RIANA MURTI HANDAYANI
POTENSI CENDAWAN ENDOFIT DALAM UPAYA
PENGENDALIAN PENYAKIT ANTRAKNOSA
(Colletotrichum capsici) PADA TANAMAN CABAI MERAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Potensi Cendawan Endofit dalam Upaya Pengendalian Penyakit Antraknosa (Colletotrichum capsici) pada Tanaman Cabai Merah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2016
Riana Murti Handayani
NIM A352110051
_________________________
RINGKASAN
RIANA MURTI HANDAYANI. Potensi Cendawan Endofit dalam Upaya Pengendalian Penyakit Antraknosa (Colletotrichum capsici) pada Tanaman Cabai Merah. Dibimbing oleh WIDODO dan SURYO WIYONO.
Cabai merah (Capsicum annuum L.) merupakan komoditas pertanian penting. Produktivitas tanaman cabai masih rendah dibandingkan nilai produksi optimumnya, disebabkan oleh adanya serangan penyakit. Antraknosa merupakan salah satu penyakit yang menyerang cabai merah. Penyakit ini disebabkan oleh cendawan Colletotrichum capsici, dan dapat mengakibatkan kerusakan serta kehilangan hasil panen hingga 100%. Usaha pengendalian penyakit antraknosa dengan menggunakan pestisida sintetis belum dapat menanggulangi masalah tersebut. Oleh karena itu, perlu dilakukan alternatif upaya pengendalian antraknosa, diantaranya adalah dengan pengendalian hayati menggunakan cendawan endofit.
Cendawan endofit telah diketahui mampu membantu tanaman inang dalam membangun ketahanan terhadap patogen tanaman. Tujuan penelitian ini adalah untuk mengetahui kemampuan cendawan endofit asal tanaman cabai merah dalam mengendalikan perkembangan penyakit antraknosa. Penelitian ini terdiri atas 5 tahapan, yaitu 1) Eksplorasi cendawan endofit dari tanaman cabai, 2) Uji patogenisitas, 3) Identifikasi, 4) Uji efektivitas penghambatan perkembangan penyakit antraknosa pada buah, 5) Pengujian in vitro dan in vivo untuk mengetahui mekanisme cendawan endofit dalam mengendalikan penyakit antraknosa.
Hasil isolasi diperoleh 237 isolat cendawan endofit, yang terdiri atas 24 isolat dari Cianjur, 65 isolat dari Bogor, dan 148 isolat dari Garut. Isolasi cendawan endofit tersebut menunjukkan bahwa sebagian besar diperoleh dari daun dibandingkan bagian tanaman lainnya. Cendawan endofit hasil isolasi umumnya berpotensi patogen, hanya berkisar antara 4-12% yang berpotensi non patogenik. Hasil uji patogenisitas terdapat 14 isolat cendawan endofit yang berpotensi non patogenik. Cendawan endofit E9D10G2 mampu menunjukkan penghambatan perkembangan antraknosa pada buah dari tanaman yang telah diberi perlakuan endofit.
Cendawan endofit E1Bh5G1 dan E3Cb7B2 memiliki kemampuan antibiosis terhadap C. capsici, masing-masing pada uji kultur ganda dan uji kultur fitrat endofit. Pengujian in vivo diketahui bahwa umumnya cendawan endofit yang diperoleh dalam penelitian ini berpotensi sebagai plant growth promoting fungi
(PGPF), dan menginduksi pembentukan peroksidase tanaman.
Penelitian lebih lanjut diperlukan untuk mengetahui ketepatan waktu dalam aplikasi cendawan endofit pada tanaman cabai merah, dan mengetahui mekanisme lainnya yang mendukung keefektivan cendawan endofit dalam mengendalikan penyakit antraknosa. Hasil penelitian ini diharapkan dapat menjadi alternatif pengembangan teknologi dalam bidang pertanian. Teknologi yang memanfaatkan interaksi tanaman dengan cendawan endofit dalam pengendalian penyakit tanaman, serta dapat menunjang peningkatan produksi dan mutu cabai merah.
SUMMARY
RIANA MURTI HANDAYANI. Potency of Endophytic Fungi as Biocontrol Against of Anthracnose Caused by Colletotrichum capsici in Chili Pepper. Supervised by WIDODO and SURYO WIYONO.
Chili pepper (Capsicum annuum L.) is an important horticultural crop in Indonesia. However, the production yield is below the optimum target due to some diseases. Anthracnose disease caused by Colletotrichumcapsici could lead to lost harvest up to 100%. The use of chemically synthetic pesticides was not sufficient to control anthracnose, thus the use of endophytic fungi as biocontrol against could serve as a good alternative.
Endophytic fungi has been known to help the host plant’s survival against pathogens. Therefore, this research was aimed to study the effectiveness of chili pepper’s endophytic fungi to inhibite the anthracnose. The research was consisted
of five stages; 1) Isolation of endophytic fungi from chili pepper plant, 2) Pathogenicity test, 3) Identification, 4) Effectiveness test of anthracnose
inhibition growth in chili, 5) In vitro and in vivo study to elucidate the mechanism of disease control by endophytic fungi.
A total of 237 isolates of endophytic fungi were obtained from Bogor, Cianjur, and Garut with a composition of 24, 65, and 148, respectively. Most of the isolates used in this study were obtained from the leaves. Most of the isolates are pathogenic endophytic fungi, and only 4-12% of them are non-pathogenic. The pathogenicity test showed that 14 isolates are pathogenic. Among of the non-pathogenic obtained in this study, isolate E9D10G2 potentially suppressed chili pepper anthracnose in in planta test.
Isolate E1Bh5G1 and E3Cb7B2 exhibited antibiotic activity toward
C. capsici in dual culture and in vivo test on detached fruits respectively. Based on the in vivo study, most of the non-pathogenic endophytic fungi in this research are potentially as plant growth promoting fungi (PGPF) and showed the ability to induce plant peroxidase production.
Further research should be conducted to know the best application methods of endophytic fungi to promote chili pepper growth and understand the mechanism of anthracnose inhibition by endophytic fungi. This research might be expected for developing the technology in agricultural fields, especially in plant disease management and optimization of chili pepper quality and production.
Keywords: antibiotic mechanism, antifungal, growth inhibition, interaction ecies, between species a
environmental conditions will affect for biodiversity and living species. It is also applies to insects which environmental factors influence the life cycle of insect. Oil palm is plant of crude palm oil and palm kernel oil, it’s leading commodity non-oil sector. This plant has a production life up to 25-30 years, and the plant is cultivated as plantations.
practices and different habitat conditions of each oil palm’s age will certainly affect for existing ground vegetation. Ground vegetation at palm oil estate is one of the factors that influence the diversity of insects.
The aim of this reseach was to determine the diversity of insect at oil palm plantation. The reseach was conductedbasedage groupsof plant: 1styear, 7th years, and20th years at Rambutan EstatePTPNIII, North Sumatra, Indonesia. Ineach age group15plots(50 m x50m) was specified for insects sampling. Insectswere collectedby pitfaltrap, light trap, insectnet, yellow pan trap, andyellowstickytrap methods.
Total of15 960 insect specimentswerecollected, consisting of12orders, 120familiesand244morphospecies. Diversityof insects that foundin threeage groupsof plantsshoweda highindex. This result suggestthat age of the planthas no effect oninsectdiversity. Insect composition in three age groups of plant are different. At 1stand7th years, the ecological functions of insects dominated by
insects as hebivor, and at20th yearsecological functions of insects dominated by insect as detritivores.
Positive relationship shown by the abundance of ground vegetation around the plant oil palm for insect abundance. The higher the percentage of ground vegetation covering land, the higher abundance of insects can be found
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Fitopatologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
POTENSI CENDAWAN ENDOFIT DALAM UPAYA
PENGENDALIAN PENYAKIT ANTRAKNOSA
(Colletotrichum capsici) PADA TANAMAN CABAI MERAH
Judul Tesis : Potensi Cendawan Endofit dalam Upaya Pengendalian Penyakit Antraknosa (Colletotrichum capsici) pada Tanaman Cabai Merah
Nama : Riana Murti Handayani
NIM : A352110051
Disetujui oleh Komisi Pembimbing
Dr. Ir. Widodo, MS Ketua
Dr. Ir. Suryo Wiyono, MSc.Agr Anggota
Diketahui oleh
Ketua Program Studi Fitopatologi
Prof. Dr. Ir. Sri Hendrastuti Hidayat, MSc
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, MSc.Agr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema penelitian ini ialah biologi kontrol terhadap penyakit antraknosa pada tanaman cabai, dengan judul Potensi Cendawan Endofit dalam Upaya Pengendalian Penyakit Antraknosa (Colletotrichum capsici) pada Tanaman Cabai Merah.
Pada kesempatan ini, penulis menyampaikan penghargaan dan ucapan terima kasih kepada:
1. Dr. Ir. Widodo, MS, dan Dr. Ir. Suryo Wiyono, MSc.Agr, selaku komisi pembimbing atas segala kesabarannya memberikan bimbingan, dan saran selama pelaksanaan penelitian dan penulisan tesis.
2. Direktorat Perguruan Tinggi, Kementerian Pendidikan dan Kebudayaan, Republik Indonesia, yang telah mendanai penelitian ini dalam Program Hibah Penelitian Unggulan Strategis Nasional.
