadar Air Rerata (%)
U1 U2 U3 U1 U2 U3 U1 U2 U3 U1 U2 U3
Kacang
Komak 24.0012 22.2216 23.3025 2.0019 2.0010 2.0001 25.7888 24.0129 25.0891 10.70 10.48 10.67 10.62
Tempe Komak Mentah
23.8748 22.8936 22.4832 5.0077 5.0078 5.0064 25.6080 24.6456 24.2555 65.39 65.01 64.60 65.00
Tempe Komak kukus
26.3010 19.2656 24.8571 5.0036 5.0052 5.0036 27.9673 20.9384 26.5256 66.70 66.58 66.65 66.64
Kacang
Kedelai 23.0287 23.4444 18.3566 2.0006 2.0005 2.0008 24.7892 25.2023 20.1016 12.00 12.13 12.78 12.30 Tempe
Kedelai Mentah
23.4530 21.2079 24.3013 5.0054 5.0040 5.0054 25.2586 23.0296 26.1264 63.93 63.60 63.54 63.69
Tempe Kedelai
Kukus
23.0395 23.157 21.9184 5.0031 5.0073 5.0068 24.8180 24.9194 23.6803 64.45 64.89 64.81 64.72
Kadar air tempe komak mentah (% bb) = x 100% =
x 100%
= 65.39 %
26
Lampiran 5 Analisis kadar abu
Sampel Bobot Cawan Kosong (g) Bobot Sampel (g) Bobot Cawan + Sampel Akhir (g) Kadar Abu (%) Rerata Kadar Abu (%)
U1 U2 U3 U1 U2 U3 U1 U2 U3 U1 U2 U3
Kacang
Komak 24.0012 22.2216 23.3025 2.0019 2.0010 2.0001 24.0865 22.3175 23.3907 4.26 4.79 4.41 4.49
Tempe Komak Mentah
23.8748 22.8936 22.4832 5.0077 5.0078 5.0064 23.9013 22.9180 22.5090 0.53 0.49 0.52 0.51
Tempe Komak kukus
26.3010 19.2656 24.8571 5.0036 5.0052 5.0036 26.3240 19.2867 24.8786 0.46 0.42 0.43 0.44
Kacang
Kedelai 23.0287 23.4444 18.3566 2.0006 2.0005 2.0008 23.1221 23.5425 18.4516 4.67 4.90 4.75 4.77 Tempe
Kedelai Mentah
23.4530 21.2079 24.3013 5.0054 5.0040 5.0054 23.4917 21.2453 24.3375 0.77 0.75 0.72 0.75
Tempe Kedelai
Kukus
23.0395 23.157 21.9184 5.0031 5.0073 5.0068 23.0771 23.1900 21.9586 0.75 0.66 0.80 0.74
Kadar abu tempe komak mentah (% bb) = x 100% =
x 100%
27 Lampiran 6 Analisis kadar lemak
Sampel Bobot Labu Kosong (g) Bobot Sampel (g) Bobot Labu + Sampel Akhir (g)
Kadar Lemak (%) Rerata Kadar Lemak (%)
U1 U2 U3 U1 U2 U3 U1 U2 U3 U1 U2 U3
Kacang
Komak 30.1333 32.3452 32.9966 2.0000 2.0023 2.0022 30.1542 32.3682 33.0156 1.05 1.15 0.95 1.05
Tempe Komak Mentah
32.6573 32.6153 32.6059 2.0044 2.0020 2.0045 32.6584 32.6189 32.6116 0.05 0.18 0.28 0.17
Tempe Komak kukus
31.7700 31.7777 32.8650 2.0010 2.0050 2.0018 31.7732 31.7800 32.8684 0.16 0.11 0.17 0.15
Kacang
Kedelai 32.6098 32.6593 25.9869 2.0010 2.0022 2.0018 32.9221 32.9882 26.2792 15.61 16.43 14.60 15.55 Tempe
Kedelai Mentah
31.7700 31.7777 32.865 2.0010 2.0050 2.0018 31.8095 31.8152 32.9024 1.97 1.87 1.87 1.90
Tempe Kedelai
Kukus
32.5099 32.7582 32.865 2.0010 2.0022 2.0012 32.5423 32.8011 32.9012 1.62 2.14 1.81 1.86
h ) – ) 00
h ) 32 6584 – 32 6572)2 0044 00
28
