KAJIAN GENETIK KOMPONEN PRODUKSI DAN
KETAHANAN PECAH BUAH TOMAT
(
Solanum lycopersicum
L.) DI DATARAN RENDAH
MARLINA MUSTAFA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa disertas berjudul Kajian Genetik Komponen Produksi dan Ketahanan Pecah Buah Tomat (Lycopersicum esculentum L.) di Dataran Rendah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
MARLINA MUSTAFA. Kajian Genetik Komponen Produksi dan Ketahanan Pecah Buah Tomat (Solanum lycopersicum L.) di Dataran Rendah. Dibimbing oleh MUHAMAD SYUKUR, SURJONO H. SUTJAHJO dan SOBIR.
Pergeseran penanaman tomat dari dataran tinggi ke dataran rendah menyebabkan penurunan produksi tomat karena terbatasnya varietas unggul tomat yang adaptif pada dataran rendah. Rendahnya produksi tomat di dataran rendah disebabkan oleh meningkatnya suhu dari suhu optimum sehingga menghambat pembentukan buah dan berpengaruh terhadap penurunan kualitas buah akibat pecah buah. Perakitan varietas unggul yang adaptif dataran rendah merupakan metode yang efektif dan efisien untuk meningkatkan produksi tomat di dataran rendah. Untuk memperoleh varietas unggul diperlukan seleksi genotipe yang unggul dan tahan pecah buah yang dapat dijadikan sebagai tetua donor, dan menentukan kriteria seleksi serta pola pewarisan karakter tersebut untuk memudahkan proses seleksi.
Penelitian terdiri dari lima kegiatan yaitu (1) karakterisasi dan seleksi tiga puluh genotipe tomat berdasarkan komponen produksi, (2) karakterisasi dan seleksi tiga puluh genotipe tomat untuk ketahanan terhadap pecah buah, (3) pewarisan karakter kuantitatif dan kualitatif pada hipokotil dan kotiledon tomat, (4) studi pewarisan komponen produksi tomat di dataran rendah, dan (5) studi pewarisan pecah buah tomat di dataran rendah. Tujuan penelitian ini adalah (1) mengidentifikasi keragaman genetik tomat, karakterisasi genotipe tomat berdasarkan karakter komponen produksi, dan kriteria seleksi untuk komponen produksi yang tinggi pada tomat di dataran rendah, (2) mengidentifikasi tingkat ketahanan genotipe tomat dan kriteria seleksi terhadap pecah buah, (3) mempelajari pola pewarisan sifat kuantitatif dan kualitatif pada hipokotil dan kotiledon tomat, (4) mempelajari kendali genetik untuk pewarisan sifat komponen produksi pada tomat dan (5) mempelajari kendali genetik untuk pewarisan sifat ketahanan genotipe tomat terhadap pecah buah.
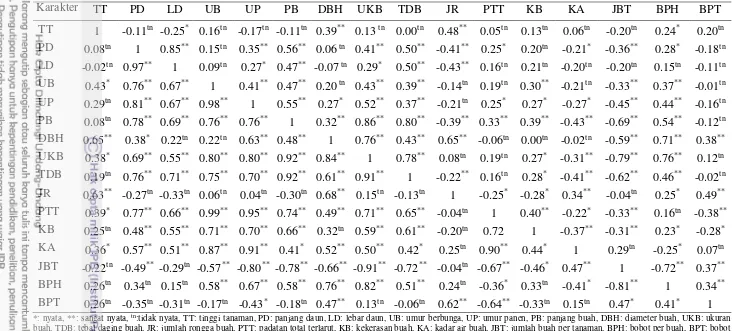
Hasil penelitian menunjukkan bahwa genotipe tomat yang diamati beragam dan mengelompok menjadi empat kelompok. Karakter yang dijadikan sebagai kriteria seleksi terhadap komponen produksi adalah karakter jumlah buah per tanaman dan bobot per buah. Karakter tersebut dipilih karena memiliki korelasi fenotipe dan genetik, pengaruh langsung yang tinggi terhadap produksi yaitu bobot buah per tanaman serta memiliki nilai heritabilitas yang tinggi.
Genotipe yang tahan terhadap pecah buah adalah IPBT4, IPBT56, IPBT60, IPBT64, IPBT83 dan IPBT85, dan genotipe yang rentan terhadap pecah buah adalah IPBT3. Karakter yang dijadikan kriteria seleksi terhadap pecah buah adalah jumlah rongga dan tebal daging buah karena memiliki korelasi fenotip dan genetik terhadap indeks pecah buah yang tinggi, memiliki pengaruh langsung terhadap pecah buah yang tinggi, serta nilai heritabilitas karakter tersebut juga tinggi.
Warna hipokotil dikendalikan oleh dua pasang gen epistasis dominan-resesif. Gen pengendali warna ungu bersifat dominan terhadap gen pengendali warna hijau pada hipokotil tomat. Tidak ada efek maternal yang berpengaruh pada pewarisan karakter hipokotil dan kotiledon. Metode pemuliaan yang tepat untuk karakter kualitatif dan kuantitatif hipokotil dan kotiledon tomat adalah pemanfaatan heterosis (hibrida).
Karakter produksi dan komponen produksi dikendalikan oleh banyak gen. Pola pewarisan karakter komponen produksi berbeda pada latar belakang populasi yang berbeda. Bobot buah per tanaman dikendalikan oleh aksi gen epistasis duplikat. Tidak ada efek maternal yang berpengaruh pada pewarisan produksi dan komponen produksi tomat.
Pecah buah tomat yang diamati pada penelitian ini terdiri dari dua yaitu pecah buah gabungan (radial dan konsentrik). Pecah buah gabungan dikendalikan oleh dua pasang gen epistasis komplementer dan pecah buah radial dikendalikan oleh dua pasang gen dengan aksi gen epistasis dominan lengkap oleh kedua tetua. Tidak ada pengaruh maternal pada pewarisan karakter pecah buah. Metode seleksi yang tepat untuk pengembangan varietas unggul tahan pecah buah adalah metode pedigree karena karakter pecah buah sudah dapat dilakukan seleksi pada generasi awal yaitu pada generasi F2.
SUMMARY
MARLINA MUSTAFA. Genetics Studies of Yield Component and Fruit Cracking Resistance of Tomatoes (Solanum lycopersicum L.) in Lowland. Suvervised by MUHAMAD SYUKUR, SURJONO H. SUTJAHJO dan SOBIR.
Tomato cultivation shifled from lowland to upland causing low production of tomato, because adaptive and superior tomato varieties were tricted in lowland. Low production of tomato in the lowland was caused by the increasing of optimum tempetarure so that inhibiting the farming of fruit and influencing against the decrease of fruit quality due to fruit cracking. Desgning adaptive varieties in the lowland was effective and efficient method to rise tomato production. To obtain the superior varieties was required great genotype selection and fruit cracking resistance that be as a parent donor and determining of selection criteria as well as the character inheritance patterns to facilitate selection prosess.
The research consisted of five activities, namely (1) the characterization and selection of thirty genotypes based on component production of tomato, (2) characterization and selection of thirty genotypes of tomato for resistance to fruit cracking, (3) inheritance character of quantitative and qualitative hypocotyl and cotyledons tomatoes, (4) inheritance of tomato components production in the lowlands, and (5) inheritance of tomatoes fruit cracking resistance in the lowlands. The aim of this research were (1) identify the genetic diversity of tomatoes, characterization of tomato genotype based characters of component production, and the selection criteria for high yield components in tomato in the lowlands, (2) identify the level of resistance genotypes of tomato and selection criteria of fruit cracking, (3) study the patterns of inheritance of quantitative and qualitative hypocotyl and cotyledons in tomatoes, (4) to study inheritance of yield component of tomatoes and (5) to study the inheritance of fruit cracking resistance of tomatoes.
This research uses thirty tomato genotype from IPB Plant Breeding Laboratory collection. Thirty tomato genotypes were selected at the first stage to identify genetic diversity and selection criteria based on yield components. These genotype were also used to select of resistant and susceptible genotypes and to determine the selection criteria for fruit cracking on the second stage. The selected genotype in the first stage would be used in the fourth experiment to study inheritance pattern of yield components characters based on the chosen character as selection criteria. The selected resistant and susceptible genotypes in the second experiment were used as a donor parent in five experiments to study inheritance pattern of fruit cracking resistance base on the selected of character selection. More effective and efficient selection in the early stage, namely sprout level which be carried out in the third experiment to study inheritance of tomato hypocotyl and cotyledons characters. Study of inheritance pattern in the third, fourth and fifth experiment was carried out with six generations method (biparental). Inheritance study for qualitative character used Mendel Analysis and for quantitative characters using the joint scaling test.
fruit. The characters were chosen due to have phenotype and genetic correlation, high direct influence against production, namely fruit weight per plant and have a high heritability.
The fruit cracking resistance genotype were IPBT4, IPBT56, IPBT60, IPBT64, IPBT83 dan IPBT85, dan susceptible genotype was IPBT3. The characters used as selection criteria against fruit cracking resistance were the number of locule and thick flesh because of owning phenotype and genetic correlation against high fruit cracking index, hashaving direct influence against high fruit cracking index, and thses characters heritability value was high.
Hypocotyl color is controlled by two pairs of epistasis dominant-recessive gene. The purple color controlling gene was dominant against the green color controlling genes of tomatoes hypocotyl. Maternal effect did not influence in inheritance of hypocotyl and cotyledons characters. Selection methods for qualitative and quantitative of hypocotyl and cotyledons tomato were heterosis utilization (hybrid).
The yield and yield components controlled by polygene. The inheritance pattern of yield components were different from distinct population background. Fruit weight per plant was controlled by duplicate epistasis gene action. There was no maternal effect on yield and yield component of tomatoes.
