PENGEMBANGAN PADI TOLERAN SALINITAS
MELALUI MUTASI DAN SELEKSI IN VITRO
:MEKANISME FISIOLOGI TOLERANSI
ROSSA YUNITA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK
CIPTA*
Dengan ini saya menyatakan bahwa disertas berjudul Pengembangan Padi Toleran Salinitas Melalui Mutasi dan Seleksi In Vitro : Mekanisme Fisiologi Toleransi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2015
ROSSA YUNITA
RINGKASAN
ROSSA YUNITA. Pengembangan Padi Toleran Salinitas Melalui Mutasi dan Seleksi In Vitro : Mekanisme Fisiologi Toleransi. Dibimbing oleh NURUL KHUMAIDA, DIDY SOPANDIE dan IKA MARISKA.
Untuk memenuhi kebutuhan beras di Indonesia, perlu dilakukan peningkatan produktivitas tanaman padi melalui perluasan areal tanam dengan pemanfaatan lahan marginal seperti tanah salin. Masalah yang dihadapi adalah ketersediaan varietas yang adaptif sangat terbatas. Penggunaan varietas toleran salinitas lebih menguntungkan karena akan menurunkan biaya produksi. Untuk itu diperlukan varietas baru yang beradaptasi di tanah salin.
Tujuan utama dari penelitian ini adalah untuk mendapatkan nomor-nomor mutan somaklon padi toleran salinitas. Penelitian ini terdiri atas enam percobaan utama. Percobaan pertama adalah studi regenerasi tunas adventif dari kalus beberapa varietas padi. Percobaan kedua adalah pengaruh iradiasi sinar gamma terhadap pertumbuhan kalus dan regenerasinya padi var. Ciherang, Inpari 13 dan Inpara 3. Percobaan ketiga adalah penentuan nilai LC50 NaCl pada kalus beberapa varietas padi. Hasil dari percobaan tersebut digunakan sebagai dasar untuk melaksanakan percobaan keempat adalah pembentukan genotipe padi toleran salinitas dari varietas Ciherang, Inpari 13 dan Inpara 3. Nomor-nomor putatif mutan somaklon yang dihasilkan selanjutnya digunakan sebagai bahan tanaman untuk percobaan studi mekanisme toleransi salinitas padi pada kondisi
in vitro dan rumah kaca dan seleksi putatif mutan somaklon pada kondisi in vitro
dan rumah kaca.
Percobaan studi regenerasi tunas adventif dari menggunakan kalus 5 varietas padi. Penelitian ini terdiri dari 4 kegiatan yaitu induksi kalus embriogenik, regenerasi tunas adventif, multiplikasi tunas, induksi akar. Perlakuan untuk induksi kalus embriogenik adalah MS + 2,4-D (0, 1, 3,5 dan 7 mg/l) + Casein hidrolisat 3 g/l. Perlakuan untuk regenerasi tunas adalah perlakuan MS + BA (0, 1 dan 5 mg/l) + Zeatin (0, 01 dan 0,3 mg/l) prolin 100 mg/l. Perlakuan untuk multiplikasi tunas adalah MS + Thidiazuron (0, 0,1 dan 0,3 mg/l) dan perlakuan untuk perakaran adalah MS + IBA (0, 1, 2 dan 3 mg/l). Hasil penelitian adalah formulasi media induksi kalus embriogenik dari embrio zigotik varietas Ciherang, Inpari 13 dan Pokkali adalah MS + 2,4-D 3mg/l + Chasein Hidrolisat 3 g/l, untuk varietas Inpara 3 dan IR 29 adalah MS + 2,4-D 5 mg/l Chasein Hidrolisat 3 g/l. Media regenerasi tunas adventif pada padi varietas Ciherang, Inpari 13 dan Pokkali adalah MS + BA 3 mg/l + Zeatin 0,3 mg/l + prolin 100 mg/l, dan untuk varietas Inpari 3 dan IR 29 adalah MS + BA 3 mg/l + Zeatin 0,1 mg/l. Media multiplikasi dan perakaran pada semua varietas berturut turut adalah MS + Thidiazuron 0,3 mg/l, MS +IBA 2 dan 3 mg/l.
Pada percobaan penentuan nilai LC50 NaCl pada kalus beberapa varietas padi bertujuan untuk mengetahui nilai LC50 NaCl. Perlakuannya yang diberikan yaitu konsentrasi NaCl (0, 50, 100, 150 dan 200 mM). Hasil yang peroleh adalah nilai LC50 NaCl dari kalus Ciherang adalah 85,79 mM, Inpari 13 adalah 90,91 mM, Inpara 3 adalah 91,77 mM dan, IR29 adalah 70,67 mM.
Tujuan percobaan pembentukan genotipe padi toleran salinitas dari varietas Ciherang, Inpari 13 dan Inpara 3 adalah mendapatkan nomor-nomor mutan somaklon padi toleran salinitas hasil mutasi induksi dan seleksi in vitro. Varietas yang digunakan adalah Ciherang, Inpari 13, Inpara 3. Penelitian ini terdiri 4 kegiatan utama yaitu induksi kalus embriogenik, induksi mutasi dan seleksi in vitro, regenerasi tunas, aklimatisasi. Hasil dari penelitian adalah diperoleh 29 nomor putatif mutan somaklon yang berasal dari padi varietas Ciherang. 39 nomor yang berasal dari padi varietas Inpari 13 dan 42 nomor yang berasal dari padi varietas Inpari 13.
Tujuan dari percobaan studi mekanisme toleransi salinitas padi pada kondisi in vitro dan rumah kaca dan seleksi putatif mutan somaklon pada kondisi in vitro dan rumah kaca adalah untuk mendapatkan nomor-nomor mutan somaklon padi toleran salinitas yang telah diseleksi secara in vitro dan rumah kaca dan mendapatkan informasi mekanisme toleransi salinitas tanaman padi pada kondisi in vitro dan rumah kaca. Percobaan ini terdiri dari dua kegiatan yaitu seleksi toleransi cekaman salinitas putatif mutan somaklon berdasarkan SES (Standard Evaluation Score) dan studi mekanisme toleransi salinitas hasil seleksi in vitro. Hasil penelitian adalah diperoleh nomor-nomor putatif mutan somaklon padi toleran salinitas hasil seleksi in vitro dan di rumah kaca yaitu CH-30 dan CH-4-2 (asal Ciherang); II-13-42, II-13-7, II-13-10, II-13-13 II-13-2 (asal Inpari 13) dan IA-3-21 (asal Inpara 3). Mekanisme toleransi salinitas pada nomor-nomor putatif mutan somaklonal yang toleran terhadap NaCl hasil diseleksi secara in vitro maupun di rumah kaca menunjukkan mekanisme yang sama. Beberapa parameter yang diuji memperlihatkan hasil akumulasi prolin yang lebih tinggi, kandungan K, Mg dan Ca pada daun yang cenderung tetap dan kandungan Na yang lebih rendah, kerapatan stomata yang lebih renggang dan perbandingan panjang dan lebar stomata lebih besar. Uraian diatas menunjukkan putatif mutan somaklon padi yang dihasilkan memiliki mekanisme toleransi ekslusi.
SUMMARY
ROSSA YUNITA. Salinity Tolerant Rice Development Through In Vitro Mutation and Selection: Mechanism of Tolerance Physiology. Supervised by NURUL KHUMAIDA, DIDY SOPANDIE and IKA MARISKA.
To meet the demand for rice in Indonesia, is necessary to improve the productivity of rice plants through the expansion of harvest area to use marginal land such as saline soils. The problem is the availability of very limited adaptive varieties. The use of saline-tolerant varieties are more profitable because it will reduce production costs. It is necessary for new varieties to be able adapt in saline soil
The main objective of this study was to obtain numbers of salinity tolerant mutants somaclone. This study consists of six main experiment. The first experiment is a study of adventitious shoot regeneration from callus several rice varieties. The second experiment is the effect of gamma irradiation on the growth of callus and regeneration of rice var. Ciherang, Inpari 13 and Inpara 3. The third experiment is to determine the LC50 value of NaCl on some varieties of rice callus. Results of these experiments are used as the basis for implementing the fourth experiment was the formation of salinity tolerant rice genotypes of Ciherang, Inpari 13 and Inpara 3. The numbers putative mutants generated is then used as plant material for experiment study of salinity tolerance mechanisms of rice in vitro conditions and greenhouse and selection of putative mutants somaclone in vitro conditions and greenhouse
Experiment study of regeneration of adventitious buds of using 5 rice varieties callus. This study consists of four activities; they are embryogenic callus induction, regeneration of adventitious buds, shoots multiplication and root induction. Embryogenic callus induction was treated MS + 2,4-D (0, 1, 3.5 and 7 mg/l) + Casein hydrolyzate 3 g/l. Regeneration of shoots was treated MS + BA (0, 1 and 5 mg/l) + Zeatin (0, 01 and 0.3 mg/l) Proline 100 mg/l. Shoot multiplication was treated MS + Thidiazuron (0, 0.1 and 0.3 mg/l) and rooting was treated MS + IBA (0, 1, 2 and 3 mg/l). Results of the study are embryogenic callus induction media formulations of zygotic embryos Ciherang, Inpari 13 and Pokkali wasMS + 2,4-D 3mg/l + Chasein hydrolyzate 3 g/l, for a variety Inpara 3 and IR 29 was MS + 2.4 -D 5 mg/l Chasein hydrolyzate 3 g/l. Media regeneration of adventitious LD50 value of callus 5 rice varieties. The treatment in this experiment is a gamma-ray irradiation at a dose of 10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 Gy. The results obtained in this experiment in the LD50 value for callus Ciherang was 22.468 Gy, for Inpari 13 was 23.124 Gy and Inpara 3 was 21.0305 Gy.
obtained are LC50 values of callus Ciherang NaCl is 85.79 mM, Inpari 13 is 90.91 mM, Inpara 3 is 91.77 mM and, IR29 is 70.67 mM.
