PERAN BAKTERI KITINOLITIK DAN FUNGI MIKORIZA
ARBUSKULAR DALAM PENGENDALIAN BUSUK PANGKAL
BATANG KELAPA SAWIT
NILDAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Peran Bakteri Kitinolitik dan Fungi Mikoriza Arbuskular dalam Pengendalian Busuk Pangkal Batang Kelapa Sawit adalah karya saya dengan arahan dari Komisi Pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Januari 2011
NILDAYANTI. The Role of Chitinolytic Bacterial and Arbuscular Mycorrhizal Fungi to Control Basal Stem Rot Palm Oil. Under the direction of MEITY SURADJI SINAGA and GIYANTO
Basal stem rot (BSR) is the most serious disease of palm oil that caused by Ganoderma boninense Pat. The objectives of these researches were to find chitinase-producing bacterial which inhibit G.boninense growth and to determine the relationship between mycorrhizal and chitinolytic bacterial in controlling the BSR. The experimenthals were conducted in laboratory, secreen house dan fields. The research activities were: (i) exploration of chitinolytic bacterial as antagonistic agents of pathogens (ii) evaluate the pathogenicity, index chitinolitic of the chitinolytic bacterial and their antagonism effect against G. boninense, (iii) study on the application of chitinolytic bacterial and AMF in controlling the oil palm BSR through split plot experimenthal design. There were seven isolate B13, B14, B16, B42, Al11, TB41 and L34 have been selected as the best chitinolase-producing bacterial and they were not pathogenic to plants. The study showed that selected bacterial have differences on chitinolytic index. All of them suppressed the growth of G. boninense above 95% in vitro antagonism test using dual culture method on liquid media. However the antagonism test on PDA only isolate of AL11, TB41 and L34 had higher antagonism capacity against the pathogen. The severity of basal stem rot that expressed by the root necrotics were reducted since the oil palm seedling showed high degrees of mycorrhizal root colonization. The application of combination of chitinolytic bacterial and AMF suppressed the disease severity (95%) higher compare to the single one (55% - 70%).
NILDAYANTI. Peran Bakteri Kitinolitik dan Fungi Mikoriza Arbuskular dalam Pengendalian Busuk Pangkal Batang Kelapa Sawit. Dibimbing oleh MEITY SURADJI SINAGA dan GIYANTO.
Ganoderma boninense Pat. merupakan penyebab penyakit busuk pangkal batang (BPB), yang menjadi kendala utama dalam pengembangan dan peningkatan produksi kelapa sawit di Asia Tenggara termasuk Indonesia. Hingga saat ini usaha-usaha pengendalian BPB belum berhasil menekan secara konsisten perkembangan penyakit dalam populasi tanaman kelapa sawit. Oleh karena itu dilakukan suatu studi yang bertujuan mendapatkan bakteri kitinolitik yang mampu mengekspresikan enzim kitinase yang dapat menghambat pertumbuhan Ganoderma boninense penyebab busuk pangkal batang (BPB) serta melihat pengaruh kombinasi antara mikoriza dan bakteri kitinolitik terhadap penyakit BPB.
Kandidat bakteri kitinolitik diisolasi dari rizosfir kelapa sawit dan diseleksi pada media koloidal kitin. Uji patogenesitas isolat bakteri posistif kitinolitik dilakukan pada tanaman tembakau. Setelah itu dilakukan uji antagonisme terhadap G. boninense dengan metode kultur ganda pada media cair dan padat, uji indeks kitinolitik, uji serta evaluasi keefektifan bakteri kitinolitik dan FMA dalam menekan keparahan penyakit BPB pada bibit kelapa sawit. Pengujian pada bibit sawit, peubah yang diamati ialah persen keparahan penyakit pada akar, berat basah akar dan tajuk, tinggi tanaman, asosiasi FMA pada akar, dan aktivitas enzim peroksidase (POD).
Hasil eksplorasi diperoleh 7 isolat bakteri positif kitinolitik yaitu isolat B13, B14, B16, B42, AL11, TB41, dan L34, serta semuanya tidak bersifat fitopatogenik. Dari 7 isolat bakteri tersebut hanya isolat B16 yang bersifat gram positif. Nilai indeks kitinolitik 7 isolat bakteri tersebut pada 7 hsi menunjukkan nilai indeks yang berbeda-beda. Hasil uji potensi antagonis 7 isolat bakteri kitinolitik terhadap G. boninense dengan metode kultur ganda menunjukkan hasil yang berbeda. Uji in vitro pada media cair, kemampuan 7 isolat bakteri uji hingga 95% dalam menekan pertumbuhan G. boninense dibandingkan dengan kontrol. Sedangkan pada media padat hanya isolat AL11, TB41 dan L34 yang menunjukkan penekanan yang relatif tinggi yaitu 52% , 44% dan 41%. Isolat B14, B13, B16 dan B42 hanya mampu menghambat pertumbuhan G. boninense kurang dari 10%.
Isolat bakteri TB41 dan AL11 digunakan untuk mengevaluasi keefektifan pengendalikan BPB pada bibit kelapa sawit. Gejala nekrotik pada akar merupakan gejala awal infeksi G. boninense. Dalam percobaan ini hingga 2 bulan setelah inokulasi (bsi) belum menunjukkan gejala busuk pada pangkal batang. Pengamatan pada akar bibit sawit yang diinokulasi G. boninense menunjukkan nekrotik berwarna coklat kehitaman. Nekrotik ini meluas seiring dengan perkembangan penyakit yang merupakan indikator perkembangan penyakit.
tertinggi ditunjukan oleh perlakuan kombinasi M1B2W2P1 dengan peresentase penekanan mencapai 95% walaupun secara statistik tidak berbeda dengan perlakuan kombinasi lainnya. Meskipun demikian, perlakuan tunggal isolat bakteri kitinolitik dan FMA juga mampu menekan keparahan penyakit. Adanya aplikasi bakteri kitinolitik meningkatkan kolonisasi FMA pada akar. Aktivitas enzim peroksidase yang tinggi tidak selalu diikuti ketahanan tanaman yang tinggi terhadap G. boninense. Aplikasi FMA dan bakteri kitinolitik baik secara tunggal maupun kombinasi meningkatkan berat tajuk dan tingi tanaman.
©
Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-undang1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB. 2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
ARBUSKULAR DALAM PENGENDALIAN BUSUK PANGKAL
BATANG KELAPA SAWIT
NILDAYANTI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Entomologi/Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
dalam Pengendalian Busuk Pangkal Batang Kelapa Sawit
Nama : Nildayanti
NRP : A352080021
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Meity Suradji Sinaga, M.Sc. Dr. Ir. Giyanto, M.Si. Ketua Anggota
Diketahui
Ketua Mayor Fitopatologi Dekan Sekolah Pascasarjana
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
Segala puji bagi Allah SWT, Tuhan seru sekalian alam yang setia membimbing hamba-hamba-Nya. Atas bantuan dan tuntunan-Nya penyusunan tesis dengan judul “Peran Bakteri Kitinolitik dan Fungi Mikoriza Arbuskular dalam Pengendalian Busuk Pangkal Batang Kelapa Sawit” dapat diselesaikan.
Penulis menyampaikan terima kasih yang tidak terhingga kepada semua pihak yang langsung maupun tidak langsung, turut andil dan memotivasi penyelesaian tesis ini, antara lain kepada :
1. Prof. Dr. Ir. Meity Suradji Sinaga, M.Sc. dan Dr. Ir. Giyanto, M.Si. selaku pembimbing yang dengan sabar dan cermat membimbing dan mengarahkan penulis dalam penyelesaian penelitian dan penulisan tesis ini
2. Seluruh Dosen Fitopatologi yang telah berkenan mentransfer dan membuka cakrawala ilmu pengetahuan kepada penulis
3. Teman-teman Mahasiswa Program Magister Fitopatologi dan Entomologi yang sering berbagi wacana
4. Ayahanda (almarhum) semoga Allah menerima semua amal ibadah beliau, Ibunda dan Kakak-kakak tercinta yang tak henti-hentinya mendukung penulis secara materiil maupun spirituil dalam penyelesaian tesis ini.
Akhirnya semoga tesis ini membawa manfaat untuk pengembangan ilmu pengetahuan. Amin.
Bogor, Januari 2011
Penulis dilahirkan di Ujung Pandang pada tanggal 17 Agusutus 1975 dari ayah Jeppe (Alm) dan ibu Mala. Penulis adalah anak bungsu dari sembilan bersaudara. Tahun 1996 penulis diterima sebagai mahasiswa Jurusan Ilmu Hama dan penyakit Tumbuhan, Fakultas Pertanian, Universitas Hasanuddin (Unhas) melalui tes UMPTN. Gelar Sarjana diperoleh pada tahun 2001. Sejak tahun 2006, penulis menjadi staf pengajar di Jurusan Budidaya Tanaman Perkebunan, Politeknik Pertanian Negeri Pangkep. Tahun 2008 penulis terdaftar sebagai mahasiswa program Magister Sains pada Program Studi Fitopatologi, Sekolah Pascasarjana, Institut Pertanian Bogor dengan beasiswa program pascasarjana (BPPS) dari Direktorat Jendral Pendidikan Tinggi, Departemen Pendidikan Nasional Republik Indonesia.