3. Prof. Dr. Ir. Sri Hendrastuti Hidayat, MSc atas bantuannya dan sebagai ketua tim peneliti dalam Program Hibah Penelitian Unggulan Strategis Nasional, Direktorat Perguruan Tinggi, Kementerian Pendidikan dan Kebudayaan, Republik Indonesia.
4. Dr. Ir. Kikin H. Mutaqin, MSi selaku dosen penguji luar komisi yang telah memberikan wawasan baru bagi kami.
5. Bapak Rebo dan keluarga, staf dan keluarga besar CV WiSH Indonesia-Bogor, serta Laboratorium Mikologi Tumbuhan, Departemen Proteksi Tanaman IPB, atas segala bantuan, dan semangatnya.
6. Ayah (Wahyono), Ibu (Rubiyati), dan adik-adik (Rizki, Tatag), serta Mas Gani atas segala doa, dukungan dan kasih sayangnya yang tulus.
7. Bapak Katiman dan Ibu Suparni, Prof. Dr. Ir. Ari Purbayanto, MSc, serta Prof. Dr. Ir. Roedhy Poerwanto, MSc dan Dr. dr. Sri Budiarti beserta keluarga, atas doa, dukungan, dan semangat yang telah diberikan.
8. Prof. Dr. Ir. Meity Suradji Sinaga, MSc, Prof. Dr. Ir. Sandra Arifin Aziz, MS, Dr. Ir. Efi Toding Tondok, Dr. Ir. Hermanu Triwidodo, MSc, serta Dr. Ir. Gayuh Rahayu, atas motivasi, bimbingan, serta semangat yang telah diberikan selama ini.
9. Bapak, Ibu dosen pengajar IPB yang telah memberikan banyak ilmu selama penulis menempuh pendidikan di program S2.
10. Sahabat-sahabatku dan semua pihak yang selalu memberikan semangat dan doa serta bantuannya, sehingga penelitian dan tulisan ini terselesaikan dengan baik.
Semoga tesis ini bermanfaat.
Bogor, Februari 2016
DAFTAR ISI
Penyakit Antraknosa (Colletotrichum spp.) pada Cabai 3 Patogenesis dan Siklus Hidup Colletotrichum spp. 4 Pengendalian Penyakit Antraknosa (Colletotrichum spp.) 4
Cendawan Endofit 5
Cendawan Endofit sebagai Agens Pengendalian Hayati 6
BAHAN DAN METODE 7
Kemampuan Cendawan Endofit Menghambat Perkembangan Gejala
Antraknosa pada Buah secara In planta 14
Kemampuan Cendawan Endofit Menghambat Pertumbuhan Koloni
DAFTAR TABEL
1. Isolat cendawan endofit dari beberapa bagian tanaman cabai merah
sehat 12
2. Patogenisitas cendawan endofit asal berbagai bagian tanaman cabai
merah 13
3. Patogenisitas cendawan endofit asal tanaman cabai merah dari berbagai
daerah 13
4. Diameter gejala antraknosa dan kejadian penyakit pada buah dari
tanaman perlakuan 14
5. Kemampuan penghambatan cendawan endofit terhadap pertumbuhan cendawan C. capsici pada interaksi koloni ganda 15 6. Pengaruh kultur filtrat cendawan endofit dalam pengendalian penyakit
antraknosa 15
DAFTAR GAMBAR
1. Bagan alir penelitian 11
2. Uji patogenisitas cendawan endofit, (A) benih tidak dapat berkecambah, (B) Kontrol, (C) benih berkecambah dan terdapat gejala nekrotik 13 3. Keragaan pertumbuhan tanaman cabai dengan perlakuan cendawan
endofit 16
4. Nilai uji aktivitas enzim (UAE) peroksidase pada perlakuan bibit
tanaman cabai merah 16
DAFTAR LAMPIRAN
1. Lokasi pengambilan sampel tanaman cabai sehat (A) Cianjur, (B) Bogor,
(C) Garut 27
2. Beberapa isolat cendawan endofit yang telah diisolasi 28 3. Hasil isolasi cendawan patogen penyebab antraknosa pada cabai merah,
(A, B) struktur morfologi C. capsici pengamatan dengan mikroskop
compound pada perbesaran 400x, dan (C) koloni cendawan C. capsici
pada media PDA 29
4. Gejala yang muncul pada kecambah cabai merah dalam uji patogenisitas, (A dan B) kecambah nekrotik, (C) benih tidak berkecambah, (D) kecambah tidak berkembang, (E) Kecambah pada kontrol, dan (F) Kecambah pada perlakuan cendawan endofit non patogenik 29 5. Hasil identifikasi cendawan endofit berpotensi non fitopatogenik 30 6. Uji kultur ganda (A) mekanisme antibiosis, (B) mekanisme lisis 33 7. Keragaan pertumbuhan tanaman dengan perlakuan endofit pada 35 hari
setelah pindah tanam 34
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L.) merupakan suatu komoditas pertanian yang penting (KEMENTAN 2015). Produktivitas cabai merah nasional pada tahun 2014 mengalami peningkatan dari tahun 2013 sebesar 8.16 ton per hektar dengan total luas panen 124.110 hektar, menjadi 8.35 ton per hektar dengan total luas panen 128.734 hektar (BPS 2015). Besarnya produktivitas cabai merah berdasarkan data tersebut masih di bawah potensi produksinya yang dapat mencapai 20 ton per hektare (Nurahmi et al. 2011). Produktivitas yang masih rendah tersebut dapat disebabkan oleh beberapa faktor antara lain anomali iklim, hama, serta penyakit pada tanaman cabai. Penyebab penyakit pada tanaman cabai yaitu virus, bakteri, nematoda dan cendawan. Cendawan penyebab penyakit pada tanaman cabai diantaranya yaitu genus Colletotrichum. Penyakit antraknosa disebabkan oleh cendawan Colletotrichum spp., dicirikan dengan adanya bercak coklat kehitaman pada permukaan buah, yang selanjutnya meluas menjadi busuk lunak, pada bagian tengah bercak terdapat kumpulan titik-titik hitam yang terdiri dari sekelompok seta dan konidium cendawan (Agrios 2005).
Usaha pengendalian penyakit antraknosa secara hayati perlu dilakukan sebagai upaya alternatif selain penggunaan pestisida sintetis yang terkadang masih kurang efektif dalam pengendalian organisme pengganggu tanaman. Cendawan endofit merupakan mikroorgnisme yang hidup di dalam jaringan tanaman secara mutualistik, komensalistik, atau parasitik dengan inangnya (Schulz et al. 1999). Beberapa penelitian sebelumnya telah membuktikan bahwa mikroorganisme endofit mampu membantu tanaman inangnya dalam membangun ketahanan terhadap patogen (Rodriguez dan Redman 2008). Cendawan endofit juga memiliki kemampuan menghasilkan metabolit sekunder yang menekan perkembangan beberapa cendawan patogen, sehingga dapat mengendalikan penyakit tanaman (Schulz et al. 2002).
Penelitian sebelumnya telah menunjukkan bahwa beberapa cendawan endofit mampu mengendalikan cendawan penyebab penyakit antraknosa. Istikorini (2008) mengungkapkan bahwa cendawan endofit yang diisolasi dari akar tanaman cabai, yaitu Trichoderma sp. dan Gliocladium sp. mampu menghambat cendawan
Colletotrichum sp. secara langsung. Penggunaan elisitor berupa konidia Fusarium oxysporum CB5 lebih efektif menghambat pertumbuhan koloni C. acutatum. Penelitian Wilia (2010) menyebutkan bahwa penekanan penyakit antraknosa oleh cendawan Acremonium sp., F. oxysforum CT1, dan F. solani CJ1 lebih efektif dibandingkan dengan penekanan penyakit oleh Trichoderma sp. dan Gliocladium
2
Tujuan Penelitian
1. Eksplorasi cendawan endofit asal tanaman cabai.
2. Memperoleh cendawan endofit yang potensial dalam menghambat perkembangan penyakit antraknosa (C. capsici) pada tanaman cabai merah (C. annuum L.).
3. Mengetahui mekanisme cendawan endofit potensial dalam menekan perkembangan penyakit antraknosa (C. capsici) pada tanaman cabai merah (C. annuum L.).
Hipotesis
Cendawan endofit yang berasal dari tanaman cabai merah mampu menghambat perkembangan penyakit antraknosa yang disebabkan oleh
Colletotrichum capsici.
Manfaat Penelitian
3
TINJAUAN PUSTAKA
Penyakit Antraknosa (Colletotrichum spp.) pada Cabai
Colletotrichum sp. merupakan penyebab penyakit antraknosa, selain juga dilaporkan sebagai busuk merah tebu, penyakit buah kopi, busuk mahkota pada stroberi dan pisang, serta bercak coklat kacang tunggak (Waller et al. 2002). Penyakit antraknosa merupakan salah satu penyakit yang banyak terjadi dan merugikan cabai merah. Penyakit antraknosa disebabkan oleh cendawan
Colletotrichum sp., dengan gejala yang diawali oleh bercak coklat kehitaman pada permukaan buah, yang selanjutnya meluas menjadi gejala busuk lunak. Pada gejala ini akan muncul kumpulan titik-titik hitam yang merupakan tubuh buah cendawan tersebut. Gejala yang timbul pada persemaian jika terbawa benih, dapat berupa kegagalan perkecambahan dan menyebabkan kelayuan. Serangan pada tanaman dewasa dapat menyebabkan mati pucuk, serta busuk kering pada daun dan batang. Tingkat serangan antraknosa akan sangat parah ketika musim hujan, dan dapat menyebabkan kehilangan hasil mencapai 50-100% (Hariati 2007; Pakdeevaraporn
et al. 2005).