The purpose of this experiment of studies the mechanism of salinity tolerance of rice at in vitro conditions and greenhouse and selection of putative mutants somaclone at in vitro conditions and greenhouse are get the numbers of mutant somaclone tolerant salinity and to get information mechanisms of tolerance salinity of rice at in vitro and greenhouse conditions . This experiment consists of two activities, the selection of salinity tolerance putative mutant somaclone based SES (Standard Evaluation Score) and the study of mechanisms of salinity tolerance in vitro selection results. The research resulted in obtained putative numbers salinity tolerant rice mutant somaclone selection results in vitro and in the greenhouse is the CH-30 and CH-4-2 (Ciherang origin); 42, II-13-7, II-13-10, II-II-13-2 13-13 (derivat from Inpari 13) and IA-3-21 (derivat from Inpara 3). Salinity tolerance mechanism in numbers putative mutant somaclon tolerant NaCl selected resulted in vitro and in greenhouses showed the same mechanism. The resulted of parameters showed higher proline accumulation, content of K, Mg and Ca in the leaves tend to remain and lower Na content, stomatal density of the lighter and the length and width ratio greater stomata. The Description above shows the putative mutant rice produced somaclone have tolerance mechanisms of exclusion.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
PENGEMBANGAN PADI TOLERAN SALINITAS
MELALUI MUTASI DAN SELEKSI IN VITRO
:MEKANISME FISIOLOGI TOLERANSI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
Penguji pada Ujian Tertutup:
1. Dr. Ir. Hajrial Aswidinoor. M.Sc.
Staf Pengajar Pada Departemen Agronomi dan Hortikultura, Fakultas Pertanian, IPB
2. Dr. Ir. Ali Jamil, M.P
Kepala Balai Besar Penelitian Tanaman Padi. Badan Litbang Pertanian, Kementrian Pertanian
Penguji pada Ujian Terbuka:
1. Dr. Ir. Ali Jamil, M.P.
Kepala Balai Besar Penelitian Tanaman Padi. Badan Litbang Pertanian, Kementrian Pertanian
2. Prof. Dr. Ir. Sudirman Yahya, M.Sc.
Staf Pengajar Pada Departemen Agronomi dan Hortikultura, Fakultas Pertanian, IPB
HALAMAN PENGESAHAN
Judul Penelitian : Pengembangan Padi Toleran Salinitas Melalui Mutasi dan Seleksi In Vitro : Mekanisme Fisiologi Toleransi
Nama : Rossa Yunita
NRP : A 263120051
Disetujui:
Ketua Komisi Pembimbing
Dr. Ir. Nurul Khumaida, MSi Ketua
Prof. Dr. Ir. Didy Sopandie, M.Agr Anggota
Prof. (R). Dr. Ika Mariska Anggota
Diketahui
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dekan Sekolah Pascasarjana IPB
Dr. Ir. Yudiwanti Wahyu E K, MS Dr. Ir. DahruI Syah, MSc, Agr.
Tanggal Ujian Tertutup : 30 Juli 2015 Tanggal Lulus: 19 Oktober 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini ialah Salinitas, dengan judul : Pengembangan Padi Toleran Salinitas Melalui Mutasi dan Seleksi In Vitro : Mekanisme Fisiologi Toleransi
Penelitian dan penulisan disertasi ini dibimbing oleh Dr. Nurul Khumaida, MS. selaku ketua komisi pembimbing dan dua Angota Komisi Pembimbing yaitu Prof. Dr. Ir. Didy Sopandie, M.Agr dan Prof (R). Dr. Ika Mariska. Pada
kesempatan ini penulis menyampaikan terimakasih dan penghargaan yang tulus atas waktu dan kesempatan yang telah diluangkan dalam mengarahkan dan membimbing penulis.
Bagian dari disertasi ini telah dipublikasikan pada Jurnal Research Bioscience vol: 11 no1 halaman 04-09 tahun 2014 dengan judul “Growth and regeneration of rice (Oryza sativa L.) callus in salt medium” dan Jurnal
Agrobiogen vol: 28 no: 3 halaman 101-108 dengan judul “Pengaruh Iradiasi sinar gamma terhadap pertumbuhan dan regenerasi kalus padi varietas Ciherang dan Inpari 13”.
Penelitian dan penyelesaian disertasi ini didanai oleh Program Dana Bantuan Penelitian Petugas Belajar Badan Litbang Pertanian, karena itu penulis menyampaikan banyak terimakasih kepada Badan Litbang Pertanian, Kementrian Pertanian. Ucapan terimakasih Penulis sampaikan kepada Kepala Balai Besar BB-Biogen yang telah memberikan fasilitas untuk penelitian.
Terimakasih penulis ucapkan kepada teman–teman Peneliti di BB-Biogen pada umum dan khusunya di Kelti Biologi Sel dan Jaringan. Ungkapan terima kasih juga disampaikan setinggi-tingginya kepada Ayahanda H. Asri Harun, Ibunda Hj. Rosliani, Suami tercinta Abdul Aziz, SSos, MSi, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Oktober 2015
DAFTAR ISI
Pengaruh Kondisi Salin Terhadap Proses Fisiologi Tanaman 5
Mekanisme Fisiologi Toleransi Salinitas Pada Tanaman 6
Mutasi Induksi 6
Seleksi In vitro 7
3. REGENERASI TUNAS ADVENTIF DARI KALUS BEBERAPA VARIETAS PADI
4. PENGARUH IRADIASI SINAR GAMMA TERHADAP PERTUMBUHAN KALUS DAN REGENERASINYA PADI
6. PEMBENTUKAN GENOTIPE PADI TOLERAN SALINITAS PADI VARIETAS CIHERANG, INPARI 13 DAN INPARA 3
36
Abstrak 36
Abstract 36
Pendahuluan
Metode Penelitian 37
Hasil dan Pembahasan 38
Kesimpulan 42
7. STUDI MEKANISME TOLERANSI SALINITAS PADI PADA
KONDISI IN VITRO
43
Abstrak 43
Abstract 43
Pendahuluan 43
Metode Penelitian 44
Hasil dan Pembahasan 47
Simpulan 66
8. SELEKSI PUTATIF MUTAN SOMAKLON PADI DI RUMAH
KACA DAN MEKANISME TOLERANSI SALINITAS
67
Abstrak 67
Abstract 67
Pendahuluan 67
Metode Penelitian 68
Hasil dan Pembahasan 69
Simpulan 92
9. PEMBAHASAN UMUM 93
10. SIMPULAN DAN SARAN 97
DAFTAR TABEL
Nomor Halaman 1 Pengaruh konsentrai 2,4-D terhadap pembentukan kalus pada
beberapa varietas padi, umur 8 minggu setelah kultur... 12 2. Regenerasi kalus embriogenik membentuk tunas adventif
beberapa varietas padi (Ciherang, Inpari 13, Inpara 3, Pokkali
dan IR 29), umur 8 minggu setelah kultur... 14 3. Rerata jumlah tunas yang terbentuk pada media yang
mengandung Thidiazuron dari bebera varietas padi, umur 2
minggu setelah kultur .. ... 16 4. Rerata jumlah dan panjang akar yang terbentuk pada media
yang mengandung IBA dari berberapa varietas padi, umur 2
minggu... 17 5. Regenerasi kalus padi membentuk tunas adventif dari kalus
padi varietas Ciherang setelah perlakuan iradiasi sinar gamma 25 6. Regenerasi kalus padi membentuk tunas adventif varietas
Inpari 13 setelah perlakuan iradiasi sinar gamma... 26 7. Regenerasi tunas dari kalus padi varietas Inpara 3 yang telah
diberi perlakuan iradiasi sinar gamma ... 27 8. Persentase kalus padi mencoklat pada media yang mengandung
NaCl dengan konsentrasi berbeda ... 31 9. Nilai lethal konsentrasi 50 (LC50) beberapa kalus varietas padi.. 32 10. Persentase jumlah kalus membentuk tunas dan jumlah tunas per
kalus padi ... 34 11. Persen kalus dan kalus embriogenik padi yang terbentuk dari
eksplan embrio zigotik dewasa... 39 12. Seleksi in vitro kalus mutan pada media yang mengandung
NaCl dengan konsentrasi LC50 padi... 40 13. Regenerasi kalus hasil seleksi membentuk spot hijau dan tunas
adventif padi ... 41 14. Aklimatisasi planlet padi putatif mutan somaklon hasil seleksi
