The identification of several mutations and genes involved in sporogenesis and gametogenesis has initiated a genetic framework for understanding gametophyte biogenesis. Recent advances include the molecular characterization of genes required for sporocyte formation and meiosis. These studies have revealed some unexpected interactions linking
development of sporophytic cells and tissues with initiation and progression of gametophyte development in angiosperms.
Addresses
Institute of Molecular Agrobiology, 1 Research Link, The National University of Singapore, Singapore 117604
*e-mail: [email protected] †e-mail: [email protected]
Current Opinion in Plant Biology2000, 3:53–57
1369-5266/00/$ — see front matter © 2000 Elsevier Science Ltd. All rights reserved.
Abbreviations ant aintegumenta GC generative cell
gem1 gemini pollen 1 gf/gfa gametophytic factor
sap sterile apetala SPL SPOROCYTELESS VC vegetative cell
Introduction
The alternation between a diploid sporophytic generation and a haploid gametophytic generation is fundamental to the life cycle of plants. In higher plants, the formation of the gametophyte from the sporophyte is the result of two sequential processes, sporogenesis and gametogenesis. The molecular and genetic mechanisms controlling these processes are as yet poorly understood. In recent years, genetic studies have begun to reveal mutations and, in some cases, the corresponding genes that control these processes. Genes controlling ovule and pollen develop-ment have been reviewed extensively [1,2•–5•,6••]. In this
article, we review progress over the past year in the area of male and female gametophyte development, focusing on genetic studies from the model plant Arabidopsis thaliana.
Formation of sporocytes
Sporogenesis starts with the differentiation of hypodermal cells of the ovules and anthers to form archesporial cells, which subsequently differentiate into sporocytes and under-go meiosis. In the ovule, the archesporial cell is a single cell that differentiates from the hypodermal layer at the distal end of the nucellus. In maize the recessive mac1 mutation results in the formation of multiple archesporial cells in ovules [7]. It is likely that Mac1 regulates archesporial cell specification in the ovule primordium through a signal that suppresses differ-entiation of neighboring hypodermal cells. Thus far, no similar mutation has been identified in Arabidopsis.
Recently, a gene required for the initiation of sporogenesis,
SPOROCYTELESS (SPL), has been identified in
Arabidopsis [8••]. In spl mutants, subepidermal cells of
anther and ovule primordia are able to form archesporial cells but subsequently fail to form megasporocytes or microsporocytes. In splovules, integument development is unaffected; however, in spl anthers, formation of the anther walls and tapetum is also disrupted. The archesporial cells divide to form sporogenous cells and primary parietal cells but further development of both cell layers is arrested. The SPL gene has been cloned and shown to encode a nuclear protein with limited similarities to MADS box transcription factors. SPL expression is restricted to sporogenous cells and microsporocytes in anthers and to megasporocytes in the ovules. Thus the primary role of SPL may be to promote the formation of male and female sporocytes and the defect in anther wall development is likely to be indirect. The implication is that development of the anther walls and tapetum is dependent upon signals from the microsporocytes; in their absence, sporophytic development of the anther cannot proceed.
Interactions between the megasporocyte and
sporophytic tissues
Sporophytic tissues such as integuments (Figure 1) play a role in the progression of the meiotic divisions of the megas-porocyte as defects in integument development in
Arabidopsis result in meiotic arrest. This is demonstrated by the aintegumenta(ant) and bel1mutants in which the integu-ments are either missing (ant) or abnormal (bel1) [9–11]. The exact nature of this role is unclear, however, and it is possible that integuments simply play a supportive (physiological or physical) role in promoting meiosis. Recently, a mutant called sap(sterile apetala) has been described in which megas-porogenesis is arrested during or just after the first meiotic division, although integument development is normal [12]. Surprisingly, while the SAP gene is expressed initially in the
Genetics of gametophyte biogenesis in
Arabidopsis
Wei-Cai Yang
*
and Venkatesan Sundaresan
†
Figure 1
Schematic drawing of a 7-cell ovule near maturity in Arabidopsis. Chalaza
Antipodals
Funiculus Central cell
Egg cell
Micropyle Synergids
Inner integument
Outer integument
nucellar region (archesporial cell and nucellar cells), later expression is predominantly in the developing integuments. The sap phenotype and the expression pattern of SAPcould be taken to mean that signals from the integuments, regulat-ed or mregulat-ediatregulat-ed by the SAP gene product, are required for the meiotic progression of the megasporocyte.