Antraknosa pada cabai disebabkan oleh beberapa spesies Colletotrichum
spp., antara lain Colletotrichum gloeosporioides, C. acutatum, C. dematium,
C. capsici, C. truncatum dan C. coccodes (Kim et al. 1999; Pakdeevaraporn et al. 2005; Than et al. 2008; Sharma et al. 2014). Cendawan C. gloeosporioides
memiliki dua strain yaitu strain R dan G. Strain R hanya menyerang buah cabai masak yang berwarna merah, sedangkan strain G dapat menyerang semua tanaman, termasuk buah cabai yang masih berwarna hijau maupun buah yang berwarna merah C. gloeosporioides dan C. capsici (Kim et al. 1985). Jenis patogen tersebut dapat bertahan di benih dalam waktu yang cukup lama dengan membentuk acervulus, sehingga merupakan penyakit tular benih. Buah yang terserang C. capsici menjadi busuk dengan warna seperti terbakar sinar matahari yang diikuti busuk basah berwarna hitam. Gejala berwarna hitam karena adanya seta yaitu bagian cendawan yang terbentuk pada aservulus. Cendawan ini pada umumnya menyerang buah cabai menjelang masak ketika buah mulai berwarna kemerahan (Mahasuk et al. 2008).
Colletotrichum capsici,C. gloeosporioides, dan C. acutatum merupakan tiga dari beberapa spesies Colletotrichum yang dilaporkan sebagai penyebab penyakit antraknosa di Asia (Montri et al. 2009; Kanchana-udomkan et al. 2004; Raj et al.
2014). C. capsici merupakan spesies Colletotrichum yang banyak ditemukan pada penelitian tersebut. C. capsici menginfeksi buah cabai, dan dapat bertahan pada benih dengan membentuk aservulus dan mikrosklerotia (Raj dan Chrishtopher 2009). Infeksi C. capsici yang terjadi pada stadia tanaman dewasa lebih tinggi dibandingkan pada pada fase awal pertumbuhan tanaman cabai (Raj et al. 2013).
4
Patogenesis dan Siklus Hidup Colletotrichum spp.
Spesies cendawan Colletotrichum ini banyak yang bersifat tular benih dan bertahan hidup dengan baik sebagai saprob pada sisa tanaman mati yang berada di tanah. Spora seksual dan aseksualnya dapat disebarkan melalui percikan air atau bersamaan uap air yang terkandung dalam angin (Garg et al. 2013; Nicholson dan Moraes 1980). Kondisi permukaan tanaman yang basah secara langsung berpengaruh terhadap perkecambahan spora cendawan, proses infeksi dan pertumbuhan patogen pada tanaman inang. Pada umumnya infeksi terjadi selama cuaca hangat dan basah, pada kisaran suhu 27 oC dengan kelembaban tinggi (80%) yang optimum bagi perkembangan penyakit antraknosa (Than et al. 2008).
Karakteristik hemibiotropik dari beberapa spesies Colletotrichum me- rupakan gabungan fase biotrofik awal yang singkat, di mana sel inang tetap hidup, diikuti dengan perkembangan nekrotrofik yang sangat merusak ditandai dengan meluasnya daerah jaringan mati. Pada proses kolonisasi, keberhasilan kolonisasi terjadi dengan terbentuknya struktur infeksi selama awal dan pasca fase invasi pada proses infeksi. Infeksi terjadi dengan apresorium yang berkembang dari perkecambahan spora pada permukaan jaringan tanaman, yang diikuti penetrasi dipicu oleh perubahan turgor pada kutikula (Deising et al. 2000) serta dalam beberapa kasus infeksi ini juga melalui hifa infektif pada sel epidermis inang (Bailey et al. 1992). Perkecambahan konidia Colletotrichum, terjadi pada saat patogen menginvestasi sel epidermis inang, dengan membentuk tabung kecambah pendek yang dikenal sebagai apresorium segera setelah pengenalan inang. Apresorium dewasa memiliki lapisan melanin dalam dinding sel apressorial dan senyawa osmotik aktif yang disintesis pada konsentrasi tinggi (Mendgen dan Deising 1993; Deising et al. 2000). Infeksi yang muncul dalam jaringan tanaman dibantu oleh produksi efektor penginduksi virulensi terhadap inang (Kleemann et al. 2012; O'Connell et al. 2012). Koloni baru yang muncul dalam beberapa kasus akan memasuki fase biotrofik pada jaringan terinfeksi tanpa menunjukkan gejala eksternal dan dalam jangka waktu yang singkat (1-3 hari) (O’connel et al. 2000). Gejala eksternal yang tidak muncul tersebut dapat lebih lama dengan melibatkan faktor dan proses yang menyebabkan patogen tidak aktif (Prusky dan Plumbley 1992). Cendawan memasuki fase nekrotrofik yang menyebabkan kematian pada sel tumbuhan dan munculnya lesio akibat serangan patogen (Colletotrichum sp.). Jika terdapat infeksi laten, serangan yang tertunda tersebut akan menyebabkan buah tampak sehat dan gejala penyakit akan muncul pada pasca panen sehingga kualitas buah menurun selama masa penyimpanan (Prusky dan Plumbley 1992). Strategi biotrofik pada spesies Colletotrichum yang hadir dengan tanpa gejala juga dapat menjadikan Colletotrichum sp. bersifat endofitik dalam jaringan tanaman hidup (Lu et al. 2004; Joshee et al. 2009; Rojas et al. 2010; Yuan et al. 2011). Fase teleomorf dari cendawan ini adalah Glomerella. Beberapa spesies dari cendawan antraknosa mampu menghasilkan jumlah spora aseksual (konidia) yang sangat banyak dari genus mitosporik atau tidak sempurna(Agrios 2005).
Pengendalian Penyakit Antraknosa (Colletotrichum spp.)
5 kualitas buah akibat pemanenan tersebut. Beberapa penelitian telah dilakukan untuk mengendalikan penyakit antraknosa, diantaranya melalui upaya perakitan varietas tahan (Wusani 2004), bioteknologi dalam rekayasa genetik tanaman (Mahasuk et al. 2008), modifikasi lingkungan, pemanfaatan ekstrak tanaman (Rahman et al. 2011), serta pemanfaatan agens hayati (Istikorini 2008; Wilia 2010).
Pada penelitian Chaisemsaeng et al. (2013) telah diketahui bahwa khamir isolat HS6 memiliki kemampuan penghambatan tertinggi (46.25%) terhadap pertumbuhan C. capsici. Penelitian lain melaporkan khamir antagonis (Candida quercitrusa isolat L2) yang diisolasi dari cabai, menunjukkan penghambatan sebesar 66.40% terhadap C. capsici (Chanchaichaovivat et al. 2007). Aktivitas antifungi dari 24 ekstrak tanaman dan metabolit sekunder dari 5 strain Trichoderma
telah terbukti dapat menekan perkecambahan konidia dan perkembangan tabung kecambah Colletotrichum capsici (Rahman et al. 2011). Cendawan endofit
Trichoderma sp. dan Gliocladium sp. dilaporkan mampu menghambat cendawan
C. acutatum secara langsung, demikian juga dengan elisitor berupa konidia
Fusarium oxysporum CB5 efektif dalam menghambat pertumbuhan koloni
C. acutatum (Istikorini 2008). Patogen penyebab penyakit antraknosa secara in vitro dapat ditekan pertumbuhannya oleh cendawan Acremonium sp., F. oxysforum
CT1, dan F. solani CJ1, serta tidak terjadi mekanisme antibiosis dalam penghambatan pertumbuhan C. acutatum oleh cendawan endofit tersebut (Wilia 2010).
Cendawan Endofit
Cendawan endofit merupakan simbion mutualis, yang hidup dalam jaringan tanaman. Endofit didefinisikan sebagai organisme yang hidup dalam periode waktu tertentu dan dalam siklus hidupnya mengkolonisasi jaringan hidup tanaman inangnya tanpa menimbulkan gejala apapun (Schulz dan Boyle 2005). Keragaman endofit selain ciri morfospesiesnya, dapat diketahui berdasarkan beberapa karakteristiknya. Karakteristik cendawan endofit meliputi interaksi mutualistik dan sifat simbiosis antara inang dengan endofit, kemampuan endofit dalam menginduksi ketahanan pada inang, relatifitas dari infeksi utama dalam hubungannya dengan evolusi terkait asal cendawan endofit, kelimpahan, keragaman, atau komposisi cendawan endofit pada inang, mutualistik vertikal dan horizontal cendawan endofit, regulasi intrinsik cendawan endofit pada tanaman terkait hubungannya dengan kemampuan kompetisi, antagonisme, dan sifat interaksi lainnya (Arnold et al. 2003)
6
(Krauss et al. 2007). Cendawan endofit mampu menghasilkan perubahan fisiologis dan ekologis tanaman, yang berdampak pada peningkatan vigor tanaman (Gundel
et al. 2011; Uchitel et al. 2011). Cendawan endofit membantu tanaman dalam mengatasi stres abiotik dan biotik (Rasmussen et al. 2008; White dan Torres 2010).
