Karakter Genetik Protein Membran Virus Avian Influenza
Subtipe H5N1
N.L.P.
I
NDI
D
HARMAYANTI
1
,
D.A.
H
EWAJULI
1
,
A.
K
R
ATNAWATI
1
,
R.
I
NDRIANI
1
dan D
ARMINTO
2
1
Balai Besar Penelitian Veteriner JL RE Martadinata 30, Bogor
2Pusat Penelitian dan Pengembangan Peternakan JL Raya Pajajaran, Bogor
(Diterima dewan redaksi 26 Juli 2010)
ABSTRACT
D
HARMAYANTI
,
N.L.P.I.,
D.A.
H
EWAJULI
,
A.
R
ATNAWATI
,
R.
I
NDRIANI
and
D
ARMINTO
. 2010. Genetic characteristic of protein
membran of avian influenza viruses H5N1 subtype.
JITV
15(3): 231-239.
In 2006-2008 there were findings about the antigenic drift on AI virus due to vaccination and the AI H5N1 subtype viruses
which was similar to H5N1 viruses in human. The findings indicated that the AI viruses continue and undergoing to mutate and
try to adapt with their environment. The objective of this study to characterize the mutation of recent AI viruses (2009) on the
membran protein namely Hemagglutinin (HA), Neuraminidase (NA) and Matrix 2 (M2). In this study RT-PCR – sequencing
methods and genetic analysis for the protein membran of AI viruses were used. Result revealed that there were specific mutation
belong to AI 2009 viruses on HA and NA protein such as AI virus mutation in 2008 which isolated from backyard chicken. The
mutations were non synonimous and not caused by immunological pressure. Furthermore, M2 analysis indicated that the viruses
were resistant to amantadine.
Key Words:
Mutation, AI Subtype H5N1 Viruses, Membran Protein
ABSTRAK
D
HARMAYANTI
,
N.L.P.I.,
D.A.
H
EWAJULI
,
A.
R
ATNAWATI
,
R.
I
NDRIANI
dan
D
ARMINTO
. 2010. Karakter Genetik Protein Membran
virus avian influenza subtipe H5N1.
JITV
15(3): 231-239.
Ditemukannya virus AI yang mengalami
antigenic drift
akibat vaksinasi pada tahun 2006, 2007 dan 2008 serta beberapa
virus yang mempunyai kemiripan dengan virus H5N1 pada manusia memperlihatkan virus AI subtipe H5N1 di Indonesia sedang
dan terus bermutasi dan berusaha beradaptasi dengan lingkungannya. Penelitian ini bertujuan untuk mengetahui karakter mutasi
pada tiga protein membran yaitu Hemagglutinin (HA), Neuraminidase (NA) dan Matrix 2 (M2) pada virus AI subtipe H5N1
yang diisolasi pada tahun 2009. Metode yang digunakan dalam penelitian ini adalah propagasi virus pada telur
specific pathogen
free
(SPF) dan karakterisasi dilakukan dengan RT-PCR sekuensing pada tiga protein membran virus AI subtipe H5N1 tahun
2009 yaitu HA, NA dan M2 dan analisis genetika. Hasil penelitian memperlihatkan mutasi spesifik terjadi pada virus AI tahun
2009 yaitu pada gen HA dan pada gen NA seperti yang dimiliki oleh virus AI tahun 2008 asal ayam buras. Virus yang dianalisis
menunjukkan bahwa mutasi yang terjadi bersifat non sinonim dan tidak disebabkan karena tekanan vaksinasi. Enam virus tahun
2009 yang dianalisis juga menunjukkan resisten terhadap amantadin yang ditunjukkan oleh adanya mutasi pada protein M2.
Kata Kunci
: Mutasi, Virus AI Subtipe H5N1, Protein Membran
PENDAHULUAN
Virus influenza adalah virus yang mempunyai
materi genetik RNA untai tunggal berpolaritas negatif
dengan genom bersegmen. Virus influenza A da B
mempunyai delapan segmen, sedangkan virus influenza
C mempunyai tujuh segmen (M
CGEOCH
et al.,
1976;
D
ESSELBERGER
et al
., 1980). Segmen-segmen tersebut
secara bebas diselubungi oleh nukleoprotein dan
dihubungkan dengan polimerase kompleks. Partikel
virus terdiri dari RNA viral (vRNA), Nukleoprotein
(NP) dan komplek polimerase yang disebut dengan
partikel ribonukleprotein (RNP) (K
ATES
et al
., 1962).
Tiga protein virus terdapat dalam membran adalah dua
glikoprotein permukaan (
spike
) yaitu hemaglutinin
(HA) dan neuraminidase (NA) dan protein
membran-channel
, (M2) (Z
EBEDEE
dan L
AMB
, 1988).
Haemagglutinin (HA) homotrimer yang mempunyai
aktifitas pengikatan terhadap reseptor dan aktivitas fusi
membran yang diaktivasi dengan pH rendah dalam
endosom selama masuknya virus ke dalam sel
(W
HARTON
et al.,
1990). Neuraminidase homotetramer
yang mempunyai aktifitas menghancurkan reseptor
untuk membebaskan virus baru dari permukaan sel
yang terinfeksi (C
OLMAN
, 1989), sedangkan M2 adalah
membran-spanning
yang juga menyediakan sinyal
untuk transpor ke permukaan sel yang berbentuk
tetramer membran
channel
(S
UGRUE
dan H
AY
, 1991;
P
INTO
et al.,
1992; W
EBSTER
et al
., 1992).
Setelah lebih dari enam tahun bersirkulasi di
Indonesia, karakter molekuler virus AI di Indonesia
telah mengalami perubahan. Virus AI yang diisolasi
dari unggas yang divaksinasi AI mengalami
antigenic
drift
yang cukup ektenstif jika dibandingkan dengan
virus AI yang diisolasi dari ayam yang tidak divaksinasi
AI (D
HARMAYANTI
, 2009). Penelitian D
HARMAYANTI
(2009) mengidentifikasi adanya motif tertentu yang
dimiliki oleh virus AI yang kemungkinan dapat
ditularkan ke manusia. Mutasi yang diamati pada virus
AI sepanjang tahun 2003-2008 adalah mutasi non
sinonim yang terjadi pada gen HA dan M. Penelitian ini
bertujuan untuk mengetahui karakter mutasi pada tiga
protein membran yaitu HA, NA dan M pada virus AI
subtipe H5N1 yang diisolasi pada tahun 2009. Hasil
penelitian diharapkan dapat memberikan gambaran dan
informasi terbaru tentang karakter genetik virus AI
subtipe H5N1 tahun 2009.
