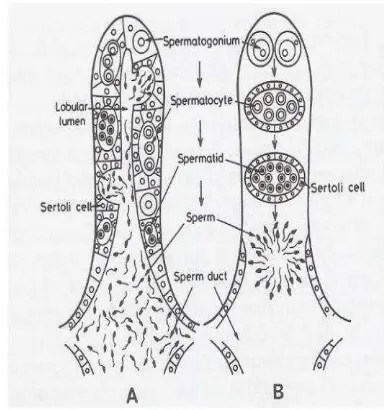
Pengaruh Penambahan Serum dan atau DNase dalam Medium Disosiasi terhadap Jumlah dan Viabilitas Spermatogonia Ikan Gurame (Osphronemus gouramy Lac.)
Teks penuh
Gambar


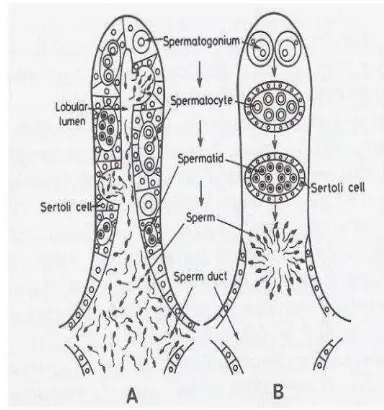
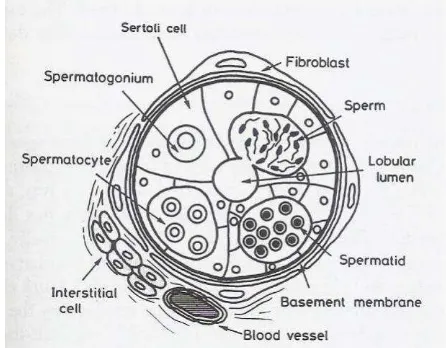

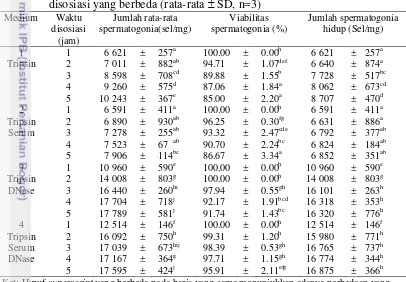
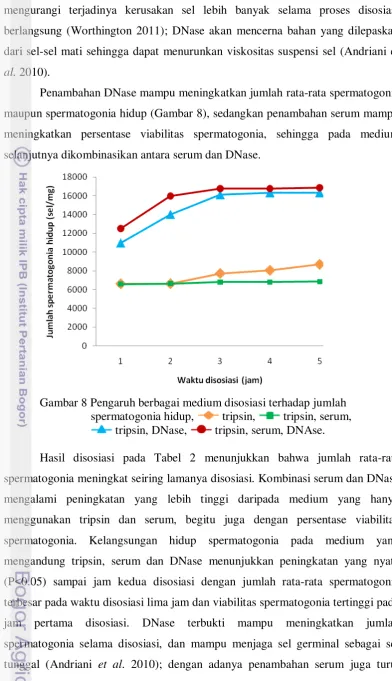


Dokumen terkait
Program HIV termasuk di dalamnya promosi tes HIV bagi pasangan populasi kunci dan serodiskordan perlu dikerangkakan dalam bingkai kesehatan seksual dan reproduksi agar
Total padatan adalah bagian padat yang terdiri dari bahan yang dicampurkan dan nutrisi yang terkandung didalamnya yaitu lemak, protein, karbohidrat, mineral,
Dengan ini saya menyatakan bahwa segala pernyataan dalam tesis ANALISIS KINERJA PETERNAK AYAM BROILER PADA POLA KEMITRAAN DAN POLA MANDIRI DI KABUPATEN LANGKAT (Studi Kasus
Berdasarkan paparan data dan pembahasan tentang peningkatan kemampuan merancang pembuktian hipotesis dalam pembelajaran menggunakan pendekatan saintifik pada
[r]
Mardalis, Metode Penelitian : Suatu Pendekatan Proposal , Jakarta : Bumi Aksara, 1999, Ritzer George & Douglas J. Goodman., Teori Sosiologi Modern, Handbook of
Menimbang : bahwa untuk melaksanakan ketentuan pasal 7 ayat (3) Peraturan Menteri Dalam Negeri Nomor 17 Tahun 2007 tentang Pedoman Teknis Pengelolaan Barang
Ada satu hal yang menarik dari yang dilakukan Ron Clark demi menarik perhatian siswa-siswinya. Sudah sering Ron Clark tidak dipedulikan oleh siswa-siswinya. Ron Clark sendiri