in vitro padi ... 41 15. Kriteria skoring berdasarkan Modifikasi SES (IRRI 1997) ... 45 16. Hasil skoring beberapa varietas padi dan nomor-nomor putatif
mutan somaklon yang di seleksi secara in vitro... 46 17. Kandungan Kalium pada daun tiga varietas padi dan putatif
mutan somaklonnya yang diseleksi secara in vitro... 50 18. Kandungan Kalium (%) pada akar tiga varietas padi dan putatif
mutan somaklonnya... 51 19. Kandungan Natrium (%) pada daun tiga varietas padi dan putatif
mutan somaklonnya ... 53 20. Kandungan Natrium (%) pada akar tiga varietas padi dan putatif
mutan somaklonnya ... 54 21. Nilai rasio K/Na pada daun tiga varietas padi dan putatif mutan
somaklonnya ... 56 22. Kandungan kalsium (%) pada daun dari tiga varietas padi dan
23. Kandungan Kalsium (%) pada akar tiga varietas padi dan putatif
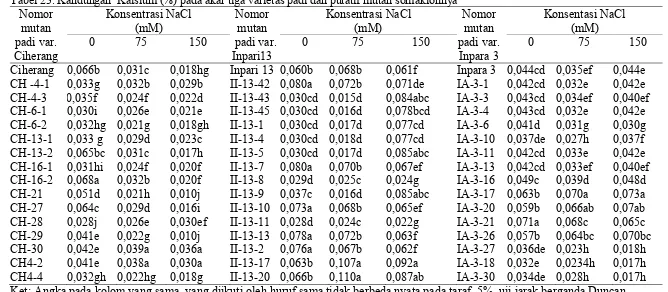
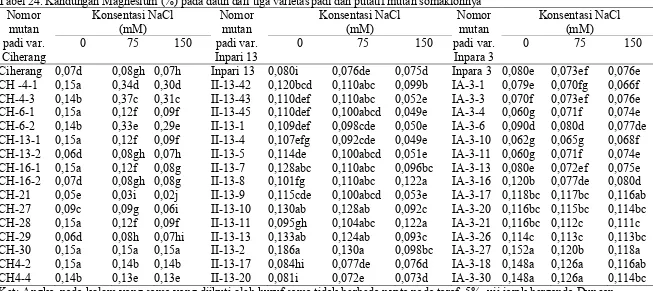
mutan somaklonnya ... 59 24. Kandungan Magnesium (%) pada daun dari tiga varietas padi dan
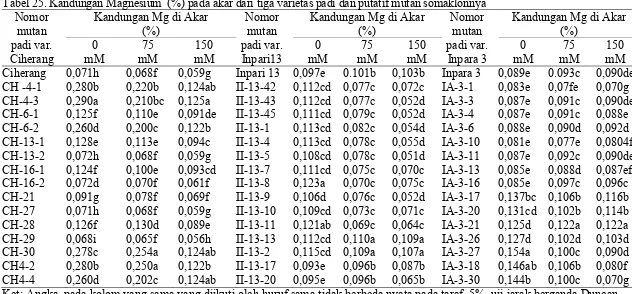
putatif mutan somaklonnya ... 61 25. Kandungan Magnesium (%) pada akar dari tiga varietas padi dan
putatif mutan somaklonnya ... 62 26. Kerapatan stomata pada tiga varietas padi dan putatif mutan
somaklonnya pada pembesaran 40 x 10 kali ... 64 27. Perbandingan panjang dan lebar stomata pada tiga varietas padi
dan putatif mutan somaklonnya pada pembesaran 40 x 10 kali... 65 28. Hasil skoring beberapa varietas padi dan nomor-nomor putatif
mutan somaklon hasil seleksi di rumah kaca... 70 29. Kandung prolin (mg/g) pada daun dari ketiga varietas padi
(Ciherang, Inpari 13 dan Inpara 3) dan putatif mutan somaklon.... 72 30. Kandungan Kalium (%) pada daun tiga varietas padi dan putatif
mutan somaklonnya ... 74 31. Kandungan Kalium (%) pada akar tiga varietas padi dan putatif
mutan somaklonnya ... 75 32. Kandungan Natrium (%) pada daun tiga varietas padi dan putatif
mutan somaklonnya... 77 33. Kandungan Natrium (%) pada akar tiga varietas padi dan putatif
mutan somaklonnya... 78 34. Nilai rasio K/Na pada daun tiga varietas padi dan putatif mutan
somaklonnya hasil seleksi di rumah kaca... 81 35 Kandungan Kalsium (%) pada daun dari tiga varietas padi dan
putatif mutan somaklonnya... 83 36 Kandungan Kalsium (%) pada akar tiga varietas padi dan putatif
mutan somaklonnya ... 84 37 Kandungan Magnesium (%) pada daun dari tiga varietas padi
dan putatif mutan somaklonnya... 86 38 Kandungan Magnesium (%) pada akar dari tiga varietas padi dan
putatif mutan somaklonnya... 87 39 Kerapatan stomata pada tiga varietas padi dan putatif mutan
somaklonnya pada pembesaran 40 x 10 kali... 89 40 Perbandingan panjang dan lebar stomata pada tiga varietas padi
DAFTAR GAMBAR
Gambar Halaman
1. Bagan alur penelitian 4
2. Kalus embriogenik padi Varietas Ciherang (a) Inpari 13 (b)
dan Inpara 3 (c) ... 13 3. Regenerasi kalus embriogenik membentuk kalus nodular
dan tunas beberapa varietas padi (a) Ciherang (b) Inpari
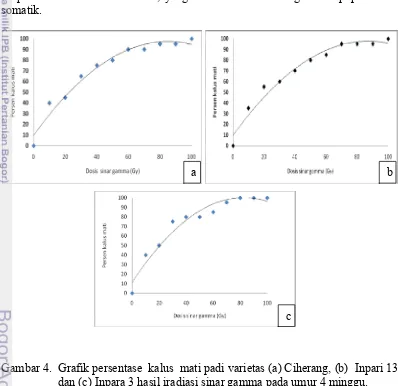
13 (c) Inpara 3 (d) Pokkali (e) IR 29 ... 13 4. Grafik persentase kalus mati padi varietas (a) Ciherang
(b) Inpari 13 dan (c) Inpara 3 hasil iradiasi sinar gamma
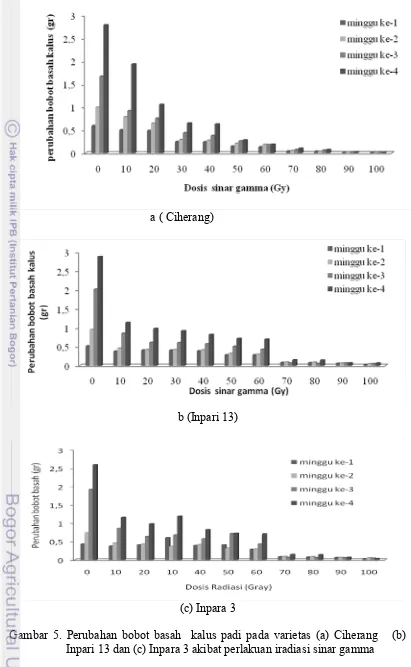
pada umur 4 minggu... 21 5. Perubahan bobot basah kalus padi pada varietas (a)
Ciherang dan (b) Inpari 13 dan (c) Inpara 3 akibat
perlakuan iradiasi sinar gamma ... 23 6. Penampilan kalus padi Inpari 13 setelah diberi perlakuan
iradiasi sinar gama (a) 0 Gy, (b) 20 Gy dan (c) 100 Gy ... 24 7. Perubahan berat basah kalus padi akibat perlakuan NaCl
pada padi varietas (a) Ciherang, (b) Inpari 13, (c)
Inpara 3, (d) Pokkali dan (e) IR 29 ... 33 8. Kalus embriogenik dari embrio zigotik padi varietas (a)
Ciherang, (b) Inpari 13 dan (c) Inpara 3 ... 39 9. Respon kalus akibat pemberian Iradiasi sinar gamma dan
seleksi pada media NaCl . (a) Ciherang, (b) Inpari 13, (c)
Inpara 3 ... 40 10. Proses aklimatisasi nomor-nomor putatif mutan
somaklon. (a) Planlet di kultur pada media aquades, (b) Planlet di kultur pada media yosida, (c) Planlet di kultur
pada media lumpur ... 42 11. Keragaan tanaman tetua yang di kulturkan pada media
mengandung NaCl. (a) 0 mM NaCl, (b) 75 mM NaCl dan
(c) 150 mM NaCl ... 47 12. Kandung prolin (mg/g) pada daun padi varietas Ciherang,
dan putatif mutan somaklon ... 48 13. Kandung prolin (mg/g) pada daun padi varietas Inpari 13
dan putatif mutan somaklon... 48 14. Kandung prolin (mg/g) pada daun padi varietas Inpara 3
dan putatif mutan somaklon... 49 15. Keragaan putatif mutan somaklon yang bersifat peka
(CH-13-2, II-13-17, IA-3-1) dan yang bersifat toleran
(CH-4-2, II-13-42, IA-3-21) ... 69 16. Kandungan Kalium (K) pada daun pada varietas padi dan
putatif mutan somaklonya (A) Ciherang, (B) Inpari 13,
(C)Inpara 3 ... 76 17. Kandungan Natrium pada daun beberapa varietas padi dan
nomor putatif mutan somaklon (A) Ciherang, (B) Inpari
padi dan nomor putatif mutan samaklonya (A) Ciherang
(B) Inpari 13, (C) Inpara 3 ... 82 19. Kandungan Magnesium pada daun beberapa varietas padi,
dan nomor putatif mutan samaklonya (A) Ciherang, (B)
Inpari 13, (C) Inpara 3 ... 85 20 Keragaan stomata daun padi putatif mutan somaklon yang
bersifat peka yaitu (a) CH-13-2, (b) II-13-17, (c) IA-3-1 dan yang bersifat toleran yaitu (d) CH-4-2, (e) II-13-42,
(f) IA-3-21... 91 21 Kandungan prolin pada daun padi varietas (A) IR 29 dan
(B)Pokkali yang diseleksi dengan NaCl di rumah kaca
I PENDAHULUAN
Latar Belakang
Padi (Oryza sativa L) merupakan komoditas yang penting bagi rakyat Indonesia karena berperan sebagai sumber bahan pangan utama (Staple food). Kebutuhan akan beras sebagai bahan pangan pokok terus meningkat sejalan dengan meningkatnya jumlah penduduk dan kesejahteraan masyarakat. Pada tahun 2014 kebutuhan masyarakat Indonesia akan komoditas ini sebesar 124 kg/orang/tahun dan produksi padi nasional hanya mencapai 70,8 juta ton (Badan Pusat Statistik Republik Indonesia 2015). Untuk memenuhi kebutuhan masyarakat Indonesia akan komoditas ini perlu upaya peningkatan produksi beras nasional yang dinamis.
Pada kondisi seperti itu, peluang yang dapat dilakukan untuk meningkatkan produksi padi di Indonesia adalah dengan meningkatkan pemanfaatan lahan sub optimal seperti lahan sawah tadah hujan, lahan kering, dan lahan salin. Lahan salin di Indonesia cukup luas sehingga berpotensi dikembangkan sebagai sarana perluasan areal panen padi, dengan luas lahan salin bertipe gambut sekitar 0,44 juta ha (Alihamsyah et al. 2002). Total luas tanah salin di Indonesia belum diketahui secara tepat karena Indonesia terdiri dari pulau-pulau. Luas lahan salin diperkirakan akan terus bertambah terutama di daerah pesisir pantai karena terjadinya perubahan iklim global dan naiknya permukaan air laut (Ismail 2007).