MATERI DAN METODE
Virus AI
Virus AI subtipe H5N1 yang digunakan untuk studi
ini dipropagasi pada telur
specific pathogen free
(SPF)
berembrio umur 9-11 hari. Cairan alantois hasil panen
dari telur SPF berembrio yang telah diinfeksi, diuji
lebih lanjut yaitu dikarakterisasi dengan RT-PCR dan
sekuensing.
Ekstraksi RNA dan RT-PCR
Ekstraksi RNA virus yang berasal dari cairan
alantois terinfeksi dilakukan dengan menggunakan
QIAmp RNA viral mini kit (Qiagen) (D
HARMAYANTI
,
2009). Reaksi RT-PCR dilakukan dengan menggunakan
Supercript III one step RT-PCR system (Invitrogen).
Sekuensing DNA
Strategi dan set primer untuk mengamplifikasi gen
HA dan M sesuai dengan H
OFFMAN
(2003); K
OMADINA
(2007, komunikasi pribadi) dan D
HARMAYANTI
(2009).
Fragmen DNA hasil amplifikasi dipurifikasi dengan
QIAquick PCR purification
(Qiagen). Reaksi
sekuensing dilakukan dengan menggunakan reagen Big
Dye Terminator v 3.1 (Applied Biosystem) dan DNA
sekuensing dilakukan dengan mesin Genetic Analyzer
3130 (Applied Biosystenm). Hasil sekuensing dianalisis
dengan menggunakan Finch TV
(http://www.geospiza.com/Products/finchtv.shtml) dan
pembuatan
multiple aligment
dengan menggunakan
BioEdit, versi 7
(http://www.mbio.ncsu.edu/BioEdit/bioedit.html).
Pohon filogenetika dihasilkan dengan MEGA 4
(http://www.megasoftware.net).
HASIL DAN PEMBAHASAN
Sebanyak enam virus yang berhasil dianalisis
menunjukkan bahwa hasil analisis genetika pada gen
HA1 memperlihatkan bahwa virus AI tahun 2009
mempunyai mutasi asam amino yang tidak dimiliki oleh
virus AI tahun sebelumnya (2003-2008). Mutasi asam
amino terjadi pada urutan 9 yaitu asam amino A
menjadi S (A9S). Mutasi lainnya adalah asam amino
pada urutan 69 yaitu asam amino R menjadi K (R69K)
dan asam amino R menjadi M pada posisi 205
(R205M). Mutasi yang terjadi adalah mutasi non
sinonim (Gambar 1). Protein HA virus influenza
mempunyai glikoprotein trimerik yang mempunyai 3-9
N-linked glycosylation sequons
perunit, tergantung pada
galur virus (S
CHULZE
, 1997). Pada penelitian ini virus
mempunyai sebanyak 7 tempat glikosilasi pada protein
HA1 yaitu pada posisi 11, 23, 84, 154, 165, 193 dan
286 (Gambar 1). Hal berbeda jika dibandingkan dengan
virus yang mengalami
antigenic drift
yaitu virus yang
Tabel 1
. Isolat AI subtipe H5N1 yang digunakan pada penelitian
Nama Isolat
Patogenesitas Asal
sampel
A/Chicken/West Java/Bgr-Cmgg/2009
Mortalitas tinggi
Wabah unggas, disekitar kasus manusia terinfeksi H5N1
A/Chicken/West Java/Smi-Dmn/2009 Mortalitas tinggi
Wabah unggas
A/Chicken/West Java/Smi-Endo/2009 Mortalitas tinggi
Wabah unggas
A/Chicken/West Java/Smi-Mgg/2009 Mortalitas tinggi
Wabah unggas
A/Chicken/West Java/Indramayu-Indr/2009
Mortalitas tinggi
Wabah unggas
diisolasi dari unggas yang divaksinasi (Smi-Hj18/07,
Smi-Sud1/07, SMI-M1/08, SMI-M6/08, SMI- Biot/08
termasuk virus Pwt-Wij/06 dan Smi-Pat/06) hanya
memiliki sebanyak 4 tempat glikosilasi karena tidak
mempunyai tempat glikosilasi pada posisi 84, 165 dan
193 (D
HARMAYANTI
, 2009). Hasil penelitian ini
menunjukkan bahwa seluruh virus AI 2009 yang
dianalisis tidak mengalami penurunan atau peningkatan
jumlah glikosilasi sehingga kemungkinan tidak
menciptakan populasi virus yang mengalami
peningkatan afinitas terhadap reseptor dan tidak
menghasilkan populasi virus yang lebih tahan terhadap
netralisasi daripada parentalnya (S
CHULZE
, 1997), tidak
seperti halnya virus yang diisolasi dari ayam yang
divaksinasi AI yang telah terbukti lebih tahan terhadap
netralisasi daripada induknya. Berdasarkan sekuen asam
amino pada
cleavage site
HA, enam virus yang
digunakan pada penelitian ini mempunyai rangkaian
asam amino yang menandakan virus termasuk
kelompok
highly pathogenic
dan mempunyai substitusi
pada posisi -6 protein HA1 yaitu Arginin (R)ÆSerin
(S) (Gambar 1). Variasi pada motif dapat terjadi
berkaitan dengan geografi asal isolat, tidak
dihubungkan dengan perubahan virulensi dan infeksi
pada manusia (W
RITTING
C
OMMITTEE
O
F
S
ECOND
W
ORLD
O
RGANIZATION
C
ONSULTATION
, 2008). Virus
yang dianalisis pada penelitian ini tidak mengalami
mutasi pada asam amino posisi 222 dan 224 sehingga
masih mengenal
avian receptor
(
α
2,3) dan belum
mengenal
human receptor
(
α
2,6) (S
TEVENS
et al.,
2006). Substitusi sebuah asam amino pada protein
hemaglutinin dapat mengubah spesifisitas pengikatan
sialyl-linkage
Neu5Ac2-3Gal (Ac2-3) menjadi
Neu5Ac2-3Gal (Ac2-6) atau sebaliknya. Substitusi
Ser205Tyr yang terletak pada antigenik D dari
hemaglutinin virus H3 (jaraknya jauh dari
reseptor
binding site
), menghasilkan perubahan spesifisitas
pengikatan reseptor dari 2-3 ke 2-6. Perubahan
Leu226Gln pada
pocket receptor binding site
juga
mengubah spesifisitas pengikatan reseptor 2-6 ke 2-3.