Cendawan Endofit sebagai Agens Pengendalian Hayati
Pengendalian hayati didefinisikan sebagai pemanfaatan organisme hidup, hidup selain ketahan genetik tanaman, untuk menekan aktivitas dan populasi satu atau lebih patogen tanaman, serta membantu dalam pembentukan ketahanan inang terhadap suatu penyakit (Cook dan Baker 1993). Beberapa cendawan dalam perannya di alam, hidup di dalam tanaman tanpa menimbulkan kerusakan jaringan atau gejala akibat adanya infeksi dari cendawan tersebut (Hyde dan Soytong 2008). Kohabitasi tanaman dan cendawan tersebut berkembang menjadi asosiasi mutualistik yang spesifik, asosiasi tersebut dapat menghasilkan keunggulan kompetitif. Penelitian Saikkonen et al. (2010) menunjukkan bahwa beberapa spesies Neotyphodium dapat membantu tanaman dalam membangun ketahanan terhadap cendawan lainnnya dan serangga, serta tanaman toleran terhadap kekeringan. Interaksi ini secara signifikan dapat mempengaruhi kesehatan tanaman dengan berbagai mekanisme. Mekanisme pengendalian dapat dipahami melalui cara interaksi organisme tersebut. Organisme yang bersifat biotrof dan hidup dalam tanaman, secara langsung memanfaatkan fotosintat tanaman inang. Interaksi tersebut belum dapat diungkapkan dengan pasti, akan tetapi pada interaksi tertentu cendawan endofit berperan dalam efisiensi fotosintesis (Yan et al. 2013).
Beberapa cendawan endofit mampu mengurangi infeksi patogen dalam pengendalian penyakit. Dingle dan Mc Gee (2003) mengemukakan bahwa filtrat kultur Chaetomium dan Phoma sp. dapat mengaktivasi reaksi pertahanan aktif dari tanaman, sehingga mampu membatasi penyebaran dan replikasi Puccinia recondita
f.sp tritici. Cendawan endofit dapat menghasilkan senyawa metabolit yang berperan melindungi inang. Li et al. (2004) menunjukkan bahwa kultur cendawan endofit
Neotyphodium sp. yang diisolasi dari rumput Poa ampla Merr. dapat membantu mengekskresikan enzim kitinase, yang berperan sebagai enzim hidrolitik dalam menyediakan nutrisi, pertumbuhan atau pertahanan bagi endofit. Kapang endofit
Monotospora sp. yang berasal dari tanaman Cynodon dactylon mampu meng-hasilkan enzim lakase (Wang et al. 2006). Metabolit sekunder yang dihasilkan oleh kapang endofit pun bervariasi dan memiliki aktivitas biologis yang bervariasi pula, sehingga pemanfaatan cendawan endofit atau pun senyawa metabolit yang dihasil-kan banyak dimanfaatdihasil-kan di bidang pertanian, industri, maupun farmasi (Tan dan Zou 2001; Zhang et al. 2006).
7
multiflorum, yaitu endofit mutualistik (Neotyphodium occultans) dan patogenik (Claviceps purpurea). Neotyphodium merupakan cendawan endofit yang membentuk simbiosis sistemik dan tanpa gejala dengan rumput pada musim dingin. Infeksi alami berpengaruh dalam menghasilkan interaksi simbiotik atau pun
non-simbiotik dengan tanaman pada berbagai kondisi stres terhadap herbisida
N. occultans secara signifikan mengurangi 70% dari infeksi yang ditimbulkan oleh
C. purpurea (Pérez et al. 2013). Penelitian Pérez et al. (2013) menunjukkan bahwa cendawan endofitik dapat menjadi simbion konstitutif yang bersifat sistemik dan dapat memediasi interaksi antara inang dengan patogen, pada tingkat stres tanaman terhadap lingkungan.
BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilakukan di Laboratorium Mikologi Tumbuhan, Departemen Proteksi Tanaman, Institut Pertanian Bogor (PTN-IPB), Lahan percobaan di CV WiSH Indonesia, serta pertanaman penduduk di Desa Loa, Kecamatan Tamansari, Bogor. Penelitian dilaksanakan pada bulan Mei 2013 sampai dengan Desember 2015.
Bahan
Bahan yang digunakan dalam penelitian ini yaitu tanaman dengan buah cabai merah yang tidak menunjukkan gejala antraknosa, yang berasal dari daerah Cianjur, Garut, dan Bogor. Cendawan patogen (C. capsici) yang diperoleh dari hasil isolasi asal buah cabai yang bergejala antraknosa.
Metode
Isolasi Cendawan
Isolasi cendawan patogen. Cendawan patogen yang digunakan dalam penelitian ini berasal dari buah cabai merah yang menunjukkan adanya gejala antraknosa. Cabai merah bergejala antraknosa disterilisasi permukaan dengan dicelup dalam larutan NaOCl 1% selama 30 detik, dan dibilas dengan air destilata steril, kemudian dikeringkan di atas kertas tisu steril. Bagian buah dipotong dengan ukuran 1x1 cm, dan ditanam pada media Potato Dextrose Agar (PDA), kemudian diinkubasikan pada suhu ruang (25-30 oC). Pengamatan dilakukan setiap hari terhadap pertumbuhan miselium pada potongan jaringan tanaman. Cendawan yang tumbuh pada potongan jaringan, kemudian disubkultur sehingga diperoleh koloni murni. Koloni cendawan yang telah murni diamati di bawah mikroskop dan diidentifikasi.
8
dalam etanol 70% (1 menit), kemudian dengan NaOCl 1% (daun dan buah) dan 3% (akar, batang, dan cabang) selama 3 menit dan etanol 70% selama 30 detik. Setelah disterilisasi kemudian dibilas dengan air destilata steril dan dikeringkan di atas kertas tisu steril. Setelah dipotong dengan ukuran 0.5-1 cm, bagian tanaman tersebut ditanam pada media Malt Extract Agar (MEA) 10%, dan diinkubasikan pada suhu ruang (25-30 oC). Pengamatan dilakukan setiap hari terhadap pertumbuhan miselium pada potongan jaringan tanaman. Uji awal kesterilan jaringan tanaman dilakukan dengan cara membuat goresan air destilata steril pada bilasan terakhir di permukaan media MEA 10% dan selanjutnya diinkubasi selama 3-5 hari. Uji kesterilan dilakukan untuk meyakinkan bahwa cendawan yang muncul berasal dari dalam jaringan tanaman. Hasil isolasi cendawan endofit tidak dapat digunakan jika pada media uji kesterilan masih ditemukan kontaminan (modifikasi Rodriguez dan Redman 2008).
Uji Patogenisitas
Seleksi cendawan non patogenik dilakukan dengan metode penanaman biji cabai di atas permukaan koloni isolat cendawan endofit (7 hari setelah masa inkubasi koloni cendawan) pada media Potato Dextrose Agar (PDA). Biji yang akan ditanam, direndam dalam air destilata steril selama 15 menit, kemudian disterilisasi permukaan dengan larutan NaOCl 1% selama 2 menit, dan dibilas sebanyak tiga kali dengan air destilata steril, kemudian dikeringanginkan di atas kertas tisu steril. Pengamatan patogenisitas cendawan endofit dilakukan pada 14 hari setelah penanaman benih pada media PDA dengan koloni cendawan endofit. Patogenisitas cendawan endofit terhadap benih cabai dan perkecambahan cabai diketahui berdasarkan penghambatan cendawan endofit terhadap perkecambahan benih cabai, daya kecambah benih cabai, serta gejala nekrotik yang ditimbulkan pada kecambah cabai. Cendawan endofit non patogenik tidak akan menimbulkan gejala penyakit atau kerusakan jaringan pada tanaman tersebut (Istikorini 2008). Identifikasi
Isolat-isolat cendawan endofit yang diperoleh tersebut diidentifikasi berdasarkan karakter morfologi secara mikroskopik yang meliputi bentuk konidia, bentuk hifa, dan beberapa karakter lainnya yang dicocokkan dengan pustaka acuan identifikasi menurut Barnett dan Hunter (1998), dan Watanabe (2002).
Penyiapan Pembibitan Cabai, Inokulasi Cendawan Endofit dan Uji Ketahanan Buah terhadap C. capsici secara In planta
ULTRA-9 TURRAX T18 Basic) dengan kecepatan 3500-24000 rpm selama 5 menit atau hingga didapatkan suspensi yang relatif homogen. Kerapatan propagul cendawan endofit yang digunakan adalah 106cfu ml-1.
Inokulasi cendawan endofit dilakukan 2 kali, yaitu inokulasi dengan perendaman benih selama 12 jam sebelum ditanam, dan penyiraman suspensi cendawan endofit pada bibit cabai berumur 3 minggu setelah semai. Perlakuan dengan air destilata steril untuk perendaman dan penyiraman digunakan sebagai perlakuan kontrol. Benih cabai yang telah diberi perlakuan ditanam pada baki semai dengan media semai terdiri atas sekam bakar, pupuk kandang, dan media tanam komersil dengan perbandingan 1:1:1 (v/v). Tanaman persemaian yang telah berumur 4 minggu setelah semai dipindah tanam ke lahan pertanaman. Perawatan tanaman dilakukan dengan penyiraman dan pemupukan tanaman secara berkala, sampai tanaman berbuah.