Data dari FAO menunjukan hampir 50% lahan irigasi mengalami masalah salinitas dan akibatnya setiap tahun beberapa ratus hektar sawah ditinggalkan. Di Indonesia saat ini salinitas tanah telah menjadi suatu masalah serius terutama di daerah sentra produksi padi, yang terletak di pinggir laut seperti daerah pantai utara Jawa, Sulawesi Selatan dan pantai Sumatra, khususnya pantai Timur dan Barat Nanggroe Aceh Darussalam dimana masalah salinitas ditimbulkan karena bencana tsunami (FAO 2005).
Salinitas tinggi mempengaruhi tanaman dalam dua hal, yaitu efek osmotik dan efek cekaman ion (Munns 2002). Cekaman osmotik, yaitu terjadinya penurunan potensial osmotik larutan tanah sehingga mengurangi ketersediaan air (Sopandie 2014). Konsentrasi garam yang tinggi di dalam tanah mengganggu kapasitas akar untuk mengambil air, kondisi tersebut menyebabkan dehidrasi parsial sel dan hilangnya turgor sel karena berkurangnya potensial air di dalam sel (Yuniati 2004). Efek selanjutnya toksisitas dimana terjadi peningkatan konsentrasi ion yang bersifat racun bagi tanaman. Efek cekaman ion ini yang merupakan faktor utama yang dapat menghambat pertumbuhan padi (Yang et al.
2008). Akumulasi Na+ dan Cl+ yang berlebihan dalam sitoplasma menyebabkan perubahan metabolisme dalam sel mengakibatkan penghambatan proses fisiologis dan biokimia, sehingga pertumbuhan dan perkembangan tanaman akan terhambat (Munns & Tester 2008). Pada tanaman padi keracunan garam dapat ditandai dengan terhambatnya pertumbuhan, berkurangnya anakan, hingga akhirnya akan menurunkan produksi gabah mencapai 50%. (Hakim et al. 2012, Aref & Rad 2012 , Rad et al. 2012).
dan kultur in vitro dapat meningkatkan toleransi tanaman terhadap salinitas dan kekeringan dalam waktu relatif lebih cepat dibandingkan menggunakan pemuliaan konvensional. Metode ini juga sudah dicobakan untuk merakit tanaman toleran pada kondisi salin, seperti pada kentang (Sabbah & Tal 1990), gandum (Barakat & Abdel-latif 1996), dan bunga matahari (Alvares et al. 2003).
Margava (1986) menyatakan bahwa mekanisme fisiologi toleransi populasi sel dan plantlet terhadap cekaman pada kondisi in vitro merupakan interaksi dari karakter biokimia, fisiologi dan morfologi. Untuk mengetahui mekanisme adaptasi fisiologi kondisi in vitro dapat dilakukan dengan melakukan analisis rasio K+/Na+, kandungan ABA maupun senyawa osmotik yang kompatibel seperi gula, prolin, poliol dan glisin betain.
Teknik seleksi in vitro telah banyak dimanfaatkan untuk mengembangkan tanaman toleran salinitas, akan tetapi belum banyak dipelajari tentang mekanisme toleransi salinitas tanaman padi pada kultur in vitro. Pada penelitian ini dipelajari mekanisme toleransi tanaman padi pada kultur in vitro, khususnya pada fase plantlet. Dengan diketahuinya mekanisme fisiologis yang terjadi pada kondisi cekaman in vitro maka dapat dilakukan penapisan dini pada populasi sel somatik dan plantlet sehingga akan mempercepat perolehan galur-galur harapan baru yang toleran salinitas.
Studi mengenai mekanisme tolerasi terhadap cekaman dalam kondisi in vitro belum banyak dilakukan. Penelitian ini merupakan suatu inovasi yang dapat memberikan kontribusi dalam pembangunan pertanian khususnya untuk pengembangan tanaman pangan di sub optimal.
Mekanisme toleransi tanaman di lapang pada kondisi salin sangat bervariasi. Sebelum melakukan kegiatan pemulian untuk menghasilkan varietas-varietas yang toleran salinitas, perlu dipahami dahulu tentang mekanisme toleransi salinitas pada tanaman tersebut. Hal ini penting untuk menetapkan metode untuk merakit varietas dan menentukan karakter seleksi yang akan di gunakan dalam kegiatan pemuliaan.
Mekanisme toleransi tanaman terhadap kondisi salinitas meliputi mekanisme ekslusi dan inklusi (Marscher 2012). Mekanisme ekslusi adalah mekanisme toleransi untuk mencegah defisit air secara internal dengan cara mensintesis solut organik dan meningkatkan sukulensi pada daun. Mekanisme inklusi merupakan mekanisme toleransi salinitas dengan mencegah toksisitas ion-ion melalui jaringan yang mempunyai toleransi tinggi terhadap salinitas (Sopandie 2014). Mekanisme ini diantaranya dengan cara kompartementasi garam, sintesis solut kompatibel, ekskresi garam. Respon mekanisme toleransi salinitas pada tanaman padi sangat bervariasi dapat inklusi atau ekslusi ( Hu et al. 2012).
yang dilepas tahun 2009. Varietas ini memiliki keunggulan di samping produktivitas tanaman yang tinggi dengan rata-rata hasil panen sebesar 6,59 t/ha atau setara dengan potensi hasil 8,0 t/ha, varietas ini juga mempunyai umur yang sangat genjah (sekitar 103 hari) (Balai Besar Penelitian Tanaman Padi, 2009). Inpara 3 merupakan padi rawa yang mempunyai keungulan toleran rendaman selama 6 hari dan toleran terhadap Fe dan Al. Tanaman ini memiliki fenotipe yang lebih baik dari pada varietas yang telah dilepas sebelumnya. Verietas Pokkali merupakan varietas yang toleran pada kondisi salin sehingga dimanfaatkan sebagai kontrol toleran, sedangkan untuk kontrol peka umumnya digunakan IR 29.
Perumusan Masalah
Peningkatan produktivitas tanaman dapat ditempuh antara lain melalui perluasan areal panen antara lain pada lahan marginal, sehingga untuk mencapai tujuan tersebut diperlukan teknologi budidaya dan varietas yang adaptif. Masalah yang dihadapi saat ini antara lain ketersediaan varietas yang adaptif sangat terbatas. Pengunaan varietas yang toleran terhadap tanah salin akan lebih menguntungkan untuk jangka panjang. Di samping itu dengan tersedianya varietas yang adaptif tidak memerlukan biaya yang tinggi untuk pengelolaan tanah salin. Dengan demikian perlu teknologi terobosan untuk menghasilkan varietas baru yang dapat beradaptasi di tanah salin.
Mutasi induksi yang dikombinasi dengan teknik seleksi in vitro merupakan salah metode alternatif untuk merakit varietas yang toleran pada kondisi salin. Dengan mengunakan teknik tersebut pada penelitian ini dihasilkan nomor nomor putatif mutan somaklon toleran salinitas.
Pada penelitian ini juga akan dipelajari bagaimana mekanisme toleransi tanaman mutan padi yang dihasilkan, pada kondisi pertumbuhan di lapangan (setelah diaklimatisasi). Serta akan dilakukan perbandingan, apakah mekanisme toleransi yang terjadi pada kondisi lingkungan in vitro akan sama dengan mekanisme toleransi yang terjadi pada lingkungan ex vitro setelah aklimatisasi.
Tujuan Penelitian
Tujuan umum penelitian ini adalah untuk mendapatkan nomor-nomor mutan somaklon padi toleran salinitas dan secara khusus penelitian ini bertujuan untuk: 1. Mendapatkan metode regenerasi kalus membentuk plantlet beberapa varietas
padi yaitu Ciherang, Inpari 13, Inpara 3, Pokkali dan IR 29.
2. Mendapatkan Nilai LD50 iradiasi sinar gamma dari kalus padi varietas Ciherang, Inapari 13 dan Inpara 3
3. Mendapatkan nilai LC50 NaCl dari kalus padi varietas Ciherang, Inpari 13 dan Inpara 3
4. Mendapatkan informasi mekanisme toleransi salinitas tanaman padi pada kondisi in vitro
5. Mendapatkan informasi mekanisme toleransi salinitas tanaman padi pada kondisi di rumah kaca
Manfaat Penelitian
2. Diperoleh informasi mekanisme toleransi salinitas tanaman padi pada kondisi
in vitro dan di rumah kaca.
Kebaruan
1. Metode kombinasi mutagen fisik dan seleksi in vitro untuk menghasilkan putatif mutan somaklon padi yang toleran salinitas dengan mengunakan LD50 radiasi sinar gamma dan LC 50 NaCl.
2. Keeratan hubungan mekanisme toleransi cekaman salinitas yang diuji pada kondisi in vitro dan di rumah kaca.
Ruang Lingkup Penelitian
Penelitian ini terdiri atas 6 percobaan yaitu: (1) studi regenerasi tunas adventif dari kalus beberapa varietas padi, (2) Pengaruh irradiasi sinar gamma terhadap pertumbuhan dan regenerasi kalus padi varietas Ciherang, Inpari 13 dan Inpara 3, (3) Penentuan nilai LC50 NaCl pada kalus beberapa varietas padi, (4) pembentukan fenotipe padi toleran salinitas padi varietas Ciherang, Inpari 13 dan Inpara 3, (5) studi mekanisme toleransi salinitas padi pada kondisi in vitro dan (6) seleksi putatif mutan somaklon padi di rumah kaca dan mekanisme toleransi salinitas. Secara jelas ruang lingkup penelitian disampaikan pada Gambar1.
