OPTIMALISASI RANSUM KOMPLIT BERBASIS JERAMI DAN DEDAK PADI
DENGAN SUPLEMEN KAYA NUTRIEN DITINJAU DARI
FERMENTABILITAS DAN KECERNAAN
in vitro
REGINA FIDELIA
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Optimalisasi Ransum Komplit Berbasis Jerami dan Dedak Padi dengan Suplemen Kaya Nutrien Ditinjau dari Fermentabilitas dan Kecernaan in vitro adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
REGINA FIDELIA. Optimalisasi Ransum Komplit Berbasis Jerami dan Dedak Padi dengan Suplemen Kaya Nutrien Ditinjau dari Fermentabilitas dan Kecernaan in vitro. Dibimbing oleh ANITA SARDIANA TJAKRADIDJAJA dan SURYAHADI.
Tujuan penelitian ini adalah untuk mempelajari optimalisasi ransum komplit berbasis jerami padi yang diberi suplemen kaya nutrien (SKN) ditinjau dari fermentabilitas dan kecernaan in vitro. Percobaan fermentabilitas menggunakan rancangan acak kelompok berpola faktorial 4×3 dengan empat ulangan. Faktor P adalah ransum komplit jerami padi dan SKN (60% : 40%): P1 = jerami padi + SKN A1 (dedak padi, tepung ikan, tepung daun singkong, molases, tepung daun lamtoro,tepung daun turidan mineral mix), P2 = jerami padi + SKN A2 (dedak padi, tepung ikan, tepung daun singkong, tepung daun gamal, molases dan mineral mix), P3 = jerami padi + SKN A3 (SKN A2 + ampas teh), dan P4 = jerami padi + SKN A4 (SKN A3 + tepung daun kembang sepatu). Faktor B adalah waktu inkubasi: 1, 3, dan 5 jam. Percobaan kecernaan menggunakan rancangan acak kelompok dengan tiga perlakuan pakan (P1, P2, P3, P4) dan empat ulangan. Hasil penelitian menunjukkan penggunaan SKN yang berbeda tidak mempengaruhi semua peubah yang dianalisis, namun populasi protozoa, konsentrasi VFA, dan DBK dipengaruhi oleh waktu inkubasi.
Kata kunci: degradabilitas, dedak padi, fermentabilitas, jerami padi, suplement kaya nutrien (SKN)
ABSTRACT
REGINA FIDELIA.Optimilization of rice straw and bran based diet with nutrient rich supplement on in vitro fermentability and digestibility. Supervised by ANITA SARDIANA TJAKRADIDJAJA and SURYAHADI.
The aim of this experiment was to study optimilization of rice and bran straw based diet with nutrient rich supplement (NRS) on in vitro fermentability and digestibility. The fermentability experiment used factorial randomized block design 4×3 with four replicates. The ration treatments (60% rice straw : 40% nutrient rich supplement (NRS)) were factor P: P1 = Rice straw + NRS A1 (rice bran, fish meal, cassava leaf meal, Leucaena leaf meal, Sesbania leaf meal molases and mineral mix), P2 = Rice straw + NRS A2 (rice bran, fish meal, cassava leaf meal, Gliricidia sepium leaf meal, molases and mineral mix), P3 = Rice straw + NRS A3 (concentrate A2 + tea waste), and P4 = Rice straw + NRS A4 (concentrate A3 + Hibiscus rosasinensis leaf meal). Factor B was incubation times: 1, 3, and 5 hours. The digestibility experiment used randomized block design with four ration treatments (P1, P2, P3, P4) and four replications. The result showed that supplementation did not influence all variables measured, but protozoal population, VFA concentration and dry matter degradability were affected by incubation time.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada
Departemen Ilmu Nutrisi dan Teknologi Pakan
OPTIMALISASI RANSUM KOMPLIT BERBASIS JERAMI DAN DEDAK
PADI DENGAN SUPLEMEN KAYA NUTRIEN DITINJAU DARI
FERMENTABILITAS DAN KECERNAAN
in vitro
REGINA FIDELIA
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
Judul Skripsi : Optimalisasi Ransum Komplit Jerami Padi dengan Konsentrat Berbasis Dedak Padi dan Tepung Daun Ditinjau dari
Fermentabilitas dan Kecernaan in vitro Nama : Regina Fidelia
NIM : D24090110
Disetujui oleh
Ir Anita S Tjakradidjaja, MRurSc Pembimbing I
Dr Ir Suryahadi, DEA Pembimbing II
Diketahui oleh
Prof Dr Ir Panca Dewi MHKS, MS Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Januari 2013 ini ialah suplemen kaya nutrien, dengan judul Optimalisasi Ransum Komplit Berbasis Jerami dan Dedak Padi dengan Suplemen Kaya Nutrien Ditinjau dari Fermentabilitas dan Kecernaan in vitro.
Jerami padi merupakan sumber energi yang baik bagi ternak ruminansia. Namun kecernaan yang rendah menjadi permasalahan dalam penggunaan jerami padi. Suplementasi protein dan energi dalam ransum berbasis jerami padi dapat melengkapi kebutuhan nutrien pakan yang dibutuhkan oleh ternak sapi potong, sehingga performa produksi sapi potong meningkat.
Skripsi ini merupakan salah satu syarat untuk kelulusan dan memperoleh gelar Sarjana Peternakan, Institut Pertanian Bogor. Kritik dan saran sangat diharapkan penulis untuk menyempurnakan skripsi ini.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
METODE PENELITIAN 2
Bahan 2
Alat 2
Komposisi Suplemen Kaya Nutrien (SKN) 3
Lokasi dan Waktu Penelitian 4
Prosedur Percobaan 4
Pengambilan Cairan Rumen 4
Pembuatan Larutan McDougall 4
Pencernaan Fermentatif 4
Pengukuran NH3 5
Pengukuran VFA 5
Perhitungan Populasi Bakteri Total 6
Perhitungan Populasi Protozoa Total 6
Analisis Sintesis Protein Mikroba 6
Pengukuran Degradabilitas Bahan Kering dan Bahan Organik 7 Pengukuran Koefisien Cerna Bahan Kering dan Bahan Organik 7
Peubah yang Diamati 8
Analisis Data 8
HASIL DAN PEMBAHASAN 9
Kandungan Nutrien Ransum 9
Konsentrasi NH3 (Amonia) 10
Konsentrasi VFA 12
Populasi Bakteri Total 14
Populasi Protozoa Total 15
Sintesis Protein Mikroba 16
Degradabilitas Bahan Kering (DBK) dan Bahan Organik (DBO) 18
Koefisien Cerna Bahan Kering dan Bahan Organik 19
SIMPULAN DAN SARAN 21
Simpulan 21
Saran 21
DAFTAR PUSTAKA 21
LAMPIRAN 25
DAFTAR TABEL
1 Komposisi suplemen kaya nutrien 3
2 Kandungan nutrien suplemen kaya nutrien berdasarkan perhitungan 3 3 Kandungan nutrien ransum perlakuan berdasarkan perhitungan 10 4 Pengaruh perlakuan terhadap rataan konsentrasi amonia 11 5 Pengaruh perlakuan terhadap rataan konsentrasi VFA total 13 6 Pengaruh perlakuan terhadap rataan populasi bakteri total 15 7 Pengaruh perlakuan terhadap rataan populasi protozoa total 16 8 Pengaruh perlakuan terhadap rataan sintesis protein mikroba 17 9 Pengaruh perlakuan terhadap rataan degradabilitas bahan kering dan
bahan organik 18
10 Pengaruh perlakuan terhadap rataan koefisien cerna bahan kering
dan koefisien cerna bahan organik 20
DAFTAR LAMPIRAN
1
Sidik ragam pengaruh perlakuan terhadap konsentrasi amonia 25 2 Sidik ragam pengaruh perlakuan terhadap konsentrasi VFA total 25 3 Uji ortogonal polinomial pengaruh waktu inkubasi terhadapkonsentrasi VFA 25
4 Sidik ragam pengaruh perlakuan terhadap populasi bakteri total 26 5 Sidik ragam pengaruh perlakuan terhadap populasi protozoa total 26 6 Uji ortogonal polinomial pengaruh waktu inkubasi terhadap protozoa
total 26
7 Sidik ragam pengaruh perlakuan terhadap sintesis protein mikroba 27 8 Sidik ragam pengaruh perakuan terhadap degradibilitas bahan kering 27 9 Uji ortogonal polinomial pengaruh waktu inkubasi terhadap
degradibilitas bahan kering 27
10 Sidik ragam pengaruh perlakuan terhadap degradibilitas bahan
organik 28
11 Sidik ragam pengaruh perlakuan terhadap koefisien cerna bahan
kering 28
12 Sidik ragam pengaruh perlakuan terhadap koefisien cerna bahan
PENDAHULUAN
Pakan adalah semua bahan yang biasa diberikan dan bermanfaaat bagi ternak serta tidak menimbulkan pengaruh negatif terhadap tubuh ternak. Pakan yang diberikan harus berkualitas tinggi yaitu mengandung zat-zat yang diperlukan oleh tubuh ternak seperti air, karbohidrat, lemak, protein dan mineral. Permasalahan yang sering terjadi adalah ketersediaan pakan yang tidak selalu kontinyu, di sisi lain limbah pertanian seperti jerami padi merupakan salah satu produk samping pertanian yang tersedia cukup melimpah. Jerami padi yang dihasilkan dapat mencapai 5 ton ha-1 setiap kali panen (Haryanto et al. 2002). Kandungan bahan kering (BK) jerami padi segar sekitar 40% sampai 45%, sehingga potensi produksi BK jerami padi adalah 2.0 sampai 2.25 ton ha-1. Adapun kebutuhan ternak sapi adalah sekitar 6 sampai 7 kg BK jerami padi ekor-1 hari-1 untuk sapi dengan bobot hidup 250 kg. Dengan demikian produksi jerami padi (per satuan BK) dapat digunakan sebagai pakan untuk 375 ekor sapi. Namun demikian penggunaan jerami padi secara langsung sebagai pakan tunggal tidak dapat memenuhi pasokan nutrien yang dibutuhkan ternak. Hal tersebut disebabkan kandungan protein rendah, serat kasar tinggi, kecernaan rendah dan dinding selnya tersusun oleh selulosa, hemiselulosa, lignin dan silika (Budiman 2007). Kandungan serat kasar yang tinggi dapat menghambat mikroba rumen dalam mencerna pakan. Oleh karena itu, penelitian dan pengembangan terus dilakukan untuk meningkatkan kualitas jerami padi agar dapat dimanfaatkan sebagai bahan pakan secara optimal, terutama untuk ternak ruminansia.