The observed tomato fruit cracking in this research refersed to two, namely combined fruit cracking (radial and concentric) and radial fruit cracking. The joint fruit cracking was controlled by two pairs of complementer epistasis genes and radial fruit cracking was controlled by two pairs of gene with complete dominant epistasis gene action by both parent donors. There was no maternal effect on inheritance of fruit cracking. The accurate selection method used to develop superior varieties of fruit cracking resistance was pedigree method because fruit cracking could be conducted through selection of early generation, namely F2 generation.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
KAJIAN GENETIK KOMPONEN PRODUKSI DAN
KETAHANAN PECAH BUAH TOMAT
(
Solanum lycopersicum
L.) DI DATARAN RENDAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Penguji pada Ujian Tertutup:
Dr Awang Maharijaya, SP MSi
(Staf peengajar pada Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor)
Dr Willy Bayuardi Suwarno, SP MSi
(Staf peengajar pada Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor)
Penguji pada Sidang Promosi :
Dr Willy Bayuardi Suwarno, SP MSi
(Staf peengajar pada Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor)
Dr Ir Yul Harry Bahar, MSi
PRAKATA
Alhamdulillahi rabbil’alamin. Puji syukur hanya untuk Allah SWT, dengan nikmat dan karunia-Nya, sehingga penulis untuk menyelesaikan disertasi ini yang merupakan kelengkapan tugas akhir padaProgram Doktor Sekolah Pascasarjana Institut Pertanian Bogor, dengan judul Kajian Genetik Komponen Produksi dan Ketahanan Pecah Buah Tomat (Solanum lycopersicum L.) di Dataran Rendah.
Penghargaan dan ucapan terima kasih penulis sampaikan kepada:
1. Prof Dr Muhamad Syukur, SP MSi, Prof Dr Ir Surjono H. Sutjahjo, MS dan dan Prof Dr Ir Sobir, MSi selaku komisi pembimbing atas segala bimbingan, arahan, dan saran selama penelitian hingga penyusunan disertasi ini.
2. Dirjen Dikti yang telah memberikan beasiswa pendidikan melalui program beasiswa unggulan Dikti
3. Rektor Universitas Hasanuddin dan Dekan Fakultas Pertanian yang telah memberikan rekomendasi untuk memperoleh beasiswa pada tahun 2011. 4. Rektor Universitas Sembilanbelas Nopember Kolaka yang telah memberikan
izin untuk menyelesaikan studi ini.
5. Rektor Institut Pertanian Bogor, Dekan Sekolah Pascasarjana dan Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman IPB yang telah menerima penulis untuk melanjutkan sekolah di IPB.
6. Dr Awang Maharijaya, SP, MSi dan Dr Willy Bayuardi Suwarno, SP, MSi, yang bersedia menjadi penguji luar komisi pada ujian pra kualifikasi dan ujian tertutup, yang banyak memberikan saran untuk perbaikan disertasi ini. 7. Dr Ir Suwardi, MAgr sebagai perwakilan dekan dan Dr Ir M Rahmat
Suhartanto, MSi sebagai perwakilan ketua Program Studi Pemuliaan dan Bioteknologi Tanaman saat ujian tertutup.
8. Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman saat ini Dr Ir Yudiwanti Wahyu EK, MS dan periode sebelumnya yaitu Dr Ir Trikoesoemaningtyas, MSc atas segala arahan dan motivasinya selama studi hingga penyelesaian disertasi ini.
9. Para dosen program studi pemuliaan dan bioteknologi tanaman IPB yang dengan keikhlasannya telah memberikan ilmu pengetahuan kepada penulis. Semua itu diberikan dengan dedikasi yang tinggi.
10. Pak Undang SP., M.Si dan pak Darwa atas bantuannya dalam memelihara tanaman selama penelitian.
11. Tim tomat (Helfi, Yuni Safitri, Estriana, Zamroh), Tim Solanaceae di Laboratorium Pemuliaan Tanaman IPB, atas segala bantuan tenaga, sharing informasi, dukungan, saran, kerjasama dan persaudaaran yang baik selama penelitian hingga penyusuan disertasi ini
12. Teman-teman PBT 2011, Anneke Pesik, Fitri Rachmawati, Agus Zainuddin, Heni Safitri, Anggelita Pujilestari, Rini Hermanasari, Walter Ajambang Nchu atas persaudaraan, kerjasama dan dukungannya selama studi.
13. Teman-teman Rumana Sulawesi Selatan secara umum dan tim pikitamijikoto satu secara khusus, atas persaudaraannya, atas bantuannya selama pelaksanaan penelitian.
15. Suamiku tercinta Lutfi Hair Djunur, ST, MT terima kasih atas segala doa dan kasih saying, perhatian dan kesabaran serta dukungan selama menyelesaikan studi.
16. Teristimewa kepada Ayahanda Mustafa Sirupa (alm) dan Ibunda tercinta Hj Rabbi, SPdI, atas segala doa restu, kasih sayang, perhatian dan kerja kerasnya, serta kesabaran dan dukungannya selama ini.
17. Kakakku tercinta Mardiana Mustafa, SKM., dan suami Drs. H. Hamka L., MS, terima kasih atas perhatian, pengertian dan kasih sayangnya serta segala bantuannya dalam penyelesaian studi, serta kepada ponakan-ponakanku atas segala keceriaan yang menjadi penyemangat menyelesaikan studi.
18. Keluarga besar Mertua saya Djunaid Wadda, atas segala perhatian, kasih sayang dan doanya yang menjadi motivator dalam menyelesaikan studi. 19. Keluarga besar Aliya Candik, Amir Candik, Arif Candik, atas segala
dukungan dan bantuannya selama studi.
20. Keluarga Besar Kadang (Alm), atas segala perhatian, motivasi dan doanya selama studi.
Kepada semua pihak yang telah membantu selama penelitian hingga penulisan studi ini, penulis ucapkan terima kasih. Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2016
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR x
DAFTAR LAMPIRAN xii
1 PENDAHULUAN 1
1.1 Latar Belakang 1
1.2 Tujuan Penelitian 2
1.3 Hipotesis 3
1.4 Ruang Lingkup Penelitian 3
2 TINJAUAN PUSTAKA 5
2.1 Botani dan Potensi Tanaman Tomat 5
2.2 Budidaya Tanaman Tomat di Dataran Rendah 6
2. 3 Pecah Buah pada Tomat 7
2.4 Studi Pewarisan Menggunakan Populasi Dasar 11
Daftar Pustaka 13
3 KARAKTERISASI DAN SELEKSI GENOTIPE TOMAT (Solanum lycopersicum L.) BERDASARKAN KARAKTER KOMPONEN
PRODUKSI 17
Abstrak 17
Abstract 18
3.1 Pendahuluan 19
3.2 Metode 19
3.3 Hasil dan Pembahasan 28
3.4 Simpulan 37
Daftar Pustaka 38
4 KARAKTERISASI DAN SELEKSI GENOTIPE TOMAT UNTUK
KETAHANAN TERHADAP PECAH BUAH 40
Abstrak 40
Abstract 41
4.1 Pendahuluan 42
4.2 Metode 42
4.3 Hasil dan Pembahasan 46
4.4 Simpulan 51
Daftar Pustaka 52
5 PEWARISAN SIFAT KARAKTER KUANTITATIF DAN
KUALITATIF PADA HIPOKOTIL DAN KOTILEDON TOMAT 55
Abstrak 55
Abstract 56
5.2 Metode 58
5.3 Hasil dan Pembahasan 62
5.4 Simpulan 69
Daftar Pustaka 70
6 STUDI PEWARISAN TOMAT TERHADAP KOMPONEN
PRODUKSI DI DATARAN RENDAH 73
Abstrak 73
Abstract 74
6.1 Pendahuluan 75
6.2 Metode 75
6.3 Hasil dan Pembahasan 78
6.3.7 Simpulan 97
Daftar Pustaka 98
7 STUDI PEWARISAN KETAHANAN TOMAT TERHADAP PECAH
BUAH 100
Abstrak 100
Abstract 101
7.1 Pendahuluan 102
7.2 Metode 103
7.3 Hasil dan Pembahasan 105
7.4 Simpulan 136
Daftar Pustaka 138
8 PEMBAHASAN UMUM 140
9 SIMPULAN DAN SARAN 145
9.1 Kesimpulan 145
9.2 Saran 145
DAFTAR PUSTAKA 146
LAMPIRAN 154
DAFTAR TABEL
3.1 Sumber Keragaman dan nilai harapan 27
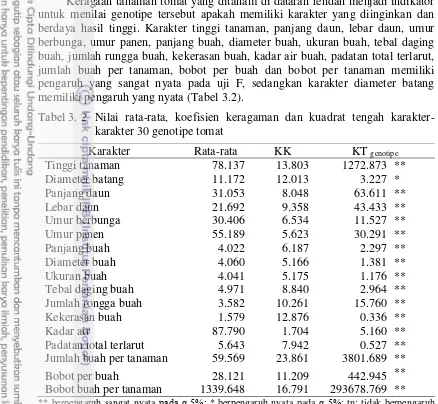
3.2 Nilai rata-rata, koefisien keragaman dan kuadrat tengah
karakter-karakter 30 genotipe tomat 28
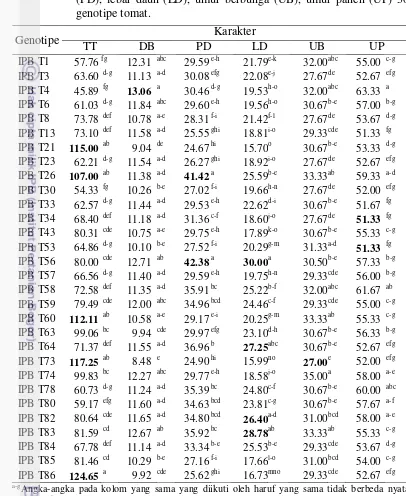
3.3 Keragaan tinggi tanaman (TT), diameter batang (DB), panjang daun (PD), lebar daun (LD), umur berbunga (UB), umur panen (UP) 30
genotipe tomat. 29
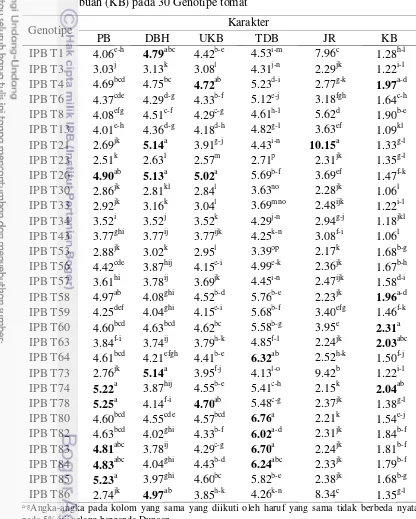
3.4 Keragaan panjang buah (PB), diameter buah (DBH), ukuran buah (UKB), tebal daging buah (TDB), jumlah rongga buah (JR),
kekerasan buah (KB) pada 30 Genotipe tomat 30
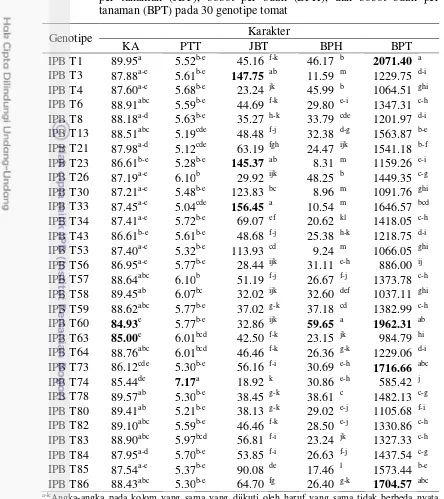
3.5 Keragaan kadar air (KA), padatan total terlarut (PTT), jumlah buah per tanaman (JBT), bobot per buah (BPH), dan bobot buah per
