Cricula trifenestrata (Lepidoptera: Saturniidae):
PERKEMBANGAN EMBRIO, PENUNDAAN PENETASAN
TELUR, DAN PEMECAHAN DORMANSI PUPA
TEGUH LARASATI ANDRIANI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis berjudul ”Cricula trifenestrata (Lepidoptera: Saturniidae): Perkembangan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa”, adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
Teguh Larasati Andriani
ABSTRACT
TEGUH LARASATI ANDRIANI. Cricula trifenestrata (Lepidoptera:
Saturniidae): Embryonic Development, Postponement of Egg Hatching, and Breaking of Pupal Dormancy. Under direction of DEDY DURYADI S and DAMIANA RITA E.
Cricula trifenestrata (Lepidoptera: Saturniidae) is a pledge silkworm because of its golden cocoon. The research aimed to know the embryonic development of C. trifenestrata and its bearing with the postponement of egg hatching and the influence of light intensity and pupal condition on pupal dormancy. The wet mount of each egg stadia (1-8 days) was made by soaking the egg in 5 M NaOH for 3 days. The postponement of egg hatching was done by preserve each stadia of egg for 3, 7, 14, 21, and 28 days in regrigerator (8 0C). Breaking of pupal dormancy was done by place unaked and naked pupae into incubator with 8, 18 and 23 watt lamps. Complete embryonic development of C. trifenestrata occured at the 6th day after oviposition. The postponement of egg hatching of 5, 6 and 7 days old egg could be done for three days. The 5 and 6 days old egg even could be postponed until seven days. The highest percentage of egg hatching was at 6 days old egg. Light intensity didn’t give significantly response to break pupal dormancy. Naked pupa gave positive response for breaking pupal dormancy. The naked pupa that placed incubator with 18 watts lamp showed the fastest breaking pupal dormancy.
RINGKASAN
TEGUH LARASATI ANDRIANI. Cricula trifenestrata (Lepidoptera:
Saturniidae): Perkembangan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa. Dibimbing oleh DEDY DURYADI S dan DAMIANA RITA E.
Cricula trifenestrata (Lepidoptera: Saturniidae) dapat diunggulkan sebagai ulat sutera yang menghasilkan benang berwarna khas yaitu kuning keemasan (‘golden silk’) dan lebih berpori serta tidak mudah kusut dibandingkan dengan sutera biasa. Kokonnya berbentuk seperti jarring dan tidak berbau. Permintaan kokon jauh lebih tinggi daripada ketersediaannya di alam. Oleh karena itu banyak penelitian yang harus dilakukan untuk meningkatkan kualitas dan kuantitas kokon dari ulat sutera jenis ini.
Permasalahan utama dalam domestikasi ulat sutera untuk keperluan komersial adalah penyimpanan telur yang aman untuk menjamin ketersediaan telur. Di alam waktu penetasan berlangsung serempak sehingga perlu diatur waktu penetasannya agar usaha budidaya dapat optimal. Diperlukan informasi yang akurat mengenai perkembangan embrio dari serangga ini terutama pada saat setelah telur dikeluarkan dari induk betina hingga menetas berkembang menjadi larva. Informasi ini akan sangat berguna untuk menentukan fase perkembangan embrio mana yang dapat dihentikan secara sementara. Apabila informasi ini tepat maka sangat berguna bagi
usaha penyimpanan telur dalam berbagai kondisi seperti pada suhu rendah (5 - 10 0C). Dormansi yang terjadi pada pupa C. trifenestrata juga menjadi masalah
karena menyebabkan siklus hidup menjadi lebih panjang. Perlu dipikirkan cara pemecahan dormansi yang terjadi pada pupa C. trifenestrata.
Uji pendahuluan pembuatan preparat embrio dilakukan dengan menggunakan larutan HCl 37%, NaOH 5 M, laktofenol, dan asam asetat. Telur direndam dalam larutan kimia dengan lama perendaman 1 - 5 hari. Tujuan perendaman dalam larutan kimia adalah untuk membuat cangkang telur menjadi transparan dan lunak sehingga embrio dapat dikeluarkan dari cangkangnya. Dari uji pendahuluan ini akan disimpulkan larutan kimia mana yang paling baik dapat digunakan dalam pembuatan preparat embrio C. trifenestrata.