Teknologi suplementasi merupakan satu alternatif yang diharapkan dapat meningkatkan kemampuan mikroba, terutama bakteri rumen, dalam meningkatkan kapasitas ternak untuk mencerna jerami padi. Suplemen yang digunakan dalam penelitian ini adalah suplemen dengan campuran bahan pakan yang mudah didapatkan seperti tepung ikan, molases, ampas teh, daun singkong, daun gamal, dan daun kembang sepatu. Selain itu, pembuatan ransum komplit jerami yang berbasis dedak padi ditambah suplemen kaya nutrien (SKN) berupa tepung daun dalam bentuk konsentrat diharapkan dapat mengatasi masalah ketersediaan pakan yang tidak selalu kontinyu dan memperbaiki nilai nutrien ransum jerami padi.
2
gamal lebih mudah didegradasi oleh mikroba rumen (Smith dan Van Houtert 2000). Dua bahan yang lainnya, yaitu ampas teh dan daun singkong. Ampas teh mengandung tanin yang dapat melindungi protein sehingga dapat meningkatkan suplai protein bypass (Lubis 1963). Walaupun daun singkong mengandung HCN, tetapi efek negatif HCN diharapkan dapat diturunkan dengan proses pengeringan yang dilakukan di penelitian ini. Daun singkong tidak mengandung tanin sehingga tidak terjadi perlindungan protein dan protein daun singkong akan lebih mudah didegradasi di dalam rumen (Sigit 1983). Daun kembang sepatu juga digunakan dalam penelitian ini sebagai sumber saponin yang merupakan upaya defaunasi untuk meningkatkan suplai protein bakteri (Putra 2006).
Penggunaan SKN yang dicampur dedak padi dalam bentuk konsentrat diharapkan dapat membantu memperbaiki penampilan sapi potong, karena pakan jerami dan dedak padi yang diberikan oleh peternak tidak mencukupi kebutuhan sapi potong. Penambahan SKN pada pakan sapi potong sangat baik dan dapat mencukupi kebutuhan sapi potong. Hal ini sudah terlihat dari keberhasilan penelitian sebelumnya (Suryahadi et al. 2012), namun formulasi baru dan penggunaan SKN yang ditingkatkan jumlahnya dan dicampurkan dengan dedak padi yang lalu diintegrasikan dengan jerami padi sebagai ransum komplit sangat diperlukan untuk memperkuat keberhasilan penelitian ini.
Penelitian ini bertujuan untuk mempelajari optimalisasi ransum komplit jerami padi dengan konsentrat berbasis dedak padi dan tepung daun ditinjau dari fermentabilitas dan kecernaan in vitro.
METODE PENELITIAN
Bahan
Bahan yang digunakan adalah cairan rumen segar sapi potong yang berasal dari rumah potong hewan (RPH) di Bubulak, jerami padi, dedak padi, tepung daun singkong, tepung ikan, molases, tepung daun turi, tepung daun lamtoro, tepung daun gamal, ampas teh, tepung daun kembang sepatu, mineral mix, plastik kemasan, label, larutan McDougall dengan pH 6.5 sampai 6.9, larutan pepsin HCl 0.2%, aquadest, larutan HgCl2 jenuh, larutan Na2CO3 jenuh, larutan H2SO4 0.005
N, asam borat berindikator merah metil dan hijau bromo kresol, larutan HCl 0.5 N, larutan H2SO4 15%, larutan NaOH 0.5 N, larutan indikator phenolphtalein
0.1% (PP), larutan garam formalin (formal saline), medium brain heart infusion (BHI), gas CO2, trichloro acetic acid (TCA), dan sulfo salicylic acid (SSA).
Alat
Peralatan yang digunakan adalah seperangkat alat-alat percobaan fermentasi dan kecernaan in vitro (timbangan digital, tabung fermentor, tutup karet berventilasi, shaker waterbath, tabung gas CO2, cawan porselen, oven 105 oC,
3
Komposisi Suplemen Kaya Nutrien (SKN)
Ransum komplit jerami padi dan SKN disusun dengan komposisi jerami padi dan SKN (60%:40%). Ransum P1 = jerami padi dan SKN A1, P2 = jerami padi dan SKN A2, P3 = jerami padi dan SKN A3 dan P4 = P3 + tepung daun kembang sepatu. SKN yang dibuat dalam penelitian ini terdiri dari bahan pakan yang mudah didapatkan di sekitar lokasi peternakan dan dapat digunakan sebagai pakan alternatif. SKN yang digunakan (Tabel 1) adalah :
SKN A1 = dedak padi, tepung ikan, tepung daun (singkong, turi dan lamtoro), molases dan mineral mix.
SKN A2 = dedak padi, tepung ikan, tepung daun (singkong dan daun gamal), molases dan mineral mix.
SKN A3 = SKN A2 + ampas teh.
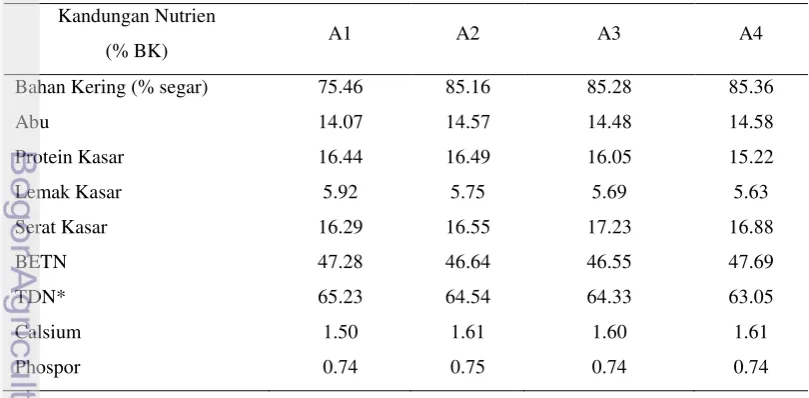
SKN A4 = SKN A3 + tepung daun kembang sepatu. Tabel 1 Komposisi suplemen kaya nutrien
Bahan pakan A1 A2 A3 A4
% BK
Dedak padi 55 55 55 55
Tepung ikan 10 10 10 10
Tepung daun singkong 15 15 15 15
Tepung daun turi 9 - - -
Tepung daun lamtoro 5 - - -
Tepung daun gamal - 14 9 4
Tepung daun kembang sepatu - - - 5
Ampas teh - - 5 5
Molases 5 5 5 5
Mineral mix (campuran mineral) 1 1 1 1
Tabel 2 Kandungan nutrien suplemen kaya nutrien berdasarkan perhitungan Kandungan Nutrien
(% BK) A1 A2 A3 A4
Bahan Kering (% segar) 75.46 85.16 85.28 85.36
Abu 14.07 14.57 14.48 14.58
Protein Kasar 16.44 16.49 16.05 15.22
Lemak Kasar 5.92 5.75 5.69 5.63
Serat Kasar 16.29 16.55 17.23 16.88
BETN 47.28 46.64 46.55 47.69
TDN* 65.23 64.54 64.33 63.05
Calsium 1.50 1.61 1.60 1.61
Phospor 0.74 0.75 0.74 0.74
4
Adapun rumus TDN yang digunakan (Sutardi 2001 dalam Irawan 2002) adalah : 1. Untuk pakan dengan SK<18 % dan PK<20 %
TDN = 2.79 + 1.17 PK + 1.74 Lemak – 0.295 SK + 0.810 Beta-N 2. Untuk pakan dengan SK<18 % dan PK>20 %
TDN = 25.6 + 0.530 PK + 1.70 Lemak – 0.474 SK + 0.732 Beta-N 3. Untuk pakan dengan SK>18 % dan PK<20 %
TDN = 70.6 + 0.259 PK + 1.01 Lemak – 0.760 SK + 0.0991 Beta-N 4. Untuk pakan dengan SK>18 % dan PK>20 %
TDN = 3.17 + 0.640 PK + 2.08 Lemak – 0.0675 SK + 0.940 Beta-N
Lokasi dan Waktu Penelitian
Penelitian dimulai dari bulan Januari 2013 hingga Juli 2013. Penelitian dilaksanakan di Laboratorium Biokimia, Mikrobiologi, dan di Laboratorium Nutrisi Ternak Perah, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor, Bogor.
Prosedur Percobaan
Pengambilan Cairan Rumen
Air hangat (suhu 39 °C) diisikan ke dalam termos dan dibawa ke rumah potong hewan (RPH) di Bubulak. Isi rumen diambil dari sapi yang baru dipotong, kemudian disaring menggunakan kain penyaring. Cairan rumen yang sudah disaring lalu dimasukkan ke dalam termos setelah air hangat di dalam termos dibuang. Cairan rumen dalam termos tersebut kemudian dibawa ke Laboratorium Biokimia dan Mikrobiologi Nutrisi.
Pembuatan Larutan McDougall
Pembuatan Larutan McDougall (1 liter) dilakukan dengan komposisi berikut : NaHCO3 (9.8 g); Na2HPO4.7H2O (4.6325 g); KCl (0.57 g); NaCl (0.47 g);
MgSO4.7H2O (0.12 g); CaCl2.2H2O (0.04 g). Seluruh bahan hingga homogen
dengan menggunakan alat magnetik stirrer. Selanjutnya campuran dilarutkan kembali sambil dialiri dengan CO2. CaCl2.2H2O ditambahkan paling akhir setelah
bahan lainnya larut sempurna.
Pencernaan Fermentatif
Percobaan in vitro dilakukan dengan menggunakan metode Tilley dan Terry (1963) yang dimodifikasi oleh Sutardi (1979). Dalam metode ini digunakan tabung fermentor (50 ml) yang diisi dengan 1 g sampel, 12 ml larutan buffer McDougall dan 8 ml cairan rumen segar. Tabung lalu dikocok dengan dialiri CO2
5 sama dengan kondisi di dalam rumen dan diinkubasi selama 1; 3; dan 5 jam. Proses fermentasi dihentikan dengan meneteskan larutan HgCl2 jenuh sebanyak 2
tetes. Sebelum fermentasi dihentikan, sampel diambil untuk analisis bakteri total dan protozoa total. Setelah itu, tabung fermentor disentrifuse pada kecepatan 3000 rpm selama 15 menit. Supernatan diambil untuk analisis konsentrasi NH3 dan
VFA, sedangkan residu diambil untuk analisis DBK dan DBO.