Perubahan ini sangat penting karena berpengaruh pada
kemampuan infeksi virus pada inangnya. Pada
penelitian ini, dikarenakan virus masih mengenal
avian
receptor
(Ac2-3) sehingga infeksi pada manusia
kemungkinan tertular dari unggas yang terlebih dahulu
terinfeksi virus H5N1.
Pada
multiple alignment
protein NA menunjukkan
kekhasan virus AI 2009 yaitu perubahan asam amino
pada posisi 163 yaitu V menggantikan L (V143L).
Selain itu mutasi juga terjadi pada posisi 189 yaitu asam
amino S digantikan oleh G (S189G), asam amino E
digantikan oleh asam amino K pada posisi 259
(E259K), dan asam amino G menggantikan E (G382E)
(Gambar 2). Tiga mutasi ini serupa dengan virus AI
tahun 2008 yang diisolasi dari ayam buras tanpa
vaksinasi AI yaitu isolat A/Ck/West Java/Smi-Acl/08
dan A/Ck/Banten/Srg-Fadh/08. Pada protein NA, semua
virus H5N1 Indonesia mempunyai delesi 20 asam
amino pada regio
stalk
yaitu pada posisi 48-67. Tempat
glikosilasi pada regio
stalk
dari protein neuraminidase
berperan dalam menjaga struktur tetramer dari protein
(L
UO
et al.,
1993). Semua virus pada penelitian ini tidak
mempunyai tempat glikosilasi pada
stalk
protein
neuraminidase karena delesi di daerah ini, sehingga
pada protein NA hanya memiliki 3 tempat glikosilasi
yaitu pada posisi 88, 146 dan 235. Delesi pada daerah
ini akan meningkatkan retensi virion pada membran
plasma (
M
ATROSOVICH
et al.,
1999).
Mutasi yang terjadi tidak seperti virus yang diisolasi
dari ayam yang divaksinasi AI secara intensif dan tidak
terjadi pada epitop pengenalan antibodi
(D
HARMAYANTI
,
2009), sehingga besar kemungkinan
mutasi yang terjadi bukan akibat tekanan vaksinasi
sehingga asal virus diperkirakan adalah virus yang
bersirkulasi di lingkungan. Mutasi asam amino pada
protein permukaan virus yaitu HA dan NA yang terjadi
pada virus tahun 2009 mengindikasikan bahwa virus
terus berusaha beradaptasi dengan lingkungan
sekitarnya.
Indikasi adanya mutasi pada gen NA adalah hal
baru, karena sebelumnya D
HARMAYANTI
(2009) tidak
menemukan adanya mutasi pada gen NA pada
virus-virus tahun 2003-2008, sehingga paparan mutasi hanya
terbatas pada gen HA saja serta beberapa virus
memperlihatkan mutasi pada gen internal. Hal ini
tentunya sangat menarik karena paparan mutasi pada
virus AI tahun 2009 telah mengakibatkan perubahan
asam amino pada glikoprotein permukaan virus yaitu
Neuraminidase selain Hemagglutinin. Dengan
perkataan lain bahwa hasil penelitian ini
menggambarkan virus AI sedang dan terus bermutasi.
Hasil analisis filogenetika, enam virus 2009 yang
dianalisis pada aras gen HA (Gambar 3) dan NA
(Gambar 4) menunjukkan bahwa virus tahun 2009
membentuk kelompok tersendiri meskipun masih dalam
kelompok besar virus AI subtipe H5N1 asal Indonesia,
dan berbeda dengan kelompok virus yang mengalami
antigenic drift
yang berasal dari flok ayam yang
divaksinasi AI.
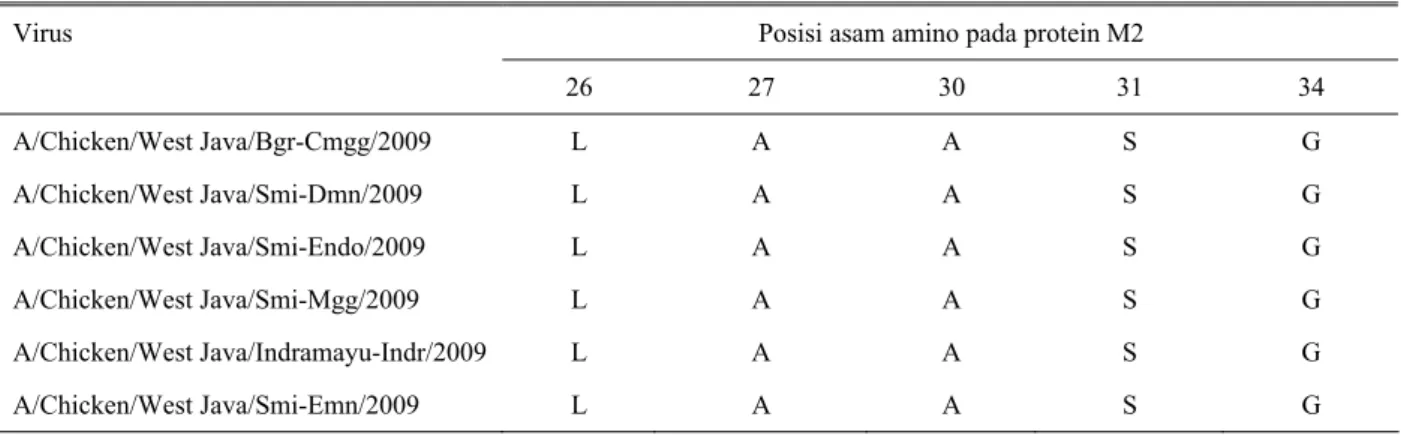
Substitusi asam amino tunggal pada asam amino
26(LeuÆPhe), 27 (ValÆAla atau Thr), 30 (AlaÆThr
atau Val), 31 (SerÆAsn atau Arg) dan 34 (GÆE)
dalam domain transmembran M2 diimplikasikan
dengan hilangnya sensitivitas bloker M2 yang
mengakibatkan resisten terhadap amantadin (H
AY
et al.,
1985; P
INTO
et al.,
1992; S
UZUKI
et al.,
2003). Analisis
pada protein M2 memperlihatkan bahwa enam virus
tahun 2009 yang digunakan pada penelitian ini
memiliki substitusi pada asam amino posisi 27 yaitu A
(ValÆAla; V27A) yang menunjukkan virus tersebut
resisten terhadap amantadin (Tabel 2). D
HARMAYANTI
10 20 30 40 50 60 70 80 90 100 110 120 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Ck/East Java/BL-IPA/03
DQICIGYHANNSTEQVDTIMEKNVTVTHAQDILEKTHNGKLCDLDGVKPLILRDCSVAGWLLGNPMCDEFINVPEWSYIVEKANPANDLCYPGNFNDYEELKHLLSRINHFEKIQIIPKS
Ck/West Java/1074/03
...L...