DAFTAR ISI
Halaman
DAFTAR TABEL ... xiv
DAFTAR GAMBAR ... xv
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 3
Manfaat ... 3
TINJAUAN PUSTAKA ... 5
Penyakit Busuk Pangkal Batang (BPB) ... 5
Potensi Bakteri Kitinolitik dan Mikoriza untuk Pengendalian G. boninense ... 7
Bakteri Kitinolitik ... 7
Mikoriza ... 7
BAHAN DAN METODE ... 11
Waktu dan Tempat Penelitian ... 11
Eksplorasi Bakteri Kitinolitik …….. ... 11
Uji Indeks Kitinolitik ... 12
Uji Potensi Antagonis Bakteri Kitinolitik terhadap G. boninense ... 12
Uji Keefektifan Bakteri Kitinolitik dan FMA dalam Menekan Keparahan Penyakit dan Pengaruh terhadap Pertumbuhan Bibit Sawit ... 13
Penyiapan Substrat Inokulum G. boninense ... 13
Penyiapan Formula FMA ... 13
Penyiapan Bakteri Kitinolitik ... 13
Aplikasi FMA, Bakteri Kitinolitik dan G. boninense pada Bibit Sawit ... 14
Pengamatan terhadap Tingkat Keparahan Penyakit BPB ... 15
Tingkat Asosiasi FMA pada Akar Bibit Sawit ... 15
Aktivitas enzim Peroksidase ... 17
HASIL DAN PEMBAHASAN ... 19
Eksplorasi Bakteri Kitinolitik ... 19
Evaluasi Potensi Antagonisme dan Indeks Kitinolitik ... 20
Pengaruh Aplikasi Bakteri Kitinolitik dan FMA terhadap BPB ... 23
Pengaruh Aplikasi Bakteri Kitinolitik dan FMA terhadap Pertumbuhan Bibit Sawit………... 30
SIMPULAN DAN SARAN ... 33
Simpulan ... 33
Saran ... 33
DAFTAR TABEL
Halaman
1. Perlakuan uji pada bibit sawit ... ... 14 2. Skoring bercak nekrotik akar berdasarkan persentase nekrotik akar kelapa sawit ... 15 3. Persentase penghambatan pertumbuhan G. boninense dengan
perlakuan bakteri kitinolitik pada media padat 4 hsi dan media cair 14 hsi ... 21 4. Sidik ragam perlakuan aplikasi bakteri kitinolitik, FMA dan
kombinasinya terhadap keparahan penyakit ... 24 5. Persentase kolonisasi FMA, aktivitas enzim peroksidase, dan
persentase penekanan penyakit pada akar bibit sawit pada berbegai
DAFTAR GAMBAR
Halaman 1. Isolat bakteri positif kitinolitik dengan zona bening pada media koloidal kitin ... 19 2. Penekanan pertumbuhan G. boninense oleh isolat bakteri kitinolitik pada media cair dan media padat ... 20 3. Indeks kitinolitik 7 isolat bakteri pada 7 his dan pertumbuhan
koloni bakteri kitinolitik ………. 22
PENDAHULUAN
Latar Belakang
Ganoderma boninense Pat. merupakan penyebab penyakit busuk pangkal batang (BPB), yang menjadi kendala utama dalam pengembangan dan peningkatan produksi kelapa sawit di Asia Tenggara termasuk Indonesia (Idris et al. 2004). Menurut Sinaga et al. (2003) pengembangan kelapa sawit secara besar-besaran di Indonesia dan Malaysia, dan iklim yang cocok untuk G. boninense, menyebabkan perkembangan BPB menjadi sangat pesat. Serangan G. boninense pada perkebunan sawit terus meningkat terutama pada kebun-kebun peremajaan dan timbulnya gejala juga semakin dini dengan tingkat serangan yang lebih berat. Zakaria et al. (2005) melaporkan bahwa pada kebun peremajaan kedua, kematian tanaman akibat BPB dapat mencapai 60%.
Penyakit BPB saat ini menjadi sangat penting pada perkebunan kelapa sawit di Indonesia karena tanaman yang terserang G. boninense cepat atau lambat akan berakhir dengan kematian. Anggapan bahwa G. boninense hanya menyerang tanaman tua merupakan suatu anggapan yang keliru karena saat ini G. boninense juga menyerang tanaman yang belum berproduksi. Penyakit ini menyebabkan busuk pangkal batang namun untuk menimbulkan gejala pada permukaan tanaman memerlukan periode inkubasi yang cukup lama. Gejala awal di pembibitan dapat diamati pada akar berupa nekrotik. Nekrotik pada akar ini meluas seiring dengan perkembangan penyakit, dan bila infeksi berlanjut sampai pangkal batang dapat menimbulkan pembusukan dan akhirnya tanaman mati.
alternatif pengendalian lain yang efisien dan efektif sehingga perkembangan penyakit dapat ditekan, antara lain dengan menggunakan mikrob agens antagonis (Sinaga et al. 2003; Susanto 2002)
Bakteri kitinolitik berpotensi untuk dimanfaatkan sebagai agens hayati. Bakteri ini memiliki kemampuan menghasilkan enzim kitinase untuk menghidrolisis polimer kitin dalam memperoleh karbon, nitrogen, dan energi. Kemampuan itu menyebabkan bakteri kitinolitik dapat menekan pertumbuhan dan perkembangan fungi patogen, nematoda atau serangga hama karena kitin merupakan komponen struktural dari sebagian besar dinding sel organisme tersebut (Adams 2004). Penggunaan bakteri kitinolitik sebagai agens pengendali patogen diantaranya dilaporkan oleh El-Tarabily et al. (2000), Chung et al. (2005), Suryanto et al. (2006), dan Kamil et al. (2007).
Mikroorganisme lain yang berpotensi untuk digunakan sebagai agens pengendali hayati G. boninense adalah fungi mikoriza arbuskular (FMA). FMA telah banyak dilaporkan memiliki kemampuan sebagai agens biokontrol terhadap patogen tular tanah seperti P. parasitica (Cordier et al. 1998 dan Pozo et al. 2002), Gaeumannomyces graminis var. tritici (Khaosaad et al. 2007). Sclerotuim rolfsii (Ozgonen et al. 2010). Hasil penelitian Hashim (2003) menunjukkan bahwa perlakuan mikoriza di pembibitan sawit mampu menekan penyakit yang disebabkan G. boninense mencapai 80%.
Tujuan
Penelitian bertujuan mendapatkan bakteri kitinolitik yang mampu mengekspresikan enzim kitinase yang dapat menghambat pertumbuhan Ganoderma boninense penyebab busuk pangkal batang (BPB) serta melihat pengaruh kombinasi antara mikoriza dan bakteri kitinolitik terhadap penyakit BPB.
Manfaat
TINJAUAN PUSTAKA
Penyakit Busuk Pangkal Batang (BPB)
Penyakit busuk pangkal batang (BPB) merupakan penyakit utama pada perkebunan sawit di Indonesia dan Malaysia (Idris et al. 2004). Hasil penelitian Abadi (1987), BPB di Sumatera Utara disebabkan oleh G. boninense, demikian pula di Malaysia. Ho & Nawawi (1985) melaporkan bahwa ratusan tubuh buah yang dikumpulkan dari berbagai tempat di Malaysia, semuanya adalah spesies G. boninense.
Tanaman kelapa sawit dewasa yang terserang G. boninense menunjukkan gejala awal pada pelepah daun berwarna pucat seperti kekurangan unsur hara. Tetapi hanya terbatas pada beberapa pelapah yang terdapat di pucuk. Daun akan mengalami nekrotik dimulai dari daun tua kemudian ke yang lebih muda dan tidak membuka serta lebih banyak dari biasanya. Pelepah daun patah dan menggantung. Gejala ini sering muncul pada tanaman yang sudah tua yaitu tanaman yang berumur lebih dari 15 tahun. Gejala yang paling khas dari penyakit ini adalah terjadinya pembusukan pada pangkal batang serta diikuti robohnya pohon dan adanya basidiospora sebagai tanda penyakit. Gejala internal yaitu berupa zona-zona yang mengalami pembusukan ditandai dengan adanya garis-garis berwarna coklat kehitaman. Kerusakan pada akar, terlihat adanya perubahan warna jaringan utama stele yang kemudian lapuk. Di permukaan kulit akar yang sakit tidak pernah ditemukan massa miselium. Diduga miselianya hanya terdapat di dalam jaringan akar sakit, terutama stele yang mengalami perubahan warna (Abadi 1987; Susanto et al. 2005).
semakin banyak. Kondisi ini akan mengancam kelangsungan hidup tanaman kelapa sawit muda yang baru saja ditanam untuk menggantikan tanaman yang telah mati (Sinaga et al. 2003).
Berbagai usaha telah dilakukan untuk pengendalian penyakit BPB, namun hingga saat ini belum dapat dikatakan berhasil. G. boninense bersifat soil borne dan kemampuan bertahan dalam kondisi kurang optimal yang tinggi merupakan salah satu faktor yang mempersulit usaha pengendalian baik secara kultur teknis, mekanis, maupun kimiawi. Pengendalian menggunakan fungisida baik dengan metode absorpsi akar maupun penyiraman fungisida ke dalam tanah kurang efektif, karena pengaruh sifat fisik dan kimia tanah atau terdegradasi oleh mikroflora di dalam tanah sebelum mencapai sasaran. Selain itu efek samping yang ditimbulkan dapat membahayakan lingkungan. Keragaman mikroorganisme yang berpotensi sebagai angens biokontrol dapat berkurang karena penggunaan fungisida secara terus menerus (Sinaga et al. 2003; Susanto 2002).
Beberapa pendekatan pengendalian mulai dikembangkan diantaranya pengendalian hayati dan penggunaan tanaman tahan. Menurut Agrios (2005) penggunaan tanaman tahan adalah cara yang paling mudah, murah, dan aman sepanjang varietas dari tanaman tahan yang dimaksud tersedia. Namun diperlukan waktu yang cukup lama untuk dapat menghasilkan suatu varietas tanaman tahan merupakan kendala tersendiri. Penggunaan varietas tahan yang tidak tepat dapat menimbulkan resistensi varietas mudah patah dan dapat menimbulkan masalah yang lebih berat, terlebih jika penggunaannnya bersamaan dengan aplikasi pestisida secara berlebihan seperti yang biasa dilakukan di perkebunan (Sinaga et al. 2003).
berpotensi mengendalikan patogen-patogen yang bersifat tular tanah termasuk G. boninense (Cook & Baker 1996; Sinaga et al. 2003).