Pengukuran NH3
Konsentrasi NH3 diukur dengan menggunakan metode Mikrodifusi Conway
(General Laboratory Procedure, Department of Dairy Science University of Wisconsin 1969). Vaselin digunakan untuk mengolesi bagian bibir dan tutup cawan Conway. Supernatan (1 ml) diletakkan di salah satu bagian cawan Conway, dan disebelah bagian ini ditempatkan larutan Na2CO3 jenuh (1 ml). Kedua larutan
tersebut diupayakan untuk tidak bercampur sebelum penempatan larutan asam borat berindikator merah metil dan hijau bromo kresol (1 ml) di bagian tengah cawan. Setelah itu, cawan ditutup rapat hingga kedap udara, dan digoyang dengan hati-hati agar supernatan dan larutan Na2CO3 jenuh dapat bercampur. Amonia
yang terdesak dari supernatan akan ditangkap oleh asam borat, proses ini berlangsung dalam suhu ruang. Setelah 24 jam, tutup cawan dibuka, asam borat berindikator dititrasi dengan larutan H2SO4 0.005N sampai terjadi perubahan
warna dari biru menjadi merah. Konsentrasi NH3 dihitung berdasarkan rumus
berikut:
Pengukuran VFA
Konsentrasi VFA diukur dengan menggunakan Teknik Destilasi Uap (General Laboratory Procedure, Department of Dairy Science University of Wisconsin 1969). Sebanyak 5 ml supernatan yang sama dengan analisa NH3
dimasukkan ke dalam tabung destilasi, lalu ditambahkan larutan H2SO4 15% (1
ml). Labu kemudian segera ditutup dengan tutup karet yang mempunyai lubang yang dapat menghubungkan dengan labu pendingin. Segera setelah penambahan larutan H2SO4 15% ke dalam supernatan, tabung destilasi dimasukkan ke dalam
labu penyulingan yang berisi air mendidih (dipanaskan terus selama destilasi). Uap air panas akan mendesak VFA yang akan terkondensasi di dalam pendingin. Cairan yang terbentuk lalu ditampung dalam labu Erlenmeyer yang berisi 5 ml NaOH 0.5 N sampai mencapai 250 ml. Indikator PP ditambahkan sebanyak 2 tetes ke dalam destilat yang lalu dititrasi dengan HCl 0.5 N sampai warna titrat berubah dari merah jambu menjadi tidak berwarna. Rumus berikut digunakan untuk menghitung konsentrasi VFA.
Keterangan :
6
Perhitungan Populasi Bakteri Total
Perhitungan populasi bakteri total dilakukan menggunakan metode Ogimoto and Imai (1981). Populasi bakteri dihitung dengan metode pencacahan koloni bakteri hidup. Prinsip perhitungannya adalah cairan rumen diencerkan secara serial, lalu disimpan dalam tabung Hungate. Sampel (cairan rumen yang telah mengalami perlakuan inkubasi) dipipet 0.05 ml dan dimasukkan ke dalam media pengencer. Pengenceran dilakukan sebagai berikut: 0.05 ml kultur bakteri dimasukkan ke dalam 4.95 ml media pengencer. Selanjutnya dari media pengencer diambil kembali 0.05 ml, lalu dimasukkan ke dalam 4.95 ml media pengencer berikutnya, sehingga mendapat pengenceran 10-2, 10-4, 10-6, dan 10-8. Dari masing-masing seri tabung pengenceran diambil sebanyak 0.1 ml, kemudian dimasukkan ke dalam media agar dan diputar sambil dialiri air pada roller, supaya medium dapat menjadi padat secara merata. Selanjutnya bakteri diinkubasi selama 24 jam. Perhitungan populasi bakteri dilakukan dengan rumus:
Populasi bakteri (CFU ml-1) = n x 10x 0.05 x 0.1 Keterangan :
n = jumlah koloni yang terdapat pada tabung seri pengenceran ke-x
Perhitungan Populasi Protozoa
Perhitungan populasi protozoa dilakukan dengan menggunakan counting chamber dari sampel yang diperoleh. Sebelumnya sampel yang didapat dari proses fermentasi dicampur dengan larutan garam formalin (TBFS) dengan rasio 1 : 1. Larutan garam formalin dibuat dari campuran larutan formalin 4% ditambah larutan garam NaCl fisiologis 0.9% dalam 100 ml larutan (Ogimoto dan Imai 1981). Sampel yang sudah dicampur tersebut lalu dihomogenkan. Sampel diambil dan diteteskan pada counting chamber sebanyak 2 tetes dan ditutup dengan gelas penutup (cover glass) sampai rata. Counting chamber yang digunakan mempunyai ketebalan 0.1 mm, dengan luas kotak terkecil 0.0625 mm dan terdapat 16 kotak. Pada perhitungan populasi protozoa, kotak yang dibaca sebanyak 5 kotak. Populasi protozoa dilakukan dengan mikroskop lensa obyektif (perbesaran 40x dan okuler 10x). Perhitungan populasi protozoa adalah:
Protozoa ml cairan rumen-1 = 1000 x FP x C 0.1 x 0.0625 x 16 x 5 Keterangan :
FP = faktor pengencer
C = jumlah protozoa terhitung dalam counting chamber
Analisis Sintesis Protein Mikroba
7 dan endapan ditambah dengan aquades (3 ml), kemudian ditambahkan campuran TCA-SSA (6 ml). Campuran ini dihomogenkan lagi dengan vortex (2 menit), lalu disentrifuge (kecepatan 3000 rpm; 15 menit). Supernatannya dibuang dan endapannya dianalisis dengan metode Kjehldal mikro.
Pengukuran Degradabilitas Bahan Kering dan Bahan Organik
Metode Tilley dan Terry (1963) yang dimodifikasi oleh Sutardi (1979) digunakan untuk mengukur tingkat degradabilitas BK dan BO (DBK dan DBO). Untuk mengukur peubah ini, digunakan residu yang diperoleh dari sampel yang sama dari proses fermentasi. Residu yang diperoleh lalu dikeringkan di dalam oven 105 °C selama 24 jam untuk mengetahui bobot BK residu. Setelah itu, sampel diabukan di dalam tanur (600 °C selama 6 jam) untuk mendapatkan bobot abu, dan bobot BO yang diperoleh dengan mengurangi bobot BK dengan bobot abu. Sampel blanko diperlakukan dengan cara yang sama dengan sampel perlakuan pakan, hanya di dalam sampel blanko tidak diisi dengan sampel pakan. Degradabilitas bahan kering (DBK) dan bahan organik (DBO) dapat dihitung dengan rumus:
Pengukuran Koefisien Cerna Bahan Kering dan Bahan Organik
Pengukuran kecernaan BK dan BO (KCBK dan KCBO) dilaksanakan melalui dua tahap, yaitu tahap fermentasi dan tahap kecernaan berdasarkan metoda Tilley dan Terry (1963) yang dimodifikasi oleh Sutardi (1979). Komposisi tabung fermentor sama dengan untuk pengukuran fermentasi, hanya proses fermentasi dilakukan selama 24 jam sebagai tahap pertama. Setelah 24 jam proses fermentasi dihentikan dengan menambah larutan HgCl2 jenuh (2 tetes).
8
Peubah yang Diamati
Peubah yang diamati yaitu konsentrasi NH3, konsentrasi VFA total,
degradabilitas bahan kering (DBK), dan degradabilitas bahan organik (DBO), koefisien cerna bahan kering (KCBK), koefisien cerna bahan organik (KCBO), populasi protozoa, populasi bakteri total, dan sintesis protein mikroba.
Analisis Data
Data dianalisis dengan menggunakan sidik ragam (ANOVA) dan untuk mengetahui perbedaan antara perlakuan diuji dengan ortogonal kontras (Steel dan Torrie, 1993).
Rancangan percobaan yang digunakan untuk percobaan fermentabilitas adalah rancangan acak kelompok (RAK) berpola faktorial 4×3. Faktor P adalah ransum berbasis jerami dan dedak padi yang disuplementasi dengan SKN. Faktor B adalah waktu inkubasi fermentasi in vitro 1; 3; 5 jam. Cairan rumen dari empat ekor sapi potong digunakan sebagai ulangan atau kelompok. Adapun perlakuan ransum yang diterapkan adalah :
P1 = jerami padi + SKN A1 (dedak padi, tepung ikan, tepung daun singkong, tepung daun lamtoro,tepung daun turidan mineral mix) P2 = jerami padi + SKN A2 (dedak padi, tepung ikan, tepung daun
singkong, tepung daun gamaldan mineral mix) P3 = jerami padi + SKN A3 (SKN A2 + ampas teh) P4 = jerami padi + SKN A4 (SKN A3 + tepung daun
kembang sepatu)
Model matematika yang digunakan adalah :
Yijk = μ + τi + αj + ßk + αjßk + εijk
Keterangan :
Yijk = nilai pengamatan kelompok ke-i, perlakuan ke-j dan waktu inkubasi ke-k
μ = nilai rataan umum
τi = pengaruh kelompok (cairan rumen) ke-i
αj = pengaruh perlakuan ransum yang diberi suplemen ke-j
ßk = pengaruh perlakuan waktu inkubasi ke-k
αjßk = pengaruh interaksi perlakuan ransum yang diberi suplemen dan waktu
inkubasi
εijk = galat percobaan untuk kelompok ke-i, pengaruh perlakuan ransum yang
diberi suplemen ke-j dan pengaruh perlakuan waktu inkubasi ke-k
Rancangan percobaan yang digunakan untuk percobaan kecernaan adalah rancangan acak kelompok (RAK) dengan 4 perlakuan dan 4 ulangan.
Model matematika dari rancangan adalah : Yij= μ + αi + ßj+ εij
Keterangan :
Yij = nilai pengamatan pada perlakuan ke-i dan ulangan ke-j
9 αi = pengaruh perlakuan ransum yang diberi suplemen ke-i
ßj = pengaruh kelompok ke-j
εij = eror perlakuan ke-i dan ulangan ke-j
HASIL DAN PEMBAHASAN
Kandungan Nutrien Ransum
Penelitian ini menggunakan bahan pakan yang berasal dari limbah pertanian yaitu jerami padi, dedak padi. Selain itu digunakan beberapa bahan lain, yaitu tepung ikan, molasses, tepung daun (gamal, kembang sepatu, singkong, turi, lamtoro), dan ampas teh yang belum dimanfaatkan secara optimal, sehingga penggunaannya dapat menjadi pakan alternatif yang menguntungkan. Ketersediaan yang melimpah pada suatu daerah dapat memudahkan peternak untuk mendapatkan pakan yang sesuai dengan kebutuhan nutrisi ternak. BATAN (2005) menambahkan bahwa suplemen yang tersusun dari kombinasi bahan limbah sumber protein dengan tingkatan jumlah tertentu dapat mendukung pertumbuhan, perkembangan dan kegiatan mikroba secara efisien di dalam rumen. Menurut Yulistiani et al. (2003), berbagai teknologi untuk memperbaiki kualitas nutrien jerami padi telah tersedia, antara lain melalui perlakuan secara fisik, kimiawi, biologis, suplementasi, atau kombinasi di antaranya. Astuti (2006) menyebutkan bahwa suplementasi secara keseluruhan dapat memberikan pengaruh yang baik terhadap peningkatan protein mikroba, daya cerna dan konsumsi pakan sehingga dapat diperoleh keseimbangan yang baik antara asam amino dan energi di dalam zat-zat makanan yang terserap. Dalam percobaan ini, perbaikan penggunaan jerami padi dilakukan dengan upaya penggunaan konsentrat yang berbasis dedak padi dan tepung daun.