MD/Jakarta/DKI-Uwit/04 ...S...
MD/West Java/Bgr-Cw/05
...K...S...
Ck/Jakarta/DKI31/05
...
MD/West Java/Bgr-Cw/05
...K...S...
Ck/West Java/Smi-Hay/05
...T...S...
MD/Jakarta/Sum106/06
...T...S...
Duck/Jakarta/Slmt306/06
...S...
MD/West Java/Bks3/07
...T...S...
Ck/Pessel/BPPVRII/07
...T...S...
Ck/Inhu/BPPVRII/07
...S...
Ck/Jakarta/DKI-Nurs/07
...K.Q...S.T...S...K...R...
Ck/West Java/Smi-Hj18/07
...K.Q...S.T...S...K...R...
Ck/West Java/Smi-Sud1/07
...K.Q...S.T...S...K...R...
Ck/West Java/Smi-Acul/08
...T...S...
Ck/Banten/Srg-Fadh/08
...K...T...S...
Ck/West Java/Smi-M1/08
...N...K.Q...S.T...S...K...R...
Ck/West Java/Smi-M6/08
...N...K.Q...G...S.T...S...K...R...
Ck/West Java/Smi-Biot/08
.H...N...K.Q...S.T...S...K...R...
Ck/West Java/Bgr-Cmgg/09
...K...T...S...
Ck/West Java/Smi-Dmm/09
...T...L.K...T....D..S...
Ck/West Java/Smi-Emn/09
...E...K...T...S...
Ck/West Java/ Smi-Endo/09
...D...K...T...S...
Ck/West Java/Indramayu/09
...K...T...S...
Ck/West Java/Smi-Mgg/09
...R...K...T...S...
130 140 150 160 170 180 190 200 210 220 230 240 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Ck/East Java/BL-IPA/03
SWSDHEASSGVSSACPYQGKSSFFRNVVWLIKKNSAYPTIKRSYNNTNQEDLLVLWGIHHPNDAAEQTRLYQNPTTYISVGTSTLNQRLVPKIATRSKVNGQSGRMEFFWTILKPNDAIN
Ck/West Java/1074/03
...T...L...D...H...
MD/Jakarta/DKI-Uwit/04 ...L.R...NT...A...
MD/West Java/Bgr-Cw/05
...L.SP...T...K...N...
Ck/Jakarta/DKI31/05
...L.R...T...I...
MD/West Java/Bgr-Cw/05
...L.SP...T...K...N...
Ck/West Java/Smi-Hay/05
...L.SP...T...K...I...
MD/Jakarta/Sum106/06
...L.SP...T...K...I...
Duck/Jakarta/Slmt306/06
...L.SP...NT...K...S...
MD/West Java/Bks3/07
...L.SP...T...K..K...NEE...I...
Ck/Pessel/BPPVRII/07
...L.SP...K...I...
Ck/Inhu/BPPVRII/07
...L.SP...NT...K...
Ck/Jakarta/DKI-Nurs/07
D...L...L.SP...T...K..K...S.NVE...I....I...H...D...N...T..
Ck/West Java/Smi-Hj18/07
D...-...L.SP...TQ...T..I..K..K...I...S.NVE...I....I...T..H...D...N...T..
Ck/West Java/Smi-Sud1/07
D...-...L.SP...TQ...T..I..K..K...I...S.NVE...I....I...T..H...D...N...T..
Ck/West Java/Smi-Acul/08
...A....L.SP...T...K...TI...
Ck/Banten/Srg-Fadh/08
...L.SP...T...KT...I...ND....M...I...
Ck/West Java/Smi-M1/08
D.P...T.-...L.SP...TQ...T..I..K..K...I...S.NVE..KS....SI....I...T..H...D...N...T..
Ck/West Java/Smi-M6/08
D.P...T.-...L.SP...TQ...T..I..K..K...I...S.NVE..KS....SI....I...T..H...D...N...T..
Ck/West Java/Smi-Biot/08
D.P...T.-...L.SP...TQ...T..I..K..K...I...S.NVE..KS....SI....I...T..H...D...N...T..
Ck/West Java/Bgr-Cmgg/09
...L.SL...T...KT...I...V....NE....M...I...
Ck/West Java/Smi-Dmm/09
...M...L.SP...T...KT...I...NE....M...I...
Ck/West Java/Smi-Emn/09
...L.SP...T...KT...I...NE....M...D..I...S...
Ck/West Java/ Smi-Endo/09
...L.SP...T...KT...I...NE.D..M...I...
Ck/West Java/Indramayu/09
...L.SP...T...KT...I...NE....M...I...
Ck/West Java/Smi-Mgg/09
...L.SP...T...KT...I...NE....MRN...I...S...
250 260 270 280 290 300 310 320 330 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . .
Ck/East Java/BL-IPA/03
FESNGNFIAPEYAYKIVKKGDSAIMKSELEYGNCNTKCQTPMGAINSSMPFHNIHPLTIGECPKYVKSNRLVLATGLRNSPQRERRRKKRGLFGAIA
Ck/West Java/1074/03
...
MD/Jakarta/DKI-Uwit/04 ...
MD/West Java/Bgr-Cw/05
...
Ck/Jakarta/DKI31/05
...
MD/West Java/Bgr-Cw/05
...
Ck/West Java/Smi-Hay/05
...S...
MD/Jakarta/Sum106/06
...S...
Duck/Jakarta/Slmt306/06
...
MD/West Java/Bks3/07
...S...
Ck/Pessel/BPPVRII/07
...
Ck/Inhu/BPPVRII/07
...G...
Ck/Jakarta/DKI-Nurs/07
...D...S...
Ck/West Java/Smi-Hj18/07
...D...S...
Ck/West Java/Smi-Sud1/07
...D...S...
Ck/West Java/Smi-Acul/08
...S...