Buah cabai yang dihasilkan oleh tanaman dengan perlakuan endofit dipanen. Buah dicuci bersih dan disterilisasi permukaan menggunakan NaOCl 1%, serta dibilas dengan air destilata steril sebanyak tiga kali, kemudian dikeringanginkan di atas kertas buram steril. Buah yang telah disterilisasi permukaan tersebut dilukai dengan jarum, kemudian ditetesi suspensi konidia C. capsici (kerapatan 106
cfu ml-1) sebanyak 20 l pada setiap titik pelukaan. Pengamatan dilakukan terhadap jumlah titik inokulasi yang bergejala antraknosa dan diameter gejala yang timbul pada titik pelukaan buah tersebut.
Kejadian penyakit diamati 7 hari setelah inokulasi. Pengamatan dilakukan terhadap 3 buah cabai setiap perlakuan yang diulang 5 kali. Kejadian penyakit antraknosa dihitung dengan menggunakan rumus berikut:
KP =N ×n %
Keteranganμ
KP = Kejadian penyakit
n = Jumlah titik luka yang bergejala
N = Jumlah titik luka pada buah yang diamati
Uji Daya Penghambatan Cendaw an Endofit terhadap Pertumbuhan Cendawan C. capsici secara In vitro
Uji daya hambat dengan metode kultur ganda.Medium yang digunakan untuk uji ini adalah media PDA. Masing-masing cendawan non patogenik (diameter 0.5 cm) diletakkan pada media PDA dalam cawan Petri (diameter 9 cm) dengan jarak 2.2 cm dari pinggir cawan, kemudian dipasangkan dengan potongan media yang ditumbuhi C. capsici (diameter 0.5 cm) pada jarak ± 4 cm dari cendawan endofit. Masing-masing perlakuan terdiri atas 5 ulangan. Pengamatan dilakukan pada 7 hari masa inkubasi, dengan mengukur diameter koloni cendawan patogen. Pengamatan dilakukan terhadap diameter koloni pertumbuhan cendawan C. capsici
dan lebar zona penghambatan antara kedua ujung koloni cendawan. Persentase penghambatan cendawan patogen (Colletotrichum sp.) dihitung dengan persamaan sebagai berikut:
PR =(dkd− dp)
10
Keterangan:
PR = Persentase penghambatan relatif
dk = Diameter koloni patogen yang tumbuh sebagai kontrol
dp = Diameter koloni patogen yang tumbuh pada uji kultur ganda Uji kemampuan filtrat kultur cendawan endofit terhadap penyakit antraknosa pada buah cabai. Inokulum cendawan endofit yang telah diinkubasi selama 7 hari pada medium PDA, dibiakkan ke dalam media PDB dan digoyang dengan penggojok (shaker) pada kecepatan 100 rpm selama 14hari. Kultur filtrat yang dihasilkan oleh cendawan endofit dalam media PDB (14 hari) kemudian disaring menggunakan saringan milipore 0.22 m, kemudian disentrifugasi dengan kecepatan 6000 rpm selama 1 menit dan diambil supernatannya. Supernatan tersebut diencerkan dengan air destilata steril sampai konsentrasi larutan 5% (v/v). Suspensi patogen diperoleh dengan cara menuangkan air destilata (3-5 ml) ke dalam koloni C. capsici yang telah diinkubasi selama 7 hari pada medium PDA. Suspensi disaring dengan kain kasa(3 lapis) untuk memisahkan hifa dan miselia, kemudian disentrifugasi 1500 rpm selama 1 menit, dan dibuang cairannya.
Buah cabai dari pertanaman dicuci bersih dan disterilisasi permukaannya menggunakan NaOCl 1% dengan direndam selama 2 menit, serta dibilas dengan air destilata steril sebanyak tiga kali. Buah yang telah disterilisasi permukaannya direndam (selama 5 menit) dalam kultur filtrat 5% (v/v), dan dikeringanginkan. Buah yang telah direndam tersebut, kemudian dilukai dengan jarum, dan diteteskan suspensi konidia C. capsici (kerapatan 106 cfu ml-1) sebanyak 20 l pada tiap titik luka. Pengamatan dilakukan terhadap jumlah titik bergejala dan diameter dari gejala yang timbul di titik pelukaan pada buah tersebut, pada 7 hari setelah inokulasi. Karakterisasi Mekanisme Agens Hayati terhadap Pertumbuhan Cendawan C. capsici secara In vivo
Evaluasi adanya induksi resistensi pada tanaman cabai. Evaluasi adanya induksi resistensi pada tanaman cabai dilakukan melalui analisis fitokimia enzim peroksidase (modifikasi Hammerschmidt et al. 1982). Enzim peroksidase diekstraksi dari bagian batang tanaman yang sudah diberi perlakuan digerus dalam buffer fosfat 0.01 M, pH 6.0 dengan perbandingan 1:4 (g ml-1) menggunakan mortar. Hasil gerusan disaring dengan kain, kemudian disentrifugasi dengan kecepatan 5 000 rpm selama 30 menit pada suhu 4 oC. Supernatan diencerkan dengan larutan bufer fosfat 0.01 M, pH 6.0 (1:3) dan dihomogenkan (sebagai sumber enzim).
Analisis kuantitatif enzim peroksidase dilakukan dengan metode spektrofotometri. Blanko yang digunakan dalam analisis ini adalah larutan pereaksi tanpa sumber enzim. Larutan pereaksi terdiri atas 5 ml larutan pirogallol 0.5 M dan 0.5 ml H2O2 1%. Larutan pirogallol 0.5 M terbuat dari 10 ml pirogalol 0.5 M, 12.5
ml bufer fosfat 0.066 M, pH 6.0 dan air destilata hingga volume 100 ml. Larutan pereaksi ditambahkan pada 0.2 ml larutan sumber enzim. Campuran tersebut selanjutnya dihomogenkan selama 5 sampai 10 detik dan diamati pada panjang gelombang ( ) 420 nm. Nilai absorban diamati setiap 30 detik selama 150 detik.
11
Optical Density) AOD = b) dari suatu pengamatan dihitung melalui persamaan regresi (Y=a+bx).
Unit aktivitas enzim (UAE) dihitung dengan persamaan :
UAE = AOD x sediaan enzim (ml) / bobot basah kontrol (g)
Kemampuan cendawan endofit dalam memacu pertumbuhan tanaman cabai. Bibit tanaman cabai yang telah diinokulasi cendawan endofit, dan berumur 35 hari setelah tanam, diambil sebanyak 5 tanaman. Sampel tanaman tersebut diukur tinggi tajuk, dan panjang akar, serta jumlah daun.
Gambar 1 Bagan alir penelitian Eksplorasi Cendawan Endofit
Isolasi
Uji Patogenisitas Koleksi
Tidak diuji
Plant Growth Promoting
Fungi
Isolat cendawan Potensial (Agens Biokontrol Antraknosa)
Uji Efikasi
Uji Kultur Filtrat
Uji Buah (Pascapanen) Evaluasi
Mekanisme
Uji Peroksidase Uji Kultur Ganda
(Uji Antagonis) Patogenik
12
HASIL
Isolasi Cendawan
Sampel buah cabai merah sakit bergejala antraknosa diperoleh dari pertanaman di daerah Gunung Bunder (Bogor) dan berhasil teridentifikasi sebagai
Colletotrichum capsici. Colletotrichum capsici (Syd.) Butler dan Busby memiliki konidia agak meruncing seperti bulan sabit (falcate) berukuran panjang 16-30 µm dan banyak seta (Sutton 1992).
Pada penelitian ini telah dilakukan pengambilan sampel dan isolasi cendawan. Sampel tanaman cabai diperoleh dari daerah Cianjur (Mangunkerta, Varietas Tanjung), Bogor 1 (Gunung Bunder, Varietas TW), Bogor 2 (Situ Gede, Varietas TM 99), Garut 1 (Desa Panjiwangi, Varietas TM 99) dan Garut 2 (Desa Rancabango, Varietas TM 99), masing-masing lokasi 10 tanaman cabai merah, dengan umur tanaman antara 3-3.5 bulan. Lokasi pengambilan sampel umumnya merupakan dataran tinggi (604-800 mdpl).
Cendawan endofit yang berhasil diisolasi dari wilayah Cianjur, Garut, dan Bogor berjumlah 237 isolat, yang terdiri atas 24 isolat dari Cianjur, 65 isolat dari Bogor, dan 148 isolat dari Garut (Tabel 1). Hasil isolasi cendawan endofit dalam penelitian ini menunjukkan bahwa sebagian besar cendawan endofit pada tanaman cabai ditemukan dari daun dibandingkan bagian tanaman lainnya (Tabel 1).