In wild-type ovules, degeneration of the nucellar tissue occurs during embryo-sac development. When megasporo-genesis is arrested, however, as in sap and spl mutants, the nucellus persists through ovule development, suggesting that formation of the functional megaspore may be required for nucellar degeneration. In spl mutants, the nucellus becomes arrested initially but later starts to divide to form an abnormal finger-like structure shortly after the completion of integument development. This may imply that signals from the developing integument are responsi-ble for suppression of nucellar growth.
A role for ethylene in the ovule?
In orchids, the ovary is immature and lacks ovules at anthe-sis, and pollination triggers ovule development. It was shown that pollination induces ethylene production but a direct causality between ethylene and ovule development could not be established [13]. More definitive evidence comes from tobacco, where ovule development is complet-ed before pollination. When pistil-specific ethylene production is abolished by gene silencing of the ethylene forming enzyme, ACC (1-aminocyclopropane-1-carboxy-late) oxidase, or its action is inhibited by silver thiosulfate, ovules are arrested and megasporocytes are unable to start or complete meiosis; consequently no embryo sacs are formed [14•]. Furthermore, application of exogenous
ethyl-ene can restore ovule development in the arrested ovary. These data demonstrate that ethylene is essential for the progression of meiosis and ovule development in tobacco. It is not known whether this is also the case for Arabidopsis, although it has been noted that the constitutive triple response 1 (ctr1) ethylene-response mutant is defective in female gametophyte development [2•]. A closer examination of
the flower phenotypes of other Arabidopsis mutants defec-tive in the ethylene pathway might be informadefec-tive.
Genes involved in meiosis
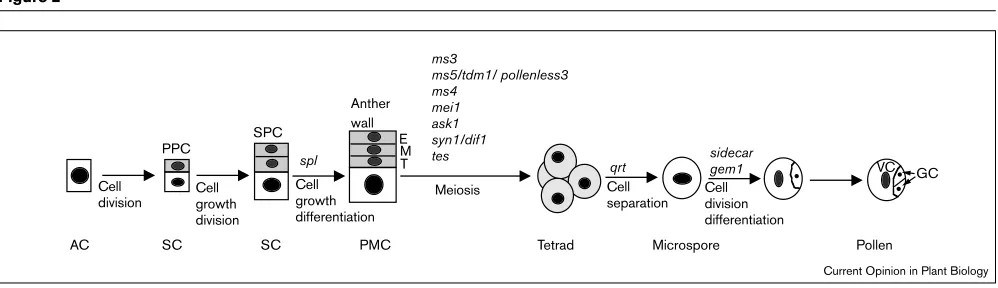
Several genes and mutations that affect male meiosis have been reported recently [15–18] (Figure 2). The MALE STERILE 5 (MS5) (THREE DIVISION MUTANT 1/POL-LENLESS3) gene has been cloned and found to encode a protein with limited homologies to a synaptonemal complex protein and to cell cycle regulatory proteins; the homology to cell cycle proteins is consistent with the observed pheno-type, in which the microsporocytes undergo an extra division after meiosis II [15,18,19•,20]. Two mutations that
affect chromosomal separation during meiosis have been identified. The Arabidopsis skp1-like (ask1) mutation is defective in chromosomal separation in anaphase I of meio-sis in the anther, resulting in the formation of a variable number (two to seven) of microspores with abnormal kary-otypes [21]. The ASK1 gene product shows high homology with the human and yeast S-PHASE KINASE ASSOCIAT-ED PROTEIN 1 (SKP1) protein, which is involved in targeting of specific proteins for ubiquitin-mediated degra-dation. Mutations in the SYNAPSIS 1/DETERMINATE INFERTILE 1 (SYN1[DIF1]) gene disrupt chromosomal segregation through meiosis I and II in both male and female meiosis, resulting in non-disjunction and sterility.
SYN1(DIF1) encodes a putative cohesin, which is a homolog of proteins required for sister chromatid cohesion in fission yeast, the Rec8p/Rad21p cohesins [22,23]. Mutations in
MEIOSIS 1(MEI1) affect male meiosis, resulting in tetrads abnormal in number, size and DNA content. The MEI1
gene has been cloned and shown to encode an 89 residue peptide with similarity to human acrosin-trypsin inhibitor [17]. Another gene, TETRASPORE(TES), is likely to act later as it is required for cytokinesis but not for the nuclear divisions in meiosis [16].