Lampiran 7 Analisis kadar protein
Sampel Bobot Sampel (g) V HCl (ml) % N
Kadar Protein (%) Rerata Kadar Protein (%)
U1 U2 U3 U1 U2 U3 U1 U2 U3 U1 U2 U3
Kacang
Komak 0.5011 0.5001 0.5006 16.2 16.4 16.35 4.5645 4.6301 4.6114 28.53 28.94 28.82 28.76
Tempe Komak Mentah
0.5001 0.5013 0.5010 7.9 7.7 8.2 2.2304 2.1687 2.3109 13.94 13.55 14.44 13.98
Tempe Komak kukus
0.5042 0.5044 0.5036 7.5 7.3 7.6 2.1002 2.0434 2.1308 13.13 12.77 13.32 13.07
Kacang
Kedelai 0.5019 0.5004 0.5028 18.42 18.42 18.5 5.1621 5.1973 5.1950 29.68 29.88 29.87 29.81 Tempe
Kedelai Mentah
0.5042 0.5044 0.5036 11.85 11.95 12.3 3.3183 3.3450 3.4485 19.08 19.23 19.83 19.38
Tempe Kedelai
Kukus
0.5042 0.5044 0.5036 11.3 11.22 11.15 3.1643 3.1407 3.1260 18.19 18.06 17.97 18.08
N 7 λ – 0) 0 008 4 0070 500 00 = 2.2304
29 Lampiran 8 Analisis kadar karbohidrat
Sampel Kadar Air (%) Kadar Abu (%) Kadar Lemak (%) Kadar Protein (%)
Kadar Karbohidrat (%) Rerata Kadar Karbohidrat
(%)
U1 U2 U3 U1 U2 U3 U1 U2 U3 U1 U2 U3 U1 U2 U3
Kacang
Komak 10.70 10.48 10.67 4.26 4.79 4.41 1.05 1.15 0.95 28.53 28.94 28.82 55.46 54.64 55.15 55.08
Tempe Komak Mentah
65.39 65.01 64.60 0.53 0.49 0.52 0.05 0.18 0.28 13.94 13.55 14.44 20.09 20.77 20.16 20.34
Tempe Komak kukus
66.70 66.58 66.65 0.46 0.42 0.43 0.16 0.11 0.17 13.13 12.77 13.32 1955 20.12 19.70 19.70
Kacang
Kedelai 12.00 12.13 12.78 4.67 4.90 4.75 15.61 16.43 14.60 32.26 32.48 32.47 35.46 34.06 34.97 34.97 Tempe
Kedelai Mentah
63.93 63.60 63.54 0.77 0.75 0.72 1.97 1.87 1.87 20.74 20.91 21.55 12.59 12.87 12.59 12.59
Tempe Kedelai
Kukus
64.45 60.89 64.81 0.75 0.66 0.80 1.62 2.14 1.81 19.78 19.63 19.54 13.40 12.68 13.04 13.04
Kadar karbohidrat tempe komak mentah (% bb) = 100 % - (kadar air + kadar abu + kadar protein + kadar lemak ) = 100 % - (65.39 + 0.53 + 0.05 + 13.94) %
Lampiran 9 Derajat hidrolisis
31 Total protein tempe komak mentah =
=
= 998.50 mg Derajat hidrolisis tempe komak mentah = T
N 00
= λ8 42
32
Lampiran 10 Analisis protein terlarut
33 Lampiran 11 Korelasi antara kosentrasi hidrolisat protein sampel dengan aktivitas
penghambatan ACE
Konsentrasi hidrolisat protein kacang komak (mg/g)
y = 33,673x + 28,493
Konsentrasi hidrolisat protein tempe komak mentah (mg/g)
y = 4,0271x + 32,174
34
Konsentrasi hidrolisat protein kacang kedelai (mg/g)
y = 5,1989x + 41,209
0,00 1,00 2,00 3,00 4,00 5,00
A
Konsentrasi hidrolisat protein tempe kedelai mentah (mg/g)
y = 4,0271x + 32,174
35 - Kaptropil
y = 4,0271x + 32,174 R² = 0,9969
0 10 20 30 40 50 60 70
0 1 2 3 4 5 6 7
A
kt
iv
itas
Pen
g
h
am
b
atan
A
CE