Potensi Bakteri Kitinolitik dan Mikoriza untuk Pengendalian G. boninense
Bakteri Kitinolitik
Pemanfaatan mikrob kitinolitik merupakan salah satu cara pengendalian hayati yang efektif untuk fungi patogen tanaman. Mikrob ini dapat digunakan langsung sebagai agens antagonis atau menggunakan protein murni dari mikrob kitinolitik atau melalui manipulasi gen (Tahtamouni et al. 2006; Singh et al. 1999). Enzim kitinase yang diproduksi mikrob kitinolitik dapat menghidrolisis struktur kitin, yang merupakan struktur utama penyusun dinding sel, spora, tabung kecambah, dan miselia fungi, sehingga fungi tidak mampu menginfeksi tanaman. Salah satu penyakit yang dapat dikendalikan dengan mikrob kitinolitik adalah penyakit karat daun kedelai yang disebabkan oleh fungi Phakopsora pachyrhizi Syd. Penyakit ini sukar dikendalikan dengan varietas tahan karena pergeseran ras patogen yang cepat sementara sumber ketahanan kedelai sangat terbatas (Priyatno et al. 2001).
Pengendalian hayati fungi dengan menggunakan mikroorganisme kitinolitik didasarkan pada kemampuan mikroorganisme menghasilkan enzim kitinase dan β -1,3-glukanase yang dapat melisis sel fungi (El-Katatny et al. 2000). Kemampuan kitinolitik bakteri sebagai agens biokontrol patogen dilaporkan oleh Inbar & Chat (1991), yang menggunakan Aeromonas caviae untuk mengendalikan beberapa fungi patogen tanaman. Selain A. caviae beberapa bakteri kitinolitik dilaporkan memiliki potensi sebagai pengendali hayati misalnya P. syringae, Burkholderia cepasia, Bacillus subtilis, Agrobacterium radiobacter, Enterobacter cloacae dan Streptomyces griseoviridis, Actinomycetes, (Fravel et al. 1998; Tahtamouni et al. 2006).
Mikoriza
(Cook & Baker 1996). Mikoriza adalah mikrob yang berpotensi sebagai agens biokontrol terhadap patogen tular tanah.
Fungi mikoriza arbuskular (FMA), memiliki pengaruh yang luas terhadap patogen dan mikrob non-patogenik di dalam tanah. Selain berpotensi dalam pengendalian hayati, juga mampu meningkatkan penyerapan hara esensial terutama fosfor (P) oleh akar tanaman. Selain itu, FMA mampu meningkatkan kandungan klorofil dan zat perangsang tumbuh, sehingga tanaman terhindar dari stres lingkungan terutama saat dipindahkan ke lapangan (Hasanuddin & Gonggo 2004; Widiastuti et al. 2005).
Sharda & Rodrigues (2009) merangkum beberapa hasil penelitian mengemukakan bahwa tanaman yang berasosiasi dengan FMA akan mengalami perubahan dalam morfologi pada akar dan jaringan akar. Terjadi lignifikasi dinding sel akar tanaman inang akan menghambat infeksi patogen akar. Pada tanaman yang bermikoriza, mengandung isoflavonoid lebih tinggi sehingga tanaman lebih tahan terhadap serangan patogen karena senyawa tersebut dapat menghambat pertumbuhan patogen tanah. Perubahan pada unsur kimia dan jaringan tanaman, pengurangan stres abiotik, dan perubahan populasi mikroorganisme pada rizosfir (Artursson et al. 2006). Kontribusi FMA sebagai agens biokontrol penyakit tanaman, terutama patogen tular tanah karena memiliki kemampuan untuk mengeluarkan zat antibiotik (Azcon-Anguilar & Barea 1996) dan atau menghasilkan metabolit sekunder seperti fitoaleksin (Harrison & Dixon 1993) dan senyawa fenolik (Grandmaison et al. 1993).
Imas et al. (1989) mengemukakan bahwa mikoriza dapat berfungsi sebagai pelindung terhadap infeksi patogen akar dengan mekanisme sebagai berikut: (1) adanya selaput hifa (mantel) dapat berfungsi sebagai barier masuknya patogen, (2) mikoriza menggunakan hampir semua kelebihan karbohidrat dan eksudat lainnya, sehingga tercipta lingkungan yang tidak cocok untuk patogen, (3) akar tanaman yang sudah diinfeksi fungi mikoriza, tidak dapat diinfeksi oleh fungi patogen yang menunjukkan adanya kompetisi.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan dari Juni 2009 sampai Juli 2010 di Laboratorium Bakteriologi Tumbuhan dan Mikologi Tumbuhan, dan lahan percobaan Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
1. Eksplorasi Bakteri Kitinolitik
Bakteri kitinolitik diisolasi dari rizosfir tanaman kelapa sawit yang diambil dari PTPN XIV (Sulawesi Selatan) dan PT Astra Agrolestari (Sulawesi Barat). Akar digerus kemudian dilakukan pencawanan untuk setiap seri pengenceran. Sedangkan suspensi tanah dibuat dengan menambahkan 5g tanah ke dalam 50 ml phospat buffer saline, diratakan dengan menggunakan inkubator bergoyang (shaker) dengan kecepatan 150 rpm selama 1 jam, dan dilakukan pencawanan untuk setiap seri pengenceran pada media nutrien agar. Koloni bakteri yang muncul diinokulasikan pada media luria-bertaini (LB) 10% dengan penambahan serbuk kitin 1% (b/v) sebagai induksi awal bakteri kitinolitik dan ditumbuhkan selama ±12 jam. Setelah itu sebanyak 10 µl kultur bakteri diteteskan ke dalam media kitin dan diinkubasi pada suhu ruang. Koloni yang menghasilkan zona bening pada media kitin adalah bakteri kitinolitik yang dipilih untuk pengujian berikutnya yaitu uji patogenitas isolat bakteri dengan metode hipersensitif pada tanaman tembakau. Pengujian ini dilakukan dengan menyuntikan suspensi bakteri pada daun tembakau dengan menggunakan syringe steril. Hasil positif ditunjukkan dengan adanya gejala nekrotik pada daun tembakau yang disuntik dengan suspensi bakteri setelah 24 jam (Schaad et al. 2001).
diangkat-angkat keatas dari tetesan tersebut. Reaksi positif ditunjukkan dengan larutan KOH 3% menjadi kental dan cairan mengikuti jarum ose sampai 0.5-2 cm atau lebih, bila jarum tersebut di angkat. Hal tersebut menunjukan bahwa bakteri uji adalah gram negatif sebaliknya jika slime atau cairan kental tidak berbentuk atau tidak mengikuti jika jarum ose diangkat maka reaksi adalah negatif, dan bakteri diindikasikan bersifat gram positif (Schaad et al. 2001).
2. Uji Indeks Kitinolitik
Koloni bakteri kitinolitik ditumbuhkan pada media LB 10% dengan penambahan serbuk kitin 1%. Setelah itu 10 μl dari masing-masing kultur bakteri yang positif kitnolitik diteteskan pada media kitin dan diinkubasikan selama 7 hari pada suhu ruang. Lebarnya zona bening menunjukan aktifitas kitinase dari masing-masing isolat. Nilai aktivitas hidrolisis secara kualitatif diperoleh dengan membagi diameter zone bening disekitar koloni dengan diameter koloni isolat.
3. Uji Potensi Antagonis Bakteri Kitinolitik terhadap G. boninense
Uji potensi antagonis bakteri kitinolitik terhadap G. boninense dilakukan dengan metode kultur ganda pada media cair dan media padat. Pengujian pada media padat dilakukan di media PDA pada petri yang berdiameter 9 cm. Potongan biakan G. boninense dan suspensi bakteri ditempatkan pada posisi berlawanan masing-masing berjarak 3 cm dari tepi petri. Pengamatan dilakukan setiap hari selama 4 hari. Pengujian pada media cair dilakukan pada media PDB. Inolukum G. boninense dimasukkan dalam media PDB yang mengandung 106cfu/100 ml suspensi bakteri kitinolitik. Sebagai kontrol inolukum G. boninense ditumbuhkan pada media PDB tanpa perlakuan bakteri kitinolitik. Pengamatan dilakukan 14 hsi dengan menimbang berat basah fungi G. boninense yang tumbuh.
Persentase penekanan pertumbuhan patogen pada media cair dan media padat dihitung dengan menggunakan rumus sebagai berikut.
%
tanpa bakteri kitinolitik (kontrol), dan Ro adalah jari-jari koloni patogen yang mengarah ke bakteri kitinolitik atau berat basah patogen yang tumbuh pada media dengan perlakuan bakteri kitinolitik.
Pengujian potensi antagonis bakteri kitinolitik terhadap G. boninense pada media cair dan media padat dilakukan dalam percobaan Rancangan Acak Lengkap (RAL) yang terdiri dari 9 perlakuan dan 3 ulangan. Data hasil pengamatan dianalisis dengan sidik ragam (ANOVA) dan perbedaan nilai tengah antar perlakuan diuji menggunakan uji BNT pada 0,05.
4. Uji Keefektifan Bakteri Kitinolitik dan FMA dalam Menekan Keparahan
Penyakit dan Pengaruh terhadap Pertumbuhan Bibit Sawit
4.1. Penyiapan Substrat Inokulum G. boninense
Isolat fungi G. boninense yang digunakan merupakan koleksi dari Pusat Penelitian Kelapa Sawit (PPKS) Medan, yang diremajakan kembali dengan menggunakan mediaPDA. Pembiakan substrat inokulum G. boninense dilakukan dengan cara potongan batang karet yang berukuran 2 cm x 2 cm x 2 cm direndam selama semalam lalu dimasukkan ke dalam plastik polipropilen tahan panas yang bagian atasnya diberi ring paralon diameter 1 inci dan ditutup kapas, kemudian disterilisasi dengan autoklaf pada suhu 121 oC selama 60 menit. Biakan G. boninense yang berumur 14 hari pada media PDA diinfestasikan pada substrat yang telah steril dan diinkubasi pada suhu ruang selama 5 minggu, hingga mengkolonisasi seluruh bagian substrat, dan selanjutnya dapat langsung digunakan sebagai sumber inokulum di dalam polybag.