Studi embrio dilakukan pada telur C. trifenestrata mulai hari pertama hingga hari kedelapan setelah oviposisi. Setiap stadia umur (n = 5) dari perkembangan telur (embrio) dibuat preparatnya dengan cara merendam telur dalam larutan NaOH 5 M selama tiga hari pada suhu ruangan yaitu 28 0C. Embrio yang telah dikeluarkan dari cangkangnya kemudian disimpan dalam alkohol 70% yang berfungsi sebagai pengawet preparat sebelum difoto. Pemotretan dilakukan dengan menggunakan mikroskop stereo Nikon WD 70 Plan Apo dan kamera Nikon D100 dengan adapter BR2.
disucihamakan dengan formalin 4% selama lima detik, kemudian dibilas dengan air mengalir. Telur yang telah disucihamakan diletakkan di dalam cawan petri. Jumlah telur setiap cawan petri adalah 30 butir. Telur tersebut disimpan dalam lemari pendingin pada suhu 8 0C selama 3, 7, 14, 21 dan 28 hari. Keempat puluh perlakuan tersebut diulang tiga kali. Selanjutnya dihitung mengenai jumlah dan persentase telur yang menetas setelah diberikan perlakuan.
Kokon yang digunakan dalam percobaan pemecahan dormansi adalah kokon yang berasal dari alam yang tidak keluar menjadi imago setelah satu bulan. Kokon dimasukkan dalam kotak inkubasi dengan ukuran 40 cm x 40 cm x 100 cm. Pada percobaan ini digunakan tiga kotak inkubasi dengan penyinaran lampu yang berbeda yaitu 8, 18, dan 23 watt. Lama penyinaran setiap harinya adalah selama 12 jam. Parameter yang diamati adalah (a) lamanya waktu munculnya imago dari kokon atau pupa, (b) efektivitas perlakuan, (c) suhu pada tiap perlakuan, dan (d) intensitas cahaya. Kelembaban udara di dalam kandang diatur menjadi 80%. Efektivitas perlakuan dihitung dengan cara membandingkan jumlah imago yang keluar dari kokon/pupa setelah diberikan perlakuan dengan jumlah keseluruhan kokon/pupa pada tiap perlakuan.
Lama perendaman dalam larutan kimia mempengaruhi bentuk embrio. Perendaman dalam larutan kimia yang terlalu lama akan membuat embrio menjadi hancur, sebaliknya bila terlalu cepat perendamannya cangkang belum lunak hingga embrio belum dapat dikeluarkan dari cangkangnya. Hasil terbaik dalam percobaan ini adalah perendaman dengan larutan NaOH 5 M selama 3 hari. Oleh karena itu metode ini dipilih untuk pembuatan preparat embrio C. trifenestrata. Pada pengamatan embrio hari pertama terlihat bahwa telah terjadi proses pembelahan yang diikuti dengan pembentukan blastoderm (blastula). Gastrulasi sebagai bagian dari diferensiasi berlangsung pada hari kedua. Pada embrio hari ketiga hingga kelima terjadi perkembangan diferensiasi dan organogenesis. Embrio secara sempurna terbentuk lengkap pada hari keenam. Pada stadia ini peruasan menghasilkan kepala, thoraks dan abdomen. Struktur ‘appendages’ seperti alat mulut dan tungkai telah terbentuk. Stadia hari berikutnya (tujuh dan delapan) merupakan pematangan embrio hingga saat penetasan telur terjadi.
Hal ini diduga karena telur pada stadia ini telah memiliki embrio dengan struktur yang lengkap. Pada lama penyimpanan tujuh hari, stadia telur umur tujuh hari tidak menetas. Diduga hal yang sama terjadi pada kematian embrio hari ke 8 pada penyimpanan tiga hari. Kerusakan embrio karena makin menipisnya cadangan makanan dari kuning telur.
Berdasarkan hasil uji anova, intensitas cahaya tidak memberikan respon yang nyata terhadap pemecahan dormansi pupa C. trifenestrata. Namun demikian terlihat bahwa ada kecenderungan pemberian intensitas yang berbeda akan memberikan waktu keluarnya imago yang berbeda. Intensitas cahaya yang memberikan respon tercepat terhadap keluarnya imago adalah 240 lux (lampu 18 watt). Kokon atau pupa yang mendapatkan perlakuan penyinaran dengan lampu ini cenderung lebih cepat keluar menjadi imago dibanding dengan penyinaran lampu yang lain (8 dan 23 watt). Pemecahan dormansi pupa juga dipengaruhi oleh kondisi pupa. Kondisi pupa yang tidak terbungkus kokon memberikan respon positif untuk pemecahan dormansi pupa. Pupa yang tidak terbungkus kokon lebih cepat keluar menjadi imago dibanding yang terbungkus kokon. Diduga pupa yang tidak terbungkus kokon lebih kuat menerima rangsangan dari lingkungan berupa cahaya dibanding yang pupa yang terbungkus kokon. Salah satu rangsangan untuk memecahkan dormansi pada pupa adalah cahaya. Cahaya akan diteruskan ke otak melalui kutikula pada pupa. Pada pupa yang terbungkus kokon, sebelum menuju kutikula pupa cahaya harus melewati kokon sebagai barier. Selanjutnya proses endokrin yang terjadi adalah proses pengaktifan hormone ekdison yang akan menyebabkan terjadinya pergantian kulit dari pupa menjadi imago. Pada perhitungan efektivitas perlakuan, terlihat perlakuan dengan 23 watt dengan kondisi pupa terbungkus kokon menunjukkan efektivitas perlakuan tertinggi (65%). Namun beberapa kokon/pupa ditemukan mati dan tidak menjadi imago karena parasitoid dan kering. Bila pupa yang mati karena parasitoid tidak dimasukkan dalam perhitungan maka efektivitas perlakuan tertinggi adalah pada perlakuan lampu 8 watt dengan kondisi pupa terbungkus kokon (100%). Namun bila dilihat jumlah ulangannya, maka perlakuan lampu 23 watt dengan kondisi pupa terbungkus kokon lebih banyak dari perlakuan 8 watt dengan kondisi pupa terbungkus kokon. Sehingga dapat disimpulkan perlakuan yang paling efektif adalah perlakuan lampu 23 watt dengan kondisi pupa terbungkus kokon. Parasitoid yang ditemukan menyerang pupa C. trifenestrata adalah dari famili Ichneumonidae dan Sarchopagidae.