2 TINJAUAN PUSTAKA
Pengaruh Kondisi Salin Terhadap Proses Fisiologi Tanaman
Salinitas dapat didefinisikan sebagai adanya kandungan garam yang terlarut dalam konsentrasi berlebih pada larutan tanah, sehingga dapat mengganggu pertumbuhan dan perkembangan tanaman ( Munns & Tester, 2008). Pengaruh salinitas pada tanaman mencakup tiga aspek, yaitu: (1) keracunan yang disebabkan kandungan garam yang berlebih, dimana konsentrasi NaCl yang tinggi dapat merusak permeabilitas membran sel (Carden et al. 2003, (2) peningkatan potensial osmotik pada larutan tanah karena bertambahnya konsentrasi garam didalam larutan tanah dapat meningkatkan potensial osmotik larutan tanah. Oleh sebab itu salinitas dapat menyebabkan tanaman sulit menyerap air sehingga terjadi kekeringan fisiologis (Munns & Tester 2008); (3) Penurunan dalam penyerapan unsur-unsur hara penting bagi tanaman (FAO 2005). Kondisi salinitas akan mengurangi ketersediaan K+ dan Ca++ dalam larutan tanah dan menghambat proses transportasi kedua unsur hara tersebut kedaerah pertumbuhan tanaman sehingga akan mengurangi kualitas pertumbuhan baik organ vegetatif maupun reproduktif (Carden et al. 2003).
Pengaruh salinitas pada tanaman padi memberikan pengaruh yang berbeda pada setiap fase pertumbuhan. Pada fase perkecambahan, kondisi cekaman salinitas dapat menurukan kemampuan benih untuk berkecambah sehingga persentase bibit yang dihasilkan berkurang serta pertumbuhan bibit juga terhambat (Hakim et al. 2012). Kondisi salin pada fase tillering dan fase inisiasi malai menunjukkan penurunan pada jumlah gabah permalai. Sedangkan pada pada fase generatif, salinitas dapat mempengaruhi proses pengisian malai sehingga jumlah gabah hampa meningkat. Secara umum kondisi salin dapat menurunkan produksi padi (Rad et al. 2012).
Mekanisme Fisiologi Toleransi Salinitas pada Tanaman
Toleransi salinitas untuk tanaman yang dibudidayakan ditanah salin mengacu pada bagaimana tanaman tersebut memiliki kemampuan untuk bertahan dan berproduksi dengan baik bila ditanam pada lahan dengan kondisi salin. Menurut Marschner (2012) mekanisme toleransi salinitas pada tanaman meliputi mekanisme inklusi dan ekslusi.
Mekanisme inklusi, yaitu suatu mekanisme yang dialami oleh tanaman dimana tanaman tersebut mencegah terjadinya keracunan ion-ion garam dengan cara sintesis solut kompatibel, kompertementasi garam dalam vakuola serta retranslokasi garam melalui floem, eksresi garam dengan menggugurkan daun daun tua (Marschner 2012).
yang terletak di membran vakuola, H + akan ditransport ke luar dari dalam vakuola sementara Na + diangkut ke dalam vakuola. Cara ini akan menyebabkan Na + terakumulasi dalam vakuola. Kekuatan transfer berasal dari gradien konsentrasi proton dan potensial elektrokimia ( Bing et al. 2008, Ren et al.
2005).
Pada kondisi cekaman salinitas, tanaman mudah kehilangan air dan pada kondisi ekstrim dapat menyebabkan kematian. Tekanan osmotik dilingkungan bercekaman salinitas lebih tinggi dari pada tekanan osmatik dalam sel tanaman, hal ini akan mengakibatkan tanaman sulit menyerap air, sehingga mengakibatkan fenomena defisit air. Untuk menghindari dehidrasi dan menjaga keseimbangan potesnial air, sel tanaman akan meningkatkan konsentrasi zat terlarut intraseluler dan mengurangi potensi osmotik intraseluler untuk mengatur beda potensial osmotik antara sel. Untuk mempertahankan lingkungan sitoplasma yang stabil, tanaman mengatur konsentrasi relatif dari ion anorganik (terutama K + dan Na +) untuk menyesuaikan turgor sel, volume sel, nilai pH intraseluler, dan parameter fisiologis penting lainnya (Demidchik et al. 2002). Tanaman juga menghasilkan senyawa osmotik kompatibel sitoplasma seperti prolin, glisin, gula alkohol dan lain-lain untuk menjaga tekanan osmotik tinggi dalam sel (Hu et al. 2000).
Mekanisme ekslusi adalah mekanisme untuk mencegah masuknya Na kedalam jaringan tanaman dan mencegah terjadinya defisit air secara internal dengan cara sintesis solut organik dan meningkatkan sukulensi (Sopandie 2014). Untuk mempertahankan stabilitas lingkungan mikro di dalam sitoplasma, tanaman mengatur konsentrasi relatif dari ion anorganik (terutama K + dan Na +) untuk mempertahankan turgor sel, volume sel, nilai pH intraseluler dan parameter fisiologis penting lainnya. Pada proses fisiologis, sel-sel tumbuhan mengakumulasi K+ dan mengarahkan Na+ keluar sel, yang mengarah kepada peningkatan ratio nilai K+ /Na (Bing et al. 2008). Banyak penelitian menunjukan bahwa dengan adanya kondisi garam yang tinggi kebanyakan tanaman mulai mengakumulasikan beberapa metabolit dengan berat molekul yang rendah dalam sitoplasma seperti prolin, glisin, gula alkohol dan lain-lain untuk menjaga tekanan osmotik tinggi dalam sel, sehingga memungkinkan bagi tanaman untuk menyerap air di bawah kondisi garam yang tinggi ( Hu et al.
2012).
Cekaman garam pada kondisi in vitro akan menghambat pertumbuhan tanaman padi, baik pada kalus maupun tunas hal ini karena konsentrasi garam yang tidak seimbang di dalam media (Htwe et al. 2011). Tanaman yang toleran pada kondisi ini akan melakukan mekanisme tertentu untuk bertahan pada kondisi salin. Mekanisme toleransi pada kalus yaitu dengan meningkatnya akumulasi prolin dalam sel (Htwe et al. 2011). Pada fase bibit akumulasi prolin lebih rendah Adanya akumulasi prolin pada kalus dan plantlet menunjukan adanya potensi genetik untuk mendapatkan tanaman yang toleran salinitas.
Mutasi Induksi
di alam (spontaneous mutation) dan dapat terjadi melalui induksi. Keduanya dapat menimbulkan variasi genetik untuk dijadikan dasar seleksi tanaman (Soeranto, 2003).
Mutasi induksi merupakan salah satu metode untuk meningkatkan keragamman yang dapat diterapkan dalam pemuliaan tanaman. Mutasi induksi dapat dilakukan dengan perlakuan bahan mutagen tertentu. Mutagen yang sering digunakan dalam pemuliaan tanaman yaitu mutagen fisik (radiasi sinar X, sinar gamma) dan mutagen kimia. Penggunaan mutagen dapat mengganggu sistem pembelahan sel, sehingga dapat merubah aktivitas gen, bahkan dapat terbentuk protein baru. Teknik tersebut dapat mempercepat diperolehnya varietas baru dengan berbagai sifat atau karakter yang diinginkan. Sifat-sifat baru yang terbentuk pada mutan akan beragam karena mutasi yang terbentuk bersifat acak.
Mutasi induksi menggunakan radiasi sinar-X dan sinar gamma merupakan metode yang paling banyak digunakan untuk merakit varietas unggul baru, dalam kurun waktu 70 tahun terakhir telah dilepas 2250 varietas mutan diseluruh dunia. (Maluszynki et al. 2000). Iradiasi dapat menyebabkan perubahan genetik dalam sel somatik (mutasi somatik), dapat diturunkan dan dapat menyebabkan terjadinya perubahan fenotipe. Iradiasi sinar gamma sering digunakan dalam usaha pemuliaan tanaman karena dapat meningkatkan variasi – variasi genetik sehingga dapat menghasilkan mutan-mutan baru.
Respon tanaman terhadap efek iradiasi sinar gamma, selain dipengaruhi oleh jenis kultur yang digunakan, juga tergantung dari laju dosis iradiasi yang digunakan (Soeranto 2003). Dosis rendah dapat menyebabkan perubahan abnormal pada fenotipe tanaman dan bersifat dapat balik sedangkan pemberian iradiasi pada dosis yang tinggi dapat mengakibatkan kematian pada jaringan yang diradiasi. Sensitivitas terhadap radiasi dapat diukur berdasarkan nilai LD (lethal dose). LD (lethal dose) adalah dosis yang dapat menyebabkan kematian dari tanaman yang diradiasi. Tingkat sensitivitas tanaman dipengaruhi oleh jenis tanaman, fase tumbuh, ukuran, bahan yang akan dimutasi, serta sangat bervariasi antar jenis tanaman dan antar genotipe (Banerji & Datta 1992). Dari beberapa hasil penelitian menunjukkan bahwa dosis optimum dalam induksi mutasi yang dapat menghasilkan mutan terbanyak diperoleh pada LD50 (Datta 2001). Variabilitas mutan tertinggi terdapat pada mutan hasil iradiasi sinar gamma sekitar LD20 dan LD50 (Soeranto 2012).
Pengunaan iradiasi sinar gamma telah digunakan untuk merakit galur-galur padi yang toleran pada kondisi stress abiotik dan telah banyak dilakukan untuk mendapatkan galur-galur yang toleran salinitas. Perlakuan mutagen fisik dapat merubah karakter fisiologis tanaman sehingga tanaman padi relatif toleran terhadap kondisi salin. (Saleem et al. 2005, Song et al. 2012).
Seleksi In vitro
Keragamman genetik yang ditimbulkan oleh variasi somaklonal dan induksi mutasi bersifat acak. Untuk mengidentifikasi keragamman somaklonal maupun induksi mutasi kearah perubahan yang diinginkan, dapat digunakan metode seleksi in vitro. Melalui metode tersebut, seleksi ketahanan terhadap cekaman abiotik seperti salinitas dapat digabungkan dalam media kultur in vitro
Seleksi in vitro merupakan metode yang lebih efektif dan efisien karena kondisi seleksi dapat dibuat homogen, tempat yang dibutuhkan relatif sedikit, dan efektivitas seleksi tinggi. Intensitas seleksi dapat diperkuat dan dibuat lebih homogen. Penggunaan teknik kultur in vitro akan menghasilkan variasi dari populasi sel somatik melalui seleksi pada media yang sesuai. Populasi sel somatik dapat diseleksi dalam media yang mengandung komponen seleksi sehingga akan meningkatkan frekuensi varian dengan sifat yang diinginkan (Biswan et al. 2002).