4.2. Penyiapan Formula FMA
Formula FMA diperoleh dari Balai Pengkajian Bioteknologi, Badan Pengkajian dan Penerapan Teknologi (BPPT), Pusat pengembangan Ilmu Pengentahuan dan Teknologi (Puspiptek), Serpong dengan kandungan Gigaspora sp. dan Glomus sp.
4.3. Penyiapan Bakteri Kitinolitik
menggunakan inkubator bergoyang (shaker) dengan kecepatan 150 rpm selama ± 12 jam.
4.4. Aplikasi FMA, Bakteri Kitinolitik dan G. boninense pada Bibit Sawit
Bibit kelapa sawit yang telah berumur 3 bulan dipindahkan ke Polybag besar berukuran 25 cm x 30 cm (pre-nursery). Media tanam yang digunakan adalah tanah yang terlebih dahulu difumigasi menggunakan Vapam (Sodium methyldithiocarbamate) selama 10 hari dalam kantong plastik besar (4 g Vapam/50 kg tanah), setelah itu kantong plastik dibuka dan diangin-anginkan selama 3 hari dan siap digunakan sebagai media tanam bibit sawit. Waktu inokulasi bakteri kitinolitik dan FMA dilakukan sesuai dengan perlakuan yang telah ditentukan. Inokulasi FMA baik secara tunggal maupun kombinasi dengan bakteri kitinolitik sebanyak 15 g/polybag dan suspensi bakteri sebanyak 106 cfu/ml dengan dosis 38,33 ml/kg tanah. Inokulasi G. boninense dilakukan dengan cara biakan G boninense sebanyak 2 potong diinokulasi dengan cara kontak langsung pada perakaran kelapa sawit. Tanaman kelapa sawit dipelihara selama 2 bulan, setelah diinokulasi patogen, kemudian dibongkar setiap unitnya untuk diamati pada bagian akar dan pangkal batang.
Pengujian dilakukan dengan dengan menggunakan petak terpisah yang terdiri atas 20 perlakuan dan 5 ulangan. Data hasil pengamatan dianalisis dengan sidik ragam (ANOVA) dan perbedaan nilai tengah antar perlakuan diuji menggunakan uji BNT 0,05. Susunan perlakuaan pada penelitian ini adalah sebagai berikut:
Tabel 1 Perlakuan uji pada bibit sawit
Kode perlakuan Keterangan: Mo= tanpa FMA, M1= dengan FMA, Bo= tanpa bakteri, B1= isolat
4.4.1. Pengamatan terhadap Tingkat Keparahan Penyakit BPB
Tingkat keparahan penyakit (KpP) diukur berdasarkan nekrotik yang terjadi pada akar. Keparahan penyakit dihitung menggunakan rumus Townsen & Hüberger dalam Ariffin et al. (2000)
dimana KpP adalah persen keparahan penyakit, ni adalah jumlah tanaman dalam skor yang sama, vi adalah nilai skor penyakit, Z adalah nilai skor tertinggi yang digunakan, dan N adalah jumlah tanaman yang diamati. Metode pemberian skor skala 0 hingga 4 digunakan untuk menentukan persen KpP.
Tabel 2 Skoring bercak nekrotik akar berdasarkan persentase nekrotik akar kelapa sawit
Nilai Skor Nekrotik akar (%) Keterangan
0 0 ≤ x < 1 Tidak ada nekrotik atau nekrotik bukan
4.4.2 Pengamatan terhadap Berat Basah Akar, Tajuk, dan Tinggi Bibit Sawit
Penentuan berat akar kelapa sawit dilakukan secara kuantitatif yaitu dengan menimbang akar kelapa sawit yang telah dicuci bersih menggunakan timbangan digital, sedangkan tinggi tajuk diukur secara manual menggunakan mistar berukuran 100 cm.
4.4.3 Tingkat Asosiasi FMA pada Akar Bibit Sawit
jam, perlakuan ini sama dengan pemanasan pada suhu 121 oC selama 5 menit di dalam autoklaf. Akar yang masih berwarna gelap dimasukkan ke dalam larutan alkalin hydrogen peroksida 0.5% H2O2 (v/v) selama 10 menit, selanjutnya direndam dalam larutan 1% HCl (v/v) selama 10 menit, dan diwarnai menggunakan larutan trypan blue 0.05% (w/v) di dalam larutan lactid acid glycerol yang merupakan campuran asam laktat, gliserol, dan air dengan perbandingan 1:1:1 (v/v), lalu dipanaskan pada suhu 121 oC selama 15 menit, kemudian dicuci dibawah air mengalir hingga bersih sebelum dipindahkan ke dalam larutan yang berbeda. Akar yang telah diwarnai disimpan di dalam larutan 50 % gliserol (v/v) (Brundett et al. 1996).
Infeksi akar oleh FMA dihitung menggunakan prosedur the gridline intersection method (Brundett et al. 1996). Akar yang telah diwarnai disebar secara acak pada cawan petri berdiameter 14 cm yang telah diberi kotakan berukuran 0.8 cm x 0.8 cm sebanyak 100 kotak. Potongan akar yang melewati gridline dihitung sebagai infeksi, apabila ditemukan struktur mikoriza (spora, hifa, vesikel, atau arbuskula) pada jaringan yang ditandai dengan bagian berwarna lebih gelap dibandingkan dengan bagian lainya. Pengamatan dilakukan menggunakan mikroskop binokuler dengan cara mengamati seluruh kotakan gridline secara vertikal dan horizontal (Newman 1996; Tennant 1975; Giovannetti & Mosse 1980 dalam Brundett et al. 1996). Tingkat asosiasi mikoriza dihitung menggunakan rumus berikut :
olonisasi mikori a akar terinfeksi mikori aakar interseksi
Aktivitas enzim Peroksidase
Aktifitasenzim peroksidase (POD) pada akar tanaman diukur menggunakan menggunakan spektrofotometer. Ekstraksi dan kuantifikasi enzim POD dilakukan pada akhir percobaan menurut metode Karthikeyan et al. (2005) yang dilakukan di Laboratorium Rekayasa Bioproses Pusat Antar Universitas (PAU) IPB.
Sampel akar yang akan dianalisis dari setiap perlakuan terlebih dahulu dihomogenkan menggunakan nitrogen cair, kemudian sebanyak 1 gr sampel ditambah 2 ml 0.1M buffer fosfat pH 7.0 pada suhu 4 oC lalu disentrifugasi selama 20 menit pada kecepatan 10000 rpm dan sepernatannya digunakan sebagai sumber enzim yang akan dianalisis.
HASIL DAN PEMBAHASAN
Eksplorasi Bakteri kitinolitik
Hasil eksplorasi bakteri kitinolitik dari rizosfir kelapa sawit di beberapa lokasi perkebunan kelapa sawit Sulawesi Barat dan Sulawesi Selatan diperoleh 77 isolat bakteri. Diantara 77 isolat bakteri yang diperoleh, 7 isolat bakteri positif bersifat kitinolitik. Aktifitas kitinolitik ditunjukkan dengan terbentuknya zona hidrolisis/zona bening di sekitar koloni bakteri pada medium yang mengandung koloidal kitin pada 7 hari setelah inkubasi (hsi) (Gambar 1).
Gambar 1 Isolat bakteri positif kitinolitik dengan zona bening pada media koloidal kitin, tanda panah menunjukkan koloni bakteri, dan tanda panah kiri kanan menunjukkan zona bening aktivitas enzim kitinase yang dihasilkan bakteri.
Pleban et al. (1997) melaporkan bahwa zona bening yang terbentuk di sekitar koloni bakteri menandakan adanya aktivitas kitinolitik di medium pertumbuhan yang mengandung kitin. Untuk mengetahui produksi kitinase dari suatu bakteri kitinolitik dapat dilihat dari perubahan warna medium menjadi lebih transparan dengan terbentuknya zona bening disekeliling koloni bakteri. Warna medium transparan disebabkan oleh terdegradasinya kitin pada media tumbuh karena adanya aktivitas enzim kitinase yang dihasilkan oleh bakteri ke medium. Kitinase merupakan enzim ekstraseluler yang berperan penting dalam menghidrolisis kitin, enzim ini dihasilkan di dalam sel bakteri, tetapi dikeluarkan ke dalam medium tumbuhnya (Tsujibo et al. 1996; Wijaya 2002; Dewi 2008).
B13 B14 B16 B42
Hasil pengujian gram mengunakan KOH 3% menujukkan bahwa dari ketujuh isolat uji hanya isolat B16 yang gram positif, sedangkan isolat lainnya (B13, B14, B42, AL11, TB41, dan L34) adalah gram negatif. Hasil uji patogenesitas dengan metode hipersensitif pada tanaman tembakau menunjukkan bahwa ketujuh isolat bakteri kitinolitik bereaksi negatif (tidak menimbulkan nekrotik). Berdasarkan hasil ini, dapat disimpulkan bahwa ketujuh isolat bakteri uji tidak fitopatogenik.