10
Komposisi SKN yang dibuat merupakan hasil modifikasi dari SKN yang digunakan oleh Suryahadi et al. (2012). Berdasarkan hasil analisis proksimat, konsentrat memiliki kandungan PK berkisar 15.22 sampai 16.49% dan kandungan TDN berkisar antara 63.05% sampai 65.23% (Tabel 2). Pada penelitian ini telah dilakukan perbaikan terhadap kandungan protein dari konsentrat yang diujikan sebelumnya sebesar 5.26%, dan kandungan energi sebesar 10.32% (Saputra 2011). Dengan demikian, SKN dapat digunakan sebagai suplemen dalam ransum yang akan sedikit meningkatkan kandungan PK dan TDN, dan pemakaian suplemen telah ditingkatkan jumlahnya dan diintegrasikan dengan dedak padi dalam bentuk ransum komplit jerami padi (Tabel 3).
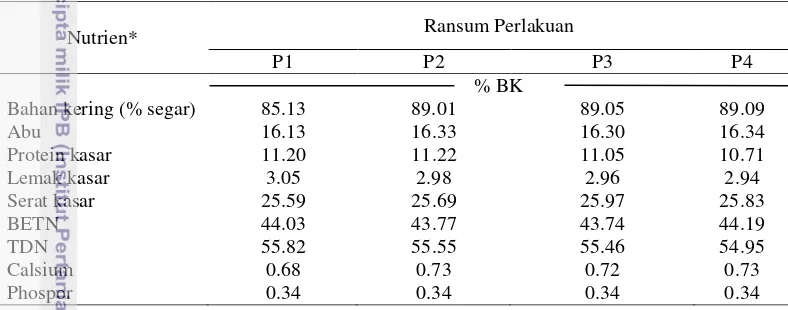
Tabel 3 Kandungan nutrien ransum perlakuan berdasarkan perhitungan
Nutrien* Ransum Perlakuan
P1 P2 P3 P4
% BK
Bahan kering (% segar) 85.13 89.01 89.05 89.09
Abu 16.13 16.33 16.30 16.34
Protein kasar 11.20 11.22 11.05 10.71
Lemak kasar 3.05 2.98 2.96 2.94
Serat kasar 25.59 25.69 25.97 25.83
BETN 44.03 43.77 43.74 44.19
TDN 55.82 55.55 55.46 54.95
Calsium 0.68 0.73 0.72 0.73
Phospor 0.34 0.34 0.34 0.34
*Perhitungan berdasarkan data Sutardi (1980); P1 = jerami padi + SKN A1; P2 = jerami padi + SKN A2; P3 = jerami padi + SKN A3; P4 = jerami padi + SKN A4.
Penggunaan SKN di dalam ransum dasar jerami padi menunjukkan kandungan PK dalam ransum penelitian berkisar 10.71% sampai 11.22% dan kandungan TDN berkisar 54.95% sampai 55.82% (berdasarkan perhitungan). Kandungan PK terendah dimiliki oleh Ransum P4 yang merupakan ransum dengan penambahan SKN A4 (daun gamal, daun kembang sepatu, dan ampas teh) sebesar 10.71%, sedangkan kandungan PK untuk P1, P2, dan P3 berturut-turut adalah 11.20%, 11.22%, dan 11.05%. Perbedaan bahan yang digunakan dalam ransum dapat menghasilkan kandungan PK dan TDN yang berbeda pula.
Konsentrasi NH3 (Amonia)
Konsentrasi (amonia) merupakan hasil fermentasi dan degradasi pakan di dalam rumen. Konsentrasi amonia menunjukkan sifat degradabilitas protein bahan makanan di dalam rumen (Sutardi 1980). Amonia merupakan sumber nitrogen utama dan penting untuk sintesis protein mikroba (Sakinah 2005).
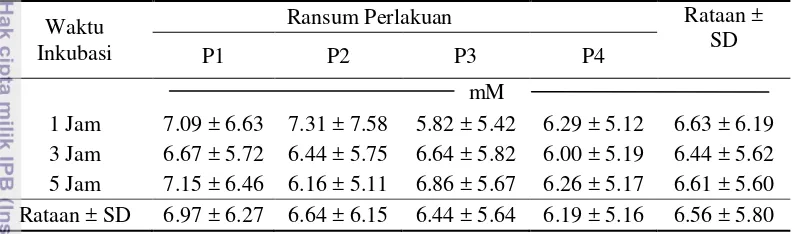
11 amonia untuk pembentukan protein mikroba, yaitu 4 sampai 12 mM (Sutardi 1979) atau 6 sampai 12 mM (McDonald et al. 2002). Konsentrasi amonia percobaan ini lebih tinggi bila dibandingkan dengan hasil penelitian yang dilakukan oleh Saputra (2011) yang menghasilkan konsentrasi amonia sekitar 2.68 mM sampai 3.61 mM. Rataan konsentrasi amonia tertinggi pada ransum P4 (3.61 mM) yaitu ransum komplit dan terendah pada ransum P1 (2.68 mM) yaitu ransum jerami padi tanpa penambahan suplemen kaya nutrien (Saputra 2011). Tabel 4 menunjukkan data konsentrasi amonia.
Tabel 4 Pengaruh perlakuan terhadap rataan konsentrasi amonia
Waktu
Adanya penambahan SKN yang berbeda pada setiap perlakuan tidak menyebabkan perbedaan yang nyata terhadap konsentrasi amonia. P1 menggunakan SKN A1 (daun lamtoro dan daun turi) seperti yang digunakan pada penelitian sebelumnya (Saputra 2011); penggunaan daun turi dan lamtoro diganti dengan daun gamal dalam P2; sebagian daun gamal diganti dengan ampas teh dalam P3; dan dalam P4, penggunaan daun gamal semakin berkurang dan diganti dengan ampas teh dan kembang sepatu. Dengan tidak adanya perbedaan diantara ke empat ransum tersebut, menunjukkan bahwa beberapa bahan yang ditambahkan atau digantikan seperti lamtoro, turi, gamal, ampas teh dan kembang sepatu, dapat digunakan sebagai suplemen yang dapat menunjang ketersediaan amonia. Tidak adanya perbedaan yang nyata dalam konsentrasi amonia diantara perlakuan ransum dapat disebabkan kandungan protein setiap perlakuan yang tidak signifikan yaitu berkisar 10.71% sampai 11.22%. Perbaikan terhadap kandungan protein dari SKN yang diujikan sebelumnya mengakibatkan perbaikan pula pada konsentrasi amonia yaitu sebesar 3.36 mM. Kandungan protein ransum yang tinggi dan proteinnya mudah didegradasi akan menghasilkan konsentrasi NH3 yang relatif tinggi di dalam rumen (McDonald et al. 2002).
12
teh dan daun kembang sepatu ke dalam ransum perlakuan diduga dapat meningkatkan konsentrasi amonia. Penambahan ampas teh pada ransum P3 menghasilkan konsentrasi amonia yang tidak berbeda bila dibandingkan dengan P1, P2 dan P4, hal ini diakibatkan oleh penggunaan ampas teh dalam ransum yang relatif rendah sehingga kadar tanin pun jumlahnya sedikit dan tidak bersifat racun. Menurut (Wahyuni 2008), penambahan ampas teh dapat digunakan untuk pencernaan pasca rumen atau bypass karena tanin yang terkandung dalam ampas teh dapat mengikat protein agar tidak didegradasi terlalu banyak oleh mikroba di dalam rumen sehingga protein mikroba dapat lolos ke usus halus sehingga dapat digunakan untuk pertumbuhan ternak. Penggunaan 100% kembang sepatu dapat meningkatkan konsentrasi amonia dan penggunaan kombinasi ampas teh dan daun kembang sepatu dengan perbandingan 1:1 secara in vitro menunjukkan hasil yang terbaik bagi produksi amonia (Setiani 2002). Kembang sepatu dapat menghasilkan konsentrasi amonia yang cukup baik. Protein kembang sepatu lebih mudah dihidrolisis menjadi asam amino daripada protein ampas teh, hal ini dikarenakan tingkat degradasinya cukup tinggi (Despal 1993), sehingga dapat tercukupi untuk pembentukan protein mikroba.
Waktu inkubasi 1, 3, dan 5 jam tidak mempengaruhi konsentrasi amonia. Hal ini tidak sebanding dengan pernyataan Nuraeni (1993) yang menyatakan bahwa konsentrasi amoniadapat dipengaruhi oleh waktu inkubasi. Sutardi (1980) menyatakan bahwa pada 1 sampai 1.5 jam waktu inkubasi pakan merupakan awal penentu dari kadar NH3 suatu pakan yang berasal dari solubilitas dari pakan itu
sendiri. Hal ini diduga karena kelarutan pakan antar perlakuan relatif sama.
Konsentrasi VFA
Karbohidrat di dalam rumen diubah menjadi volatile fatty acid (VFA) yang merupakan sumber energi utama bagi ternak ruminansia. Produksi gas yang dihasilkan didalam rumen menunjukkan terjadinya proses fermentasi bahan pakan oleh mikroba rumen yaitu menghidrolisa karbohidrat menjadi monosakarida dan disakarida yang kemudian difermentasi lebih lanjut menjadi asam lemak terbang (VFA) dan gas metan dan CO2 (Astuti 2006). McDonald et al. (2002) menyatakan
bahwa proporsi molar dari VFA terdiri dari 65% asetat, 21% propionat dan 14% butirat, proporsi ini sangat dipengaruhi oleh jenis pakan yang dikonsumsi. VFA sangat penting karena merupakan sumber energi yang memenuhi sekitar 50 sampai 70% dari kebutuhan energi ternak ruminansia (Damron 2006).