Ck/Banten/Srg-Fadh/08
...S...
Ck/West Java/Smi-M1/08
...D...I...S...
Ck/West Java/Smi-M6/08
...D...I...S...
Ck/West Java/Smi-Biot/08
...D....D...I...S...
Ck/West Java/Bgr-Cmgg/09
...D...S...
Ck/West Java/Smi-Dmm/09
...S...S...
Ck/West Java/Smi-Emn/09
...D...S...
Ck/West Java/ Smi-Endo/09
...D...S...
Ck/West Java/Indramayu/09
...S...
Ck/West Java/Smi-Mgg/09
...G...S...
Gambar 1.
Multiple alignment
protein HA1 virus AI subtipe H5N1 tahun 2003-2009
Keterangan:
Tempat glikosilasi ditandai dengan kotak tertutup dan residu asam amino didaerah
cleavage site
HA ditunjukkan dengan
kotak tertutup dengan warna abu-abu. Penomoran asam amino berdasarkan virus BL-IPA/03. Ck : Chicken; MD :
Muscovy duck
10 20 30 40 50 60 70 80 90 100 110 120 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
Goose/Guangdong/1/96 MNPNQKIITIGSICMVVGIISLMLQIGNIISIWVSHSIQTGNQHQAEPCNQSIITYENNTWVNQTYVNISNTNFLTEKAVASVTLAGNSSLCPISGWAVHSKDNGIRIGSKGDVFVIREP Ck/East Java/BL-IPA/03 ...I..V...M...S---T....P...R...S... Ck/West Java/1074/03 ...I..V...M...S---...P...R...S... MD/JakartDKI-Uwit/04 ...I..V...M...D...S---.I.N.P...N...R...S... Ck/Banten/Pdgl-Kas/04 ...I..V...M...D...S---.I.N.P...N...R...S... Ck/JakartDKI31/05 ...I..V...M....I...S---...P...R...S... MD/West Java/Bgr-Cw/05 ...AI..V...M...S---...PP...R...N... Ck/West Java/Smi-Hay/05 ...I..V...M...S---...P...R...N... MD/JakartSum106/06 ...I..V...M...S---...P...R...N... Duck/JakartSlmt306/06 ...I..V...M...S---...P...R...N... MD/West Java/Bks3/07 ...I..V...M...K...S---...P...R...N... Ck/Pessel/BPPVRII/07 ...I..V...M...K...S---...P...I...R...N... Ck/Inhu/BPPVRII/07 ...I..V...M...K...S---...P...I...R...N... Ck/JakartDKI-Nurs/07 ...I..V...M...S---...P...R...N... Ck/West Java/Smi-Hj18/07 ...I..V...M..T...S---...P.N...R...N... Ck/West Java/Smi-Sud1/07 ...I..V...M...S---...P.N...R...N... Ck/West Java/Smi-Acul/08 ...AT...I..V...M...K...S---...P...R...N...R... Ck/Banten/Srg-Fadh/08 ...A...I...M...K...S---...P...T...R...N... Ck/West Java/Smi-M1/08 ...I..V...M...S---...S.N..T...R...N... Ck/West Java/Smi-M6/08 ...I..V...M...S---...S.N...R...N... Ck/West Java/Smi-Biot/08 ...I..VG...M...S---...S.N...R...N... Ck/West Java/Smi-Dmn/09 ...I..V...MR...I...K...T.S---...P...R...N... Ck/West Java/Smi-Emn/09 ...I..V...M....I...K...T.S---...P...R...N... Ck/West Java/Smi-Endl/09 ...I..V...M....I...K...T.S---...P...R...N... Ck/West Java/Smi-Mgg/09 ...I..V...M....I...K...T.S---T....P....T...R...N... Ck/West Java/Indramayu/09 ...S...I..V...M...K...T.S---...P...T...R...N...
130 140 150 160 170 180 190 200 210 220 230 240 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | Goose/Guangdong/1/96 FISCSHLECRTFFLTQGALLNDKHSNGTVKDRSPHRTLMSCPVGEAPSPYNSRFESVAWSASACHDGTSWLTIGISGPDNGAVAVLKYNGIITDTIKSWRNNILRTQESECACVNGSCFT Ck/East Java/BL-IPA/03 ... Ck/West Java/1074/03 ... MD/JakartDKI-Uwit/04 ... Ck/Banten/Pdgl-Kas/04 ... Ck/JakartDKI31/05 ... MD/West Java/Bgr-Cw/05 ... Ck/West Java/Smi-Hay/05 ... MD/JakartSum106/06 ...N...S..E... Duck/JakartSlmt306/06 ...N...E...K... MD/West Java/Bks3/07 ...E... Ck/Pessel/BPPVRII/07 ...E... Ck/Inhu/BPPVRII/07 ...E... Ck/JakartDKI-Nurs/07 ...E... Ck/West Java/Smi-Hj18/07 ...E... Ck/West Java/Smi-Sud1/07 ...E... Ck/West Java/Smi-Acul/08 ...G...E... Ck/Banten/Srg-Fadh/08 ...G...E... Ck/West Java/Smi-M1/08 ...E... Ck/West Java/Smi-M6/08 ...E... Ck/West Java/Smi-Biot/08 ...E... Ck/West Java/Smi-Dmn/09 ...L...G...E... Ck/West Java/Smi-Emn/09 ...L...G...E...A Ck/West Java/Smi-Endl/09 ...L...G...E...A Ck/West Java/Smi-Mgg/09 ...H...L...G...E...V Ck/West Java/Indramayu/09 ...L...G...E...