©
Hak Cipta milik IPB, tahun 2009 Hak cipta dilindungi Undang-Undang1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
Cricula trifenestrata (Lepidoptera: Saturniidae):
PERKEMBANGAN EMBRIO, PENUNDAAN PENETASAN
TELUR, DAN PEMECAHAN DORMANSI PUPA
TEGUH LARASATI ANDRIANI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Bio Sains Hewan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul : Cricula trifenestrata (Lepidoptera: Saturniidae): Perkembanngan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa
Nama : Teguh Larasati Andriani NIM : G 352070241
Disetujui Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA Dr. drh. Damiana Rita E, MS Ketua Anggota
Diketahui
Koordinator Mayor Biosains Hewan Dekan Sekolah Pascasarjana
Dr. Bambang Suryobroto Prof. Dr. Ir. Khairil Anwar Notodiputro, MS
Cricula trifenestrata (Lepidoptera: Saturniidae):
PERKEMBANGAN EMBRIO, PENUNDAAN PENETASAN
TELUR, DAN PEMECAHAN DORMANSI PUPA
TEGUH LARASATI ANDRIANI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis berjudul ”Cricula trifenestrata (Lepidoptera: Saturniidae): Perkembangan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa”, adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
Teguh Larasati Andriani
ABSTRACT
TEGUH LARASATI ANDRIANI. Cricula trifenestrata (Lepidoptera:
Saturniidae): Embryonic Development, Postponement of Egg Hatching, and Breaking of Pupal Dormancy. Under direction of DEDY DURYADI S and DAMIANA RITA E.
Cricula trifenestrata (Lepidoptera: Saturniidae) is a pledge silkworm because of its golden cocoon. The research aimed to know the embryonic development of C. trifenestrata and its bearing with the postponement of egg hatching and the influence of light intensity and pupal condition on pupal dormancy. The wet mount of each egg stadia (1-8 days) was made by soaking the egg in 5 M NaOH for 3 days. The postponement of egg hatching was done by preserve each stadia of egg for 3, 7, 14, 21, and 28 days in regrigerator (8 0C). Breaking of pupal dormancy was done by place unaked and naked pupae into incubator with 8, 18 and 23 watt lamps. Complete embryonic development of C. trifenestrata occured at the 6th day after oviposition. The postponement of egg hatching of 5, 6 and 7 days old egg could be done for three days. The 5 and 6 days old egg even could be postponed until seven days. The highest percentage of egg hatching was at 6 days old egg. Light intensity didn’t give significantly response to break pupal dormancy. Naked pupa gave positive response for breaking pupal dormancy. The naked pupa that placed incubator with 18 watts lamp showed the fastest breaking pupal dormancy.
RINGKASAN
TEGUH LARASATI ANDRIANI. Cricula trifenestrata (Lepidoptera:
Saturniidae): Perkembangan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa. Dibimbing oleh DEDY DURYADI S dan DAMIANA RITA E.
Cricula trifenestrata (Lepidoptera: Saturniidae) dapat diunggulkan sebagai ulat sutera yang menghasilkan benang berwarna khas yaitu kuning keemasan (‘golden silk’) dan lebih berpori serta tidak mudah kusut dibandingkan dengan sutera biasa. Kokonnya berbentuk seperti jarring dan tidak berbau. Permintaan kokon jauh lebih tinggi daripada ketersediaannya di alam. Oleh karena itu banyak penelitian yang harus dilakukan untuk meningkatkan kualitas dan kuantitas kokon dari ulat sutera jenis ini.