Tabel 1 Isolat cendawan endofit dari beberapa bagian tanaman cabai merah sehat
Asal
Isolat Akar BatangJumlah isolat dari bagian tanamanCabang Daun Buah Total Cianjur 7 (2λ.17)1 6 (25.00) 1 (4.17) 6 (25.00) 4 (16.70) 24 (λ.λ5)
Bogor 13 (20.00) 13 (20.00) 7 (10.77) 28 (43.08) 4 (6.15) 65 (26.70) Garut 23 (15.54) 10 (6.67) 4 (2.70) 83 (56.08) 28 (18.λ0) 1 48 (63.35)
Total 43 (18.10) 2λ (12.24) 12 (5.06) 117 (4λ.37) 36 (15.20) 237 (100.00)
Keterangan : 1 Angka di dalam kurung menunjukkan persentase.
Uji Patogenisitas
13 terdapat beberapa cendawan endofit yang berpotensi non patogenik, masing-masing ditemukan 8% dan 12% (Tabel 3).
Tabel 2 Patogenisitas cendawan endofit asal berbagai bagian tanaman cabai merah
Patogenisitas Akar BatangJumlah isolat dari bagian tanaman Cabang Daun Buah Total Potensi patogen 41 (λ5.35)1 28 (λ3.33) 17 (λ4.44) 55 (88.8λ) 1λ (λ0,48) 161 (λ2.00)
Non patogenik 2 (4.65) 2 (6.67) 1 (5.56) 7 (11.11) 2 (λ.52) 14 (8.00)
Total 43(100.00) 30 (100.00) 18(100.00) 63 (100.00) 21 (100.00) 175(100.00)
Keterangan : 1 Angka di dalam kurung menunjukkan persentase
Tabel 3 Patogenisitas cendawan endofit asal tanaman cabai merah dari berbagai daerah
Patogenisitas Cianjur Asal isolatBogor Garut
Potensi patogen 22 (100.00)1 38 (88.37) 101 (λ1.82)
Non patogenik 0 (0.00) 5 (11.63) λ (8.18)
Keterangan : 1 Angka di dalam kurung menunjukkan persentase
Hasil uji patogenisitas cendawan endofit terhadap benih menunjukkan beberapa respon yang beragam. Respon tersebut diantaranya yaitu benih tidak dapat berkecambah, benih berkecambah dan terdapat gejala berupa nekrosis pada kecambah, serta benih berkecambah tanpa adanya gejala nekrosis (Gambar 2). Perkecambahan benih dengan perlakuan cendawan endofit pada umumnya menunjukkan daya kecambah yang lebih kecil dari kontrol.
Gambar 2 Uji patogenisitas cendawan endofit, (A) benih tidak dapat berkecambah, (B) Kontrol, (C) benih berkecambah dan terdapat gejala nekrotik
14
Identifikasi Cendawan Endofit
Cendawan endofit non patogenik merupakan yang berhasil ditemukan ber-potensi sebagai kandidat agens pengendalian hayati. Hasil identifikasi menunjuk-kan bahwa dari 14 cendawan endofit non patogenik 2 diantaranya tidak dapat digunakan lebih lanjut, yaitu isolat E6D9B2 dan isolat E8B3G2. Cendawan endofit E6D9B2 dan E8B3G2 diidentifikasi sebagai Paecilomyces sp.
Uji lanjut dalam mengetahui potensi cendawan endofit dari tanaman cabai merah dilakukan terhadap 12 isolat cendawan endofit yang teridentifikasi yaitu sebagai cendawan dengan miselia steril (E1Bh5G1, E2A3B2, E5BH4G1, E7B8B2, E11D9B2, E13A7G1, E14D2G2), Curvularia sp. (E3Cb7B2), Verticillium sp. (E9D10G2, E12D6G2), Colletotrichum gloeosporioides (E4D5G1), Fusarium solani non patogenik (E10D6G1) (Lampiran 5).
Kemampuan Cendawan Endofit Menghambat Perkembangan Gejala Antraknosa pada Buah secara In planta
Tingkat insidensi penyakit dan diameter gejala antraknosa pada buah yang berasal dari tanaman perlakuan tidak berbeda nyata dengan kontrol. Namun dari hasil uji pada buah menunjukkan adanya potensi penghambatan oleh isolat E9D10G2 (Tabel 4).
Tabel 4 Diameter gejala antraknosa dan insidensi penyakit pada buah dari tanaman perlakuan
Perlakuan Diameter gejala antraknosa (cm) Insidensi penyakit (%)
Kontrol 0.64 ± 0.28 44.44
Kemampuan Cendawan Endofit Menghambat Pertumbuhan Koloni Cendawan C. capsici, dan Perkembangan Gejala Antraknosa secara In vitro
pada Buah
15 dalam menghambat pertumbuhan C. capsici. Persentase nilai penghambatan ditunjukkan dalam interaksi tersebut (Tabel 5). Mekanisme antibiosis pada interaksi in vitro tersebut terjadi pada interaksi antara isolat E1Bh5G1 dengan C. capsici
(Tabel 6).
Tabel 5 Kemampuan penghambatan cendawan endofit terhadap pertumbuhan cendawan C. capsici pada interaksi koloni ganda
Perlakuan Penghambatan Relatif (%) Antibiosis/Lisis
Kontrol 0 -
Tabel 6 Pengaruh kultur filtrat cendawan endofit dalam pengendalian penyakit antraknosa
Perlakuan Diameter gejala antraknosa (cm) Insidensi penyakit (%)
Kontrol 0.37±0.11 92.59
Kemampuan Cendaw an dalam Memacu Pertumbuhan Tanaman Cabai
16
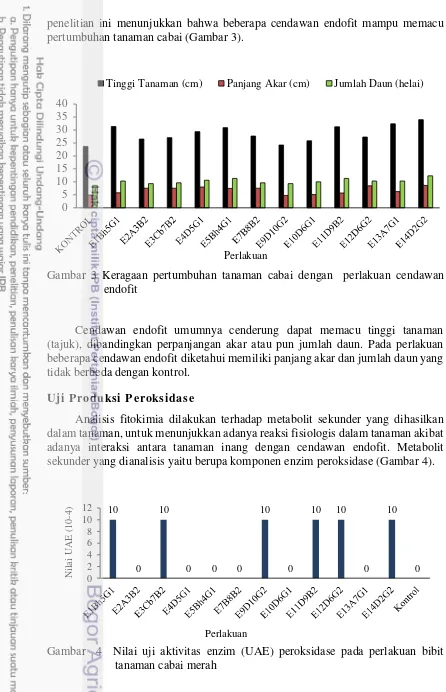
penelitian ini menunjukkan bahwa beberapa cendawan endofit mampu memacu pertumbuhan tanaman cabai (Gambar 3).
Gambar 3 Keragaan pertumbuhan tanaman cabai dengan perlakuan cendawan endofit
Cendawan endofit umumnya cenderung dapat memacu tinggi tanaman (tajuk), dibandingkan perpanjangan akar atau pun jumlah daun. Pada perlakuan beberapa cendawan endofit diketahui memiliki panjang akar dan jumlah daun yang tidak berbeda dengan kontrol.
Uji Produksi Peroksidase
Analisis fitokimia dilakukan terhadap metabolit sekunder yang dihasilkan dalam tanaman, untuk menunjukkan adanya reaksi fisiologis dalam tanaman akibat adanya interaksi antara tanaman inang dengan cendawan endofit. Metabolit sekunder yang dianalisis yaitu berupa komponen enzim peroksidase (Gambar 4).
Gambar 4 Nilai uji aktivitas enzim (UAE) peroksidase pada perlakuan bibit
Tinggi Tanaman (cm) Panjang Akar (cm) Jumlah Daun (helai)
17
PEMBAHASAN
Hasil eksplorasi cendawan endofit dalam penelitian ini menunjukkan bahwa jumlah terbanyak cendawan endofit yang diperoleh berasal dari daun dibandingkan bagian tanaman lainnya (Tabel 1). Mikroorganisme filoplan, cendawan, khamir dan bakteri telah dilaporkan dapat dalam mengendalikan Colletotrichum gloeosporioides secara alami (Stirling et al. 2000). Inacio et al. (2010) menyatakan bahwa daun memiliki topografi dan sifat fisik yang sering berubah, serta mampu menyediakan habitat yang beragam bagi bermacam-macam komunitas mikroorganisme. Peneliti lain melaporkan bahwa kolonisasi cendawan endofit dan distribusinya bergantung pada jenis tanaman dan musim pada saat pengambilan sampel (Kim et al. 2013). Metode isolasi, letak geografis asal tanaman, serta bagian yang diisolasi juga turut mempengaruhi keragaman spesies dari cendawan endofit (Hyde dan Soytong 2008).
Uji patogenisitas pada 175 cendawan endofit terhadap benih cabai secara in vitro menunjukkan beberapa respon yang beragam. Respon tersebut yaitu, benih tidak dapat berkecambah, benih berkecambah dan menunjukkan gejala nekrosis pada kecambah, serta benih berkecambah tanpa adanya gejala nekrosis. Perkecambahan benih dengan perlakuan cendawan endofit pada umumnya menunjukkan daya kecambah yang lebih kecil dibandingkan kontrol. Berdasarkan uji patogenisitas, sebagian besar cendawan endofit yang diperoleh dalam penelitian ini masih berpotensi sebagai patogen (Tabel 2 dan Tabel 3). Cendawan endofit tersebut beberapa diantaranya berasal dari cendawan patogen yang berkembang menjadi patogen avirulen, kemudian berevolusi menjadi simbion mutualistik. Schulz dan Boyle (2005) melaporkan bahwa cendawan endofit dalam interaksinya dengan tanaman berperan menyediakan nutrisi dan sebagai pelindung dari lingkungan eksternal yang tidak menguntungkan dan kompetisi dengan mikro-organisme lainnya.