Interestingly, mutations of MS5, TES, MEI1 and ASK1
cause defects only in male meiosis, with MEI1 and ASK1 possibly involved in proteolytic reactions required for
Figure 2
AC SC
PPC
SPC
PMC SC
Anther wall
spl
Cell
division Cellgrowth division
Cell growth differentiation
Tetrad Microspore Pollen
ms3
ms5/tdm1/ pollenless3 ms4
mei1 ask1 syn1/dif1
tes sidecar
gem1 qrt
Meiosis Cell
separation Celldivision differentiation
VC GC E
M T
Current Opinion in Plant Biology
Schematic drawing depicting the sequential development events leading to pollen formation in Arabidopsis.The mutations and genes involved in each step are also indicated. AC, archesporial cell; E,
progression through the meiotic cell cycle. These obser-vations suggest that different genes are employed during male and female meiosis in plants. Compared to genes that specifically control male meiosis, few genes or muta-tions have so far been identified that specifically affect female meiosis. In the sap1 mutant discussed earlier, only a female meiotic defect is observed, but because of the failure of anther development prior to meiosis, it is not clear if the defect is actually female specific. Recently, however, a mutant called dyad has been iden-tified which results in female specific arrest of meiosis at the dyad stage, that is after meiosis I (I Siddiqui, per-sonal communication).
Establishment of functional megaspore
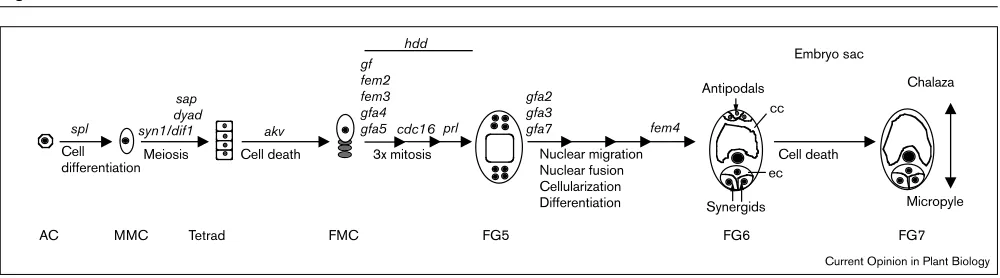
Meiosis of the megasporocytes results in the formation of four megaspores of which only the chalazal spore becomes functional, while the others degenerate (Figure 3). The cell polarity of the megasporocyte has long been considered important for the specification of the functional spore [24]. In Arabidopsis, polarity of the megasporocyte is evident from the accumulation of plastids at the chalazal end and endoplasmic reticulum and vesicles at the micropylar end [25]. In addition, the presence of plasmodesmata between the chalazal megaspore and its neighboring nucellar cells suggests that there is cell–cell communication between these two distinct cell types. We have recently identified a mutation, antikevorkian(akv) that prevents degeneration of the remaining megaspores (WC Yang, V Sundaresan, unpublished data). In plants carrying the akv mutation, any one of the surviving megaspores appears capable of forming an embryo sac and, in some cases, all four megaspores appear to develop equally and form multiple embryo sacs. The AKV gene may be required for positional signaling from the chalazal megaspore to promote degeneration of the other spores, resulting in a single embryo sac per ovule.
Female gametophyte development
Several mutations affecting nuclear division and polar nuclear fusion and cell specification associated with
megagametogenesis have been described [2•,26]. The
majority of the mutants show defects in nuclear division. These include defects in the first division as in female gametophyte 2 (fem2), fem3, gametophytic factor(gf), gameto-phytic factor 4(gfa4) and gfa5, defects in the second or third division as in cell division cycle 16 (cdc16) (HS Kwee, V Sundaresan, unpublished data) and prolifera(prl) [27], or all three divisions as in hadad(hdd) [28]. It is likely that some of these mutations involve genes that are required for operation of the cell cycle machinery, as exemplified by
prland cdc16. In gfa2, gfa3and gfa7mutants the two polar nuclei are able to migrate but fail to fuse, while gfa3 and
gfa7mutants are also defective in nuclear division as some ovules contain five nuclei or lack an embryo sac. In fem1
mutants, the embryo sac degenerates at anthesis and the degeneration of the central vacuole precedes the cell dis-integration, while the fem4 mutants seems to have pleiotropic defects in cell morphology ([26]; Figure 3). Further phenotypic characterization of the mutants and isolation of corresponding genes should reveal details of the mechanisms controlling megagametogenesis in plants.