tanaman (BPT) pada 30 genotipe tomat 31
3.6 Nilai Heritabilitas beberapa karakter tomat 33
3.7 Korelasi fenotipe dan genetik antar karakter terhadap pecah buah
tomat 35
3.8 Pengaruh langsung dan tidak langsung masing-masing karakter
terhadap bobot buah per tanaman tomat 36
4.1 Persentase jumlah buah pecah per tanaman (JBP), persentase bobot buah pecah per tanaman (BBP), dan indeks pecah buah (IPB) pada
30 Genotipe tomat. 46
4.2 Nilai Heritabilitas beberapa karakter tomat 46
4.3 Nilai korelasi antar karakter terhadap pecah buah tomat dan nilai
heritabilitas karakter 47
4.4 Pengaruh langsung dan tidak langsung masing-masing karakter
terhadap indeks pecah buah tomat 50
5.1 Koefisien komponen genetik dalam uji skala gabungan 61 5.2 Nilai χ2 hitung warna hipokotil tomat pada populasi P1 (IPB T64),
P2 (IPB T3), BCP1 (F1 x IPB T64), BCP2 (F1 x IPB T3) dan F2
IPBT64 x IPB T3 63
5.3 Uji pengaruh tetua betina pada karakter hipokotil dan kotiledon
tomat 64
5.4 Potensi rasio, nilai skewness dan kurtosis pada populasi F2 karakter
panjang dan lebar kotiledon tomat 66
5.5 Hasil uji skala persilangan IPB T64 x IPB T3 66 5.6 Uji kecocokan model genetik beberapa karakter tomat 67 5.7 Pendugaan komponen genetik beberapa karakter tomat 67 5.8 Komponen ragam dan heritabilitas beberapa karakter tomat 68 6.1 Nilai tengah populasi karakter panjang buah tomat 78 6.2 Uji pengaruh tetua betina pada karakter panjang buah tomat 78 6.3 Aksi gen dan jumlah gen pengendali karakter panjang buah tomat 79 6.4 Uji kecocokan model genetik karakter panjang buah tomat 79 6.5 Pendugaan komponen genetik karakter panjang buah tomat 80 6.6 Komponen ragam dan dan nilai duga heritabilitas arti luas dan
6.8 Uji pengaruh tetua betina pada karakter diameter buah tomat 81 6.9 Aksi gen dan jumlah gen pengendali karakter diameter buah tomat 82 6.10 Uji kecocokan model genetik karakter diameter buah tomat 83 6.11 Pendugaan komponen genetik karakter diameter buah tomat 83 6.12 Nilai duga heritabilitas arti luas dan heritabilitas arti sempit karakter
diameter buah tomat 84
6.13 Nilai tengah populasi karakter ukuran buah tomat 84 6.14 Uji pengaruh tetua betina pada karakter ukuran buah tomat 85 6.15 Aksi gen dan jumlah gen pengendali karakter ukuran buah tomat 85 6.16 Uji kecocokan model genetik karakter ukuran buah tomat 86 6.17 Pendugaan komponen genetik karakter ukuran buah tomat 86 6.18 Nilai duga heritabilitas arti luas dan heritabilitas arti sempit karakter
ukuran buah tomat 88
6.19 Nilai tengah populasi karakter bobot per buah pada tomat 87 6.20 Uji pengaruh tetua betina karakter bobot per buah pada tomat 87 6.21 Aksi gen dan jumlah gen pengendali karakter bobot per buah pada
tomat 88
6.22 Uji kecocokan model genetik karakter bobot per buah pada tomat 88 6.23 Pendugaan komponen genetik karakter bobot per buah pada tomat 89 6.24 Nilai duga heritabilitas arti luas dan heritabilitas arti sempit karakter
bobot per buah pada tomat 90
6.25 Nilai tengah populasi karakter jumlah buah per tanaman pada tomat 90 6.26 Uji pengaruh tetua betina pada karakter jumlah buah per tanaman
pada tomat 90
6.27 Aksi gen dan jumlah gen pengendali karakter jumlah buah per
tanaman pada tomat 91
6.28 Uji kecocokan model genetik karakter jumlah buah per tanaman
pada tomat 91
6.29 Pendugaan komponen genetik karakter jumlah buah per tanaman
pada tomat 92
6.30 Nilai duga heritabilitas arti luas dan heritabilitas arti sempit karakter
jumlah buah per tanaman pada tomat 93
6.31 Nilai tengah populasi karakter bobot buah per tanaman pada tomat 93 6.32 Uji pengaruh tetua betina pada karakter bobot buah per tanaman
pada tomat 94
6.33 Aksi gen dan jumlah gen pengendali karakter bobot buah per
tanaman pada tomat 94
6.34 Uji kecocokan model genetik karakter bobot buah per tanaman pada
tomat 95
6.35 Pendugaan komponen genetik karakter bobot buah per tanaman pada
tomat 96
6.36 Nilai duga heritabilitas arti luas dan heritabilitas arti sempit karakter
bobot buah per tanaman pada tomat 97
7.1 Nilai tengah populasi karakter lebar daun tomat
7.6 Komponen ragam dan heritabilitas karakter lebar daun tomat 108 7.7 Nilai tegah populasi karakter panjang buah tomat 108 7.8 Uji pengaruh tetua betina pada karakter panjang buah tomat 109 7.9 Aksi gen, dan jumlah gen pengendali karakter panjang buah tomat 109 7.10 Uji kecocokan model genetik karakter panjang buah tomat 110 7.11 Pendugaan komponen genetik karakter panjang buah tomat 110 7.12 Komponen ragam dan heritabilitas karakter panjang buah tomat 111 7.13 Nilai tengah populasi karakter tebal daging buah pada tomat 112 7.14 Uji pengaruh tetua betina pada karakter tebal daging buah pada
tomat 112
7.15 Aksi gen, dan jumlah gen pengendali karakter tebal daging buah
pada tomat 113
7.16 Uji kecocokan model genetik karakter tebal daging buah pada tomat 113 7.17 Pendugaan komponen genetik karakter tebal daging buah pada tomat 114 7.18 Komponen ragam dan heritabjilitas karakter tebal daging buah pada
tomat 115
7.19 Nilai tengah populasi karakter jumlah rongga buah tomat 115 7.20 Uji pengaruh tetua betina pada karakter jumlah rongga buah tomat 116 7.21 Aksi gen, dan jumlah gen pengendali karakter jumlah rongga buah
tomat 116
7.22 Uji kecocokan model genetik karakter jumlah rongga buah tomat 117 7.23 Pendugaan komponen genetik karakter jumlah rongga buah tomat 117 7.24 Komponen ragam dan heritabilitas karakter jumlah rongga buah
tomat 118
7.25 Nilai tengah populasi karakter kadar air buah pada tomat 119 7.26 Uji pengaruh tetua betina pada karakter kadar air buah pada tomat 119 7.27 Aksi gen, dan jumlah gen pengendali karakter kadar air buah pada
tomat 119
7.28 Uji kecocokan model genetik karakter kadar air buah pada tomat 120 7.29 Pendugaan komponen genetik karakter kadar air buah pada tomat 120 7.30 Komponen ragam dan heritabilitas karakter kadar air buah pada
tomat 121
7.31 Nilai tengah populasi karakter jumlah buah per tanaman pada tomat 122 7.32 Uji pengaruh tetua betina pada karakter jumlah buah per tanaman
pada tomat 122
7.33 Aksi gen, dan jumlah gen pengendali karakter jumlah buah per
tanaman pada tomat 123
7.34 Uji kecocokan model genetik karakter jumlah buah per tanaman
pada tomat 123
7.35 Pendugaan komponen genetik karakter jumlah buah per tanaman
pada tomat 124
7.36 Komponen ragam dan heritabilitas karakter jumlah buah per
tanaman pada tomat 124
7.37 Nilai tengah karakter bobot per tanaman pada tomat 125 7.38 Uji pengaruh tetua betina pada karakter bobot per tanaman pada
tomat 125
7.39 Aksi gen, dan jumlah gen pengendali karakter bobot per tanaman
7.40 Uji kecocokan model genetik karakter bobot per tanaman pada tomat 126 7.41 Pendugaan komponen genetik karakter bobot per tanaman pada
tomat 127
7.42 Komponen ragam dan heritabilitas karakter bobot per tanaman pada
tomat 127
7.43 Nilai tengah populasi karakter persen buah pecah pada tomat 128 7.44 Uji pengaruh tetua betina pada karakter persen jumlah buah pecah
pada tomat 128
7.45 Aksi gen, dan jumlah gen pengendali karakter persen jumlah buah
pecah pada tomat 129
7.46 Uji kecocokan model genetik karakter persen jumlah buah pecah
pada tomat 129
7.47 Pendugaan komponen genetik karakter persen jumlah buah pecah
pada tomat 130
7.48 Komponen ragam dan heritabilitas karakter persen jumlah buah
pecah pada tomat 131
7.49 Nilai tengah populasi karakter persen bobot buah pecah per tanaman
pada tomat 131