250 260 270 280 290 300 310 320 330 340 350 360 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | Goose/Guangdong/1/96 VMTDGPSNGQASYKIFKMEKGKVVKSVELNAPNYHYEECSCYPDAGEITCVCRDNWHGSNRPWVSFNQNLEYQIGYICSGVFGDNPRPNDGTGSCGPVSPNGAYGVKGFSFKYGNGVWIG Ck/East Java/BL-IPA/03 ...D...M... Ck/West Java/1074/03 ...D...M... MD/JakartDKI-Uwit/04 ...D...M... Ck/Banten/Pdgl-Kas/04 ...D...M... Ck/JakartDKI31/05 ...D...M...A... MD/West Java/Bgr-Cw/05 ...D...M... Ck/West Java/Smi-Hay/05 ...D...M... MD/JakartSum106/06 ...D...M... Duck/JakartSlmt306/06 ...D.T...M... MD/West Java/Bks3/07 ...K...D...M... Ck/Pessel/BPPVRII/07 ...K.D...M... Ck/Inhu/BPPVRII/07 ...D...M... Ck/JakartDKI-Nurs/07 ...D...M.S... Ck/West Java/Smi-Hj18/07 ...D...M.S... Ck/West Java/Smi-Sud1/07 ...D...M.S... Ck/West Java/Smi-Acul/08 ...K...M... Ck/Banten/Srg-Fadh/08 ...K...D...M...L... Ck/West Java/Smi-M1/08 ...D...I...M.S... Ck/West Java/Smi-M6/08 ...D...I...M.S... Ck/West Java/Smi-Biot/08 ...D...I...M.S... Ck/West Java/Smi-Dmn/09 ...K...D...M... Ck/West Java/Smi-Emn/09 ...K...D...M...S... Ck/West Java/Smi-Endl/09 ...K...D...M...S... Ck/West Java/Smi-Mgg/09 ...K...D...D...M... Ck/West Java/Indramayu/09 ...V...K...D...M... 370 380 390 400 410 420 430 440 450 460 . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | Goose/Guangdong/1/96 RTKSTNSRSGFEMIWDPNGWTGTDSSFSVKQDIVAITDWSGYSGSFVQHPELTGLDCIRPCFWVELIRGRPKESTIWTSGSSISFCGVNSDTVGWSWPDD Ck/East Java/BL-IPA/03 ...S...G Ck/West Java/1074/03 ...S...G MD/JakartDKI-Uwit/04 ...S...S...G Ck/Banten/Pdgl-Kas/04 ...N...S...G Ck/JakartDKI31/05 ...P...S... MD/West Java/Bgr-Cw/05 ...S...G Ck/West Java/Smi-Hay/05 ...L...S..C... MD/JakartSum106/06 ...S..R..G Duck/JakartSlmt306/06 ...S...G MD/West Java/Bks3/07 ...S...G Ck/Pessel/BPPVRII/07 ...S...G Ck/Inhu/BPPVRII/07 ...S...G Ck/JakartDKI-Nurs/07 ...K...S...G Ck/West Java/Smi-Hj18/07 ...S...G Ck/West Java/Smi-Sud1/07 ...G...S...G Ck/West Java/Smi-Acul/08 ...E...G...S...G Ck/Banten/Srg-Fadh/08 ...E...S...G Ck/West Java/Smi-M1/08 ...S...G Ck/West Java/Smi-M6/08 ...E...S...G Ck/West Java/Smi-Biot/08 ...T...S...G Ck/West Java/Smi-Dmn/09 ...E...S...G Ck/West Java/Smi-Emn/09 ...E...Q...S...G Ck/West Java/Smi-Endl/09 ...E...Q...S...G Ck/West Java/Smi-Mgg/09 ...E...S...G Ck/West Java/Indramayu/09 ...E...S...G
Gambar 2.
Multiple alignment
protein NA virus AI subtipe H5N1 tahun 2003-2009
Keterangan:
Glikosilasi ditandai dengan kotak tertutup. Delesi 20 asam amino ditunjukkan dengan kotak tertutup warna abu-abu.
A /chicken/Kupang-3-NTT/B P P V6/2004
A /chicken/B angli B ali/B B P V6-1/2004
A /chicken/Jembrana/B P P V6/2004
A /chicken/M angarai-NTT/B P P V6/2004
A /Ck/Indo nesia/B L/2003
A /Chicken/East Java/B L-IP A /2003
A /Ck/Indo nesia/P A /2003
A /Chicken/West Java/HA M D/2006
A /chicken/Indo nesia/11/2003
A /Go o se/Guangdo ng/1/96
A /chicken/Indo nesia/7/2003
A /quail/Tasikmalaya/B P P V4/2004
A /Chicken/West Java/1074/2003
A /Chicken/Jakarta/DKI31/2005
A /Chicken/P adang/B B P VII/2006
A /chicken/Dairi/B P P VI/2005
A /Chicken/M edan/B B P V1-576/2005
A /chicken/Tebing Tinggi/B P P VI/2005
A /Chicken/P idie/B P P V1/2005
A /chicken/Deli Serdang/B P P VI/2005
A /Quail/Central Java/SM RG/2006
A /Chicken/West Java/GA RUT-M A Y/2006
A /Chicken/Indo nesia/M agelang1631-57/2007
A /Chicken/Gunung Kidul/B B VW/2006
A /Indo nesia/6/2005
A /M usco vy Duck/Jakarta/DKI-Uwit/2004
A /M ucso vy duck/West Java/B gr-Cw/2005
A /Chicken/P apua/TA 5/2006
A /Chicken/P apua/TB 1/2006
A /duck/P arepare/B B VM /2005
A /Duck/Jakarta/Slmt306/2006
A /Chicken/Inhu/B P P VRII/2007
A /Chicken/Indo nesia/B angka SelA tan1631-2
A /Chicken/P alembang/B P P V-III/2005
A /Chicken/Way Kanan/B B P VIII/2006
A /Chicken/B andar Lampung/B B P VIII/2006
A /Duck/Indramayu/B B P W109/2006
A /Indo nesia/CDC7/2005
A /Chicken/West Java/Smi-Hay/2005
A /Chicken/West Java/SM I-ENDRI1/2006
A /M ucso vy duck/West Java/B ks3/2007
A /Indo nesia/CDC1047/2007
A /Indo nesia/CDC1046/2007
A /Chicken/B anten/Srg-Fadh/2008
A
/Chicken/West Java/B o go r-Cmgg/2009
A /Chicken/West Java/Smi-Dmn/2009
A /Chicken/West Java/Indramayu-Indr/2009
A /Chicken/West Java/Smi-M gg/2009
A /Chicken/West java/Smi-Emn/2009
A /Chicken/West java/Smi-Endl/2009
A /Quail/Jakarta/JU1/2006
A /Chicken/West Java/TA SIKSOL/2006
A /Indo nesia/5/2005
A /Indo nesia/CDC370/2006
A /Chicken/West Java/TA SIK1/2006
A /Chicken/West Java/TA SIK2/2006