Male gametophyte development
After meiosis, the microspore undergoes two mitotic divi-sions in Arabidopsis, to produce the male gametophyte with two sperm cells and a vegetative cell. Curiously, disruptions of the cell cycle genes cdc16 and prl (see above) do not seem to affect male gametophyte development detectably. This could be due to functional redundancy of these genes but it may also reflect the fewer number of cell divisions that are required for male gametogenesis, so that the mutations are rescued by persistence of the parental gene product.
The first division of the microspore is highly asymmetric and results in a large transcriptionally active vegetative cell (VC) and a small transcriptionally repressed generative cell (GC) within the VC. The GC undergoes one more division resulting in the formation of tricellular pollen (Figure 2). The asymmetric division is important for the fate of daugh-ter cells as revealed by differences in cell cycle progression
Figure 3
MMC Tetrad FMC FG5 FG6 FG7
AC
3x mitosis
Meiosis Nuclear migration
Nuclear fusion Cellularization Differentiation
Cell death Cell death
gf fem2 fem3 gfa4
gfa5 fem4
gfa2 gfa3 gfa7 hdd
prl
spl akv
Antipodals
Synergids ec cc cdc16
syn1/dif1 Cell
differentiation
Chalaza Embryo sac
Micropyle dyad
sap
Current Opinion in Plant Biology
Schematic drawing depicting sequentially the development events leading to embryo sac formation in Arabidopsis. The mutations and genes involved in each step are indicated accordingly.
and cell-specific gene expression [29,30••,31]. When the
asymmetry is disrupted by in vitrocolchicine treatment in tobacco, both daughters adopt the VC fate, which can be regarded as a default state [29]. Similar results have been obtained using the gem1 (gemini pollen 1) mutation, which reduces the asymmetry of the first microspore division [30••]. The altered division patterns in microspores and
reduced female transmission suggest that GEM1 plays a role in cytokinesis, and may also act in megagametogenesis.
Analysis of the gem1 mutation has also provided insights into how the asymmetry and cell fate might be established in pollen mitosis [30••]. The smaller daughter cell
result-ing from an occasional partially asymmetric division in
gem1 pollen possesses an intermediate or mixed cell fate, manifested by vegetative gene expression (VC-like) and dispersed chromatin condensation (GC-like). In some gem1
pollen, there is also failure of cytokinesis after the first mitosis, yet increased chromatin condensation of only one of the two nuclei is observed within the same cell. Therefore it is proposed that the initial transcriptionally-repressed state of the GC nucleus is a result of differential segregation of transcriptional repressors or activators prior to cell division, which may be mediated through polarized transport coupled to nuclear migration. Once the repressed state is established, it can be maintained through chro-matin remodeling or through factors that remain bound to the chromosomes [6••,30••]. Consistent with the idea that
establishment of polarity must precede asymmetric divi-sion, the mutant sidecar pollen undergoes a premature pollen mitosis which is symmetric, presumably because polarity has not yet been established [32]. One of the daughter cells goes on to divide asymmetrically to form VC and GC, ultimately resulting in the formation of a pollen grain with an additional cell.
Cell cycle synchrony of the gametes
Coordinated cell cycle progression of male and female gametes prior to fertilization is likely to be essential for their successful fusion. In most eukaryotes, gametes remain predominantly in the G1 phase of the cell cycle
through karyogamy. As a result, the zygotic nucleus con-tains 2C DNA content. But in higher plants, the cell cycle progression of the gametes is quite variable between species, and pollen with either one or two sperm cells, arrested at either G1or G2, can be found. During
pollina-tion and pollen tube growth, further cell cycle progression of the male gametes may occur for cell cycle synchrony with the female gametes at fertilization. A recent study shows that Arabidopsis male gametes belong to tricellular-G2 type where sperm cells have entered S phase at
anthesis, progress through S phase during pollen tube growth, and are in the G2phase at fertilization [33]. These
results also imply that although Arabidopsis has distinct advantages as a model system for a molecular genetic approach, it may not be amenable for in vitrofertilization studies, as nuclei from isolated pollen require further cell cycle progression before successful fertilization.