7.50 Uji pengaruh tetua betina pada karakter persen bobot buah pecah
pada tomat 132
7.51 Aksi gen, dan jumlah gen pengendali karakter persen bobot buah
pecah pada tomat 132
7.52 Uji kecocokan model genetik karakter persen bobot buah pecah pada
tomat 133
7.53 Pendugaan komponen genetik karakter persen bobot buah pecah
pada tomat 134
7.54 Komponen ragam dan heritabilitas karakter persen bobot buah pecah
pada tomat 134
7.55 Nilai χ2 hitung ketahanan terhadap pecah buah gabungan populasi P1,
P2, BCP1, BCP2 dan F2 pada tomat 135
7.56 Nilai χ2 hitung ketahanan terhadap pecah buah radial populasi P1, P2,
BCP1, BCP2 dan F2 pada tomat 136
DAFTAR GAMBAR
1.1 Diagram Alur Penelitian 4
3.1 Letak daun pada sepertiga tanam bagian tengah (UPOV 2011) 21
3.2 Pembagian helai daun (UPOV 2011) 22
3.3 Tipe daun (IPGRI 2007) 22
3.4 Letak anak daun terhadap tulang daun utama (UPOV 2011) 22
3.5 Tipe tandan bunga (UPOV 2011) 22
3.6 Lapisan absisi pada tangkai buah (UPOV 2011) 23
3.7 Panjang pedisel (UPOV 2011) 23
3.8 Bentuk buah dalam penampang membujur (PPVT 2007) 23
3.9 Tulang buah pada ujung batang (UPOV 2011) 23
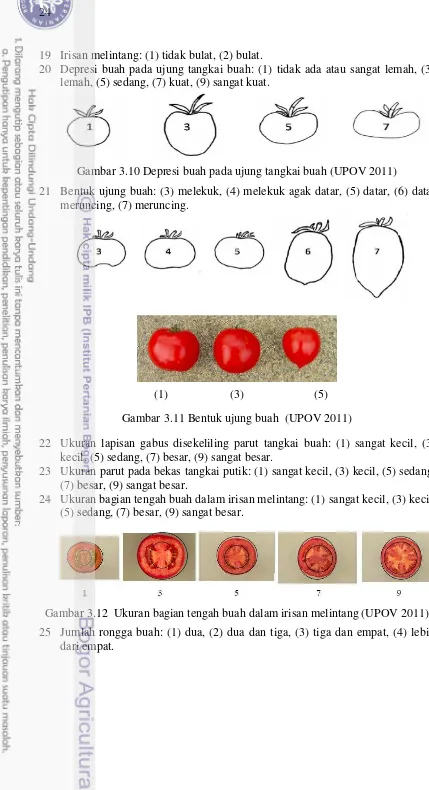
3.11 Bentuk ujung buah (UPOV 2011) 24 3.12 Ukuran bagian tengah buah dalam irisan melintang (UPOV 2011) 24
3.13 Jumlah rongga buah (UPOV 2011) 25
3.14 “Bahu buah hijau” sebelum masak (UPOV 2011) 25
3.15 Luas bahu buah hijau (UPOV) 25
3.16 Dendogram hasil analisis gerombol 30 genotipe tomat. 32 3.17 Diagram lintas beberapa karakter terhadap indeks pecah buah 34 4.1 Ilustrai scoring pecah buah, (A) tipe konsentrik, (B) tipe radial 44 4.2 Diagram lintas beberapa karakter terhadap indeks pecah buah 48
5.1 Skema persilangan 58
5.2 Warna hipokotil tomat IPBT64 (P1) x IPBT3 (P2) dan F1 IPBT64 x IPBT3. IPBT64: warna hipokotil hijau, IPBT3 warna hipokotil ungu 58 5.3 Sebaran populasi P1, P2, BCP1, BCP2, dan F2 panjang hipokotil,
panjang kotiledon dan lebar kotiledon tomat. 65
6.1 Skema persilangan 76
7.1 Skema persilangan 103
DAFTAR LAMPIRAN
1
PENDAHULUAN
1.1 Latar Belakang
Tomat merupakan salah satu sayuran yang mendapat prioritas untuk dikembangkan di Indonesia. Tomat sebagai salah satu komoditas sayuran mempunyai prospek pasar yang cerah. Tomat memiliki banyak kegunaan, baik sebagai sayuran maupun sebagai bahan baku industri makanan dan minuman. Potensi pasar tomat juga dapat dilihat dari segi harga yang terjangkau oleh seluruh lapisan masyarakat, sehingga membuka peluang yang lebih besar terhadap serapan pasar.
Apabila dilihat dari rata-rata produksinya, ternyata tomat di Indonesia masih rendah, yaitu 6.3 ton/ha jika dibandingkan dengan negara-negara Taiwan dan India yang berturut-turut 21 ton/ha dan 9.5 ton/ha. Rendahnya produksi tomat di Indonesia kemungkinan disebabkan varietas yang ditanam tidak cocok dengan kondisi lingkungan. Kebanyakan varietas tomat hanya cocok ditanam di dataran tinggi, namun seringkali terjadi penanaman tomat tanpa memperhatikan kondisi lingkungan, sehingga hasil dan kualitas buah yang dihasilkan sangat rendah. Kemampuan tomat untuk dapat menghasilkan buah sangat tergantung pada interaksi antara pertumbuhan tanaman dan kondisi lingkungannya (Wijayani dan Widodo 2005).
Di Indonesia tomat banyak diusahakan di dataran tinggi (60%) maupun di dataran rendah (40%). Hasil rata-rata pertanaman tomat di dataran rendah umumnya sekitar 6.0 ton/ha sedangkan di dataran tinggi dapat mencapai 26.6 ton/ha (Purwati 2007). Purwati (2009) melaporkan bahwa hasil tomat hibrida adaptif dataran rendah hingga tinggi yang ditanam di dataran medium (550 m dpl) hanya 1.95 kg tan-1, sedangkan potensi hasilnya bisa mencapai 3 kg tan-1 atau terjadi penurunan hasil sebesar 35%. Varietas yang beradaptasi baik pada dataran menengah hingga tinggi yang ditanam di dataran medium juga menunjukkan penurunan hasil dari 4-5 kg tan-1 menjadi 1.95 kg tan-1 atau sebesar 50-60%. Sebaliknya, jika varietas tersebut ditanam di dataran tinggi (800 m dpl) menunjukkan hasil 5.32 kg tan-1 (Soedomo 2012).
Pergeseran wilayah penanaman tomat ke dataran rendah menyebabkan resiko penurunan kualitas dan produksi buah. Rendahnya produksi di dataran rendah disebabkan oleh terbatasnya varietas unggul yang berpotensi hasil tinggi (Purnamaningsih 2008; Purwati 2007). Dataran rendah memiliki temperature yang tinggi. Temperatur yang tinggi tidak hanya berdampak pada waktu pemasakan buah tetapi juga pada laju pertumbuhan buah pada tomat (Adams, Cockshull dan Cave 2001). Peningkatan suhu 2-40C dari suhu optimum dilaporkan mempengaruhi perkembangan gamet dan menghambat pembentukan buah sehingga menurunkan produksi tomat (Peet et al. 1997; Sato et al. 2001; Firon et al. 2006).
memungkinkan masuknya pathogen, sehingga pada akhirya akan meningkatkan kehilangan panen yang cukup besar. Pecah buah pada tomat terjadi karena perubahan laju pertumbuhan buah yang cepat yang disebabkan oleh faktor genetik dan faktor lingkungan. Faktor lingkungan yang berpengaruh terhadap pecah buah adalah curah hujan, suhu, pemangkasan, intensitas cahaya, kelembaban udara, kandungan kalsium dan boron (Hudson 1956; Dorais et al. 2004; Maboko 2006; Liebisch et al. 2009). Pengaruh lingkungan tersebut sangat sulit dikontrol. Karena itu, penggunaan varietas unggul tomat dengan potensi hasil tinggi dan tahan pecah buah dipandang sebagai cara yang efektif untuk mengatasi permasalahan pada tomat yang ditanam pada dataran rendah.
Langkah awal yang harus dilakukan dalam kegiatan pemuliaan tanaman untuk perakitan varietas tahan adalah pembentukan populasi dasar dengan keragaman yang tinggi (Poespodarsono 1988). Keragaman genetik yang tinggi sangat menentukan keberhasilan pemuliaan untuk membentuk varietas unggul (Mangundidjojo 2003). Genotipe-genotipe yang telah dikoleksi kemudian dikarakterisasi, dianalisis keanekaragaman dan hubungan kekerabatannya untuk memudahkan dalam kegiatan pemuliaan tanaman. Langkah penting lainnya adalah melakukan skrining untuk memperoleh tetua donor. Selanjutnya adalah melakukan hibridisasi untuk memasukkan karakter tahan tersebut ke tetua recurrent. Seleksi diperlukan untuk memilih genotipe yang diinginkan dalam populasi bersegregasi keturunan hasil hibridisasi. Keakuratan dalam menentukan metode seleksi akan sangat menentukan keberhasilan perakitan tanaman.
Hingga saat ini informasi tentang studi pewarisan ketahanan terhadap pecah buah belum konsisten. Laporan AVRDC (1982), menyebutkan sifat pecah buah pada tomat dikendalikan oleh gen tunggal sederhana. Menurut Young (1959) pecah buah tipe radial dikendalikan oleh dua pasang gen mayor yaitu cr cr dan lr lr. Menurut Reynard, ketahanan pecah buah dikendalikan oleh dua pasang gen dengan sifat rentan dikendalikan oleh pasangan gen resesif (Pashar dan Lembeth 1960). Hasil penelitian Pashar dan Lembeth (1960) membuktikan bahwa sifat ketahanan terhadap pecah buah adalah sifat kuantitatif yang dikendalikan oleh gen major dan minor. Selanjutnya Amstrong dan Tompson (1967) dan Hernandes dan Nassar (1970) menyimpulkan bahwa sifat pecah buah pada tomat dikendalikan oleh gen ganda yang mempunyai sifat dominan sebagian. Berdasarkan hal tersebut, maka studi pewarisan tomat dengan potensi hasil tinggi dan tahan pecah buah perlu dilakukan untuk memperoleh informasi tentang kendali genetik pewarisan dan mendapatkan metode seleksi yang cepat dan akurat sehingga akan mempercepat diperolehnya tanaman tomat dengan produksi tinggi dan tahan terhadap pecah buah.