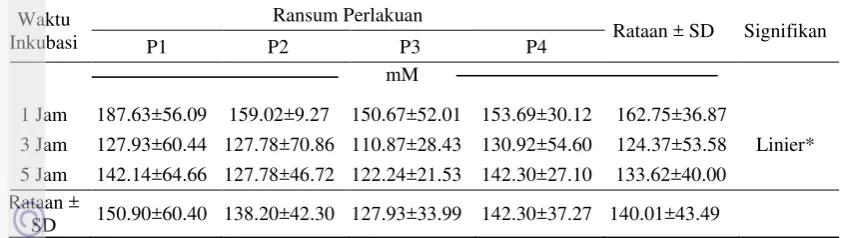
13 Tabel 5 Pengaruh perlakuan terhadap rataan konsentrasi VFA Total
Waktu
1 Jam 187.63±56.09 159.02±9.27 150.67±52.01 153.69±30.12 162.75±36.87
Linier* 3 Jam 127.93±60.44 127.78±70.86 110.87±28.43 130.92±54.60 124.37±53.58
5 Jam 142.14±64.66 127.78±46.72 122.24±21.53 142.30±27.10 133.62±40.00 Rataan ±
SD 150.90±60.40 138.20±42.30 127.93±33.99 142.30±37.27 140.01±43.49
P1 = jerami padi+SKN A1; P2 = jerami padi+SKN A2; P3 = jerami padi+SKN A3; P4 = jerami padi+SKN A4. Signifikansi: *(P<0.05) (Uji Ortogonal Polinomial).
Lamanya waktu inkubasi dapat mempengaruhi produksi VFA yang dihasilkan, proses metabolisme karbohidrat yang mudah difermentasi akan menghasilkan konsentrasi VFA yang tinggi pada awal inkubasi. Hasil ini makin diperkuat dengan pernyataan Rahmawati (2001), yaitu semakin lamanya waktu inkubasi menyebabkan konsentrasi VFA menurun karena telah digunakan oleh mikroba rumen untuk membentuk protein mikroba. Hal ini senada dengan Sutardi (1980) bahwa pada 1 sampai 1.5 jam waktu inkubasi pakan merupakan awal penentu dari kadar NH3 dan atau VFA suatu pakan yang berasal dari solubilitas
dari pakan itu sendiri. Hasil ini dapat menunjukkan bahwa penggunaan SKN dapat menjadi sumber energi (VFA) diawal waktu inkubasi.
Menurut McDonald et al. (2002), konsentrasi VFA total sangat bervariasi bergantung kepada pakan dan lama waktu setelah makan dengan konsentrasi VFA, normalnya yaitu 70 sampai 150 mM. Konsentrasi rata-rata VFA total dari seluruh perlakuan adalah 127.928 sampai 150.897 mM. Konsentrasi ini termasuk ke dalam kisaran normal VFA yang dinyatakan oleh McDonald et al. (2002). Produksi VFA dipengaruhi oleh sumber energi, produksi VFA yang tinggi merupakan kecukupan energi bagi ternak (Sakinah 2005). Sumber karbohidrat mudah tercerna yang terdapat dalam konsentrat dapat menyumbang kadar energi untuk ransum perlakuan. Penambahan konsentrat yang berbeda pada P1, P2, P3 dan P4 tidak menyebabkan perbedaan yang nyata terhadap konsentrasi VFA. Hal ini dapat dikarenakan pada seluruh perlakuan menggunakan molases sebagai bahan organik yang mudah terfermentasi. Pakan yang mengandung gula yang mudah terfermentasi (molases) akan memenuhi kebutuhan mikroba secara cepat setelah pemberian pakan (Dixon 1986). Penambahan ampas teh dan daun kembang sepatu ke dalam ransum P3 dan P4 dapat meningkatkan produksi VFA total. Hal ini didukung oleh pernyataan Setiani (2002), fermentasi karbohidrat dari kombinasi 50% ampas teh dan 50% kembang sepatu merupakan kombinasi optimal untuk menghasilkan produksi VFA tinggi. Hal ini diduga karena terjadinya degradasi sempurna fraksi-fraksi yang mudah difermentasi dari daun kembang sepatu dan ampas teh. Penambahan agen defaunasi, mineral organik dan protein bypass (ampas teh) sebenarnya bernilai positif jika digunakan untuk pencernaan di organ pasca rumen (usus halus) pada ternak ruminansia.
14
rumen sebagai sumber energi dan diserap oleh dinding rumen. Akan tetapi, penelitian ini dilakukan secara in vitro sehingga VFA tidak mungkin untuk diserap oleh dinding rumen. Aktivitas mikroba yang bervariasi dalam mencerna diduga juga dapat mempengaruhi VFA yang terbentuk.
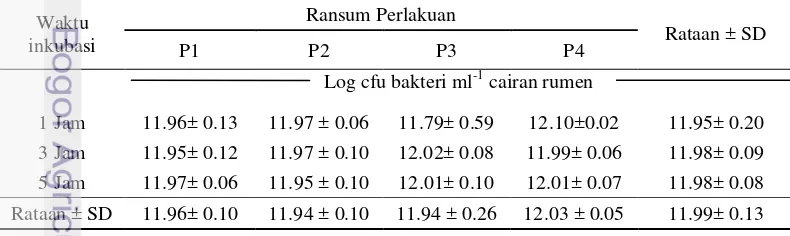
Populasi Bakteri Total
Mikroorganisme yang ada di dalam rumen dapat hidup dan melakukan aktivitasnya apabila kondisi lingkungannya mendukung. Populasi bakteri rumen total merupakan salah satu cerminan metabolisme yang terjadi di dalam rumen. Bakteri dapat hidup optimum pada pH 5.5 sampai 7.0 dalam kondisi tanpa oksigen, suhu antara 39 sampai 40 ºC dan adanya produk fermentasi pada konsentrasi sedang (Hungate 1966). Bakteri rumen dapat mentoleransi asam organik yang tinggi tanpa mempengaruhi metabolisme yang akan merugikan bakteri tersebut (Kamra 2005). Bakteri total yang terdapat dalam cairan rumen berdasarkan jenis bahan yang difermentasi dan hasil akhirnya terdiri atas bakteri pemanfaat selulosa, bakteri pemanfaat hemiselulosa, bakteri pemanfaat pati, bakteri penghasil metan, bakteri pemanfaat gula, bakteri pemanfaat asam, bakteri pemanfaat asam, dan bakteri pemanfaat lipid (Dehority dan Burk 2003).
Hasil sidik ragam menunjukkan bahwa perlakuan ransum (faktor P) tidak mempengaruhi populasi bakteri total, demikian juga dengan perlakuan waktu inkubasi (faktor B) maupun interaksi keduanya. Total bakteri yang didapatkan pada masing-masing perlakuan menghasilkan rataan berkisar 11.94 sampai 12.03 log cfu bakteri ml-1 cairan rumen, hasil rataan tersebut masih dalam kisaran normal jumlah bakteri di dalam rumen. Kamra (2005) menyatakan bakteri dalam rumen dapat mencapai 1010 sampai 1011 cfu ml-1. Berdasarkan hasil yang didapat, jumlah bakteri total pada penelitian ini menghasilkan jumlah yang lebih tinggi apabila dibandingkan dengan penelitian sebelumnya (Sari 2012) yang mendapatkan hasil total bakteri sebanyak 105cfu ml-1. Hal ini dapat disebabkan
kandungan SK ransum yang lebih tinggi pada penelitian Sari (2012), sehingga bakteri membutuhkan waktu yang lama untuk menaikkan populasinya. Tabel 6 menunjukkan data rataan populasi bakteri total.
Tabel 6 Pengaruh perlakuan terhadap rataan populasi bakteri total Waktu
inkubasi
Ransum Perlakuan
Rataan ± SD
P1 P2 P3 P4
Log cfu bakteri ml-1 cairan rumen
1 Jam 11.96± 0.13 11.97 ± 0.06 11.79± 0.59 12.10±0.02 11.95± 0.20
15 amonia yang dihasilkan pada percobaan ini tidak berbeda nyata pula antar perlakuan. Penambahan daun lamtoro pada ransum P1 yang mengandung protein yang baik karena keseimbangan asam aminonya dapat menyebabkan bakteri akan distimulasi pertumbuhannya. Wallace et al. (2003) menyebutkan bahwa bakteri selulolitik akan distimulasi pertumbuhannya dengan adanya asam amino. Menurut Danirih (2004), faktor N-NH3 merupakan faktor nutrien yang paling penting
secara kuantitatif untuk bakteri.
Mikroba rumen dapat mentolerir antinutrisi dalam ransum. Zat antinutrisi seperti tanin dan saponin yang terdapat dalam ransum P3 dan P4 disintesis oleh tanaman untuk melindungi diri dari serangan mikroba, oleh karena itu zat antinutrisi ini memiliki aktivitas antimikroba (Kamra 2005). Saponin mempunyai kemampuan untuk membentuk ikatan dengan kolesterol yang ada pada membran sel eukariotik, tetapi tidak pada sel prokariotik, sehingga adanya saponin hanya akan mempengaruhi populasi protozoa tanpa menghambat pertumbuhan bakteri (Wina et al. 2005). Tanin merupakan senyawa polifenol yang mampu mengikat protein dan membentuk senyawa komplek (Makkar 2003). Secara umum tanin mempunyai pengaruh menurunkan penggunaan pakan secara in vivo, terutama penggunaan protein dan menurunkan berbagai aktivititas enzim. Selain itu tanin juga menurunkan serangan mikrobial terhadap pakan (Suhartati 2005).
Lama waktu inkubasi juga tidak mempengaruhi populasi bakteri total. Secara umum terjadi peningkatan jumlah populasi dengan semakin lamanya waktu inkubasi. Waktu inkubasi 1 jam bakteri memiliki makanan yang cukup dari media sehingga dapat tumbuh sebagaimana seharusnya. Peningkatan populasi bakteri dengan semakin lamanya waktu inkubasi akan mampu menstimulasi pencernaan karbohidrat kompleks yang lebih baik dan lebih cepat. Menurut Saragi (2012), karbohidrat kompleks dari pakan berserat tinggi dirubah menjadi VFA oleh mikroba selulolitik. Church (1979) menyatakan bahwa makanan adalah faktor terpenting yang mempengaruhi jumlah dan proporsi relatif dari spesies yang berbeda dalam rumen. Mikroba akan beradaptasi dengan substratnya untuk mencapai keseimbangan baru. Church (1979) menyatakan bahwa konsentrasi VFA total sangat berhubungan dengan populasi mikroba rumen, khususnya bakteri selulolitik dan amilolitik.