Evaluasi Potensi Antagonisme dan Indeks Kitinolitik
Hasil uji potensi antagonisme 7 isolat bakteri kitinolitik terhadap G. boninense dengan metode kultur ganda pada media cair (PDB) dan media padat (PDA) menunjukkan hasil yang berbeda. Pada media cair, ketujuh isolat bakteri uji memiliki kemampuan menekan pertumbuhan G. boninense yang tinggi, yaitu lebih dari 95%. Pada media padat hanya isolat AL11, TB41, dan L34 yang menunjukkan penekanan yang relatif tinggi yaitu 52%, 44%, dan 41%. Sedangkan isolat B14, B13, B16, dan B42 hanya mampu menghambat pertumbuhan G. boninense kurang dari 10% (Gambar 3 dan Tabel 3).
Gambar 2 Penekanan pertumbuhan G. boninense oleh bakteri kitinolitik pada media cair (atas), tanda panah menunjukkan pertumbuhan koloni G. boninense, dan padat (bawah), tanda garis menunjukkan zona penghambatan.
Hasil analisis statistik menunjukkan bahwa pada media cair, persentase penghambatan semua isolat bakteri berbeda nyata dengan perlakuan kontrol. Kontrol Isolat B14 Isolat B42 Isolat Al11 Isolat TB41
Namun, persentase penghambatan antar masing-masing isolat bakteri tidak berbeda nyata. Sedangkan pada media padat semua isolat juga menunjukkan persentase penghambatan yang berbeda nyata dengan perlakuan kontrol, kecuali isolat B42 (Tabel 3).
Tabel 3 Persentase penghambatan pertumbuhan G. boninense dengan perlakuan bakteri kitinolitik pada media padat 4 hsi dan media cair 14 hsi
Perlakuan Persentase penghambatan (%)
Keterangan: Huruf yang sama dibelakang angka pada kolom yang sama menunjukan tidak berbeda nyata berdasar uji BNT α0.05
Perbedaan kemampuan penekanan pertumbuhan G. boninense pada media cair dan padat diduga karena adanya perbedaan mekanisme antagonisme dominan yang berperan pada media padat dan media cair. Pada media padat mekanisme antagonisme dominan yang berperan adalah enzimatis sedang pada media cair kemungkinan ada mekanisme senyawa atau aktivitas lain yang terlibat dalam proses antagonisme dari bakteri kitinolitik.
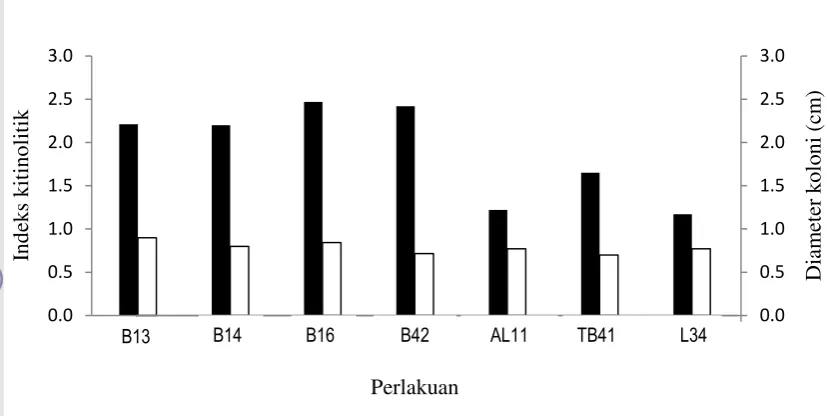
Hasil pengukuran indeks kitinolitik 7 hsi menunjukkan aktivitas kitinolitik bahwa masing-masing bakteri berbeda-beda. Nilai indeks kitinolitik tertinggi ditunjukkan pada isolat B16 dan B42, diiukiti isolat B13 dan B14. Indeks kitinolitik yang lebih rendah ditunjukkan oleh isolat TB41, AL11, dan L34. Besarnya indeks kitinolitik ini tidak berhubungan dengan kecepatan tumbuh masing-masing isolat bakteri (Gambar 2).
Gambar 3 Indeks kitinolitik 7 isolat bakteri pada 7 hsi ( ) dan pertumbuhan koloni bakteri kitinolitik ( ).
Hasil percobaan menunjukkan bahwa kemampuan menghidrolisis kitin dari 7 isolat bakteri uji berbeda-beda. Hal ini mungkin disebabkan jenis subtrat kitin yang digunakan. Menurut Joo 2005, Gohel et al. 2006, dan Brzezinska et al. 2007, kemampuan menghidrolisis kitin dari setiap strain bakteri kitinolitik salah satunya dipengaruhi oleh susbtrat kitin yang digunakan. Menurut Joo (2005), aktivitas kitinolitik Streptomyces halstedii lebih tinggi terjadi pada medium koloidal kitin yang mengandung campuran kitobiose, kitotriose, kitotetrose, dan kitopentose.
Dari hasil pengujian indeks kitinolitik pada media koloidal kitin dan pengujian potensi antogonisme pada media padat dapat dilihat bahwa nilai indeks kitinolitik yang tinggi tidak selalu diikuti dengan potensi antagonis yang tinggi terhadap G. boninense. Isolat bakteri AL11, TB41, dan L34 walaupun pada pengujian indeks kitinolitik menunjukan nilai indeks kitinolitik yang rendah namun pada pengujian kemampuan antagonis pada media padat ketiga isolat tersebut menunjukkan kemampuan antagonis yang lebih tinggi dibanding isolat lain.
Berdasarkan hasil uji antagonisme in vitro, hanya dipilih 2 isolat bakteri untuk evaluasi keefektifannya mengendalikan BPB pada bibit kelapa sawit yaitu isolat TB41 dan AL11. Pemilihan kedua isolat tersebut untuk dievaluasi keefektifannya dalam mengendalikan BPB pada bibit kelapa sawit karena kedua isolat tersebut menunjukkan kemampuan menghambat pertumbuhan G. boninense yang konsisten tinggi baik pada media cair maupun media padat.
Pengaruh Aplikasi Bakteri Kitinolitik dan FMA terhadap BPB
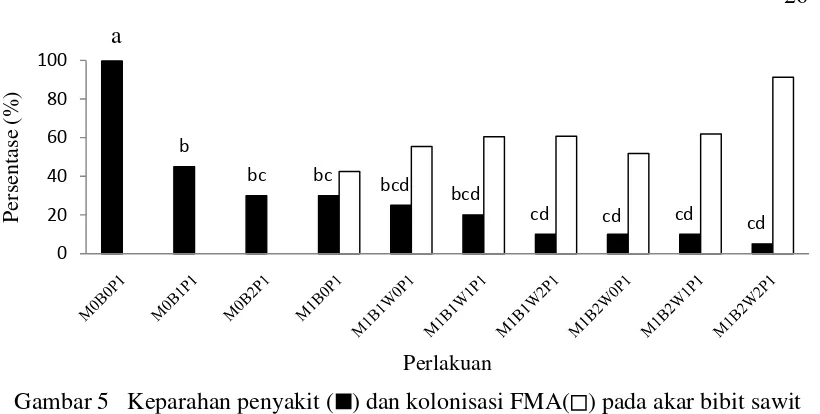
Hasil pengamatan menujukkan bahwa pada akar bibit sawit yang diinokulasi patogen terdapat gejala nekrotik berwarna coklat kehitaman. Nekrotik ini meluas seiring dengan perkembangan penyakit. Sedangkan semua tanaman tanpa inokulasi patogen tidak menunjukkan adanya gejala nekrotik pada akar. Hal ini terjadi karena pada perlakuan tanpa inokulasi patogen (kontrol) memang tidak ada inokulum patogen yang dapat menimbulkan infeksi pada bibit sawit. Adanya aplikasi bakteri kitinolitik, FMA, dan kombinasinya menurunkan pesentase akar bibit sawit yang mengalami nekrotik. Contoh akar yang mengalami nekrotik dan penilaian keparahan penyakit disajikan pada Gambar 4.
C
rapuh. Jaringan korteks berubah warna dari putih menjadi coklat kehitaman, dan mudah didisintegrasi. Pada serangan berat stele menjadi kehitaman, kadang-kadang bagian permukaan sebelah dalam endodermis ditemukan hifa berwarna keputihan yang merupakan tanda penyakit.
Dalam percobaan ini, hingga 2 bulan setelah inokulasi belum ditemukan gejala busuk pada pangkal batang. Akan tetapi, gejala nekrotik pada akar yang merupakan gejala awal adanya infeksi G. boninense sudah dapat digunakan sebagai indikator perkembangan BPB. Yulianti (2001), melaporkan bahwa inokulasi G. boninense selama 5 bulan belum menunjukkan gejala BPB pada pangkal batang dan daun. Hal yang sama juga dikemukakan oleh Sinaga et al. (2003) dan Risanda (2008), bahwa periode inkubasi G. boninense untuk dapat menimbulkan gejala pada tajuk tanaman kelapa sawit cukup lama. Menurut Sinaga et al. (2003), bahwa inisiasi gejala nekrotik pada daun dan pangkal batang muncul pada bulan ketujuh setelah inokulasi G. boninense.