(%
)
36
Lampiran 12 Aktivitas penghambatan ACE dan nilai IC50 - Kacang komak
Ulangan
Konsentrasi hidrolisat kacang komak (mg/g)
Aktivitas Penghambatan ACE (%) 45.2573 56.5247 65.2517 73.8542
Nilai IC50 1.24
- Tempe komak mentah Ulangan
Konsentrasi hidrolisat tempe komak mentah (mg/g)
Aktivitas Penghambatan ACE (%) 28.9757 36.0864 47.8032 55.3218
Nilai IC50 0.64
- Tempe komak kukus Ulangan
Konsentrasi hidrolisat tempe komak kukus (mg/g)
Aktivitas Penghambatan ACE (%) 23.8698 25.9955 43.1630 58.3396
Nilai IC50 3.51
Aktivitas Penghambatan ACE (%) 37.9300 52.3200 68.9000 79.0800
Nilai IC50 10.36
- Tempe kedelai mentah Ulangan
Konsentrasi hidrolisat tempe kedelai mentah (mg/g)
0.51 1.03 2.06 4.12
1 0.644 0.592 0.505 0.442
37
3 0.649 0.589 0.521 0.465
Rata- rata 0.648 0.593 0.511 0.450
Aktivitas Penghambatan ACE (%) 40.9892 48.2423 54.4328 61.3073
Nilai IC50 1.69
- Tempe kedelai kukus Ulangan
Konsentrasi hidrolisat tempe kedelai kukus (mg/g)
0.81 1.64 3.28 6.56
1 0.750 0.667 0.584 0.471
2 0.753 0.671 0.584 0.473
3 0.761 0.662 0.594 0.484
Rata- rata 0.755 0.667 0.587 0.476
Aktivitas Penghambatan ACE (%) 34.7525 39.3030 45.7926 58.3396
Nilai IC50 17.83
- Kaptropil
Ulangan
Konsentrasi hidrolisat kaptropil (mg/g)
0.025 0.05 0.075 0.1
1 0.489 0.342 0.258 0.187
2 0.491 0.343 0.262 0.187
3 0.487 0.340 0.260 0.182
Rata- rata 0.489 0.342 0.260 0.185
Aktivitas Penghambatan ACE (%) 56.8700 73.4800 82.6800 91.1300
38
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 11 Juli 1991 di Pematangsiantar dari Ayahanda David Sinaga dan Ibunda Heppy Saragih, sebagai anak pertama dari empat bersaudara. Penulis mulai menempuh pendidikan formal di SD Budi Mulia Pematangsiantar 1997 – 2003. Penulis melanjutkan pendidikan ke SMP Cinta Rakyat 1 Pematangsiantar 2003 – 2006, kemudian melanjutkan ke SMA Budi Mulia Pematangsiantar pada tahun 2006 – 2009. Penulis melanjutkan pendidikan di departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor melalui jalur masuk Seleksi Masuk Perguruan Tinggi Nasional (SMPTN) pada tahun 2009.
Selama mengikuti perkuliahan, penulis mengikuti berbagai organisasi kampus sebagai anggota divisi biro fundrising CREBs (Community of Reasearch and Education in Biochemistry) tahun 2011-2012, sebagai kadiv kewirausahaan di KEMAKI IPB tahun 2011-2012, sebagai sekretaris Organisasi Daerah Ikanmass 2010 - 1011. Selain itu penulis banyak mengikuti kepanitiaan yaitu IDEA (2009), ISEE (2010), Pesta Sains (2011), kadiv dekor natal civa IPB (2011), Journalistic Fair (2012), dan humas Natal Civa (2012).