Populasi Protozoa Total
16
Tabel 7 Pengaruh perlakuan terhadap rataan populasi protozoa total Waktu
P1 = jerami padi+SKN A1; P2 = jerami padi+SKN A2; P3 = jerami padi+SKN A3; P4 = jerami padi+SKN A4. Signifikansi: *(P<0.05) (Uji Ortogonal Polinomial).
Hasil sidik ragam menunjukkan bahwa populasi protozoa total dipengaruhi oleh kelompok (P<0.01) yaitu cairan rumen yang digunakan, dan waktu inkubasi (B) (P<0.05), tetapi tidak dipengaruhi oleh faktor ransum perlakuan (faktor P), dan interaksi antara kedua faktor. Uji ortogonal polinomial pada faktor B (perlakuan waktu inkubasi) menunjukkan bahwa meningkatnya waktu inkubasi dapat meningkatkan jumlah protozoa total di dalam rumen (P<0.05) yang mengikuti persamaan y = 0.0282x + 4.7411 dengan R² = 0.66.
Populasi protozoa yang dihasilkan dari penambahan SKN pada ransum P1, P2, P3 dan P4 tidak berbeda. Hal ini dapat terjadi karena protozoa banyak menempel dan mencerna pakan yang ada seperti yang dikatakan oleh Jouany (1991) bahwa sebenarnya sebagian besar mikroba rumen juga menempel pada partikel makanan dalam rumen. Selain itu adanya saponin dalam tepung daun kembang sepatu dalam konsentrat dapat menurunkan populasi protozoa (Goel et al. 2008). Walaupun penurunan yang terjadi tidak signifikan, protozoa mampu beradaptasi terhadap keberadaan saponin, sehingga saponin tidak memiliki kapasitas menekan populasi protozoa secara kuat. Wina et al. (2005) menyatakan bahwa bakteri rumen memiliki kemampuan untuk mendegradasi sebagian saponin, sehingga saponin tidak memiliki kapasitas untuk menekan populasi protozoa. Penggunaan agen defaunasi (daun kembang sepatu) dalam ransum P4 diduga dapat mengurangi jumlah populasi protozoa yang meningkat akibat sumber pati (molases) yang ditambahkan ke dalam ransum P1, P2, P3 dan P4. Populasi protozoa yang berkurang akan menyebabkan bakteri rumen banyak mendapatkan pakan untuk didegradasi maupun untuk pertumbuhan bakteri rumen itu sendiri tanpa persaingan dengan protozoa (Wahyuni 2008). Oleh karena itu, pengembangan SKN yang baru dapat menghasilkan efek yang sama dengan SKN yang digunakan oleh Saputra (2011).
Sintesis Protein Mikroba
17 perkembangannya dikendalikan oleh ketersediaan energi, kerangka karbon, nitrogen, mineral dan nutrien lainnya (Owens dan Goestch 1988). Protein mikroba mengandung 80% asam amino dengan kecernaan nyata di dalam usus sekitar 80%, sedangkan protein pakan yang tidak terdegradasi oleh mikroba rumen mempunyai kecernaan yang bervariasi 50% sampai 90%, tergantung dari jenis bahan pakan (Verite dan Peyraund 1989). Rataan sintesis protein mikroba dapat dilihat pada Tabel 8.
Hasil sidik ragam menunjukkan bahwa sintesis protein mikroba tidak dipengaruhi oleh faktor perlakuan pakan (faktor P), waktu inkubasi (faktor B), dan inkubasi antara kedua faktor, tetapi dipengaruhi oleh kelompok (P<0.01) yaitu cairan rumen. Kandungan protein yang cukup didalam ransum perlakuan akan menghasilkan konsentrasi amonia yang cukup untuk sintesis protein mikroba.
Tabel 8 Pengaruh perlakuan terhadap rataan sintesis protein mikroba Waktu
1 Jam 288.46±281.82 254.90±132.55 186.26±68.76 161.44±29.46 222.76±128.15
3 Jam 189.70±105.11 195.86±77.57 150.38±76.42 220.47±235.15 189.10±123.56 5 Jam 174.62±94.08 138.08±51.36 154.28±69.26 140.43±62.18 151.85±69.22 Rataan ±
SD 217.59±160.34 196.28±87.16 163.64±71.48 174.11±108.93 187.91±106.98
P1 = jerami padi+SKN A1; P2 = jerami padi+SKN A2; P3 = jerami padi+SKN A3; P4 = jerami padi+SKN A4.
Konsentrasi amonia yang tidak berbeda dari masing-masing perlakuan ransum menjadikan sintesis protein mikroba yang dihasilkan juga tidak berbeda nyata. Konsentrasi amonia pada penelitian ini berkisar 6.19 sampai 6.97 mM dan konsentrasi VFA berkisar 127.93 sampai 150.90 mM; konsentrasi amonia merupakan sumber nitrogen utama dan penting untuk sintesis protein mikroba (Sakinah 2005). Menurut Owens dan Goestch (1988), prekusor penyusun mikroba adalah NH3 sebagai sumber N, kerangka karbon, mineral dan unsur nutrisi
18
terfermentasi, rasio hijauan dan konsentrat, lingkungan rumen, sinkronisasi nitrogen dan energi, vitamin dan mineral (Pathak 2008).
Degradabilitas Bahan Kering (DBK) dan Bahan Organik (DBO)
Degradabilitas menunjukkan tingkat degradasi oleh mikroba rumen. Daya degradasi bahan pakan berhubungan erat dengan penyediaan zat makanan bagi ternak. Semakin besar daya degradasi suatu bahan makanan maka semakin besar pula zat makanan yang diperoleh ternak, berlaku juga sebaliknya. Rataan DBK dan DBO hasil penelitian disajikan pada Tabel 9.
Tabel 9 Pengaruh perlakuan terhadap rataan degradabilitas bahan kering dan bahan organik
5 Jam 14.44±7.08 13.61±5.90 11.26±6.10 15.62±6.75 13.73±6.46 Rataan ± SD 11.01±5.76 11.29±5.39 10.48±4.94 12.59±6.21 11.34±5.58
P1 = jerami padi+SKN A1; P2 = jerami padi+SKN A2; P3 = jerami padi+SKN A3; P4 = jerami padi+SKN A4. Signifikansi: *(P<0.05) (Uji Ortogonal Polinomial).
Hasil sidik ragam menunjukkan bahwa DBK dan DBO tidak dipengaruhi oleh ransum perlakuan (faktor P) dan interaksi antara kedua faktor, tetapi dipengaruhi oleh cairan rumen yang digunakan (P<0.01) dan pada DBK dipengaruhi pula oleh waktu inkubasi (faktor B) (P<0.05).
Hasil uji ortogonal polinomial menunjukkan bahwa DBK meningkat secara linier dengan semakin lamanya waktu inkubasi dari 1 jam ke 3 jam dan 5 jam (P<0.05). Hal ini karena BK yang terdegradasi semakin tinggi sejalan lamanya proses fermentasi, jika fermentasi terjadi lebih lama maka aktivitas mikroba rumen dalam mendegradasi pakan semakin meningkat. Kondisi ini sesuai dengan Lubis (1992) yang menyatakan pengukuran degradasi dalam rumen sangat ditentukan oleh faktor kelarutan bahan pakan dan waktu inkubasi.
19 dalam rumen yang disebabkan adanya karbohidrat yang mudah larut seperti molases dan urea. Kandungan protein dan sifat kelarutan dan ketahanan degradasi yang berbeda pada berbagai sumber protein dapat mempengaruhi kecernaan BK (Sutardi 1980), tetapi sumber protein dan jumlah yang digunakan dalam percobaan ini mempunyai efek yang tidak berbeda secara signifikan.
Rataan DBK (8.15%) dan DBO (11.34%) pada penelitian ini lebih rendah daripada yang diperoleh Saputra (2011), yaitu DBK 18.39% dan DBO 16.96%. Namun demikian, tidak terdapat perbedaan antara percobaan P1 yang nyata pada percobaan Saputra (2011) dengan perlakuan P2, P3 dan P4. Hasil ini menyatakan bahwa DBK dan DBO ransum yang digunakan oleh Saputra (2011) dan digunakan sebagai ransum kontrol pada percobaan ini tidak menyebabkan perbedaan dalam DBK dan DBO. Rendahnya DBK dan DBO pada percobaan ini dapat diakibatkan oleh SK, lignin dan silika yang berasal dari jerami padi disemua ransum perlakuan, dan adanya zat anti nutrisi tanin dari daun lamtoro, daun gamal dan ampas teh di dalam ransum percobaan dan saponin (daun kembang sepatu).
Menurut Rahmawati (2001), degradabilitas ransum berkaitan dengan komposisi nutrisi dari ransum, terutama kandungan SK. Kandungan SK yang tinggi akan menghambat gerak laju digesta di dalam alat pencernaan dan menyebabkan penurunan degradasi karbohidrat maupun zat-zat lainnya. Rendahnya DBK dan DBO tersebut juga dikarenakan adanya tanin di dalam ransum P3 dan P4. Ampas teh diduga menyebabkan degradabilitasnya menjadi lebih rendah (Lubis 1963). Hal ini sejalan dengan hasil penelitian yang diperoleh (Putri 2006), yaitu kandungan zat anti nutrisi dan adanya proses pemanasan yang dapat menyebabkan degradabilitasnya di dalam rumen menjadi lebih rendah. Wahyuni (2008) menyatakan bahwa tanin dapat memperlambat laju degradasi pakan dan menurunkan ketersediaan nutrien, akan tetapi zat makanan yang tersedia tersebut dapat memberikan proporsi yang lebih banyak untuk sintesis protein mikroba dibandingkan untuk pembentukan VFA.
Penambahan agen defaunasi (saponin) dari kembang sepatu ke dalam ransum P4 kemungkinan dapat mempengaruhi DBK melalui penurunan populasi protozoa yang meningkatkan populasi bakteri. juga oleh sifat kembang sepatu yang mudah difermentasi (Despal 1993). Meskipun demikian, suplementasi dalam percobaan ini dapat meningkatkan degradabilitas karena adanya perbaikan metabolisme dan kemampuan mikroba rumen (Suryahadi et al. 2003). Walaupun kadar SK ransum relatif tinggi, konsentrat mengandung bahan-bahan yang mudah difermentasi sehingga mikroba rumen akan mampu memfermentasi dengan lebih baik. Di dalam rumen, mikroba terlebih dahulu mencerna bahan yang mudah difermentasi seperti molases. Molases di dalam pakan berperan sebagi pelarut dan penyedia kerangka karbon yang paling mudah untuk NPN seperti urea sehingga memudahkan aktivitas fermentasi mikroba (Nurvianty 2006).