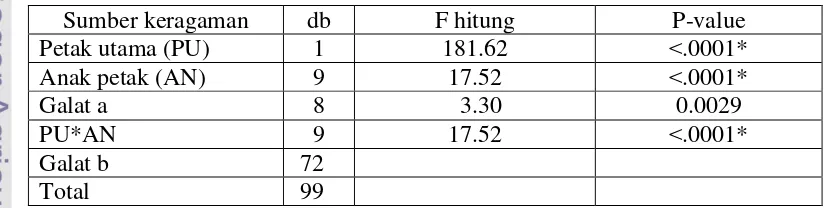
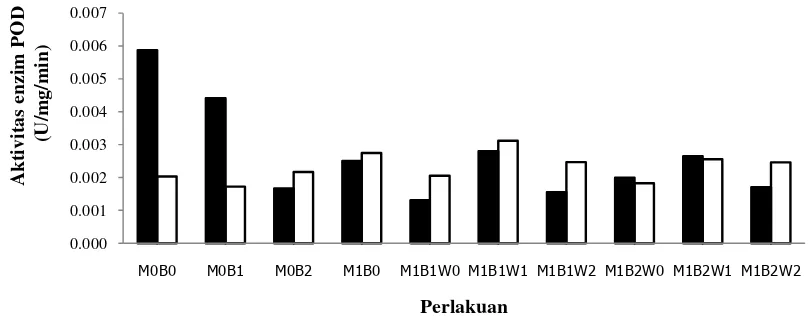
Hasil analisis statitistik menunjukkan bahwa perlakuan aplikasi bakteri kitinolitik dan FMA secara tungal dan kombinasinya berpengaruh nyata pada keparahan penyakit (Tabel 4). Hasil uji lanjut menunjukkan bahwa persen keparahan penyakit semua perlakuan berbeda nyata dengan kontrol (inokulasi G. boninense), sedangkan antar perlakuan aplikasi bakteri kitinolitik, FMA, dan kombinasinya bervariasi (Tabel 5). Perlakuan kombinasi antara bakteri kitinolitik dan FMA menunjukkan penekanan keparahan penyakit yang lebih tingi dibandingkan dengan perlakuan tunggal isolat bakteri kitinolitik dan FMA (Tabel 5 dan Gambar 5).
Keterangan: Tanda bintang (*) dibelakang nilai P-value menunjukkan berpengaruh nyata
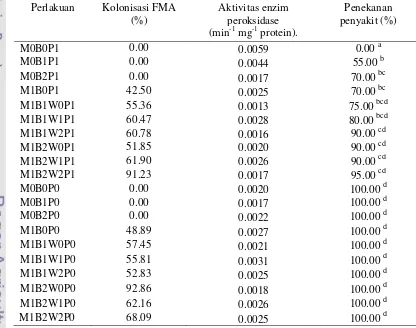
kitinolitik menghasilkan kolonisasi FMA dan persentase penekanan keparahan penyakit yang lebih tinggi dibandingkan perlakuan tunggal. Aplikasi patogen tidak mempengaruhi kolonisasi FMA pada akar. Aktivitas enzim peroksidase pada perlakuan kombinasi antara FMA dan bakteri kitinolitik tidak selalu lebih tinggi dibanding pada perlakuan tunggal, baik dengan ataupun tanpa inokulasi G. boninense. Aktivitas enzim peroksidase yang tinggi tidak selalu diikuti dengan ketahanan tanaman terhadap G. boninense. Pada bibit sawit yang aplikasikan G. boninense, waktu aplikasi bakteri kitinolitik (isolat TB41) dan FMA juga berpengaruh terhadap kemampuan menekan keparahan penyakit. Perlakuan FMA terlebih dulu memberikan hasil yang lebih baik dalam menekan keparahan penyakit, namun berbeda dengan isolat bakteri AL11 perbedaan waktu aplikasi bakteri kitinolitik dan FMA menunjukkan kemampuan yang sama dalam menekan keparahan penyakit (Tabel 5).
Tabel 5 Persentase kolonisasi FMA, aktivitas enzim peroksidase, dan persentase penekanan penyakit pada akar bibit sawit pada berbagai perlakuan
Perlakuan Kolonisasi FMA
b yang diinokulasi G. boninense.
Keterangan: Huruf yang sama diatas bar menunjukan tidak berbeda nyata berdasar uji BNT α0.05
Hasil percobaan ini membuktikan bahwa antara FMA dan isolat bakteri (TB41 dan AL11) saling kompatibel dan sinergis menekan. Hal ini dapat dilihat dari persentase penekanan keparahan penyakit dan kolonisasi FMA pada akar bibit sawit yang lebih tinggi pada perlakuan kombinasi dibanding pada perlakuan tunggal. Menurut Barea et al. (2005), beberapa bakteri yang diisolasi dari rizosfir dapat membantu perkembangan dan stabilitas FMA. Citernesi et al. (1996) dan Budi et al. (1999) melaporkan hubungan bakteri rizosfir dengan tanaman yang dikolonisasi oleh FMA menunjukkan asosiasi yang kompatibel. Beberapa bakteri dilaporkan berasosiasi dengan FMA (Bianciotto et al. 2003). Beberapa bakteri juga telah ditemukan terkait dengan struktur hifa eksternal FMA (Toljander et al. 2006) dan spora atau dinding spora (Roesti et al. 2005). Bakteri yang berasosiasi dengan FMA dapat meningkatkan potensi biokontrol dari FMA dengan menghambat pertumbuhan miselium, produksi sporangium, dan perkecambahan zoospore fungi patogen (Budi et al. 1999) serta memperbaiki struktur tanah (Andrade et al. 1997). Asosiasi antara bakteri dan FMA juga dapat meningkatkan perkecambahan spora FMA (Walley & Germida 1996; Bianciotto & Bonfante 2002; Hildebrandt et al. 2002; Xavier & Germida 2003).
Pada percobaan ini adanya aplikasi bakteri kitinolitik selain meningkatkan persen akar yang dikolonisasi oleh FMA, juga meningkatkan persen penekanan keparahan penyakit pada akar bibit sawit. Hasil ini sejalan dengan penelitian Cruz et al. (2008) yang menyatakan bahwa aktivitas biokontrol yang dilakukan
oleh FMA di dalam tanah terutama di daerah rizosfir berkaitan dengan bakteri yang berasosiasi dengan FMA. Pada penelitian ini juga terlihat bahwa semakin tinggi persen kolonisasi FMA pada akar, penekanan terhadap penyakit juga semakin tinggi. Hal yang sama dilaporkan oleh Khaosaad et al. (2007), bahwa pengaruh sistemik dari kolonisasi akar oleh FMA Glomus mosseae terhadap infeksi Gaeumannomyces graminis var. tritici (GGT) berkurang ketika tanaman barlei menunjukkan tingkat kolonisasi mikoriza yang tinggi.
Meskipun perlakuan kombinasi antara FMA dan bakteri kitinolitik menunjukkan hasil persentase penekanan penyakit yang lebih baik namun perlakukan tunggal bakteri kitinolitik dan FMA juga mampu menekan persentase keparahan penyakit BPB. Bakteri kitinolitik telah banyak digunakan sebagai agens biokontrol yang potensial karena kemampuannya untuk melisis dinding sel fungi patogen dan kemampuannya memanfaatkan hifa fungi sebagai substrat pertumbuhan (De Boer et al. 2001). Penelitian pemanfaatan bakteri kitinolitik untuk pengendalian penyakit tumbuhan diantaranya diaporkan oleh Ordentlich et al. (1987) yang melaporkan bahwa kultur filtrat bakteri kitinolitik Serratia marcescens efektif sebagai agens biokontrol terhadap Sclerotium rolfsii. Sedangkan Singh et al. (1999) melaporkan bahwa Paenibacillus spp. dan Streptomyces spp. mampu menekan layu Fusarium pada mentimun.
rolfsii, dan (Suryanto et al. 2010) melaporkan bakteri kitinolitik sebagai agens biokontrol layu Fusarium pada cabai merah.
Menurut De Boer et al. (2001) dan Gohel et al. (2006), bakteri kitinolitik memiliki beberapa mekanisme dalam menekan patogen. Dalam menekan fungi, bakteri dapat melisis hifa fungi atau menyebabkan dinding sel fungi menjadi tidak stabil. Bakteri kitinolitik akan memanfaatkan hifa fungi sebagai substrat untuk pertumbuhannya. Selain itu bakteri ini juga dapat bersimbiosis dengan akar tanaman dan menghasilkan kitinase yang berperan sebagai pertahanan diri bagi tanaman dalam melawan patogen.
Hasil penelitian tentang pemanfaatan FMA untuk pengendalian patogen telah banyak dilaporkan antara lain oleh Cordier et al. (1998) dan Pozo et al. (2002) melaporkan bahwa aplikasi FMA (G. mosseae) efektif mengendalikan Phytophthora parasitica pada tanaman tomat, Hashim (2003) melaporkan bahwa aplikasi mikoriza efektif menekan kematian akibat G. boninense pada kelapa sawit, Khaosaad et al. (2007) melaporkan bahwa aplikasi FMA secara sistemik mampu menekan Gaeumannomyces graminis var. tritici pada tanaman barlei, dan Ozgonen et al. (2010) bahwa FMA efektif untuk pengendalian busuk batang yang disebabkan oleh S.rolfsii pada kacang tanah.
Secara umum peran FMA dalam pengendalian patogen tanaman adalah adanya perubahan pada anatomi dan morfologi akar yang berasosiasi dengan FMA, perubahan populasi mikrob disekitar rizosfir tanaman yang berasosiasi dengan FMA, dan pengaktifan mekanisme pertahanan tanaman oleh adanya asosiasi dengan FMA (Azcón-Aguilar & Barea 1996). Sedangkan secara spesifik misalnya adanya aktivitas enzim-enzim yang berperan dalam biosintesis lignin (peroksidase dan polifenol oksidase) maupun enzim hidrolitik (kitinase dan glukanase), adanya pembentukan struktur seperti papila maupun deposisi material seperti kalus terutama pada tempat terjadinya infeksi patogen (Cordier et al. 1998; Pozo et al. 2002; Lee et al. 2005; Zheng et al. 2005).
0.000
M0B0 M0B1 M0B2 M1B0 M1B1W0 M1B1W1 M1B1W2 M1B2W0 M1B2W1 M1B2W2
Ak
kombinasi FMA dan bakteri kitinolitik tidak selalu lebih tinggi dibanding perlakuan tunggal (Gambar 6). Hasil pengukuran aktivitas enzim peroksidase tidak menunjukkan pola yang jelas, walaupun demikian diduga bahwa sebenarnya pembentukan lignin pada akar telah terjadi. Pola aktivitas enzim yang tidak jelas tersebut mungkin disebabkan karena analisis aktivitas ezim peroksidase dilakukan pada akhir percobaan (2 bulan setelah inokulasi patogen). Dari beberapa hasil penelitian pada umumnya mengukuran aktivitas enzim peroksidase dilakukan maksimal satu minggu setelah inokulasi petogen.