Seleksi in vitro untuk toleransi terhadap salinitas menggunakan komponen seleksi NaCl. Basu, (2002) melaporkan bahwa respon pertumbuhan kalus padi menurun tajam dengan meningkatnya konsentrasi NaCl dalam media kultur kalus padi. Pushpam and Rangaswamy (2000) menjelaskan bahwa kalus padi yang dikulturkan pada medium yang mengandung konsentrasi NaCl yang tinggi pertumbuhannya akan terhambat dari pada kalus yang dikulturkan pada media dengan konsentrasi NaCl yang lebih rendah. Penurunan pertumbuhan kalus dalam media yang mengandung NaCl disebabkan pengalihan beberapa kuantum energi untuk pertumbuhan dan metabolisme. Mereka juga melaporkan bahwa kalus padi yang dikulturkan pada media tanpa cekaman dapat tumbuh lebih cepat dibandingkan kalus yang dikulturkan pada media yang diberi cekaman. Basu et al. (2002) melaporkan bahwa retensi K+ pada kalus padi merupakan faktor penting pada mekanisme toleransi garam dimana ion tersebut akan berkolerasi positif dengan pertumbuhan. Sankar et al. (2009) memanfaatkan metode seleksi in vitro ini untuk menyeleksi cekaman garam terhadap 40 varietas hibrida. Hasil penelitian menunjukkan bahwa Hibrida GD 98029/CSSRI 13 menempati urutan pertama diikuti oleh hibrida GD 98028/CO 43, GD 98028/CSR 23, GD 98029/CSR 10, GD 98029/Nona Bokra dan Nona Bokra pada media MS yang mengandung 2,4-D 2 mg/l, kinetin 0.25 mg/l dan kasein hidrolisat 1 g/l dengan komponen seleksi NaCl konsentrasi 1,6 %.
Peningkatan keragamman genetik melalui variasi somaklonal dan induksi mutasi dan perubahannya diarahkan melalui seleksi in vitro merupakan metode yang efektif untuk merakit genotip baru yang memiliki sifat tertentu tanpa merubah terlalu banyak sifat agronomi yang baik dari tanaman induknya. Oleh karena itu, kombinasi antara induksi variasi somaklonal dan seleksi in vitro
3 REGENERASI TUNAS ADVENTIF DARI KALUS BEBERAPA VARIETAS PADI
Adventive Shoots Regeneration of Callus Some Rice Varieties
Abstrak
Regenerasi tunas dari kalus merupakan tahapan yang sangat penting untuk merakit varietas baru melalui jalur non konvensional antara lain kultur in vitro. Penelitian ini bertujuan untuk mendapatkan formulasi media untuk induksi kalus embriogenik, regenerasi dan multiplikasi tunas adventif serta induksi akar padi varietas Ciherang, Inpari 13, Inpara 3, Pokkali dan IR 29. Penelitian ini terdiri dari 4 percobaan yaitu induksi kalus embriogenik, regenerasi kalus membentuk tunas adventif, multiplikasi tunas dan induksi akar. Varietas yang dicobakan ada 5 yaitu Ciherang, Inpari 13, Inpara 3, Pokkali dan IR 29. Perlakuan yang diberikan untuk induksi kalus embriogenik MS + 2,4-D (0, 1, 3,5 dan 7 mg/l) + Casein hidrolisat 3 g/l. Untuk regenerasi tunas adalah perlakuan MS + BA (0, 1 dan 5 mg/l) + Zeatin (0, 01 dan 0,3 mg/l) prolin 100 mg/l. Untuk multiplikasi tunas digunakan MS + Thidiazuron (0, 0,1 dan 0,3 mg/l) dan tahap terakhir untuk perakaran yaitu MS+ IBA (0, 1, 2 dan 3 mg/l). Dari penelitian ini diperoleh hasil bahwa media terbaik untuk induksi kalus untuk padi varietas Ciherang, Inpari 13 dan Pokkali adalah media MS + 2,4-D 3mg/l, sedangkan untuk varietas Inpari 3 dan IR 29 adalah MS + 2,4-D 5mg/l. Untuk regenerasi kalus membentuk tunas media terbaik untuk varietas Ciherang, Inpari 13 dan Pokkali adalah media MS + BA 3 mg/l + Zeatin 0,3 mg/l, sedangkan untuk varietas Inpari 3 dan IR 29 adalah MS + 3 mg/l + Zeatin 0,1 mg/l. Media terbaik untuk multiplikasi tunas adalah MS + Thidiazuron 0,3 mg/l dan untuk induksi perakaran adalah MS + IBA 2 dan 3 mg/l
Kata kunci. kalus embriogenik , tunas adventif, Oryza sativa L, perakaran
Abstract
Shoot regeneration from callus is a very important process to assemble the new varieties through non-conventional. This study aims to gain media formulations for embryogenic callus induction, regeneration and multiplication of shoot and root induction Ciherang, Inpari 13, Inpara 3 Pokkali and IR 29. This study consists of four main activities of embryogenic callus induction, regeneration buds, shoots multiplication and root induction. This study used five variety, they are Ciherang, Inpari 13, Inpara 3 Pokkali and IR 29. The treatment for callus induction is MS + 2,4-D (0, 1, 3,5 and 7 mg/l) + Casein hidrolisat 3 g/l, for shoot regeneration is MS + BA (0, 1 and 5 mg/l) + Zeatin (0, 01 and 0,3 mg/l), for shoot multiplication is MS + Thidiazuron (0, 0,1 and 0,3 mg/l) and for root induction is MS+ IBA (0, 1, 2 and 3 mg/l). Obtained from this study showed that the best medium for callus induction for rice Ciherang, Inpari 13 and Pokkali is media MS + 2,4D 3 mg/l, while for Inpari 3 and IR 29 is MS + 2,4D 5 mg/l. For regeneration of callus forming shoots best medium for Ciherang, Inpari 13 and Pokkali is media MS + BA 3 mg/l + Zeatin 0,3 mg/l, while for Inpari 3 and IR 29 is MS + 3 mg/l + Zeatin 0.1 mg/l. The best media for shoot multiplication was MS + Thidiazuron 0,3 mg/l and for rooting induction was MS + IBA 3 mg/l
Pendahuluan
Perakitan varietas unggul melalui jalur non konvensional seperti rekayasa genetika dan kultur in vitro pada umumnya menggunakan populasi sel somatik yang dikenal dengan nama kalus (populasi sel somatik), karena lebih efektif untuk mendapatkan individu baru dengan perubahan genetic yang lebih sholid. Masalah yang sering dihadapi dalam pemulian non konvensional adalah meregenerasikan kalus membentuk tunas adventif. Apa bila metode regenerasi belum dikusasi dengan baik maka keberhasilan perakitan varietas unggul melalui jalur pemuliaan non konvensional akan sulit dicapai.
Regenerasi tunas adventif dari kalus merupakan proses yang kompleks, banyak faktor yang mempengaruhi, antara lain genotipe, umur eksplan, kondisi fisiologis pohon induk dan formulasi media tumbuh (Stonee et al. 2002), keseimbangan zat pengatur tumbuh khususnya sitokinin terhadap auksin serta struktur kalus (Ehsanpour 2002). Kalus yang baru terbentuk mempunyai respon lebih baik dibandingkan dengan kalus yang telah disub kultur dengan frekuensi tinggi dan telah mengalami periode kultur in vitro yang lama. Faktor lain yang menentukan keberhasilan regenerasi kalus adalah faktor fisik seperti kondisi lingkungan tumbuh antara lain temperatur ruangan, intensitas cahaya, lama penyinaran, kelembaban ruang kultur juga menentukan kemampuan regenerasi kalus.
Regenerasi tanaman dengan menggunakan teknik kultur jaringan dapat dilakukan melalui jalur organogenesis dan embriogenesis somatik (Jimenes 2001). Untuk kedua jalur tersebut memerlukan kalus yang bersifat embriogenik. Kalus yang embriogenik mempunyai ciri-ciri seperti warna yang putih kekuningan, mengkilat, remah dan struktur noduler (mudah dipisahkan membentuk fragment), Berbeda dengan kalus yang bersifat non embriogenik pada umunya berwarna kecoklatan, lembek dan tidak remah (Wani 2011).
Pembentukan tunas dapat dipacu dengan memanipulasi konsentrasi keseimbangan antara auksin dengan sitokinin (Gaba 2005). Beberapa jenis sitokinin yang umum digunakan dalam meregenerasikan kalus pada tanaman padi adalah BAP, Kinetin dan Zeatin. Setiap genotipe atau jaringan mempunyai respons yang berbeda terhadap zat pengatur tumbuh yang diberikan ke dalam media dan memiliki kandungan hormon tumbuh endogen yang berbeda pula (Islam et al. 2004). Banyak hasil penelitian menunjukkan bahwa zat pengatur tumbuh sangat berperan dalam menentukan arah morfogenesis. Pada penelitian Maftuchah et al. (2000) kalus dari padi varietas Indica dapat beregenerasi pada media MS + BA 0,7 mg/l + IAA 0,1 mg/l.
Tujuan Penelitian
Tujuan penelitian adalah mendapatkan formulasi media terbaik untuk induksi kalus embriogenik, regenerasi dan multiplikasi tunas serta induksi akar padi varietas Ciherang, Inpari 13, Inpara 3, Pokkali dan IR 29.