1.2 Tujuan Penelitian
Tujuan dari penelitian ini adalah
1 Mengidentifikasi keragaman genetik tomat, karakterisasi genotipe tomat berdasarkan karakter komponen produksi, dan kriteria seleksi untuk komponen produksi yang tinggi pada tomat di dataran rendah.
3 Mempelajari pola pewarisan sifat kuantitatif dan kualitatif pada hipokotil dan kotiledon tomat.
4 Mempelejari kendali genetik untuk pewarisan sifat komponen produksi pada tomat
5 Mempelajari kendali genetik untuk pewarisan sifat ketahanan genotipe tomat terhadap pecah buah.
1.3 Hipotesis
Hipotesis yang diajukan pada penelitian ini adalah:
1 Terdapat keragaman genetik, terdapat genotipe tomat yang memiliki karakter komponen produksi tinggi untuk dijadikan tetua pada pembentukan mapping population, dan beberapa karakter yang dijadikan kriteria seleksi untuk perakitan genotipe tomat unggul di dataran rendah.
2 Terdapat genotipe tomat yang sangat tahan dan genotipe yang rentan yang akan dijadikan tetua untuk analisis genetik resistensi tomat terhadap pecah buah dan minimal satu karakter sebagai kriteria seleksi ketahanan terhadap pecah buah.
3 Terdapat lebih dari satu gen yang terkait dengan sifat kualitatif dan kuantitatif hipokotil dan kotiledon tomat dengan pola pewarisan aditif, dominan, atau epistasis.
4 Terdapat lebih dari satu gen yang terkait dengan sifat komponen produksi dengan pola pewarisan aditif, dominan, atau resesif.
5 Terdapat lebih dari satu gen yang terkait dengan sifat ketahanan terhadap pecah buah dengan pola pewarisan aditif, dominan, atau resesif.
1.4 Ruang Lingkup Penelitian
Kegiatan penelitian ini terdiri dari beberapa tahapan percobaan sebagai berikut:
1 Karakterisasi dan seleksi genotipe tomat berdasarkan karakter komponen produksi
2 Karakterisasi dan seleksi genotipe tomat untuk ketahanan terhadap pecah buah
3 Studi pewarisan sifat karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon tomat
4 Studi pewarisan tomat terhadap komponen produksi di dataran rendah 5 Studi pewarisan ketahanan tomat terhadap pecah buah di dataran rendah
Plasma nutfah tomat koleksi
1. Karakterisasi dan seleksi berdasarkan komponen produksi
2. Karakterisasi dan kriteria seleksi ketahanan terhadap pecah buah radial,
konsentrik dan gabungan
1a. Analisis komponen utama dan analisis gerombol
1b. Sidik ragam, uji nilai tengah, korelasi, heritabilitas arti luas
dan analisis lintas
2a. Sidik ragam, indeks pecah buah, uji nilai tengah,
heritabilitas arti luas
2b. Korelasi dan analisis lintas
Genotipe terpilih
Genotipe terpilih
Hibridisasi genotipe terpilih
(Pembentukan populasi F1, F1R, BCP1, BCP2 dan F2)
4. Studi pewarisan terhadap komponen produksi
3. Pewarisan karakter kualitatif dan kuantitatif hipokotil dan
kotiledon
5. Studi pewarisan ketahanan pecah buah (5a. Pecah buah radial, 5b. Pecah buah konsentrik, 5c. Pecah buah gabungan)
Pengaruh maternal, derajat dominansi, aksi gen, jumlah faktor efektif, Heritabilitas arti
luas, heritabilitas arti sempit
Metode seleksi dan kendali genetik untuk pewarisan komponen produksi dan ketahanan pecah buah radial,
konsentrik dan gabungan pada tomat dataran rendah Hibridisasi genotipe terpilih
(Pembentukan populasi F1, F1R, BCP1, BCP2 dan F2)
2
TINJAUAN PUSTAKA
2.1 Botani dan Potensi Tanaman Tomat
Tanaman tomat termasuk tanaman diploid dan memiliki jumlah kromosom dasar x = 12. Jumlah kromosom normal tomat adalah 2n = 2x = 24. Tomat termasuk simple diploid genetik dengan ukuran genom yang kecil, periode reproduksi yang singkat. Diprediksikan bahwa tomat memiliki ~ 35 000 gen dengan rata-rata kerapatan gen 9,8 kb/gen (Cicedo dan Peralta 2013). Tanaman tomat tergolong ke dalam famili Solanaceae dan genus Lycopersicon. Genus Lycopersicon terdiri dari dua sub genus yang berbuah merah disebut Eulycopersicon dan yang berbuah hijau disebut sub genus Eriopersicon. Sub genus Eulycopersicon meliputi L. esculentum Mill. dan L. pimpinellifolium Jusl Mill. Eriopersicon mempunyai warna hijau yang meliputi L. cheesmanii Riley, L. glandulosum C. H. Muller, L. hirsutum Humb & Boupl dan L. peruvianum mill. Spesies esculentum merupakan spesies penting dan paling banyak dibudidayakan.
Bentuk buah tomat terdiri atas bentuk buah bulat, pear, lonjong dan oval. Warna buah merupakan produk dari kombinasi pigmen yang terdapat pada jaringan epicarp dan subepidermis. L. esculentum, L. cheesmaniae, L. galapagense, L. pimpinellifolium memiliki pigmen karatenoid buah masak berwarna merah, orange dan kuning. Warna buah masak L. esculentumi dan L. pimpinellifolium adalah merah yang merupakan akumulasi likopen, sedangkan spesies L. cheesmaniae dan L. galapagense memiliki warna buah yang kuning ketika masak. Umumnya buah tomat sebelum masak berwarna hijau, dan sebagian berwarna ungu yang merupakan akumulasi dari pigmen antosianin (Cicedo dan Peralta 2013).
Di Indonesia, tomat sudah dijadikan sebagai salah satu sayuran yang mendapat prioritas untuk dikembangkan. Tomat sebagai salah satu komoditas sayuran mempunyai prospek pasar yang baik karena merupakan salah satu jenis sayuran yang banyak dikonsumsi oleh masyarakat. Tomat memiliki banyak kegunaan, baik sebagai sayuran maupun sebagai bahan baku industri makanan dan minuman. Potensi pasar tomat juga dapat dilihat dari segi harga yang terjangkau oleh seluruh lapisan masyarakat, sehingga membuka peluang yang lebih besar terhadap serapan pasar.
Luas area budidaya tomat di Indonesia meningkat dari tahun ke tahun. Tahun 2007-2010, luas area budidaya tomat meningkat dari 51 523 hektar menjadi 61 154 hektar. Pada tahun 2011 mengalami penurunan menjadi 53 088 hektar. Luas area penanaman tomat sangat berpengaruh terhadap produksi tomat di Indonesia. Produksi tomat nasional pada tahun 2007-2011 meningkat dari 635 474 ton menjadi 954 046 ton per tahun (BPS 2012).
bagian dalam, terdiri atas jumlah biji, ketebalan daging, dan kandungan lender, 3) kualitas rasa, terdiri atas rasa manis, rasa asam, kekenyalan dan jumlah air buah.
Ameriana (1995) melaporkan bahwa persepsi konsumen mengenai kualitas buah terdiri dari kualitas eksternal dan internal (organoleptik). Kualitas eksternal yang terpenting hingga kurang penting berdasarkan persepsi konsumen adalah warna, kekerasan, bentuk dan ukuran buah. Kualitas internal yang terpenting hingga kurang penting adalah rasa manis, rasa asam, kekenyalan, dan kadar air buah. Hasil penelitian Purwati (2007) melaporkan bahwa masyarakat pada umumnya menyukai buah tomat yang warna kulitnya merah terang (dengan nilai 7-8 hasil pengukuran CBT color chart), kekerasan buah sedang (nilai 110-130 mm/50 g/10 det pengukuran dengan penetrometer), bentuk buah agak lonjong (sphericity indeks 99-199), ukuran buah agak besar (volume 80-90 cm3), rasa buah manis (gula reduksi 4.25-5.0%), tidak masam (kadar total asam 0.34-0.37%), banyak mengandung air buah (kandungan air 92-93%), dan buahnya kenyal/renyah.
Warna buah tomat adalah salah satu atribut paling penting dan kompleks. Kompleksitas warna tomat adalah karena adanya sistem pigmen karotenoid yang beragam dengan penampilan dikondisikan oleh jenis dan konsentrasi pigmen, dan tunduk pada regulasi genetik dan lingkungan (Arias et al. 2000; Lopez dan Gomez 2004). Keragaman pigmen karatenoid membantu peoses retionoid dan mengandung Provitamin A yang sangat baik untuk kesehatan. Provitamin A dalam bentuk betakaroten paling banyak terdapat pada tomat. Tomat juga merupakan sumber utama likopene yang memiliki kapasitas mengikat oksigen radikal sehingga sangat bermanfaat untuk mencegah penyakit kanker, penyakit cardiovascular dan penyakit kronik lainnya. Kandungan vitamin C juga dilaporkan sangat tinggi terdapat pada tomat, yang merupakan sumber vitamin C tertinggi setelah orange (Cicedo dan Peralta 2013).
Kualitas buah yang diinginkan untuk konsumsi rumah tangga berbeda dengan standar kualitas untuk industri. Bagi konsumen dalam negeri, tomat dikonsumsi sebagai substitusi buah-buahan dan sebagai pelengkap bumbu masak. Untuk konsumsi sebagai substitusi buah-buahan, konsumen lebih mengutamakan tomat dengan rasa manis, sedikit asam, renyah dan mempunyai kandungan air sedang. Untuk keperluan industri, kriteria tomat olahan adalah memiliki padat total terlarut tinggi (+ 4.5o Brix), pH rendah (+ 4.4), kompak, mudah dikuliti, tahan terhadap retak, dan warnanya merah cerah (Villreal 1981).