Gambar 6 Aktivitas enzim peroksidase pada perakaran bibit sawit ’tanpa
patogen’ ( ) dan ’dengan patogen’ ( ) pada berbagai perlakuan.
Peroksidase merupakan kelompok PR-protein dari golongan PR-9. Aktivitas peroksidase berhubungan dengan resistensi penyakit pada tanaman (Lin & Kao 2001; Ray etal. 1998), dan meningkat pada tanaman inang setelah infeksi patogen (Borden & Higgins 2002; Koike et al. 2001; Zheng et al. 2005). Peroxidase adalah enzim yang berpartisipasi dalam proses oksidasi reduktif polisakarida dalam dinding sel, seperti oksidasi fenol, suberisasi, dan lignifikasi sel tanaman inang (Ray et al. 1998). Peroksidase dilaporkan berperan sebagai katalis dalam tahap akhir biosintesis lignin dan hidrogen peroksida (Breusegem et al. 2001).
Menurut Breusegem et al. (2001) dan Lin & Kao (2001) resistensi tanaman terhadap patogen dikaitkan dengan induksi peroksidase dalam jaringan inang. Demikian juga Gupta et al. (1990) dan Tenaya et al. (2001) menyimpulkan dari
sebagai penanda seleksi ketahanan terhadap penyakit. Namun menurut Chen et al. (2000) dan Mohammadi & Kazemi (2002) selain peroksidase, akumulasi lignin dan senyawa fenolik juga berhubungan dengan resistensi tanaman dalam interaksinya dengan patogen tanaman, seperti Fusarium graminearum pada gandum dan Pythium aphanidermatum pada ketimun.
Seevers et al. (1971) mengemukakan bahwa peningkatan peroksidase dapat dilihat hanya sebagai sebuah gejala biokimia dan peristiwa lainnya yang sebelumnya menyebabkan resistensi. Kenyataan bahwa peningkatan peroksidase tertentu dapat memberikan beberapa informasi tentang kejadian biokimia sebelumnya atau berhubungan dengan pengembangan resistensi.
Pengaruh Aplikasi Bakteri Kitinolitik dan FMA terhadap Pertumbuhan Bibit Sawit
Hasil pengukuran kebugaran tanaman (berat basah akar, tajuk, dan tinggi tanaman) disajikan pada Tabel 6. Hasil pengamatan menunjukkan bahwa adanya aplikasi FMA dan bakteri kitinolitik meningkatkan berat tajuk dan persen penambahan tinggi tanaman walaupun ada inokulasi G. boninense. Pengaruh aplikasi FMA dan bakteri kitinolitik terhadap kebugaran tanaman ini diduga karena adanya peningkatan penyerapan hara oleh tanaman terutama yang diaplikasi FMA. Selain itu, kemampuan FMA dan bakteri kitinolitik dalam menghambat dan menekan perkembangan patogen juga faktor lain yang dapat meningkatkan kebugaran tanaman, terutama pada tanaman yang diinokulasi dengan patogen.
peningkatan percabangan akar, sekaligus membentuk pertahanan terhadap infeksi patogen.
Tabel 6 Pengaruh perlakuan bakteri kitinolitik dan FMA terhadap kebugaran tanaman berdasarkan berat basah akar dan tajuk serta tinggi tanaman
Perlakuan Berat Tajuk (g) Berat Akar (g) Penambahan Tinggi Tajuk
(%)
M0B0P1 20.90 a 8.82 abcdf 20.76 a
M0B1P1 22.10 ab 7.26 ab 33.85 bc
M0B2P1 22.45 abc 7.74 abcd 32.18 abc
M1B0P1 25.17 abcdefg 8.38 abcde 39.68 bc
M1B1W0P1 25.01 abcdefg 8.50 abcde 31.73 abc
M1B1W1P1 26.95 cdefg 8.56 abcdef 30.84 abc
M1B1W2P1 29.05 fg 9.55 cdef 28.64 ab
M1B2W0P1 25.59 abcdefg 7.93 abcde 41.46 bc
M1B2W1P1 26.43 bcdefg 8.02 abcde 43.33 c
M1B2W2P1 27.54 defg 9.39 bcdef 30.50 abc
M0B0P0 29.71 g 10.07 ef 36.58 bc
M0B1P0 23.02 abcd 6.91 a 36.22 bc
M0B2P0 23.35 abcde 8.87 abcdef 33.26 abc
M1B0P0 28.19 efg 10.77 f 33.42 abc
M1B1W0P0 26.83 bcdefg 8.54 abcdef 35.93 bc
M1B1W1P0 28.12 defg 8.37 abcde 35.48 bc
M1B1W2P0 25.77 bcdefg 7.37 abc 33.41 abc
M1B2W0P0 25.59 abcdefg 8.28 abcde 42.56 c
M1B2W1P0 26.94 cdefg 7.06 a 37.56 bc
M1B2W2P0 28.66 def 9.88 def 30.96 abc
SIMPULAN DAN SARAN
Simpulan
1. Hasil eksplorasi bakteri dari rizosfir sawit diperoleh 7 isolat bakteri kitinolitik, 2 diantaranya (isolat TB41 dan AL11) memiliki kemampuan in vitro yang konsisten menekan pertumbuhan Ganoderma boninense,
2. Aplikasi dalam bakteri kitinolitik (isolat TB41 atau AL11) yang dikombinasikan dengan FMA meningkatkan kolonisasi FMA pada akar bibit sawit dan menunjukkan penekanan keparahan penyakit yang lebih tinggi mencapai 95% dibandingkan dengan perlakuan tunggal bakteri kitinolitik atau FMA saja,
3. Bakteri kitinolitik isolat TB41 dan AL11 bersifat sinergis dengan FMA dalam membentuk pertahanan tanaman terhadap serangan G. boninense .
Saran
DAFTAR PUSTAKA
Abadi AL. 1987. Biologi Ganoderma boninense Pat. pada kelapa sawit (Elaeis guineensis Jacq.) dan pengaruh beberapa mikroba tanah antagonistik terhadap pertumbuhannya [disertasi]. Bogor: Fakultas Pascasarjana, Institut Pertanian Bogor.
Adams DJ. 2004. Fungal cell wall of chitinases and glucanases. Microbiol 150:2029-2035.
Andrade G, Mihara KL, Linderman RG, Bethlenfalvay GJ. 1997. Bacteria from rhizosphere and hyphosphere soils of different arbuscular-mycorrhizal fungi. Plant Soil 192:71-79.
Agrios GN. 2005. Plant Pathology. Ed ke-5. New York: Elsevier Academic Pr. Ariffin D, Idris AS, Singh G. 2000. Status of Ganoderma in oil palm. Di dalam:
Flood J, Bridge PD, Holderness M, editor. Ganoderma Diseases of Perrennial Crops. UK: CABI Publishing. hlm 49-68
Artursson V, Flnlayy RD, Jansson JK. 2006. Interaction between arbuscular fungi and bacteria and their potential for stimulating plant growth. Environ microbiol 8:1-10.
Azcon-Aguilar C, Barea JM. 1996. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens an overview of the mechanisms involved. Mycorrhiza 6:457-464.
Azizah H, George ST, Christine ZA. 1990. Interactions of mycorrhizal fungi with root pathogen of cocoa. Curr Trend in MycorrhizalRes. 9:78-79. Barea JM, Pozo MJ, Azcon RC. 2005. Micobial co-operation in the rhizosphere. J
Exp Bot 56:1761-78.
Bianciotto V, Bonfante P. 2002. Arbuscular mycorrhizal fungi: A specialised niche for rhizospheric and endocellular bacteria. Antonie van Leeuwenhoek 81: 365-371.
Bianciotto V, Lumini E. Bonfante P, Vandamme P. 2003. ‘Candidatus Glomeribacter gigasporarum’ gen. nov., sp. nov., an endosymbiont of arbuscular mycorrhizal fungi. Int J Syst Evol Microbiol 53:121-124.
Borden S, Higgins VJ. 2002. Hydrogen peroxide plays a critical role in the defense response of tomato to Cladosporicem fulvum. Physiol Mol Plant Pathol 61:227-236.
Breusegem FV, Vranova E, Dat JF, Inze D. 2001. The role of active oxygen species in plant signal transduction. Plant Sci 161:405-414.
Brzezinska BM, Lalke PE, Donderski W. 2007. Chitinolytic activity of bacteria and fungi isolated from shrimp exoskeletons. Oceanol Hydrobiol. 36:101-111.
Budi SW, Van Tuinen D, Martinotti G, Gianinazzi S. 1999. Isolation from the Sorghum bicolar mycorrhizosphere of a bacterium compatible with arbuscular mycorrhiza development and antagonistic toward soilborne fungal pathogens. Appl Environ Microbiol 65:5148-5150.
Chen CQ, Belanger RR, Benhamou N, Paulitz TC. 2000. Defense enzymes induced in cucumber roots by treatment with plant growth-promoting rhizobacteria (PGPR) and Pythium aphanidermatum. Physiol Mol Plant Pathol 56:13-23.
Chernin L, Ismailov Z, Haran S, Chet I. 1995. Chitinolytic Enterobacter agglomerans antagonistic to fungal plant pathogens. Appl Environ Microbiol 61:1720-1726.
Chung WC, Huang JW, Huang HC. 2005. Formulation of a soil biofungicide for control of damping-off of Chinese cabbage (Brassica chinensis) caused by Rhizoctonia solani. Biol Control 32:287-294.