Metode Penelitian
Penelitian ini dilaksanakan di Laboratorium Kultur Jaringan Kelti Biologi Sel dan Jaringan, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian. Bahan tanaman yang digunakan sebagai eksplan adalah embrio zigotik dari benih varietas Ciherang, Inpari 13, Inpara 3, Pokkali dan IR29. Penelitian ini terdiri atas 4 percobaan yaitu induksi kalus embriogenik, regenerasi kalus membentuk tunas adventif, multplikasi tunas dan induksi akar.
Induksi Kalus Embriogenik
Biji disterilisasi di dalam laminar flow menggunakan bahan sterilan seperti alkohol 70% dan klorok, setelah itu dilakukan isolasi embrio zigotik. Embrio ditambahkan sukrosa 3 %, agar 0,8 % dan pH media diatur menjadi + 5,7 dengan penambahan KOH atau HCl0,1 N. Peubah yang diamati adalah persentase eksplan membentuk kalus, persentase kalus embriogenik ( jumlah kalus embrionik/jumlah kalus terbentuk x 100).
Regenerasi Kalus Membentuk Tunas Adventif
Penelitian menggunakan Rancangan Acak Lengkap Faktorial dengan 10 ulangan dimana masing-masing ulangan terdapat 4 kalus. Apabila terdapat pengaruh yang nyata dari perlakuan terhadap peubah yang diamati, kemudian dilakukan uji lanjut Duncan pada taraf kepercayaan 95 %. Perlakuan yang diuji adalah empat taraf konsentrasi BA dengan dan tiga taraf Zeatin. Kalus embriogenik yang telah dihasilkan dipindahkan ke media regenerasi tunas. Media untuk regenerasi tunas adalah media dasar MS+ BA (0, 1, 3, 5 mg/l) + Zeatin (0, 0,1 dan 0,3 mg/l), ke dalam media perlakuan ditambahkan prolin 100 mg/l dan sukrosa 3%. Media dibuat padat dengan menggunakan agar 0,8%. Kultur diinkubasi dalam ruangan kultur pada kondisi terang selama selama 16 jam dalam sehari (dengan intensitas cahaya 1000-1400 lux) serta suhu ruang kultur 25-27oC. Peubah yang diamati yaitu persen struktur nodular yang terbentuk, rerata spot hijau per kalus, rerata tunas per kalus.
Multiplikasi Tunas
0,3% sukrosa dengan satu plantlet per botol. Kultur diinkubasi pada ruang kultur kondisi terang (dengan intensitas cahaya 1000- 1400 lux) selama selama 16 jam dalam sehari serta suhu ruang kultur 25-27oC. Peubah yang diamati yaitu jumlah tunas yang terbentuk. konsentrasi 0, 1, 2, dan 3 mg/l. Peubah yang diamati adalah jumlah dan panjang akar.
Hasil dan Pembahasan
Induksi Kalus Embriogenik
Pada minggu kedua setelah tanam eksplan embrio zigotik mulai memberikan respon dengan terjadinya pembengkakan pada eksplan dan mulai membentuk kalus. Eksplan yang dikulturkan pada media tanpa 2,4-D tidak mampu membentuk kalus, kondisi yang berbeda untuk eksplan yang dikulturkan pada media yang mengandung 2,4-D dapat membentuk kalus (Tabel 1). Beberapa hasil penelitian menunjukkan bahwa 2,4-D merupakan auksin yang efektif untuk menginisiasi terbentuknya kalus dari embrio zigotik pada berbagai varietas padi (Bona et al. 2005, Tariq et al. 2008).
Tabel 1. Pengaruh konsentrai 2,4-D terhadap pembentukan kalus pada beberapa varietas padi, umur 8 minggu setelah kultur
Kalus padi varietas Ciherang, Inpari 13 dan Pokkali yang dihasilkan pada media yang mengandung 2,4-D 3 mg/l bersifat embriogenik dimana kalus tersebut berwarna putih kekuningan, nodular dan remah (mudah dipisah-pisahkan) (Gambar 2). Menurut Wani (2011), kalus yang bersifat embriogenik memiliki ciri-ciri kalus berwana putih kekuningan, serta remah.
Gambar 2. Kalus embriogenik padi varietas (a) Ciherang, (b) Inpari 13 dan (c) Inpara 3
Eksplan padi Inpara 3 yang diberi perlakuan 2,4-D 5 mg/l memberikan respon yang paling baik bila dibandingkan dengan perlakuan lainnya, dimana eksplan yang dapat membentuk kalus adalah 84% dan yang bersifat embriogenik 68%. Peningkatan 2,4-D sampai 7 mg/l dapat menurunkan kemampuan eksplan membentuk kalus dan persentase kalus yang bersifat embriogenik juga menurun. Hal yang sama juga terjadi pada padi varietas IR 29, perlakuan 24-D 5 mg/l menghasilkan eksplan yang membentuk kalus sebesar 86% dengan jumlah kalus bersifat embriogenik sebanyak 68 %. Peningkatan konsentrasi 2,4-D menjadi 7 mg/l dapat menurunkan kemampuan eksplan membentuk kalus.
Regenerasi Kalus Membentuk Tunas Adventif
Pada minggu ke-4 setelah kultur pada media regenerasi, mulai terjadi perubahan pada kalus embriogenik yaitu, mulai terlihat adanya spot-spot hijau walaupun persentasenya masih sangat sedikit (Gambar 3). Pengammatan pada minggu kedelapan terlihat sudah muncul tunas dan pada umumnya sudah membentuk kalus nodular dan spot- spot hijau bakal tunas. Persentase kalus nodular yang membentuk spot hijau terbentuk untuk masing-masing varietas tergantung pada perlakuan kombinasi media. Pada Tabel 2 dapat diketahui bahwa pada umumnya kalus yang dikulturkan pada media tanpa BA dan Zeatin tidak mampu membentuk kalus noduler.
Gambar 3. Regenerasi kalus embriogenik membentuk kalus nodular dan tunas adventif beberapa varietas padi (a) Ciherang, (b) Inpari 13, (c) Inpara 3, (d) Pokkali dan (e) IR 29
a b c
Tabel 2. Regenerasi kalus embriogenik membentuk tunas adventif beberapa varietas padi (Ciherang, Inpari 13, Inpara 3, Pokkali dan IR 29), umur 8 minggu setelah kultur
Perlakuan
Ket: Ket: Angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda pada taraf nyata 5%, uji jarak berganda Duncan
KN : kalus nodular
Pada kalus padi varietas Ciherang, perlakuan BA tunggal hanya mampu menghasilkan kalus noduler kurang dari 50%. Untuk perlakuan kombinasi BA 5 mg/l dan Zeatin 0.1 atau 0,3 mg/l pada media kultur dapat membentuk kalus noduler sebesar 100%. Dari hasil analisis statistik menunjukan perlakuan kombinasi BA dan Zeatin memberikan pengaruh yang nyata terhadap pembentukan spot hijau dan tunas adventif. Perlakuan BA 5 mg/l dan Zeatin 0,1 mg/l memberikan hasil yang terbaik untuk pembentukan spot hijau dimana rerata jumlah tunas adventif yang dihasilkan adalah 4,6. Kombinasi BA dan Zeatin mampu meningkatkan kemampuan kalus padi untuk beregenerasi membentuk tunas adventif. Kondisi yang sama juga terjadi pada kalus tanaman mawar, pemberian Zeatin kombinasi BA pada media tumbuh dapat meningkatkan kemampuan kalus dalam membentuk tunas adventif (Kim et al. 2004).
Untuk parameter jumlah tunas adventif, hasil analisis statistik menunjukkan kombisasi BA dan Zeatin memberikan pengaruh yang nyata terhadap pembentukan tunas adventif. Pelakuan BA 3 mg/l dan Zeatin 0,3 mg/l memberikan respon terbaik dimana rerata jumlah tunasnya 3,3. Pada perlakuan BA 5 mg/l dan Zeatin 0,3 mg/l terjadi penurunan jumlah tunas adventif menjadi 1,4. Hal ini menunjukkan peningkatan konsentrasi zeatin dapat menurunkan jumlah tunas yang terbentuk. Hal yang sama juga terjadi pada kalus tanaman kapas, peningkatan kandungan Zeatin pada media tumbuh akan menurunkan kemampuan kalus dalam membentuk tunas adventif (Lashari et al. 2008).
Pada kalus padi varietas Inpari 13, media yang mengandung BA 1 mg/l atau 3 mg/l yang dikombinasikan dengan Zeatin 0,1 mg/l atau 0,3 mg/l mampu menginduksi kalus embriogenik 100%. Peningkatan BA menjadi 5mg/l menurunkan kemampuan kalus membentuk kalus nodular yaitu 80-90%. Hasil analisis statistik menunjukkan bahwa kombinasi perlakuan BA dan Zeatin memberikan pengaruh yang nyata terhadap pembentukan spot hijau dan tunas adventif. Perlakuan BA 3 mg/l dan Zeatin 0,3 mg/l memberikan pengaruh yang paling baik untuk pembentukan spot hijau dimana rerata jumlahnya yang dihasilkan adalah 5,5, perlakuan ini juga menghasilkan tunas adventif dengan jumlah yang paling banyak yaitu 5,1. Peningkatan kandungan BA menjadi 5 mg/l menurunkan kemampuan kalus membentuk spot hijau dan tunas adventif.
Semua kalus embriogenik padi varietas Inpara 3, yang ditanam pada perlakuan BA 5 mg/l dan Zeatin 0,1 dam 0,3 mg/l dapat menghasikan kalus nodular. Dengan perlakuan konsentrasi BA yang kurang dari 5 mg/l menghasilkan persentase kalus noduler menurun (Tabel 2). Dari hasil analisis statistik memperlihatkan bahwa kombinasi perlakuan BA dengan Zeatin berpengaruh yang nyata terhadap pembentukan rerata jumlah spot hijau dan tunas adventif. Pada
kandungan BA hingga 5 mg/l menurunkan persentase kalus membentuk kalus nodular. Perlakuan terbaik untuk pembentukan spot hijau dan tunas adventif adalah BA 3 mg/l dan Zeatin 0,1 mg/l, dimana rerata jumlah spot hijau yang terbentuk 3.8 dan rerata jumlah tunas adventif 2,1.