Koefisien Cerna Bahan Kering dan Koefisien Cerna Bahan Organik
20
dimanfaatkan oleh mikroba rumen dan induk semangnya (Sutardi 1977; McDonald et al. 2002). Rataan KCBK dan KCBO ditunjukkan pada Tabel 10. Tabel 10 Pengaruh perlakuan terhadap rataan koefisien cerna bahan kering dan dipengaruhi oleh faktor perlakuan pakan (faktor P). Nilai KCBK dan KCBO yang relatif sama pada P1, P2, P3, dan P4 dapat disebabkan oleh kandungan nutrien ransum perlakuan yang relatif sama (Tabel 3) seperti PK, SK dan TDN sebagaimana yang dikemukakan oleh Widodo et al. (2012). Kandungan SK pakan perlakuan P1, P2, P3 dan P4 berturut-turut 25.59%; 25.69%; 25.97% dan 25.83%. Serat kasar merupakan komponen BO yang sulit dicerna dalam rumen. Kandungan SK yang tinggi, umumnya diikuti dengan meningkatnya jumlah lignin yang mengikat selulosa dan hemiselulosa sehingga menyebabkan semakin turunnya nilai kecernaan (Tillman et al. 1998). Kecernaan BK yang relatif sama juga diduga dipengaruhi oleh kandungan PK pakan perlakuan yang relatif sama, dengan protein degradasi yang sama dalam penyediaan amonia untuk sintesis protein mikroba dan sumbangan protein untuk induk semang yang sama.
Faktor lain yang diduga menjadi penyebab nilai KCBK pakan perlakuan relatif sama yaitu kandungan TDN pakan perlakuan yang relatif sama. Pakan yang digunakan disusun dengan kandungan TDN yang relatif sama, sehingga TDN yang digunakan juga relatif sama. Kandungan TDN pada pakan perlakuan P1, P2, P3 dan P4 secara berturut-turut yaitu 55.82%, 55.55%, 55.46% dan 54.95%. TDN merupakan jumlah BO pada bahan pakan yang dapat dimanfaatkan sebagai sumber energy, baik energi untuk mikrobia rumen dan tubuh ternak dalam bentuk ATP (Tillman et al. 1998).
21 2005; Agni 2005). Tanin dapat membentuk ikatan dengan protein dan karbohidrat sehingga protein dan karbohidrat menjadi sulit dicerna (Makkar 2003).
SIMPULAN DAN SARAN
Simpulan
Ransum komplit jerami dan dedak padi yang ditambah suplemen kaya nutrien yang mengandung sumber pakan berbeda-beda menghasilkan fermentabilitas dan kecernaan in vitro yang sama. Hasil ini menunjukkan bahwa penggunaan daun lamtoro dan daun turi dapat diganti dengan daun gamal, ampas teh dan daun kembang sepatu sebagai bahan pakan alternatif. Dari penelitian ini dapat dihasilkan beberapa altermatif formula SKN. Waktu inkubasi dari 1 jam hingga 5 jam dapat menurunkan konsentrasi VFA, tetapi meningkatkan populasi protozoa dan degradabilitas BK membentuk pola linier.
Saran
Penelitian in vivo perlu dilakukan agar dapat menunjang hasil dari fermentabilitas, degradabilitas, dan kecernaan ransum komplit jerami dan dedak padi yang ditambah suplemen kaya nutrien.
DAFTAR PUSTAKA
Agni. 2005. Pemberian berbagai tingkat ampas teh (Camellia sinensis) terhadap kecernaan bahan kering, kecernaan protein dan retensi nitrogen domba lokal jantan [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Arora SP. 1995. Pencernaan Mikroba pada Ruminansia. Murwani R, penerjemah. Yogyakarta (ID): Gadjah Mada Pr.
Astuti OS. 2006. Pengaruh penambahan UMMB dan SPM terhadap kadar progesterone air susu dan performans reproduksi sapi perah laktasi [skripsi]. Bogor (ID): Institut Pertanian Bogor.
BATAN. 2005. Urea molasses multinutrient block (UMMB) [Internet]. [diunduh
2013 Okt 10]. Tersedia pada:http://www.
Infonuklir.com/Tips/atomos_ummb.htm.
Budiman. 2007. Pengaruh berbagai kombinasi jerami padi dengan daun gamal (Gliricidia maculata) terhadap kualitas silase. Bul NMT. 6(1):47-53. Chaerani L. 2004. Pemberian ransum suplemen yang mengandung ikatan ampas
tahu dengan seng dan tembaga untuk meningkatkan produksi susu sapi perah di Pangalengan [skripsi]. Bogor (ID): Institut Pertanian Bogor. Churh DC. 1979. Rumen Microbiology in Digestive Physiology and Nutrition of
Ruminant. 2 nd ed. Vol. 1.Porland. Oregon. p. 184 - 185.
22
Danirih. 2004. Evaluasi nutrisi dua macam feed block supplement (FBS) berdasarkan metabolisme dan populasi mikroba rumen [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Dehority BA, Burk A. 2003. Rumen Microbiology. Nottingham (GB): Nottingham Univ Pr.
Department of Dairy Science. 1969. General Laboratory Procedures. Madison (US): University of Wisconsin.
Despal 1993. Evaluasi nutrisi daun kembang sepatu (Hibiscus rosa-sinensis LINN) menggunakan teknik in sacco dan in vitro dengan pembanding beberapa legum pohon [skripsi]. Bogor (ID): Institut Pertanian Bogor. Dixon RM. 1986. Maximazing the rate of fibre digestion in the rumen. In :
Proceedings of The Seventh Annual Workshop of The Australian-Asian Ruminant Feeding Systems Utilizing Fibrous Agricultural Residues. [Waktu dan tempat pertemuan tidak diketahui]. Pudoc (PH): Cornell University.
Goel G, Makkar HPS, Becker K. 2008. Changes in microbial community structure, methanogenesis and rumen fermentation in response to saponin-rich fractions from different plant materials. J Appl Microbiol. 105: 770-777.
Hartadi H, Reksohadiprojo S, Tillman AD. 2005. Tabel Komposisi Pakan untuk Indonesia. Yogyakarta (ID) .Gadjah Mada University Pr.
Haryanti NP. 2005. Hubungan fermentabilitas dan kecernaan beberapa legum pohon dengan penyerapan mineral Ca dan P pada domba lokal jantan [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Haryanto B, Budiarsana IGM, Inounu I, Diwyanto K. 2002. Panduan Teknis Sistem Integrasi Padi-Ternak. Badan Litbang Pertanian. Departemen Pertanian.
Hungate RE. 1966. The Ruminant and Its Microbes. New York (US): Academic Pr.
Irawan B. 2002. Suplementasi Zn dan Cu organik pada ransum berbasis limbah agroindustri untuk pemacu pertumbuhan domba [tesis]. Bogor (ID): Institut Pertanian Bogor.
Jouany JP. 1991. Defaunation of the rumen. Di dalam : Jouany JP, editor. Rumen Microbial Metabolism and Ruminant Digestion. Paris (FR) : INRA. Hlm 239-261 .
Kalbande VH, Thomas CT. 2001. Effect of feeding bypass on rumen fermentation profile of crossbred cows. Asian Aust J Anim Sci. 14:974-978.
Kamra DN. 2005. Rumen microbial ecosystem. Current Scie. 89(1): 124–135. Lubis DA. 1963. Ilmu Makanan Ternak. Jakarta (ID): PT Pembangunan.
Lubis MH. 1992. Laju degradasi bahan kering dan bahan organik Setaria splendida, rumput lapang, dan alang-alang (Imperta cylindrica) dengan teknik in situ [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Makkar HPS. 2003. Effect and fate of tannins and strategies to overcome detrimental effect of feeding tannin-rich feeds. Small Rum Res. 49:241– 256.
23 Nuraeni S. 1993. Perlindungan protein ampas tahu dengan gambir dari degradasi dalam rumen dan efek perpaduannya dengan beberapa sumber energi [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Nurvianty AW. 2006. Uji Pakan komplit untuk ternak ruminansia secara in vitro. [skripsi]. Jakarta (ID): Universitas Nasional Jakarta.
Ogimito K, Imai S. 1981. Atlas of Rumen Microbiology. Tokyo (JP): Japan Scientific Societies Pr.
Owens FN, AL Goestch. 1988. Ruminant fermentation. Di dalam : Church DC, editor. The Ruminant Animal Physiology and Nutrition. New Jersey (US) : Engewood Cliffs. hlm 145-171.
Pathak AK. 2008. Various factors affecting microbial protein synthesis in the rumen. Veterinary World. 1(6):186-189.
Putra S. 2006. Pengaruh suplementasi agensia defaunasi segar dan waktu inkubasi terhadap degradasi bahan kering, bahan organik dan produk fermentasi secara in vitro. Anim Prod. 8 (2):121-130.
Putri HA. 2006. Fermentabilitas dan kecernaan in vitro ransum yang diberi urea molases multinutrien blok atau suplemen pakan multinutrien [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Rahmawati IGAWD. 2001. Evaluasi in vitro kombinasi lamtoro merah (Acacia villosa) dan gamal (Gliricidia maculata) untuk meningkatkan kualitas pakan pada ternak domba [skripsi]. Bogor (ID): Institut Pertanian Bogor. Sakinah D. 2005. Kajian suplementasi probiotik bermineral terhadap produksi
VFA, NH3, dan kecernaan zat makanan pada domba [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Saputra J. 2011. Kajian in vitro fermentasi dan kecernaan ransum berbasis jerami padi yang dioptimalisasi dengan penggunaan suplemen kaya nutrien [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Saragi MP. 2012. Perbaikan mutu biomineral cairan rumen dengan penambahan mineral makro terhadap aspek populasi bakteri dan protozoa rumen [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Sari F. 2012. Teknologi suplementasi dan pakan komplit pada ransum berbasis jerami padi terhadap populasi bakteri dan protozoa rumen [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Setiani E. 2002. Evaluasi in vitro kombinasi ampas the (Camellia sinensis) dengan daun kembang sepatu (Hibricus rosa-sinensis) sebagai pakan domba [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Shultz TA, Shultz E. 1969. Estimation of rumen microbial nitrogen by three analytical methods. J Dairy Sci. 53:781-784.