Citernesi AS, Fortuna P, Filippi C, Bagnoli G, Giovannetti M. 1996. The occurrence of antagonistic bacteria in Glomus mosseae pot cultures. Agron 16:671-677.
Cook RJ, Baker KF. 1996. The Nature and Practice of Biological Control of Plant Pathogens. Minnesota: APS Pr.
Cordier C, Pozo MJ, Barea JM, Gianinazzi S, Gianinazzi-Pearson V. 1998. Cell defense responses associated with localized and systemic resistance to Phytophthora parasitica induced in tomato by an arbuscular mycorrhizal fungus. Mol Plant-Microbe Interact 11:1017-1028.
Cruz AF, Horii S, Ochiai A, Yasuda A, Ishii A. 2008. Isolation and analysis of bacteri associated with spore of Gigaspora margarita. Appl Microbiol 104:1711-7.
De Boer W, Klein Gunnewiek PJA, Kowalchuk GA, Veen VJA. 2001. Growth of chitinolytic dune soil β-subclass Proteobacteria in response to invading fungal hyphae. Appl Environ Microbiol 67:3358-3362.
Dewi IM. 2008. Isolasi bakteri dan uji aktivitas kitinase termofilik kasar dari sumber air panas Tinggi Raja, Simalungan Sumatera Utara [tesis]. Medan: Pascasarja Universitas Sumatera Utara.
Edwards SG, J. Peter, Young W, Fitter AH. 1998. Interactions between Pseudomonas fluorescens biocontrol agents and Glomus mosseae, an arbuscular mycorrhizal fungus, within the rhizosphere. FEMS Microbiol Lett 166:297-303
El-Tarabily KA et al. 2000. Biological control of Sclerotinia minor using a chitinolytic bacterium and Actinomycetes. Plant Pathol 49:573-583.
Fravel DR, Onmic JR. Lewis JA. 1998. Formulation of microorganisms to contol plant disease. Di dalam Burges, editor. Formulation of Biopesticed. Dodecht: Kluwer Academic Pr. Hlm 187-202.
Gohel V, Anil S, Maisuria V, Phadnis A, Chhatpar HS. 2006. Bioprospecting and antifungal potential of chitinolytic microorganisms. African Journal of Biotechnol 5:54-72.
Gupta SK, Gupta PP, Yadava TP, Kaushik CD. 1990. Metabolic change in mustard due to Alternaria leaf blight. Indian Phytopathol 43:64-69.
Grandmaison J, Olah GM, V Calsteren, MR, Furlan V. 1993. Characterization and localization of plant phenolics likely involved in pathogen resistance expressed by endomycorrhizal roots. Mycorrhiza 3:155-164.
Handayani. 2004. Isolasi dan karakterisasi kitinase akar tusan (Pinus mercusii Jungh. et De Vriese) yang bersimbiosis dengan fungi ektomikoriza [tesis]. Yogyakarata: Program Pascasarjana, Universistas Gadjah Mada.
Harrison MJ, Dixon RA. 1993. Isoflavonoid accumulation and expression of defence gene transcripts during the establishment of vesicular-arbuscular mycorrhizal associations in roots of Mediacago tranculata. Mole Plant Microbe Interac 6:643-654.
Hasanuddin, Gonggo B. 2004. Pemanfaatan mikrobia pelarut fosfat dan mikoriza untuk perbaikan fosfor tersedia, serapan fosfor tanah (ultisol), dan hasil jagung (pada ultisol). J Ilmu-ilmu Pertanian Indones 6:8-13.
Hashim A. 2003. Ganoderma versus mycorrhiza. Oil Palm Bull 47:6-14.
Hildebrandt U, Janetta K, Bothe H. 2002. Towards growth of arbuscular mycorrhizal fungi independent of a plant host. Appl Environ Microbiol 68:1919-1924.
Ho YW, Nawawi A. 1985. Ganoderma boninense Pat. basal stem rot of oil palm (Elaeis guinensis) in Paninsular Malaysia. Pertanika 8;425-428.
Idris AS, Kushairi S, Ismail S, Arifin D. 2004. Selection for partial resistance in oil palm progenies to Ganoderma basal stem rot. J Oil Palm Res 16:12-18 Imas T, Hadioetomo RS, Gunawan AW, Setiadi Y. 1989. Mikrobiologi Tanah II.
Depdikbud Ditjen Dikti, Pusat Antar Universitas Bioteknologi, IPB.
Inbar J, Chet I. 1991. Evidence that chitinase produced by Aeromonas caviae is involved in the biological control of soil-borne plant pathogens by this bacterium. Soil Biol Biochem 23:973-978.
Joo GJ. 2005. Purification and characterization of an extracellular chitinase from the antifungal biocontrol agent Streptomyces halstedii. Biotechnol 27:1483-1486.
Karthikeyan M et al. 2005. Induction of resistance in host against the infection of leaf blight pathogen (Altenaria palandui) in onion (Allium cepa var aggregatum). Indian J Biochem Biophys 42:371-377.
Khaosaad T, Garcı´a-Garrido JM, Steinkellner S, Vierheilig H. 2007. Take-all disease is sistemically reduced in roots of mycorrhizal barley plants. Soil Biol Biochemistry 39:727–734.
Koike N, Hyakumachi M, Kageyama K, Tsuyumu S, Doke N. 2001. Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungus: Lignification and superoxide generation. Eur J Plant Pathol 107:523-533.
Lee CS, Lee YJ, Jeun YC. 2005. Observations of infection structures on the leaves of cucumber plants pre-treated with arbuscular mycorrhiza Glomus intraradices after challenge inoculation with Colletotrichum orbiculare. Plant Pathol J 21:237–243.
Lin CC, Kao CH. 2001. Cell wall peroxidase activity, hydrogen peroxide level and NaCl-inhibited root growth of rice seedlings. Plant Soil 230:135-143. McAllister CB, Romera G, Martin I, Godeas J, Ocampo JA. 1995 Interactions
between Aspergilus niger and Glomus mosseae. New Phytol 129:309-316. Mohammadi M, Kazemi H. 2002. Changes in peroxidase and polyphenol activity
in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance. Plant Sci 162:491-498.
Ordentlich A, Elad Y, Chet I. 1987. Rhizosphere colonization by Serratia marcescens for the control of Sclerotium rolfsii. Soil Biol Biochem 19:747-751.
Ozgönen H, Akgul DS, Erkilic A. 2010. The effects of arbuscular mycorrhizal fungi on yield and stem rot caused by Sclerotium rolfsii Sacc. in peanut. African J Agricultural Res 5:128-132.
Pleban S. Chernin L, Chet I. 1997. Chitinolitic activity of an endophytic strain of Bacillus cereus. Lett Appl Microbiol 25:284-288.
Pozo MJ et al. 2002. Localized versus systemic effect of arbuscular mycorrhizal fungi on defence responses to Phytophthora infection in tomato plants. J Exp Bot 53:525–534.
Pradikta AA. 2008. Peran fungi mikoriza arbuskular dan Gliocaldium virens dalam pencegahan penyakit busuk pangkal batang kelapa sawit [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Risanda D. 2008. Pengembangan teknik inokulasi buatan Ganoderma boninense Pat. pada bibit kelapa sawit (Elaeis guineensis Jacq.) [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Roesti D et al. 2005. Bacteria associated with spores of the arbuscular mycorrizal fungi Glomus geoporum and Glomus constrictum. Environ Microbiol 71:6673-6679.
Schaad NW, Jones JB, Chun W. 2001. Laboratorium Guide for Identification of Plant Pathogenic Bacteria. Ed ke-3. Minnesota: APS Pr.
Seevers PM, Daly JM, Catedral FF. 1971. The role of peroxidase isozymes in resistance to wheat stem rust disease. Plant Physiol 48:353-360.
Sinaga MS, Bonny PWS, A Susanto. 2003. Keragaman mikroorganisme rhizosfer kelapa sawit dan patogenesitas Ganoderma boninense Pat. sebagai dasar pengendalian penyakit busuk pangkal batang. Laporan Akhir Hibah Bersaing IX. Bogor: Institut Pertanian Bogor.
Sharda KW, Rodrigues BF. 2009. Applications of arbuscular mycorrhizal fungi in agroecosystems. Trop Subtrop Agroecos 10: 337-354.
Singh PP, YC Shing, CS Park, YR Chung. 1999. Biological control of Fusarium wilt of cucumber by chitinolitic bacteria. Phytopathol 89:92-99.
Suryanto D, Munir E, Yurnaliza. 2006. Eskplorasi bakteri kitinolitik: Keragaman genetik gen penyandi kitinase pada berbagai jenis bakteri dan pemanfaatannya. Laporan Akhir Hibah Bersaing. Medan: Universitas Sumatera Utara.
Suryanto D, Patonah S, Munir E. 2010. Control of Fusarium wilt of chili with chitinolytic bacteria. Hayati 17:5-8.
Susanto A. 2002. Kajian pengendalian hayati Ganoderma boninense Pat. penyebab penyakit busuk pangkal batang kelapa sawit [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Susanto A. Purba RY, Utomo C. 2005. Penyakit-penyakit infeksi pada Kelapa sawit. Medan: Pusat Penelitian Kelapa Sawit.
Tahtamouni MEW, Hameed KM, Saadoun IM. 2006. Biological Control of Sclerotinia sclerotiorum using indigenous chitinolytic Actinomycetes in Jordan. Plant Pathol 22:107-114.
Tenaya IMN, R. Setyamiharja N, Natasasmita. 2001. Correlation of capsaicin content, fructose, and peroxidase activity with antrachnose disease in chilli pepper x red pepper. Zuriat 12:73-83.
Toljander JF, Artursson V, Paul LR, Jansson JK, Finlay RD. 2006. Attachment of different soil bacteria to arbuscular mycorrhizal fungal extraradical hyphae is determined by hyphal vitality and fungal species. FEMS Microbiol Lett 254:34-40.