Terdapat perbedaan komponen kualitas buah antara tomat tipe konsumsi segar dan tipe olahan. Menurut Tigchelaar (1986) beberapa parameter kualitas tipe olahan adalah warna, pH, total keasaman, total zat terlarut dan viskositas. Sedangkan untuk tipe konsumsi segar adalah : kekerasan buah, keseragaman bentuk, warna, ukuran dan bebas dari kerusakan/kelainan fisik (defect) seperti pecah buah.
2.2 Budidaya Tanaman Tomat di Dataran Rendah
tidak sensitif terhadap panjang hari dan tandan buahnya mampu tahan terhadap panjang hari selama 7 sampai 19 jam (Gould 1974). Menurut Sunarjono (2007) musim tanam terjadi pada musim kemarau. Tomat lebih memuaskan ditanam didaerah kering dan sejuk di pegunungan daripada di dataran rendah, sehingga ukuran buah jadi lebih baik.
Penurunan daya hasil tomat di dataran rendah dipengaruhi suhu lingkungan tumbuh yang menyebabkan ukuran buah lebih kecil dan jumlah buah yang terbentuk sedikit atau fruitset bernilai kecil. Firon et al (2006) melaporkan bahwa pada kondisi suhu tinggi jumlah dan kualitas serbuk sari tomat berkurang, selanjutnya viabilitas serbuk sari juga berkurang yang akhirnya menyebabkan fruitset dan jumlah benih per buah berkurang.
Proses keberhasilan pembungaan dan pembuahan juga dipengaruhi oleh faktor abiotik, khususnya suhu udara. Tanaman tomat memerlukan suhu siang dan malam hari sebesar ±24oC dan ±18oC untuk pertumbuhan khususnya pembungaan dan pembuahaan.Suhu pada malam hari merupakan faktor kritis untuk pembentukan buah (Lai 1993). Buah yang sudah terbentuk tidak semua dapat tumbuh terus sampai menjadi matang. Faktor-faktor yang mempengaruhi tidak semua buah dapat terus tumbuh hingga matang antara lain :jumlah bunga yang dihasilkan, persentase bunga yang mengalami penyerbukan, persentase bunga yang menjadi buah, persentase buah muda yang dapat tumbuh terus sampai menjadi buah masak (Purwati 2008). Oleh sebab itu, keberhasilan pembungaan dan pembuahan bukan hanya ditentukan dari faktor lingkungan tetapi juga ditentukan dari faktor genetik.
2. 3 Pecah Buah pada Tomat
Pecah buah (fruit cracking) merupakan kelainan fisiologi yang sering terjadi pada pertanaman tomat. Berdasarkan bentuknya, pecah buah tomat dapat dibedakan menjadi burst cracking, concentric cracking, russeting, radical cracking (Peet 1992). Bentuk radial banyak dijumpai pada buah yang sudah matang penuh, dan paling banyak dijumpai. Bentuk konsentrik yang sering terjadi pada buah yang matang hijau. Bentuk konsentrik ini tidak terlalu banyak menurunkan produksi buah di lapang apabila dibanding bentuk radial (Thompson dan Kelly 1957).
Prashar dan Lambeth (1960) menduga pecah buah terjadi akibat pengembangan jaringan yang tidak terkoordinasi selama pertumbuhan atau hanya merupakan peristiwa akibat pembengkakan saja. Beberapa faktor yang diduga menjadi faktor penyebab terjadinya pecah buah adalah :
2.3.1 Genetik
dikendalikan oleh pasangan gen resesif (Pashar dan Lambeth 1960). Selanjutnya, dalam penelitian Phasar dan Lambeth (1960) telah dibuktikan bahwa sifat ketahanan terhadap pecah buah adalah sifat kuantitatif yang dikendalikan oleh gen major dan minor. Persilangan antara dua tetua yang resisten diperoleh F1 dan F2 mulai dari kelas rentan sampai resisten, dilaporkan juga bahwa pada sesama varietas resisten mempunyai susunan genetik yang berbeda.
Amstrong dan Tompson (1967), melakukan uji dialel terhadap tiga belas varietas tomat. Hasil percobaannya menunjukkan galur resisten mempunyai daya gabung umum tertinggi terhadap pecah buah. Disamping itu diketahui bahwa sifat pecah buah pada tomat dikendalikan oleh gen ganda yang mempunyai sifat dominan sebagian. Pendapat tersebut sesuai dengan hasil penelitian Hernandes dan Nassar (1970) yang menyatakan bahwa sifat ketahanan terhadap pecah buah tomat dikendalikan oleh gen ganda (poligenik) yang bersifat dominansi sebagian. 2.3.2 Pengairan yang Tidak Teratur
Pemberian air khususnya dari kondisi yang sangat kering ke kondisi yang sangat basah dapat menyebabkan terjadinya pecah buah. Mekanisme tersebut dapat dijelaskan dengan penelitian Kamimura et al. (1972). Hasil penelitian tersebut menyatakan bahwa kelembaban tanah yang tinggi akan menurunkan kekuatan perenggangan (‘tensile strength’) kulit buah tomat. Bertambahnya kecepatan pertumbuhan buah dalam kondisi seperti ini akan mendorong terjadinya keretakan kecil ini, dan akan berkembang menjadi pecah buah.
2.3.3 Temperatur dan Cahaya yang Tinggi
Meningkatnya temperatur secara drastis dapat menyebabkan terjadinya peningkatan tekanan pada kulit buah akibat adanya pulp dalam kulit buah tersebut. Tekanan tersebut akan menurunkan tingkat kekakuan dan kelenturan kulit buah, dan mendorong terjadinya pecah buah (Lang dan During 1990). Menurut hasil penelitian Awan (1983) jumlah buah yang mengalami pecah buah lebih banyak terjadi pada penanaman di dalam rumah plastik apabila dibanding dengan penanaman di lapang. Hal ini kemungkinan disebabkan karena tingginya suhu udara yang terjadi didalam rumah plastik.
Intensitas penyinaran yang tinggi secara tidak langsung akan mempengaruhi terjadinya pecah buah. Adanya penyinaran yang tinggi akan meningkatkan fotosintesis dan padatan terlarut buah. Meningkatnya padatan terlarut buah akan menurunkan potensial air buah, sehingga air dari batang maupun daun akan masuk kedalam buah. Kondisi ini akan meningkatkan tekanan terhadap kulit buah. Tekanan terhadap kulit buah akan menyebabkan terjadinya pecah buah, walaupun tidak segera terjadi. Menurut Ackley dan Krueger (1980) pada juice buah ceri, konsentrasi larutan buah yang tinggi berkaitan erat dengan kepekaan buah tersebut terhadap pecah buah.
2.3.4 Anatomi Buah
(‘elasticity’) kulit buah pada periode breaker stage sampai pink stage sangat diperlukan apabila dikaitkan dengan pecah buah. Kultivar yang resisten terhadap pecah buah paling sedikit mempunyai salah satu diantara sifat diatas.
2.3.5 Kecepatan Tumbuh Buah
Kecepatan tumbuh buah yang berlebihan dapat memacu terjadinya pecah buah. Apabila dilakukan pemangkasan atau penjarangan buah, maka persaingan antar buah dalam memperoleh assimilat akan berkurang. Akibatnya kecepatan tumbuh buah semakin bertambahnya. Menurut Peet (1992), pada buah tomat yang menerima asimilat lebih banyak akan mengalami pertumbuhan yang lebih cepat dan buah tersebut lebih rentan terhadap pecah buah.
2.3.6 Kelembaban yang Tinggi
Kelembaban udara yang tinggi dapat meningkatkan kejadian pecah buah tomat dalam rumah kaca, khususnya pada suhu yang tinggi. Tekanan air dan udara dalam buah tomat akan meningkat dengan meningkatnya suhu. Terdapat dua mekanisme dalam mengatasi kondisi ini : pertama dengan pelonggaran kulit buah, kedua dengan mengirim kembali air ke batang atau daun. Apabila kelembaban udara tinggi mekanisme kedua tidak akan terjadi karena transpirasi rendah. Dalam kondisi seperti ini mekanisme pertama yang akan terjadi. Hal ini akan mendorong terjadinya pecah buah (Peet 1992).
2.3.7 Kekuatan Dinding Sel Buah
Dari beberapa teori di atas secara tidak langsung pecah buah berkaitan erat dengan kekuatan kulit (epidermis) buah. Mekanisme penguatan dan pelenturan kulit buah dapat diperjelas dengan melihat proses yang terjadi pada dinding sel.
Mekanisme pelenturan dinding sel primer bisa terjadi pada interaksi selulosa, hemiselulosa dan hidroksiprolin. Menurut Wareing dan Philips (1981) dalam dinding sel primer serat mikro selulose terbenam dalam suatu matrik protein, pectin dan hemiselulose. Xyloglucans merupakan hemiselulosa yang berikatan kovalen dengan matrik yang lain, dan berikatan hydrogen dengan serat-serat mikro selulose. Terputusnya ikatan-ikatan hydrogen antara xyloglucans dengan serat-serat mikro selulose menyebabkan dinding sel mudah bergeser, sehingga dinding sel akan mudah meregang. Mekanisme fisik dan kimia yang menyebabkan terputusnya ikatan hidrogen ini memudahkan dinding sel bergerak sehingga lebih elastis. Hal ini merupakan faktor yang perlu dipertimbangkan dalam kasus pecah buah pada tomat.
dan Phillips, 1981). Apabila kalsium tidak terdapat dalam dinding sel, maka rigiditas akan menurun, hal ini akan memudahkan sel-sel tersebut pecah bila sel mengadakan pembesaran.
2.3.8 Auksin
Auksin adalah salah satu hormone yang tidak terlepas dari proses pertumbuhan dan perkembangan tanaman. Salah satu peran fisiologi auksin adalah pembesaran sel.
Mekanisme ekspansi sel kemungkinan disebabkan oleh longgarnya/ bergesernya (loosening) dari dinding sel primer. Menurut Moore (1979) auksin berfungsi mengaktifkan pompa ion H+ yang terletak dalam plasmalema. Hal ini akan mengakibatkan rendahnya pH pada dinding sel, sehingga ikatan hydrogen antara xyloglucan dengan serat mikroselulose pada dinding sel primer akan terputus. Dengan demikian dinding sel tersebut akan mudah mengalami pelonggaran.