Conclusions
Genetic studies have identified a large number of muta-tions that control gametophyte development from initiation to maturity. Molecular cloning of some of the genes involved has provided us some insights about sporo-cyte formation and meiosis; however, genes for many gametophytic mutations have not been isolated, and the detailed phenotypic and molecular characterization of the mutants remains a big challenge. Strategies such as combi-nations of confocal microscopy and cell type markers are starting to be used more extensively [26,28], and will be important in future studies.
Acknowledgements
We thank Megan Griffith for useful comments and Shireen Lian for help with the figures and preparation of the manuscript. Research in our laboratory is supported by grants from the National Science and Technology Board of Singapore.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as:
• of special interest
••of outstanding interest
1. Angenent GC, Colombo L: Molecular control of ovule development.
Trends Plant Sci 1996, 1:228-232.
2. Drews GN, Lee D, Christensen CA: Genetic analysis of female • gametophyte development and function.Plant Cell 1998, 10:5-17. An excellent review on the genetic control of embryo sac development, including a good classification of developmental stages and a summary of gametophytic mutations.
3. Gasser CS, Broadhvest J, Hauser BA: Genetic analysis of ovule • development.Annu Rev Plant Physiol Plant Mol Biol 1998, 49:1-24. See annotation [5•].
4. Grossniklaus U, Schneitz K: The molecular and genetic basis of • ovule and megagametophyte development.Semin Cell Dev Biol
1998,9:227-238. See annotation [5•].
5. Schneitz K, Balasubramanian K, Schiefthaler U: Organogenesis in • plants: the molecular and genetic control of ovule development.
Trends Plant Sci 1998, 3:468-472.
Together with [3•,4•], three excellent reviews about ovule development in
Arabidopsis.
6. Twell D, Park SK, Lalanne E: Asymetric division and cell fate •• determination in developing pollen.Trends Plant Sci 1998, 3:305-310. This review provides an extensive discussion about asymmetric division of microspores and its relation to vegetative and generative cell fate during pollen development.
7. Sheridan WF, Avalkina NA, Shamrov II, Batygina TB, Golubovskaya IN: The mac1 gene: controlling the commitment to the meiotic pathway in maize.Genetics 1996, 142:1009-1020.
8. Yang W-C, Ye D, Xu J, Sundaresan V: The SPOROCYTELESS gene •• of Arabidopsis is required for initiation of sporogenesis and
encodes a novel nuclear protein.Genes Dev 1999, 13:2108-2117. Mutation in the SPL gene blocks the transition from archesporial cell into megasporocyte in ovules and microsporocyte in anthers. The SPL gene was cloned by transposon tagging and encodes a putative transcription factor.
9. Reiser L, Modrusan Z, Margossian L, Samach A, Ohad N, Haughn GW, Fisher R: The BEL1 gene encodes a homeodomain protein involved in pattern formation in the Arabidopsis ovule primordium.Cell 1995, 83:735-742.
10. Klucher KM, Chow H, Reiser L, Fischer RL: The AINTEGUMENTA gene of Arabidopsisrequired for ovule and female gametophyte development is related to the floral homeotic gene APETALA2.
Plant Cell 1996, 8:137-153.
11. Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQJ, Gerentes D, Perez P, Smyth DR: AINTEGUMENTA, an APETAL2-like gene of
organ growth.Plant Cell 1996, 8:155-168.
12. Byzova MV, Franken J, Aarts MGM, de Almeida-Engler J, Engler G, Mariani C, Van Lookeren Campagne MM, Angenent GC: Arabidopsis
STERILE APETALA, a multifunctional gene regulating inflorescence, flower, and ovule development.Genes Dev 1999, 13:1002-1014. 13. Zhang XS, O’Neil SD: Ovary and gametophyte development are
coordinately regulated by auxin and ethylene following pollination.Plant Cell 1993,5:403-418.
14. De Martinis D, Mariani C: Silencing gene expression of the • ethylene-forming enzyme results in a reversible inhibition of
ovule development in transgenic tobacco plants.Plant Cell 1999, 11:1061-1071.
Blocking of ethylene biosynthesis in tobacco pistil by silencing ACC oxidase or by application of an ethylene inhibitor results in arrest of meiosis in the ovule. Application of exogenous ethylene to the silenced ovary rescues the meiotic arrest.