Sigit NA. 1983. Penilaian protein bahan makanan berdasarkan ketahanan degradasinya oleh mikroba rumen [tesis]. Bogor (ID): Institut Pertanian Bogor
24
Undegradable Dietary Protein (RUDP) dan sintesis protein mikroba. Anim Prod. 7:52–58.
Suryahadi, Bakrie B, Amrullah, Lotulong BV, Laside R. 2003. Kajian Teknik Suplementasi Terpadu untuk Meningkatkan Produksi dan Kualitas Susu Sapi Perah di DKI Jakarta. Lembaga Penelitian Institut Pertanian Bogor. Bogor (ID): Badan Penelitian dan Pengembangan Pertanian.
Suryahadi, Tjakradidjaja AS, Sunaryo D. 2012. Improving production performance of peranakan ongole cows and nutrient digestibility of rice straw based diet with energy- protein supplementation and complete feed. A Paper presented at 2nd International Seminar on Animal Industries (ISAI). Bogor (ID): Centras, LPPM Institut Pertanian Bogor.
Sutardi T. 1979. Ketahanan protein bahan makanan terhadap degradasi oleh mikroba rumen dan manfaatnya bagi produktivitas ternak. Proceeding Seminar dan Penunjang Peternakan. [Waktu dan tempat pertemuan tidak diketahui]. Bogor (ID): Lembaga Penelitian Peternakan.
Sutardi T. 1980. Sapi Perah dan Pemberian Makanannya. Departemen Ilmu Nutrisi dan Makanan Ternak. Fakultas Peternakan. Bogor (ID): Institut Pertanian Bogor.
Sutardi T. 1995. Peningkatan Produksi Ternak Ruminansia Melalui Amoniasi Pakan Serat Bermutu Rendah, Depaunasi dan Suplementasi Sumber Protein Tahan Degradasi dalam Rumen. Laporan Penelitian Hibah Bersaing 1/3 Perguruan Tinggi Tahun Anggaran 1994/1995. Bogor (ID): Institut Pertanian Bogor.
Tilley JMA, Terry RA. 1963. A two-stage tehnique for the in vitro digestion of forage crops. J British Grassland Society. 18:104-111.
Tillman AD, Hartadi H, Reksohadiprodjo S, Prawirokusumo S, Lebdosoekojo S. 1998. Ilmu Makanan Ternak Dasar. Yogyakarta: Gadjah Mada Pr.
Van Soest PJ. 1994. Nutritional Ecology of The Ruminant. 2nd Ed. Ithaca Comstock Publishing ssociates a Division of Cornell Pr.
Verite R, Peyround JL. 1989. The PDI Systems. Di dalam: Jarringe R, editor. Ruminant Nutrition. Paris (FR): INRA. hlm 33-46.
Wahyuni DS. 2008. Fermentabilitas dan degradabilitas in vitro serta produksi biomassa mikroba ransum komplit kombinasi rumput lapang, konsentrat dan suplemen kaya nutrien [skripsi]. Bogor (ID): Institut Pertanian Bogor. Wallace RJ, Atasoglu C. 2003. Metabolism and De Novo Syntheis of Amino Acids
by Rumen Microbes. Di dalam: D’Mello IPF, editor. Ed ke-2. Cambrige (US): CABI.
Widodo F, Wahyono, Sutrisno. 2012. Kecernaan bahan kering, kecernaan bahan organik, produksi VFA dan NH3 pakan komplit dengan level jerami padi
berbeda secara in vitro. J Anim Agri. 1(1): 215-230.
Widyobroto BP. 1992. Pengaruh konsentrat dalam ransum terhadap kecernaan dan sintesis N mikrobia di dalam rumen pada sapi perah produksi tinggi. Bul Peternakan Edisi Khusus : 241-249.
Wina S, Muetzel, Hoffmann EM, Makkar HPS, Becker K. 2005. The impact of saponin-containing plant material on ruminal production-A review. J Agri Food Chem. 53 : 8093–8115.
25 Lampiran 1 Sidik ragam pengaruh perlakuan terhadap konsentrasi amonia
SK db JK KT Fhit F0.05 F0.01
Total 47 1240.79 26.40
Kelompok 3 1189.11 396.37 314.19 2.89 4.44 **
Perlakuan 11 10.05 0.91 0.72 2.09 2.84 ns
Pakan 3 3.99 1.33 1.05 2.89 4.44 ns
Waktu Inkubasi 2 0.34 0.17 0.13 3.28 5.31 ns
Pakan*Waktu 6 5.72 0.95 0.76 2.39 3.41 ns
Galat 33 41.63 1.26
** sangat berbeda nyata (P<0.01), * berbeda nyata P(0.05), ns tidak signifikan; SK: sumber keragaman, Db: derajat bebas, JK: jumlah kuadrat, KT: kuadrat tengah, Fhit: nilai F.
Lampiran 2 Sidik ragam pengaruh perlakuan terhadap konsentrasi VFA total
SK Db JK KT Fhit F0.05 F0.01
Total 47 99130.18 2109.15
Kelompok 3 28664.88 9554.96 6.07 2.89 4.44 **
Perlakuan 11 18524.26 1684.02 1.07 2.09 2.84 ns
Pakan 3 3743.49 1247.83 0.79 2.89 4.44 ns
Waktu Inkubasi 2 12836.89 6418.44 4.08 3.28 5.31 *
Linier 1 6790.13 6790.13 4.31 4.14 7.47 *
Kuadratik 1 6046.76 6046.76 3.84 4.14 7.47 ns
Pakan*Waktu 6 4091.59 323.98 0.21 2.39 3.41 ns
Galat 33 56337.82 1573.97
** sangat berbeda nyata (P<0.01), * berbeda nyata P(0.05), ns tidak signifikan; SK: sumber keragaman, Db: derajat bebas, JK: jumlah kuadrat, KT: kuadrat tengah, Fhit: nilai F.
Lampiran 3 Uji ortogonal polinomial pengaruh waktu inkubasi terhadap konsentrasi VFA
Waktu Inkubasi (jam)
Komponen 1 3 5 c q JK
2604.03 1990.01 2137.89
Linier -1 0 1 -466.138 2 6790.13
Kuadratik 1 -2 1 761.898 6 6046.76
26
Lampiran 4 Sidik ragam pengaruh perlakuan terhadap populasi bakteri total
SK Db JK KT Fhit F0.05 F0.01
Total 47 1.53 0.03
Kelompok 3 0.06 0.02 0.51 2.89 4.44 ns
Perlakuan 11 0.23 0.02 0.56 2.09 2.84 ns
Pakan 3 0.06 0.02 0.53 2.89 4.44 ns
Waktu Inkubasi 2 0.01 0.004 0.11 3.28 5.31 ns
Pakan*Waktu 6 0.16 0.03 0.72 2.39 3.41 ns
Galat 33 1.24 0.04
** sangat berbeda nyata (P<0.01), * berbeda nyata P(0.05), ns tidak signifikan; SK: sumber keragaman, Db: derajat bebas, JK: jumlah kuadrat, KT: kuadrat tengah, Fhit: nilai F.
Lampiran 5 Sidik ragam pengaruh perlakuan terhadap populasi protozoa total
SK Db JK KT Fhit F0.05 F0.01
Total 47 0.98 0.02
Kelompok 3 0.44 0.15 12.37 2.89 4.44 **
Perlakuan 11 0.15 0.01 1.17 2.09 2.84 ns
Pakan 3 0.02 0.01 0.51 2.89 4.44 ns
Waktu Inkubasi 2 0.102 0.05 4.31 3.28 5.31 *
Linier 1 0.102 0.10 8.59 4.14 7.47 **
Kuadratik 1 0.0003 0.0003 0.03 4.14 7.47 ns
Pakan*Waktu 6 0.03 0.01 0.46 2.39 3.41 ns
Galat 33 0.39 0.01
** sangat berbeda nyata (P<0.01), * berbeda nyata P(0.05), ns tidak signifikan; SK: sumber keragaman, Db: derajat bebas, JK: jumlah kuadrat, KT: kuadrat tengah, Fhit: nilai F.
Lampiran 6 Uji ortogonal polinomial pengaruh waktu inkubasi terhadap populasi protozoa total
Waktu Inkubasi (jam)
Komponen 1 3 5 c q JK
76.34 77.15 78.14
Linier -1 0 1 1.81 2 0.136
Kuadratik 1 -2 1 0.17 6 0.00030
27 Lampiran 7 Sidik ragam pengaruh perlakuan terhadap sintesis protein mikroba
SK Db JK KT Fhit F0.05 F0.01
Total 47 699750.18 14888.30
Kelompok 3 308998.91 102999.64 11.51 2.89 4.44 **
Perlakuan 11 95523.75 8683.98 0.97 2.09 2.84 ns
Pakan 3 20767.30 6922.43 0.77 2.89 4.44 ns
Waktu Inkubasi 2 40262.99 20131.50 2.25 3.28 5.31 ns Pakan*Waktu 6 34493.46 5748.91 0.64 2.39 3.41 ns
Galat 33 295227.51 8946.29
** sangat berbeda nyata (P<0.01), * berbeda nyata P(0.05), ns tidak signifikan; SK: sumber keragaman, Db: derajat bebas, JK: jumlah kuadrat, KT: kuadrat tengah, Fhit: nilai F.
Lampiran 8 Sidik ragam pengaruh perlakuan terhadap degradibilitas bahan kering
SK Db JK KT Fhit F0.05 F0.01
Total 47 1822.11 38.77
Kelompok 3 587.58 195.86 7.61 2.89 4.44 **
Perlakuan 11 385.06 35.01 1.36 2.09 2.84 ns
Pakan 3 36.43 12.14 0.47 2.89 4.44 ns
Waktu Inkubasi 2 203.12 101.56 3.95 3.28 5.31 *
Linier 1 128.98 128.98 5.01 4.14 7.47 *
Kuadratik 1 74.14 74.14 2.88 4.14 7.47 ns
Pakan*Waktu 6 145.51 24.25 0.94 2.39 3.41 ns
Galat 33 849.479 25.74
** sangat berbeda nyata (P<0.01), * berbeda nyata P(0.05), ns tidak signifikan; SK: sumber keragaman, Db: derajat bebas, JK: jumlah kuadrat, KT: kuadrat tengah, Fhit: nilai F.
Lampiran 9 Uji ortogonal polinomial pengaruh waktu inkubasi terhadap degradibilitas bahan kering
Waktu Inkubasi (jam)
Komponen 1 3 5 c q JK
117.46 107.40 181.70
Linier -1 0 1 64.24 2 128.98
Kuadratik 1 -2 1 84.36 6 74.14