A /chicken/West Java/TA SIKSOB /2006
A /Chicken/M urao Jambi/B B P V-II/2005
A /Chicken/Indo nesia/P ekenbaru1631-11/200
A /Chicken/Indo nesia/P adang1631-1/2006
A /M usco vy Duck/Jakarta/HA B WIN/2006
A /Chicken/West Java/Smi-A cul/2008
A /M usco vy duck/Jakarta/Sum106/2006
A /Chicken/P essel/B P P VRII/2007
A /Chicken/Indo nesia/Semerang1631-62/2007
A /Swan/Indo nesia/M alang1631-61/2007
A /chicken/West Java/SM I-CSLK-EB /2006
A /Chicken/West Java/SM I-CSLK-EC/2006
A /Chicken/West Java/SM I-P A T/2006
A /Chicken/West Java/P WT-WIJ/2006
A /Chicken/Jakarta/DKI-Nurs/2007
A /Chicken/West Java/Smi-Sud1/2007
A /West Java/Smi-Hj18/2007
A /Chicken/West Java/Smi-B io t/2008
A /Chicken/West Java/Smi-M 1/2008
A /Chicken/West Java/Smi-M 6/2008
64
99
99
75
97
95
99
89
99
99
99
92
92
97
65
66
98
77
94
71
98
75
75
87
61
89
88
96
87
87
80
85
61
63
0.05
Virus AI H5N1
tahun 2009
Virus antigenic
drift th. 2006
Virus antigenic drift
th. 2007-2008
Gambar 3
. Filogenetika virus AI subtipe H5N1 pada aras gen HA1
Keterangan:
Virus yang berhasil diisolasi pada penelitian ditandai dengan warna biru. Nukleotida yang dianalisis pada aras gen HA1
A /Dk/Indo nesia/M S/2004
A /Chicken/East Java/B L-IP A /2003
A /Duck/IB ufeleng/B P P V1/2005
A /Duck/P ali/B B VW/2005
A /Ck/Indo nesia/P A /2003
A /Chicken/Indo nesia/Kulo n1631-47/2006
A /Chicken/Jakarta/DKI31/2005
A /Chicken/West Java/1074/2003
A /M usco vy Duck/Jakarta/DKI-Uwit/2004
A /Chicken/B anten/P dgl-Kas/2004
A /Chicken/Indo nesia/Wates83/2005
A /Duck/M adiun/B B VW1358/2005
A /Turkey/Langkat/B B P V1/2005
A /Chicken/M edan/B B P V1-498/2005
A /Chicken/Karo /B B P V1/2006
A /Chicken/Ro kan Hilli/B P P V11/2005
A /Chicken/P adang/B P P V11/2006
A /Chicken/P aulau Rampang/B P P V11/2006
A /M ucso vy duck/West Java/B gr-Cw/2005
A /Duck/Indramayu/B B VW109/2006
A /Chicken/B andar Lampung/B P P V111/2006
A /Chicken/P alembang/B P P V111/2005
A /Chicken/M urao Jambi/B P P V11/2005
A /Chicken/West Java/Smi-Hay/2005
A /Chicken/Indo nesia/So ppeng1631-71/2007
A /Swan/Indo nesia/M agelang1631-57/2007
A /Chicken/Indo nesia/Lampung1631-23/2006
A /Chicken/P essel/B P P VRII/2007
A /Chicken/Inhu/B P P VRII/2007
A /Chicken/Indo nesia/P ekenbaru1631-11/200
A /Ck/West Java/B ks 2/2007
A /Indo nesia/CDC1031/2007
A /M ucso vy duck/West Java/B ks3/2007
A /Chicken/West Java/Smi-A cul/2008
A /Chicken/B anten/Srg-Fadh/2008
A /Chicken/West Java/Smi-Dmn/2009
A /Chicken/West Java/Indramayu-Indr/2009
A /Chicken/West Java/Smi-M gg/2009
A /Chicken/West java/Smi-Emn/2009
A /Chicken/West java/Smi-Endl/2009
A /Chicken/Indo nesia/Gunung Kidul1631-33/
A /M usco vy duck/Jakarta/Sum106/2006
A /Duck/Jakarta/Slmt306/2006
A /B ird cucak wilis/Jakarta/Walko t 4/2007
A /Chicken/Jakarta/DKI-Nurs/2007
A /Ck/Jakarta/Jakbar-o nh/2007
A /Ck/Jakarta/Walko t 1/2007
A /Chicken/West Java/Smi-Hj18/2007
A /Chicken/West Java/Smi-Sud1/2007
A /Chicken/West Java/Smi-B io t/2008
A /Chicken/West Java/Smi-M 6/2008
A /Chicken/West Java/Smi-M 1/2008
A /Go o se/Guangdo ng/1/96
A /Ho ng Ko ng/483/1997
A /Ho ng Ko ng/514/97
A /Ho ng Ko ng/542/97
63
99
99
90
99
99
93
75
65
65
90
93
93
99
90
87
91
74
75
81
84
96
69
70
96
89
79
77
69
0.2
Virus AI H5N1
th 2009
Virus antigenic drift
th. 2007-2008
Gambar 4
. Filogenetika virus AI subtipe H5N1 pada aras gen NA
Keterangan:
Virus yang berhasil diisolasi pada penelitian ditandai dengan warna biru. Nukleotida yang dianalisis pada aras gen NA
adalah posisi 1-1157.
Tabel 2
. Posisi asam amino yang bertanggung jawab terhadap sensitivitas amantadin pada protein M2
Posisi asam amino pada protein M2
Virus
26 27 30 31 34
A/Chicken/West Java/Bgr-Cmgg/2009
L A A S G
A/Chicken/West Java/Smi-Dmn/2009
L A A S G
A/Chicken/West Java/Smi-Endo/2009
L A A S G
A/Chicken/West Java/Smi-Mgg/2009 L A A S G
A/Chicken/West Java/Indramayu-Indr/2009
L A A S G
A/Chicken/West Java/Smi-Emn/2009 L A A S G
bahwa dari 147 data sekuen asam amino M2 virus
influenza asal Indonesia yang dianalisis, diketahui
bahwa sebanyak 62,58%
a
tau 92 isolat virus influenza
H5N1 di Indonesia telah mengalami resistensi terhadap
amantadin, bahkan 10 virus diantaranya mempunyai
mutasi ganda yaitu pada posisi V27A dan S31N. Data
penelitian pada studi ini memperkuat penelitian
D
HARMAYANTI
et al
(2009) yaitu virus AI subtipe
H5N1 di Indonesia mengindikasikan terdapatnya
peningkatan resistensi terhadap amantadin.