15. Chaudhury AM, Lavithis M, Taylor PE, Craig S, Singh MB, Signer ER, Knox RB, Dennis ES: Genetic control of male fertility in
Arabidopsis thaliana: structural analysis of premeiotic developmental mutants.Sex Plant Reprod 1994, 7:17-28. 16. Spielman M, Preuss D, Li F-L, Browne WE, Scott RJ, Dickinson HG:
TETRASPORE is required for male meiotic cytokinesis in
Arabidopsis thaliana.Development1997, 124:2645-2657. 17. He C, Mascarenhas JP: MEI1, an Arabidopsis gene required for
male meiosis: isolation and characterization.Sex Plant Reprod
1999, 11:199-207.
18. Ross KJ, Franz P, Armstrong SJ, Vizir I, Mulligan B, Franklin FCH, Jones GH: Cytological characterization of four meiotic mutants of
Arabidopsis isolated from T-DNA transformed lines.Chromosome Res 1997, 5:551-559.
19. Sanders PM, Bui AQ, Weterings K, McIntire KN, Hsu Y-C, Lee PY, • Truong MT, Beals TP, Goldberg RB: Anther development defects in
Arabidopsis thaliana male-sterile mutants.Sex Plant Reprod
1999, 11:297-322.
Results of extensive screen for male-sterile mutants and the cloning of the
POLLENLESS3 gene. Also includes a detailed description of anther devel-opment.
20. Glover J, Grelon M, Craig S, Chaudhury A, Dennis E: Cloning and characterization of MS5 from Arabidopsis: a gene critical in male meiosis.Plant J 1998, 15:345-356.
21. Yang M, Hu Y, Lodhi M, McCombie WR, Ma H: The Arabidopsis SKP1-LIKE gene is essential for male meiosis and may control homologue separation.Proc Natl Acad Sci USA1999, 96:11416-11421.
22. Bai X, Peirson BN, Dong F, Xue C, Makaroff CA: Isolation and characterization of SYN1, a RAD21-like gene essential for meiosis in Arabidopsis.Plant Cell 1999, 11:417-430.
23. Bhatt AM, Lister C, Page T, Fransz P, Findlay K, Jones GH, Dickinson HG, Dean C: The DIF1 gene of Arabidopsis is required for meiotic chromosome segregation and belongs to the
REC8/RAD21 cohesin gene family.Plant J1999, 19:463-472. 24. Willemse MTM: Polarity during megasporogenesis and
megagametogenesis.Phytomorphology 1981, 31:124-134. 25. Bajon C, Horlow C, Motamayor JC, Sauvanet A, Robert D:
Megasporogenesis in Arabidopsis thaliana L.: an ultrastructural study.Sex Plant Reprod 1999, 12:99-109.
26. Christensen CA, Subramanian S, Drews GN: Identification of gametophytic mutations affecting female gametophyte development in Arabidopsis.Dev Biol 1998, 202:136-151. 27. Springer PS, McCombie WR, Sundaresan V, Martienssen RA:
Gene-trap tagging of PROLIFERA, an essential MCM2-3-5-like gene in
Arabidopsis.Science 1995, 268:877-880.
28. Moore JM, Calzada JPV, Gagliano W, Grossniklaus U: Genetic characterization of hadad, a mutant disrupting female gametogenesis in Arabidopsis thaliana.Cold Spring Harb Symp Quant Biol 1997, 62:35-47.
29. Eady C, Lindsey K, Twell D: The significance of microspore division and division symmetry for vegetative cell-specific transcription and generative cell differentiation.Plant Cell 1995, 7:65-74. 30. Park SK, Howden R, Twell D: The Arabidopsis thaliana
•• gametophytic mutation gemini pollen1 disrupts microspore polarity, division asymmetry and pollen cell fate.Development
1998, 125:3789-3799.
Mutation in GEM1 causes aberrant cytokinesis during pollen mitosis I, resulting in reduced asymmetry of this cell division. The phenotypic analysis of this mutation provides important insights into pollen development.
31. Xu H, Swoboda I, Bhalla PL, Singh MB: Male gametic cell-specific gene expression in flowering plants.Proc Natl Acad Sci USA
1999, 96:2554-2558.
32. Chen YCS, McCormick S: sidecar pollen, an Arabidopsis thaliana
male gametophytic mutant with aberrant cell divisions during pollen development.Development1996, 122:3243-3253. 33. Friedman WE: Expression of the cell cycle in sperm of
Arabidopsis: implications for understanding patterns of gametogenesis and fertilization in plants and other eukaryotes.