Cendawan endofit non patogenik merupakan kandidat agens hayati yang berpotensi melindungi tanaman dari gangguan biotik maupun abiotik. Cendawan
Paecilomyces yang diperoleh dalam penelitian meskipun memiliki potensi sebagai agens pengendalian hayati (Kiewnick dan Sikora 2006), tetapi juga diketahui berpotensi sebagai patogen terhadap mamalia yang dapat menyebabkan penisilikosis dan oculomycosis (Hospenthal dan Rinaldi 2008).
18
Arnold et al. (2003) menyatakan bahwa keragaman endofit dapat diketahui berdasarkan ciri morfospesies, karakteristik interaksi mutualistik vertikal dan horizontal cendawan endofit, kemampuan dalam menginduksi ketahanan inang, relatifitas dari infeksi utama, kelimpahan, keragaman, atau komposisi cendawan endofit pada inang, serta regulasi intrinsik cendawan endofit pada tanaman terkait kemampuan kompetisi, antagonis, dan sifat interaksi lainnya.
Tanaman inang dalam fase simbiotiknya dengan cendawan endofit, tanaman berfungsi sebagai tempat tumbuh bagi cendawan endofit dari pengaruh lingkungan. Serangan parasit atau hama akan menyebabkan tanaman inang membangun pertahanan, dan menyebabkan tanaman stress, sehingga cendawan endofit membantu tanaman inang membangun ketahanan dengan menghasilkan senyawa bioaktif. Senyawa bioaktif cendawan tersebut dapat merupakan hasil dari metabolisme cendawan itu sendiri, atau adanya prekusor dari tanaman inang yang dimanfaatkan oleh cendawan endofit dalam memproduksi senyawa bioaktif tersebut (Ginting et al. 2013). Isolat E3Cb7B2 dari hasil penelitian ini diidentifikasi sebagai cendawan Curvularia sp. Penelitian Siddharha et al. (2009) dengan menggunakan isolat Curvularia sp. menunjukkan bahwa cendawan tersebut menghasilkan senyawa 11-α-methoxycurvularin dan (S)-5-etil-8, 8-dimethyl-nonanal. Uji bioassay peneliti tersebut melaporkan bahwa 11-α-methoxycurvularin dapat menghambat bakteri, jamur dan larva instar 4 Spodoptera litura (Siddharha
et al. 2009).
Perbedaan kemampuan penghambatan setiap isolat pada uji buah tersebut dapat disebabkan adanya perbedaan interaksi yang muncul antara C. capsici pada buah hasil dari tanaman yang telah diperlakukan dengan suspensi cendawan endofit. Perbedaan respon yang ditimbulkan oleh cendawan endofit dapat terkait dengan karakteristik tiap-tiap isolat yang berkontribusi dalam menentukan status simbiosis dengan tanaman. Cendawan endofit umumnya pada kondisi alami dalam tanaman inang mampu mengkolonisasi inang dengan baik dan secara in vitro dapat tumbuh cepat serta memiliki kemampuan antibiosis yang relatif langka secara alaminya. Beberapa cendawan endofit secara in vitro juga cenderung tumbuh lambat, dan tidak mampu mengkolonisasi inang dengan baik (Mejía et al. 2008). Cendawan endofit secara umum mempengaruhi pertumbuhan tanaman pada pembibitan, yaitu tinggi tajuk dan jumlah daun. Kemampuan cendawan endofit dalam meningkatkan pertumbuhan tanaman tidak terlepas dari kemampuannya memproduksi sejumlah metabolit pemacu tumbuh yang tinggi. Metabolit yang telah dilaporkan mampu diproduksi oleh cendawan endofit adalah indole acetic acid
(IAA), auksin, giberelin, dan sitokinin (Dai et al. 2008; Hamayun et al. 2010; Khan
et al. 2012). Peningkatan pertumbuhan tanaman oleh cendawan endofit disebabkan pula oleh adanya peningkatan jumlah akar rambut, percabangan akar rambut, dan akar-akar lateral meningkat, sehingga perakaran tanaman akan lebih luas dan lebih dalam. Perakaran tanaman tersebut akan meningkatkan penyerapan nutrisi dan pertumbuhan tanaman akan lebih baik, lebih vigor dan lebih tahan terhadap penyakit (Vasudevan et al. 2002).
19 dimungkinkan terjadi pada pola ekspresi gen dalam menentukan status simbiosis dengan tanaman tertentu (Rodriguez et al. 2009).
SIMPULAN DAN SARAN
Simpulan
Isolasi cendawan endofit cabai merah menghasilkan 237 isolat yang terdiri atas 24 isolat dari Cianjur, 65 isolat dari Bogor, dan 148 isolat dari Garut. Cendawan endofit dalam penelitian ini menunjukkan bahwa sebagian besar cendawan endofit pada tanaman cabai diperoleh dari daun dibandingkan bagian tanaman lainnya. Cendawan endofit hasil isolasi umumnya berpotensi fitopatogen, hanya berkisar antara 4-12% yang bersifat non fitopatogenik. Perlakuan cendawan endofit E9D10G2 mampu menekan penyakit antraknosa. Cendawan endofit E1Bh5G1 dan E3Cb7B2, memiliki kemampuan antibiosis terhadap C. capsici. Mekanisme antibiosis tidak selalu menentukan efektifitas penghambatan perkembangan penyakit antraknosa, akan tetapi mekanisme antibiosis merupakan salah satu mekanisme dalam pengendalian penyakit.
Saran
DAFTAR PUSTAKA
[BPS] Badan Pusat Statistik. 2015. Produksi cabai besar, cabai rawit, dan bawang merah tahun 2014. Berita Resmi Statistik: 71/08/XVIII. Jakarta (ID): Badan Pusat Statistik.
[KEMENTAN] Kementerian Pertanian Republik Indonesia. 2015. Rencana Strategis Kementerian Pertanian 2015-2019. Jakarta (ID): Kementerian Pertanian Republik Indonesia.
Agrios GN. 2005. Plant Pathology 5th Ed. Oxford (GB) :Elsevier Academic Pr. Arnold AE, Mejía LC, Kyllo D, Rojas EI, Maynard Z, Robbins N, Herre EA. 2003.
Fungal endophytes limit pathogen damage in a tropical tress. PNAS
100:15649-15654.
Bailey JA, O’Connell RJ, Pring RJ, Nash C. 1992. Infection strategies of
Colletotrichum species. In Colletotrichum. Biology, Pathology and Control. Bailey JA, Jeger MJ, editor. Wallingford (GB): CABI.
Barnett HL, Hunter BB. 1998. Illustrated Genera of Imperfect Fungi. Fourth Edition. Saint Paul (US): The American Phytopathological Society Pr. Chaisemsaeng P, Mongkolthanaruk W, Bunyatratchata W. 2013. Screening and
potential for biological control of anthracnose disease (Colletotrichum capsici) on chili fruits by yeast isolates. J Life Sci and Technol. 4(1): 202-204.
Chanchaichaovivat A, Ruenwongsa P, Panijpan B. 2007. Screening and identification of yeast strain from fruits and vegetables: potencial for biological control of postharvest chili antrachnose Colletotrichum capsici.
Biol Cont. 42:326-335.
Clarke BB, White Jr JF, Hurley RH, Torres MS, Sun S, Huff DR, 2006. Endophyte-mediated suppression of dollar spot disease in fine fescues. Plant Dis. 90: 994-998.
Clay K, Schardl C. 2002. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. American Naturalist 160:99-127.
Cook RJ, Baker KF. 1993. The Nature and Practice of Biological Control of Plant Pathogens. St Paul (US): APS Pr.
Dai C, Yu B, Li X. 2008. Screening of endophytic fungi that promote the growth of Euphorbia pekinensis. Afr J Biotechnol. 7:3505-3510.
Deising HB, Werner S, Wernitz M. 2000. The role of fungal appressoria in plant infection. Microb and Infect. 2:1631–1641.
Dingle J, Mc Gee PA. 2003. Some endophyte fungi reduce the density of pustules of Puccinia recondi f.sp. tritici in wheat. Mycol Res. 107:310-316.
Garg R, Kumar S, Kumar R, Loganathan M, Saha S, Kumar S, Rai AB, Roy BK. 2013. Novel source of resistance and differential reactions on chilli fruit infected by Colletotrichum capsici. Aus Plant Pathol. 42:227-233. doi:10.1007/s13313-012-0194-7.
Genarro M, Gonthier P, Nicolotti G. 2003. Fungal endophytic communities in healty and declining Quercus robur L. and Q. cerris L. tress in Northern Italy.
J Phytopathol. 151:529-534.
21 inhibitory effect to Fusarium oxysporum plant pathogenic fungi. HAYATI 20 (3):127-137.
Gundel PE, Helander M, Casas C, Hamilton C, Faeth S, Saikkonen K. 2012.
Neotyphodium fungal endophyte in tall fescue (Schedonorus phoenix): a comparison of three Northern European wild populations and the cultivar Kentuky-31. Fungal Divers. 1-10.