IAA (Indol Acetic Acid) adalah auksin endogen yang terdapat dan disintesis dalam tanaman. Senyawa-senyawa lain yang mempunyai aktivitas serupa auksin dan merupakan senyawa sintetik salah satunya adalah NAA atau Naphthalene Acetic Acid (Wattimena 1988).
Menurut Yamaoto, Satch dan Watanabe (1992), NAA dapat menekan terjadinya pecah buah pada sweet cherry. Aplikasi NAA 24 hari sebelum panen pada konsentrasi 0.5, 1, dan 2 ppm dapat menekan terjadinya pecah buah hingga 13-40 % dari kontrol. Disamping itu menurut Byers et al. (1990) aplikasi GA4+7 dapat menekan terjadinya pecah buah pada apel dari 56 % menjadi 21 %. Dinyatakan pula bahwa kombinasi GA4+7, Daminozide, NAA dan Vapor Gard dapat menekan terjadinya pecah buah dari 93 % menjadi 22 %. Hal ini disebabkan karena Giberelin dapat meningkatkan aktivitas auksin dalam tanaman.
2.3.9 Kalsium
Kalsium merupakan hara makro esensial yang berfungsi dalam meningkatkan integritas sel dan permeabilitas membrane plasma. Disamping itu juga dapat mendorong perkecambahan dan pertumbuhan polen, mengaktifkan beberapa enzim dalam pembelahan dan pemanjangan sel. Kalsium juga berfungsi untuk membantu sintesis protein, transfer karbohidrat, dan keberadaannya dapat menekan efek racun dari logam berat dalam tanaman (Jones et al. 1991).
Berbeda dengan hara makro lain, proporsi yang tinggi dari total kalsium dalam jaringan tanaman terdapat pada dinding sel (apoplas). Kalsium dalam tanaman terdapat dalam berbagai bentuk yaitu : bentuk terlarut, terikat oleh pektat, oksalat, fostat. Bentuk terlarut banyak terdapat dalam vakuola demikian juga bentuk terikat pektat terdapat dalam dinding sel (lamela tengah dan permukaan luar membran plasma). Dalam kondisi suplai kalsium yang rendah bentuk pektat dapat terjadi sebanyak 50 % dari total kalsium (Marschner 1986).
Kandungan kalsium buah tomat menurut Bollard (1970) sebanyak 0.13 % berat kering, lebih rendah apabila dibandingkan dengan kandungan kalsium pada daun (6.08 %). Sedangkan kandungan unsur-unsur nitrogen, fosfor, kalium antara daun dan buah tidak jauh berbeda. Hal ini menunjukkan bahwa kalsium yang berada dalam daun tidak ditranslokasikan ke dalam buah bersama assimilat seperti terjadi pada unsur-unsur yang lain. Sehingga menurut Link (1974) dan Redmond (1975), pemberian kalsium yang efektif untuk peningkatan kualitas buah adalah dengan penyemprotan langsung terhadap buah tersebut.
Menurut Knott (1962) aplikasi kalsium pada tomat dengan penyemprotan pada daun menggunakan kalsium pada tomat menggunakan kalsium khlorida (CaCl2 36.1 % Ca) dapat dilakukan pada konsentrasi 6000-12000 ppm kalsium khlorida, dan bentuk kalsium nitrat (Ca(NO3)2) 28 % Ca) pada konsentrasi 6000-18000 ppm kalsium nitrat. Menurut Yamamoto, Satoh dan Watanabe (1992) aplikasi kalsium dalam bentuk kalsium nitrat (Ca(NO3)2) pada konsentrasi 0.5 % (5000 ppm) 7 hari sebelum panen dapat menekan pecah buah pada sweet cherry.
2.4 Studi Pewarisan Menggunakan Populasi Dasar
Menurut Mather dan Jinks (1977) terdapat dua rancangan yang dapat digunakan untuk studi pewarisan, yaitu rancangan generasi-generasi dasar dan rancangan kawin acak. Rancangan generasi-generasi dasar menggunakan populasi tetua P1, P2, F1, F1R, BCP1, BCP2 dan F2. Dengan rancangan ini dapat dilakukan pendugaan semua komponen ragam, efek maternal, interaksi antar alel, dan interaksi genotype lingkungan dan heritabilitas (Sastrosumarjo 1987; Sing dan Chaudhary 1979; Roy 2000). Rancangan persilangan kawin acak biasanya menggunakan metode dialel.
Asumsi yang digunakan untuk melakukan analisis statistik dan analisis genetik guna melacak gen-gen pengendali karakter tersebut adalah (1) tidak ada pengaruh lingkungan, (2) tidak ada pengaruh dominansi antar alel, (3) tidak adaa pengaruh epistasis, (4) gen memberikan pengaruh yang sama dan bersifat aditif untuk semua lokus, (5) tidak ada pautan gen, dan (6) tetua dalam keadaan homozigositas lengkap dan tanaman F1 dalam keadaan heterozigositas lengkap (Phoehlman 1979).
Populasi F2 sering dimanfaatkan untuk mempelajari pewarisan karakter melalui analisis segregasi. Suatu karakter yang dikendalikan oleh satu atau dua gen mayor akan menunjukkan sebaran populasi F2 diskontinu sehingga tanaman mudah diidentifikasi menjadi tahan atau rentan. Peran lingkungan biasanya kecil, maka ragam fenotip yang ditampilkan dam populasi bersegregasi sebagian besar merupakan ragam genetik dan merupakan akibat adanya pengaruh dominan (Allard 1960). Sebaliknya, jika karakter tersebut dikendalikan oleh gen minor atau poligenik maka akan menunjukkan sebaran populasi F2 yang kontinyu.
(interlokus/epistasis) (Crowder 1993). Gen yang saling berinteraksi memiliki beberapa kemungkinan pengaruh derajat dominansi yaitu derajat dominansi aditif, dominan dan epistasis. Derajat dominansi aditif menggambarkan besarnya nilai rata-rata populasi heterozigot (F1) yang sama dengan nilai rata-rata kedua tetua (MP). Derajat dominansi dominan terdiri atas tiga bentuk yaitu: dominan parsial dimana nilai rata-rata F1 melebihi nilai rata-rata kedua tetua (MP); dominan lengkap yaitu nilai rata-rata F1 setara dengan nilai rata-rata tetua; overdominan yaitu nilai rata-rata F1 melebihi nilai tetua. Derajat dominansi epistasis terjadi ketika pola perbandingan bilai rata-rata F1 beragam terhadap nilai tengah populasi tetua karena derajat dominansi pada satu lokus bersifat saling menutupi pengaruh gen pada lokus lain (Petr dan Frey 1966; Roy 2000; Mangoendidjojo 2003).
Daftar Pustaka
[AVRDC] Asian Vegetable Research and Development Center. 1982. Tomato Breeding. A progress report. Taiwan. 27 p.
Ackley WB, Krueger WH. 1980. Overheat irrigation water quality and the cracking of sweet cherries. Hort Sci. 15:289-290.
Adams SR, Cockshull KE, Cave CRJ. 2001. Effect of Temperature on the Growth and Development of Tomato Fruits. Annals of Botany. 88: 869-877.
Allard RW. 1960. Principles of plant breeding. New York (US): J Wiley & Sons 485 hal.
Ameriana M. 1995. Pengaruh petunjuk kualitas terhadap persepsi konsumen mengenai kualitas tomat. Bul Penel Hort. 27(4) : 8-14.
Amstrong RJ, Tompson AE. 1967. A dialel analysis of tomato fruit cracking. Proc Am Soc Hort Sci. 91:505-517.
Arias R, Lee T C, Logendra L, Janes H. 2000. Correlation of lycopene measured by HPLC with the L*, a*, b* colour readings of a hydroponic tomato and the relationship of maturity with colour and lycopene content. Food Chem. 48:1697–1702.
Awan PN. 1983. Evaluasi karakter hortikultur 17 nomor seleksi tahan layu bakteri. Jurusan Agronomi. [karya ilmiah]. Faperta-IPB.
Bahar H, Zen S. 1993. Parameter genetik pertumbuhan tanaman, hasil dan komponen hasil jagung. Zuriat 4:4-7.
Bollard EG 1970. The physiology and nutrition of developing fruit, p. 387-425. In A.C. Hulme (Ed.) The Biochemistry of Fruit and their Products. Volume 1. London (UK): Academic Press.
[BPS] Badan Pusat Statistik. 2012. Produksi sayuran di Indonesia [Internet]. [diunduh 2013 Januari 10]. Tersedia pada :http://www.bps.go.id/
Byers RE, Carbaugh DH, Presley CN. 1990. ‘Stayman’ fruit cracking as affected by surfactans, plant growth regulator, and other chemical. J Amer Soc Hort Sci 115:405-411.
Cicedo A, Peralta I. 2013. Basic information about tomatoes and the tomato group. Di dalam : Liedl BE, Labate JA, Stommel JR, Slade A, Kole C. Genetik Genomics and Breeding of Tomato. New York (US) : CRC Press. Crowder L. 1993. Genetika Tumbuhan. Edisi keempat. Yogyakarta (ID): Gajah
Mada University Press.
Dickinson DB, McCollum JP. 1964. The effect of calcium on cracking in tomato. Proc Am Soc Hort Sci. 84:485-490.
Dorais M, Demers DA, Papadopoulos AP, Ieperen WV. 2004. Greenhouse tomato fruit cuticule cracking. Horticultural Reviews: 30. Editor: Jules Janick. Canada (CA): John Wiley and Sons, Inc.
Firon N, Shaked R, Peet MM, Pharr DM, Zamski K, Rosenfeld L, Althan E, Pressman. 2006. Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions. Sci Hortic. 109:212-217.
Gould WA. 1974. Tomato Production, Processing and Quality Evaluation. Amerika (US): The Avi Publ. Co.
Hernandes TP, Nassar SH. 1970. Breeding tomato for radial fruit crack resistance and other character. J Am Soc Hort Sci. 95(2):223-226.