Hasil analisis statistik pada kalus padi varietas IR 29 terdapat perbedaan yang nyata dari pemberian perlakuan BA dengan Zeatin terhadap para meter rerata jumlah spot hijau dan rerata jumlah tunas adventif. Pada perlakuan BA 3 mg/l dan Zeatin 0,1 mg/l mampu membentuk kalus nodular sebesar 90% sejalan dengan persentase jumlah spot dan tunas adventif. Perlakuan yang terbaik adalah BA 3 mg/l dan Zeatin 0,1 mg/l dengan rerata jumlah spot hijau sebanyak 3,8 dan 2,1. Pada Tabel 2 menunjukkan pula bahwa tanpa pemberian Zeatin kalus padi varietas IR29 tidak mampu membentuk struktur kalus nodular, spot hijau dan tunas adventif.
Multiplikasi Tunas
Hasil analisis statistik dari rerata jumlah tunas terhadap perlakuan konsentrasi Thidiazuron memberikan pengaruh yang tidak nyata terhadap rerata jumlah tunas yang terbentuk (Tabel 3).
Tabel 3. Rerata jumlah tunas yang terbentuk pada media yang mengandung Thidiazuron dari beberapa varietas padi, umur 2 minggu setelah kultur
Konsentrasi Varietas konsentrasi Thidiazuron cenderung meningkatkan kemampuan tunas untuk bermultiplikasi. Dalam mikropropagasi atau perbanyakan tanaman melalui kultur
Induksi Perakaran
Pada umunya tunas yang berasal dari kalus embriogenik ada yang sudah membentuk meristomoid bakal akar, akan tetapi ada meristem akar yang belum memanjang membentuk akar sempurna. Untuk itu perlu dilakukan sub kultur pada media khusus untuk induksi perakaran. Tunas adventif dari semua varietas padi yang belum berakar disubkultur pada media yang mengandung IBA. Hasil analisis statistik menunjukan perlakuan konsentrasi IBA memberikan pengaruh yang tidak nyata baik terhadap panjang maupun jumlah akar.
Tabel 4. Rerata jumlah dan panjang akar yang terbentuk pada media yang mengandung IBA dari berberapa varietas padi, umur 2 minggu
Konsentrasi
Pada varietas Inpara 3 dan dan IR 29 (Tabel 4), peningkatan konsentrasi IBA hingga 3 mg/l cenderung meningkatkan pembentukan jumlah akar. Inisiasi pembentukan akar dapat dirangsang dengan pemberian auksin seperti IBA, IAA dan NAA. Diantara zat pengatur tumbuh tersebut IBA merupakan jenis auksin yang paling umum digunakan untuk menginduksi akar dibandingkan dengan jenis auksin lainnya karena translokasinya lebih lambat sehingga dapat memacu perakaran pada bagian pangkal tunas. Disamping itu IBA lebih stabil dengan tingkat toksisitas yang lebih rendah bila dibandingkan dengan NAA (George et al. 2007).
Perlakuan IBA 3 mg/l dapat menurunkan kemampuan pemanjangan akar pada tunas in vitro padi varietas Inpara 3 dan dan IR 29. Tanpa adanya auksin IBA dalam media, tunas dapat membentuk akar, kondisi ini menunjukkan semua varietas yang digunakan telah mengandung auksin alami IAA yang memadai bagi pembentukan dan pemanjangan akar.
Simpulan
1. Formulasi media terbaik untuk induksi kalus embriogenik untuk padi varietas Ciherang, Inpari 13 dan Pokkali adalah MS + 2,4-D 3 mg/l + Chasein hidrolisat 3 g/l, sedangkan untuk varietas Inpara 3 dan IR 29 adalah MS + 2,4-D 5 mg/l + Chasein hidrolisat 3 g/l.
2. Formulasi media terbaik untuk regenerasi kalus membentuk tunas adventif untuk padi varietas Ciherang, Inpari 13 dan Pokkali adalah media MS + BA 3 mg/l + Zeatin 0,3 mg/l + prolin 100 mg/l, sedangkan untuk varietas Inpara 3 dan IR 29 adalah MS + BA 3 mg/l + Zeatin 0,1 g/l.
3. Media terbaik untuk multiplikasi tunas pada semua varietas adalah MS + Thidiazuron 0,3 mg/l.
4 PENGARUH IRADIASI SINAR GAMMA TERHADAP PERTUMBUHAN KALUS DAN REGENERASI PADI VARIETAS
CIHERANG, INPARI 13 DAN INPARA 3
The Effect of Gamma Irradiation on the Callus Growth and Regeneration of Rice var. Ciherang, Inpari 13 and Inpara 3
Abstrak
Pengaruh iradiasi sinar gamma terhadap kalus padi tergantung dari dosis iradiasi yang digunakan. Tingkat sensitifitas suatu jaringan tanaman terhadap iradiasi sinar gamma berbeda-beda yang dapat diukur berdasarkan nilai LD (lethal dose). Dosis optimum untuk induksi mutasi yang menghasilkan mutan terbanyak diperoleh pada LD50. Penelitian ini bertujuan untuk menentukan nilai LD50 dan mempelajari pengaruh iradiasi sinar gamma terhadap pertumbuhan kalus padi dan regenerasi plantlet. Bahan tanaman yang digunakan pada penelitian ini adalah embrio dewasa padi varietas Ciherang dan Inpari 13 dan Inpara 3. Penelitian ini menggunakan rancangan lingkungan acak lengkap dengan perlakuan iradiasi sinar gamma pada dosis 10, 20, 30, 40, 50, 60, 70 , 80, 90 dan 100 Gy. Masing-masing perlakuan terdiri atas 10 ulangan dengan 5 kalus embriogenik per botol. Hasil penelitian menunjukan bahwa peningkatan dosis iradiasi sinar gamma mengakibatkan persentase kalus coklat meningkat, pertumbuhan kalus terhambat dan kemampuan kalus untuk beregenerasi menurun. Nilai LD50 untuk kalus varietas Ciherang adalah 22,468 Gy, varietas Inpari 13 adalah 23,124 Gy dan varietas Inpara 3 adalah 21.0305 Gy.
Kata kunci :Dosis irradiasi, letal dosis, kalus padi, sinar gamma, Oriza sativa L.
Abstrak
Effect of gamma irradiation on the callus of rice depends on the dose of irradiation used. The sensitivity of a plant tissue to gamma-ray irradiation is different. It can be measured by the value of LD (lethal dose). The optimum dose for induction of mutations that produce the most mutants obtained in LD50. This study aims to determine the LD50 value and study the effect of gamma irradiation on the growth of rice callus and regeneration of plantlets. The plant material used in this study was mature embryos Ciherang and Inpari 13 and Inpara 3. This study used a randomized complete design environment with gamma ray irradiation treatment at doses of 10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 Gy. Each treatment consisted of 10 replicates with 5 embryogenic callus per bottle. The results showed that increased doses of gamma ray irradiation resulted in increased percentage of brown callus, callus growth stunted and callus ability to regenerate decreases. LD50 value for callus Ciherang was 22.468 Gy, Inpari 13 was 23.124 Gy and varieties Inpara 3 is 21.0305 Gy
Pendahuluan
Mutasi buatan merupakan salah satu metode yang telah terbukti dapat meningkatkan keragamman genetik dan dapat diaplikasikan untuk mendukung program pemuliaan tanaman. Mutasi buatan dapat dilakukan dengan perlakuan bahan mutagen baik fisik maupun kimiawi. Mutagen fisik yaitu radiasi sinar X, sinar gamma dan mutagen kimia yaitu EMS, DEMS sering digunakan dalam pemuliaan tanaman (Kovacs & Keresztes 2002).
Dari kedua mutagen tersebut yang paling banyak digunakan adalah mutagen fisik menggunakan sinar gamma karena frekuensi hasil mutasi lebih tinggi dan lebih mudah diaplikasikan (Van Haten & Tomson 1998). Energi yang berasal dari iradiasi dapat menyebabkan perubahan struktur dan jumlah kromosom, mengganggu sistem pembelahan sel, sehingga dapat merubah aktivitas gen. Hal ini akan menyebabkan terjadinya perubahan genetik dalam sel somatik (mutasi somatik) yang berakibat terjadinya perubahan fenotipe dan sifatnya diwariskan pada turunannya. Metode ini paling banyak digunakan dan dalam kurun waktu 70 tahun terakhir telah dilepas 2250 varietas tanaman mutan diseluruh dunia (Maluszynki et al. 2000). Metode ini dapat dimanfaatkan untuk merakit varietas padi baru yang memiliki karakter adaptasi yang baik terhadap cekaman biotik dan abiotik (Maluszynki et al. 1995).
Untuk menghasilkan tanaman padi mutan, perlakuan iradiasi umumnya diberikan pada biji padi. Radiasi sinar gamma pada biji padi dapat mengakibatkan mutasi pada berbagai segmen kromosom dari sel embrio seperti daerah “scutelum” dan sumbu embrio zigotik. Hal ini memungkinkan terbentuknya individu yang bersifat kimera dimana perubahan genetik yang dihasilkan tidak terjadi pada seluruh sel dari individu tersebut dan tidak dapat diwariskan. Untuk menghasilkan varian baru, perlakuan iradiasi dapat memberikan hasil yang lebih baik apa bila dilakukan pada populasi sel somatik seperti kalus. Iradiasi pada tingkat kalus akan menghasilkan frekuensi varian yang lebih tinggi dibandingkan dengan menggunakan jaringan tanaman, karena pada tingkat kalus sel-selnya masih bersifat meristematik sehingga lebih responsif terhadap radioaktif (Van Harten, 1998). Penggunaan iradiasi sinar gamma pada kalus in vitro umumnya pada dosis rendah (Al-Safadi et al. 2000, La Vina et al. 2001). Menurut Van Harten (1998), pemberian iradiasi sinar gamma 10-30 Gy pada kalus dapat meningkatkan keragamman somaklonal.