KESIMPULAN
Hasil penelitian menunjukkan bahwa virus AI
subtipe H5N1 yang dianalisis pada penelitian ini
menunjukkan bahwa virus AI di Indonesia terus
bermutasi terutama pada protein membran virus. Mutasi
yang terjadi adalah spesifik pada virus AI tahun 2009
yaitu pada gen HA dan pada gen NA, virus AI tahun
2009 memilki mutasi yang spesifik seperti yang
dimiliki oleh virus AI tahun 2008 asal ayam buras.
Virus yang dianalisis menunjukkan bahwa mutasi yang
terjadi bersifat non sinonim dan tidak disebabkan
karena tekanan vaksinasi. Enam virus tahun 2009 yang
dianalisis juga menunjukkan resisten terhadap
amantadin yang ditunjukkan oleh adanya mutasi pada
protein M2. Meskipun wabah avian influenza subtipe
H5N1 pada unggas telah berkurang, namun virus ini
terus bermutasi sehingga monitoring sirkulasi virus ini
masih harus terus dilakukan untuk mengetahui
kemungkinan peningkatan keganasan virus serta
memperbaharui seed vaksin yang sesuai dengan virus
yang bersirkulasi di lapangan.
UCAPAN TERIMA KASIH
Terima kasih kepada Bapak Nana Suryana, SE dan
Teguh Suyatno, Amd atas bantuan teknisnya. Terima
kasih juga diucapkan kepada Drh Endri Baharianto atas
kontribusinya. Penelitian ini didanai oleh APBN-2009,
Badan LitBang Pertanian, Kementrian Pertanian.
DAFTAR PUSTAKA
C
OLMAN
,
P.M. 1989. Neuraminidase: Enzyme and antigen. In:
The Influenza Viruses. R.M. Krug (ed). Plenum Press.
New York. pp. 175-218.
D
ESSELBERGER
,
U.,
V.R.
R
ACAINELLO
,
J.J. Z
AZRA
and P.
P
ALASE
.
1980. The 3’ and 5’ terminal sequences of
influenza A,B and C virus RNA segments are highly
concerved and show partial inverted complementary.
Gene
. 8: 315-328.
D
HARMAYANTI
,
N.L.P.I. 2009. Perubahan Genom Dan
Karakter Virus Avian Influenza Subtipe H5N1 Pada
Unggas di Indonesia. Disertasi Program Doktor Ilmu
Biomedik. Fakultas Kedokteran. Universitas Indonesia.
Jakarta.
D
HARMAYANTI
,
N.L.P.I.,
F.
I
BRAHIM
and A. S
OEBANDRIO
.
2010. Amantadine resistant of Indonesian influenza
H5N1 subtype virus during 2003-2008.
Microbiol.
Indones.
5. 1: 11-16.
H
AY
,
A.J.,
A.J.
W
OLSTENHOLME
,
J.J. S
KEHEL
and M.H. S
MITH
.
1985. The molecular basis of the specific anti-influenza
action of amantadine.
EMBO J
. 4: 3021-3024.
H
OFFMANN
,
E.,
J.
S
TECH
,
Y.
G
UAN
,
,
R.G.
W
EBSTER
and D.R.
P
EREZ
. 2001. Universal primer set for the full-length
amplification of all influenza A viruses.
Arch. Virol.
146: 2275-2289.
K
ATES
,
M.,
A.C.A
LLISON
,
D.A.T
YRELL
, and A.T. J
AMES
.
1962. Origin of lipids in influenza virus.
Cold Sping
Harbor Symp Quant Biol
. 27: 293-301.
K
OMADINA
,
N. 2007. WHO Collaborating Centre for
Reference and Research on Influenza Centre, 45 Poplar
Rd., Parkville, Vic 3052, Australia.
L
UO
,
G.,
J.
C
HUNG
and P. P
ALESE
.
1993. Alterations of the
stalk of the influenza virus neuraminidase: deletions and
insertions.
Virus Res.
29: 141-153.
M
ATROSOVICH
,
M.,
N.
Z
HOU
,
Y.
K
AWAOKA
and R. W
EBSTER
.
1999. The surface glycoprotein of H5 influenza viruses
isolated from human, chickens and wild aquatic birds
have distinguishable properties.
J. Virol.
73: 1146-115.
M
CGEOCH
,
D.,
P. F
ELLNER
and C. N
EWTON
.
1976. Influenza
virus genome consists of eight distinct RNA species.
Proc. Natl. Acad. Sci.
73: 3045-3049.
P
INTO
,
L.H.,
L.J. H
OLSINGER
and R.A. L
AMB
.
1992. Influenza
virus M2 protein has ion chennel activity.
Cell
. 69:
517-528.
S
CHULZE
,
I.T. 1997. Effect of glycosilation on the properties
and functions of influenza virus hemagglutinin.
J. Infec.
Dis
. 176: S24-28.
S
TEVENS
,
A.,
O.
B
LIXT
,
T.M.
T
UMPEY
,
J.K.
T
AUBENBERGER
,
J.C. P
AULSON
and I.A. W
ILSON
.
2006. Structure and
receptor specificity of the hemagglutinin from an H5N1
influenza virus.
Science
. 312: 404-410.
S
UGRUE
,
R.J. and A.J. H
AY
.
1991. Structural characterics of
M2 protein of influenza A viruses: evidence that it
forms a tetrameric chennel.
Virology
. 180: 617-24.
S
UZUKI
,
H.,
R.
S
AITO
,
H.
M
ASUDA
,
H.
O
SHITANI
,
M. S
AITO
and
I. S
ATO
.
2003. Emergence of amantadine-resitstant
influenza a viruses: Epidemiological study.
J. Infect.
Chemother
9: 195-200.
W
EBSTER
,
R.G.,
W.J.
B
EAN
,
O.T.
G
ORMAN
,
T.M. C
HAMBERS
and Y. K
AWAOKA
.
1992. Evolution and Ecology of
influenza A viruses
. Microbiol. Rev.
56: 152-179.
W
HARTON
,
S.A.,
J.A.
H
AY
,
R.J.
S
UGRUE
,
J.J.
S
KEHEL
,
W.I.
W
EIS
and D.C. W
ILEY
.
1990. Membrane fusion by
influenza viruses and mechanism of action of
of X-ray Crystallography in the design of antivirus
agents. Academic Press. Orlando. pp. 1-12.
Z
EBEDEE
protein: monoclonal antibody restriction of virus growth
,
S.L.
and R.A. L
AMB
.
1988. Influenza virus M2