Hudson, IW. 1956. The Inheritance of resistance to fruit cracking in the tomato (Lycopersicon esculentum L). [tesis]. Oregon (US): Oregon States College. [IPGRI] International Plant Genetik Resources Institute. 1996. Descriptor for
Tomatos (Lycopersicon spp.). Italia (IT); IPGRI, AVRDC, CATIE.
Jones JB, Wolf JB, Mills HA. 1991. Plant Analysis Handbook. Athen (US): Micro-macro Publ. Inc.
Kamimura S. Yoshikawa S. Ito H. Ito K. 1972. Studies on fruit cracking in tomatoes. Bul. Hort. Res. Sta C. 7:73-138.
Knott JE. 1962. Handbook for Vegetable Growers. New York (US): John Wiley & Sons, Inc 245 p.
Lai G. 1993. Seed production techniques of tomato and brinjal in the tropics and subtropics. Di dalam Breeding of Solanaceous and cole crops. Taiwan (TW): Asian vegetable Research and Development Center.
Lang A. During H. 1990. Grape berry splitting some mechanical properties of skin. Vitis 29:61-70.
Liebisch F, Max JFJ, Heine G, Horst WJ. 2009. Blossom-end rot and fruit cracking of tomato grown in net-covered greenhouses in Central Thailand can partly be corrected by calcium and boron sprays. J Plant Nutrition and Soil Science. 172(1):140-150.
Link H. 1974. Ca-uptake and translocation by plants with special regard to apple trees. Act Hort 45:53-64.
Lopez CAF, Gomez PA 2004. Comparison of colour indexes for tomato ripening. Horticultura Brasileira 22(3), 534–37.
Maboko MM. 2006. Growth, yield and quality of tomatoes (Lycopersicum esculentum Mill) and lettuce (Lactuta sativa L.) as affected by gel-polymer soil amandment and irigation management. [disertasi]. Pretoria (AF): Faculty of Natural and Agriculture Sciences. University of Pretoria.
Mangoendidjojo W. 2003. Dasar-dasar Pemuliaan Tanaman. Yogyakarta (ID): Kanisius.
Marschner H. 1986. Mineral Nutrition of Higher Plants. London (GB). Academic Press. 674 p.
Mather K, Jinks JL. 1977. Introduction to biometrical genetik. New York (US): Cornell Univ Press.
Moore TC. 1979. Biochemistry and Physiology of Plant Hormones. New York (US): Springer-Verlag
Pashar DP, Lambeth VN. 1960. Inheritance of radial fruit cracking in tomatoes. Proc Am Soc Hort Sci. 76:530-537.
Peet MM, Willits DH, Gardner RG. 1997. Responses of ovule development and postpollen production processes in male-sterile tomatoes to chronic, sub-acute high temperature stress. Journal of Experimental Botany. 48: 101-111. Peet MM. 1992. Fruit cracking in Tomato. Hort Technology 2(2):216-223
Petr FC, Frey KJ. 1966. Genotypic correlation, dominance, and heritability of quantitative characters in oat. Crop Sci. 6:259-262.
Poespodarsono S. 1988. Dasar-dasar Ilmu Pemuliaan Tanaman. Pusat Antar Universitas. Bogor (ID): Institut Pertanian Bogor.
[PPVT] Pusat Perlindungan Varietas Tanaman. 2007. Panduan Pengujian Individual Kebaruan, Keunikan, Keseragaman dan Kestabilan Tomat. Jakarta (ID): Departemen Pertanian Repulik Indonesia.
Purnamaningsih R. 2008. Perakitan Tanaman Tomat Partenokarpi untuk Meningkatkan Produksi Tomat Di Dataran Rendah. Warta Biogen. 4 (2):11-12.
Purwati E. 2007. Varietas Unggul Harapan Tomat Hibrida (F1) dari Balitsa. IPTEK Hortikultura. 3: 34-40.
Purwati E. 2008. Hubungan antara karakteristik fenotipik buah tomat dengan jumlah biji. J Agrivigor. 7(3) : 222-229.
Purwati, E. 2009. Daya hasil tomat hibrida (F1) di dataran medium. J Hort. 19(2) : 125-130.
Redmond WJ. 1975. Transport of calcium into apple tress and its penetration into the fruit. Comm Soil Sci and Plant Anal. 6:261-272.
Roy D. 2000. Plant Breeding, Analysis and Exploitation of Variation., New Delhi (IN): Narosa Publishing House
Sastrosumarjo S. 1987. Pola pewarisan karakter resistesnsi terigu (Triticum aestivum L) terhadap kudis malai (Gibberella zeae (Schw.) Petch). [Disertasi]. Bagor [ID]: Institut Pertanian Bogor.
Sato S, Peet MM, Gardner RG. 2001. Formation of partenocarpic fruit, undeveloped flowers and aborted flowers in tomato under moderately elevated temperatures. Sci Hort. 90:243-254.
Sharpio SS, Wilk MB. 1965. An analysis of variance test for normality (completed sample). Biometrika 52:591-611.
Singh RK, Chaudhary BD. 1979. Biometrical methods in quantitative genetik analysis. New Delhi (IN): Kalyani.
Soedomo. 2012. Uji daya hasil lanjutan tomat hibrida di dataran tinggi Jawa Timur. J Hort. 22(1) : 8-13.
Sunarjono H. 2007. Bertanam 30 Jenis Sayur. Jakarta (ID): Penebar Swadaya. Susila AD. 1995. Studi pecah buah (fruit cracking) dalam rangka peningkatan
kualitas buah tomat (Lycopersicon esculentum Mill. L). [Tesis]. Institut Pertanian Bogor (ID): Bogor.
Thompson HC. Kelly WC. 1957. Vegetable Crops, New York (US): Mc Graw Hill Book
Tigchelaar EC 1986. Tomato Breeding, p. 135-171. In Bassets MJ. (Ed.) Breeding Vegetable Crops. Westport, Connecticut (US): AVI Publ. Co. Inc.
[UPOV] International Union For the Protection of New Varieties of Plants. 2011. Tomato. Geneva (CH): UPOV.
Villareal RL. 1980. Tomatoes in the tropics. Westview press boilder. Colorado (US).
Wareing PF, Phillips IDJ. 1981. Growth & Differentiation In Plants (3rd Edition)., Oxford (GB): Pergamon Press.
Wijayani A, Widodo W. 2005. Usaha meningkatkan kualitas beberapa varietas tomat dengan sistem budidaya hidroponik increasing of tomatoes quality in hydroponic culture. Ilmu Pertanian 12(1): 77 - 83
Wright S. 1968. Evolution and genetik of population. Vol 1. Genetik and biometrical foundation. Chicago and London (GB): Univ Chicago Pr. Yamamoto T, Satoh H, Watanabe S. 1992. The effect of Calcium an Naphtalene
Acetic Acid Sprays on Cracking Indeks an Natural Rain Cracking in Sweet Cherry fruits. J Japan Soc Hort Sci. 61 (3):507-511.
Yan W, Fregeau-Reid J. 2008. Breeding line selection based on multiple traits. Crops Sci. 48:417-427.
3
KARAKTERISASI DAN SELEKSI GENOTIPE TOMAT
(
Solanum lycopersicum
L.) BERDASARKAN KARAKTER
KOMPONEN PRODUKSI
Abstrak
Pergeseran budidaya tomat dari dataran tinggi ke dataran rendah menyebabkan penurunan hasil produksi tomat yang sangat signifikan terutama pada varietas tomat yang tidak mampu beradaptasi pada dataran rendah. Seleksi genotipe berdasarkan kriteria seleksi yang tepat adalah metode yang paling efektif untuk mendapatkan genotipe tomat dengan produksi yang tinggi di dataran rendah. Tujuan dari penelitian ini adalah untuk mengidentifikasi genotipe tomat yang memiliki komponen produksi tinggi pada dataran rendah, menentukan kriteria seleksi komponen produksi yang tepat di dataran rendah berdasarkan nilai heritabilitas tinggi, korelasi fenotipe dan genotipe yang tinggi terhadap komponen produksi, dan keeratan hubungan karakter dengan komponen produksi melalui analisis lintas. Penelitian ini menggunakan rancangan acak kelompok dengan tiga ulangan. Hasil analisis gerombol diperoleh kekerabatan 30 genotipe tomat yang dikelompokkan menjadi empat kelompok: kelompok I (IPBT21 dan IPBT73), kelompok II (IPBT4, IPBT6, IPBT13, IPBT64, IPBT74, IPBT80, IPBT85, dan IPBT86), kelompok III (IPBT1, IPBT8, IPBT26, IPBT43, IPBT57, IPBT58, IPBT59, IPBT60, IPBT78, IPBT82, IPBT dan IPBT84), dan kelompok IV (IPBT3, IPBT23, IPBT30, IPBT33, IPBT34, IPBT53, IPBT56 dan IPBT63). Berdasarkan ukuran buah, jumlah buah per tanaman dan bobot buah per tanaman, genotipe yang direkomendasikan sebagai tetua untuk pewarisan komponen produksi adalag genotipe IPBT1, IPBT3, IPBT26, IPBT33, IPBT73, IPBT60, IPBT73, dan IPBT78. Kriteria seleksi yang dipilih berdasarkan pada nilai heritabilitas tinggi, korelasi yang signifikan dalam fenotipe dan genotipe dan memiliki pengaruh langsung yang tinggi terhadap bobot buah per tanaman adalah karakter jumlah buah per tanaman dan berat per buah
Abstract
Shifting cultivation of tomatoes from the uplands to the lowlands causing a decrease in yield production of tomatoes for the varieties grown less does not comply with the required environmental conditions. Selection of genotypes based on appropriate selection criteria is the most effective method to obtain tomato genotypes with high production in the lowlands. The objective of this research was to identify tomato genotypes and selection criteria to determine the character of the high yield component in the lowland base on heritability, phenotype and genotype correlations, and path analysis. A randomized complete block design was used with three replications. The results of the cluster analysis obtained pattern of kinship 30 genotypes of tomato were clustered into four groups: group I (IPBT21 and IPBT73), Group II (IPBT4, IPBT6, IPBT13, IPBT64, IPBT7