Gundel PE, Rudgers JA, Ghersa CM. 2011. Incorporating the process of vertical transmission into understanding of host symbiont dynamics. Oikos 120: 1121-1128
Hamayun M, Khan SA, Khan AL, Tang DS, Hussain J, Ahmad B, Anwar Y, Lee IJ. 2010. Growth promotion of cucumber by pure cultures of gibberellins-producting Phoma sp. GAH7. J Microbiol Biotechnol. 26:889-894.
Hammerschmidt R, Nuckles EM, Kuc J. 1982. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol Plant Pathol. 20:73-82.
Hariati N. 2007. Analisis keragaman 23 genotipe cabai (Capsicum sp.) berdasarkan penampakan fenotipik serta ketahanannya terhadap penyakit antraknosa (Colletotrichum sp.) [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Hospenthal DR, Rinaldi MG. 2008. Diagnosis and Treatment of Human Mycoses.Georgiev VS, editor. New Jersey (US): Humana Pers Inc.
Hyde KD, Soytong K. 2008. The fungal endophyte dilemma. Fungal Divers.
33:163-173.
Inacio J, Ludwig W, Spencer-Martins I, Fonseca A. 2010. Assessment of phylloplane yeasts on selected Mediterranean plants by FISH with group and species oligonucleotide probes. FEMS Microbiol Ecol. 71:61-72.
Istikorini Y. 2008. Potensi cendawan endofit untuk mengendalikan penyakit antraknosa pada cabai (Capsicum annuum L.) [disertasi]. Bogor (ID) : Institut Pertanian Bogor.
Joshee S, Paulus BC, Park D, Johnston PR. 2009. Diversity and distribution of fungal foliar endophytes in New Zealand Podocarpaceae. Mycol Res. 113: 1003-1015.
Kanchana-udomkan C, Taylor PWJ, Mongkol-porn O. 2004. Development of a bioassay to study anthracnose infection of Capsicum chinense Jacq. fruit caused by Colletotrichum capsici. Thai J Agric Sci. 37: 293-297.
Khan SA, Hamayun M, Khan Al, Lee IJ, Shinwari ZK, Kim J. 2012. Isolation of plant growth promoting fungi from dicots inhabiting coastal sand dunes of Korea. J Bot. 44:1453-1460.
Kiewnick S, Sikora RA.2006. Biological control of the root-knot nematode
Meloidogyne incognita by Paecilomyces lilacinus strain 251. Bio Cont.
38:179-187.
Kim WK, Cho EK, Lee ES. 1985. Variations of the anthracnose pathogen of pepper,
Colletotrichum gloeosporioides Perez. in morphology, pathogenicity and cultural characteristics. J Plant Pathol.1:211-212.
22
Kim KD, BJ Oh, Yang J. 1999. Differential interaction of a Colletotrichum gloeosporiodes isolate with green and red pepper fruits. Phytoparasitica
27(2):1-10.
Kleemann J, Rincon-Rivera LJ, Takahara H, Neumann U, van Themaat EVL, et al.
2012. Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum.
PLoS Pathogens 8(4): e1002643. doi:10.1371/journal.ppat.1002643.
Krauss J, Harri DA, Bush L, Power SA, Muller CB. 2007. Fungal grass endophytes, grass cultivars, nitrogen deposition and the associations with colonizing insects [Internet]. [diunduh 2015 Des 15] https://www.grassland.org.-nz/publications/nzgrassland_publication_2424.pdf
Lehtonen P, Helander M, Saikkonen K. 2005. Are endophyte-mediated effects on herbivores conditional on soil nutrients?. Oecologia 142:38-45.
Li HM, Sullivan R, Moy M, Kobayashi DY, Belanger FC. 2004. Expression of a novel chitinase by the fungal endophyte in Poa ampla. Mycologia 96 (3): 526-536.
Lu G, Cannon PF, Reid A, Simmons CM. 2004. Diversity and molecular relationships of endophytic Colletotrichum isolates from the Iwokrama Forest Reserve, Guyana. Mycol Res. 108:53-63.
Mahasuk P, Khumpeng N, Wasee S, Taylor PWJ, Mongkolporn O. 2008. Inheritance of resistance to anthracnose (Colletotrichum capsici) at seedling and fruiting stages in chili pepper (Capsicum spp.). Plant Breeding 128: 701-706.
Mejía LC, Rojas EI, Maynard Z, Bael SV, Arnold AE, Hebbar P, Samuels GJ, Robbins N, Herre EA. 2008. Endophytic fungi as biocontrol agents of
Theobroma cacao pathogens. Bio Cont. 46:4-14.
Mendgen K, Deising H. 1993. Infection structures of fungal plant pathogens-a cytological and physiological evaluation. New Phytol. 124:193-213.
Montri P, Taylor PWJ, Mongkolporn O. 2009. Pathotypes of Colletotrichum capsici the causal agent of chili antrachnose, in Thailand. Plant Dis. 93:17-20.
Nicholson RL, Moraes WBC. 1980. Survival of Colletotrichum graminicola: importance of the spore matrix. Phytopathology 70:255-261.
Nurahmi E, Mahmud T, Rossiana SS. 2011. Efektifitas pupuk organik terhadap pertumbuhan dan hasil cabai merah. J Floratek. 6:158-164.
O’Connell RJ, Perfect S, Hughes B, Carzaniga R, Bailey JA, et al. 2000. Dissecting the cell biology of Colletotrichum infection processes. Di dalam:
Colletotrichum. HostSpecificity, Pathology and Host-Pathogen Interaction.
Prusky D, Freeman S, Dickman MB, editor. St Paul (US):APS Pr. hlm 57-77. O’Connell RJ, Thon MR, Hacquard S, Amyotte SG, Kleemann J, et al. 2012.
Life-style transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nature Gen. 44(9):160-167. doi:10.1038/ng.2372.
Pakdeevaraporn P, Wasee S, Taylor PWJ, Mongkolporn O. 2005. Inheritance of resistance to anthracnose caused by Colletotrichum capsici in Capsicum.
23 Pérez LI, Gundel PE, Ghersa CM, Omacini M. 2013. Family issues: fungal endophyte protects host grass from the closely related pathogen Claviceps purpurea. Fungal Ecol. 6:379-386.
Popay AJ, Bonos SA. 2008. Biotic responses in endophytic grasses. Di dalam:
Neotyphodium in Cool-Season Grasses. Roberts CA, West CP, Spiers DE, editor. Los Angeles (US): Blackwell Publishing. hlm 163-185.
Prusky D, Plumbley RA. 1992. Quiescent infections of Colletotrichum in tropical and subtropical fruit. Di dalam: Colletotrichum. Biology, Pathology and Control .Bailey JA, Jeger MJ, editor. Wallingford (UK): CABI.
Rahman MA, Rahman MM, Azad AK, Alam MF. 2011. Inhibitory effect of different plant extracts and antifungal metabolites of Trichoderma strains on the conidial germination and germ tube growth of Colletotrichum capsici
causing chili anthracnose. IJAAR 1(1):20-28. ISSN: 2223-7054.
Raj TS, Christopher DJ. 2009. Effect of bio-control agents and fungicides against
Colletotrichum capsici causing fruit rot of chilli. Ann Plant Protect Sci.
17:143-145.
Raj TS, Christopher DJ, Suji AH. 2013. Evaluation of virulence and methods of inoculation of Colletotrichum capsici (SYD) Butler and Bisby. Int J Agric Sci. 9(2):802-805.
Raj TS, Christopher DJ, Suji HA. 2014. Morphological, pathogenic and genetic variability in Colletotrichum Capsici causing fruit rot of chili in Tamil Nadu, India. Afr J of Biotechnol. 13(17):1786-1790.
Rasmussen S, Parsons AJ, Popay A, Xue H., Newman JA. 2008. Plant-endophyte-herbivore interactions: more than just alkaloids?. Plant Signal and Behav.
3:974-977.
Rodriguez R, Redman R. 2008. More than 400 million years of evolution and some plants still can't make it on their own: plant stress tolerance via fungal symbiosis. J of Exp Bot. 59(5):1109-1114.
Rodriguez RJ, White Jr JF, Arnold AE, Redman RS. 2009. Fungal endophytes: diversity and functional roles. New Phytol. 182:314-330. doi:10.1111/j.1469-8137.2009.02773.x.
Rojas EI, Rehner SA, Samuels GJ, Van Bael SA, Herre EA, et al. 2010.
Colletotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panama: multilocus phylogenies distinguish pathogen and endophyte clades. Mycologia 102:1318–1338.
Saikkonen K, Saari S, Helander M. 2010. Defensive mutualism between plants and endophytic fungi?. Fungal Divers. 41:101-113. doi:10.1007/s13225-010-0023-7.
Schulz B, Boyle C, Draeger S, Rommert SK, Krohn K. 2002. Endophytic fungi: a source of novel biologically active secondary metabolites. Mycol Res.
106:996-1004.
Schulz B, Boyle C. 2005. The endophytic continuum. Mycologia 109(6):661-686. Schulz B, Rommert AK, Dammann U, Aust HJ, Strack D. 1999. The endophyte–
host interaction: a balanced antagonism?. Mycol Res.10:1275-1283.
Sharma G, Pinnaka AK, Shenoy BD. 2014. Infra-specific diversity of
Colletotrichum truncatum associated with chili anthracnose in India based on microsatelite marker analysis. Arch of Phytopathol and Plant Prot.