Permasalahan utama dalam domestikasi ulat sutera untuk keperluan komersial adalah penyimpanan telur yang aman untuk menjamin ketersediaan telur. Di alam waktu penetasan berlangsung serempak sehingga perlu diatur waktu penetasannya agar usaha budidaya dapat optimal. Diperlukan informasi yang akurat mengenai perkembangan embrio dari serangga ini terutama pada saat setelah telur dikeluarkan dari induk betina hingga menetas berkembang menjadi larva. Informasi ini akan sangat berguna untuk menentukan fase perkembangan embrio mana yang dapat dihentikan secara sementara. Apabila informasi ini tepat maka sangat berguna bagi
usaha penyimpanan telur dalam berbagai kondisi seperti pada suhu rendah (5 - 10 0C). Dormansi yang terjadi pada pupa C. trifenestrata juga menjadi masalah
karena menyebabkan siklus hidup menjadi lebih panjang. Perlu dipikirkan cara pemecahan dormansi yang terjadi pada pupa C. trifenestrata.
Uji pendahuluan pembuatan preparat embrio dilakukan dengan menggunakan larutan HCl 37%, NaOH 5 M, laktofenol, dan asam asetat. Telur direndam dalam larutan kimia dengan lama perendaman 1 - 5 hari. Tujuan perendaman dalam larutan kimia adalah untuk membuat cangkang telur menjadi transparan dan lunak sehingga embrio dapat dikeluarkan dari cangkangnya. Dari uji pendahuluan ini akan disimpulkan larutan kimia mana yang paling baik dapat digunakan dalam pembuatan preparat embrio C. trifenestrata.
Studi embrio dilakukan pada telur C. trifenestrata mulai hari pertama hingga hari kedelapan setelah oviposisi. Setiap stadia umur (n = 5) dari perkembangan telur (embrio) dibuat preparatnya dengan cara merendam telur dalam larutan NaOH 5 M selama tiga hari pada suhu ruangan yaitu 28 0C. Embrio yang telah dikeluarkan dari cangkangnya kemudian disimpan dalam alkohol 70% yang berfungsi sebagai pengawet preparat sebelum difoto. Pemotretan dilakukan dengan menggunakan mikroskop stereo Nikon WD 70 Plan Apo dan kamera Nikon D100 dengan adapter BR2.
disucihamakan dengan formalin 4% selama lima detik, kemudian dibilas dengan air mengalir. Telur yang telah disucihamakan diletakkan di dalam cawan petri. Jumlah telur setiap cawan petri adalah 30 butir. Telur tersebut disimpan dalam lemari pendingin pada suhu 8 0C selama 3, 7, 14, 21 dan 28 hari. Keempat puluh perlakuan tersebut diulang tiga kali. Selanjutnya dihitung mengenai jumlah dan persentase telur yang menetas setelah diberikan perlakuan.
Kokon yang digunakan dalam percobaan pemecahan dormansi adalah kokon yang berasal dari alam yang tidak keluar menjadi imago setelah satu bulan. Kokon dimasukkan dalam kotak inkubasi dengan ukuran 40 cm x 40 cm x 100 cm. Pada percobaan ini digunakan tiga kotak inkubasi dengan penyinaran lampu yang berbeda yaitu 8, 18, dan 23 watt. Lama penyinaran setiap harinya adalah selama 12 jam. Parameter yang diamati adalah (a) lamanya waktu munculnya imago dari kokon atau pupa, (b) efektivitas perlakuan, (c) suhu pada tiap perlakuan, dan (d) intensitas cahaya. Kelembaban udara di dalam kandang diatur menjadi 80%. Efektivitas perlakuan dihitung dengan cara membandingkan jumlah imago yang keluar dari kokon/pupa setelah diberikan perlakuan dengan jumlah keseluruhan kokon/pupa pada tiap perlakuan.
Lama perendaman dalam larutan kimia mempengaruhi bentuk embrio. Perendaman dalam larutan kimia yang terlalu lama akan membuat embrio menjadi hancur, sebaliknya bila terlalu cepat perendamannya cangkang belum lunak hingga embrio belum dapat dikeluarkan dari cangkangnya. Hasil terbaik dalam percobaan ini adalah perendaman dengan larutan NaOH 5 M selama 3 hari. Oleh karena itu metode ini dipilih untuk pembuatan preparat embrio C. trifenestrata. Pada pengamatan embrio hari pertama terlihat bahwa telah terjadi proses pembelahan yang diikuti dengan pembentukan blastoderm (blastula). Gastrulasi sebagai bagian dari diferensiasi berlangsung pada hari kedua. Pada embrio hari ketiga hingga kelima terjadi perkembangan diferensiasi dan organogenesis. Embrio secara sempurna terbentuk lengkap pada hari keenam. Pada stadia ini peruasan menghasilkan kepala, thoraks dan abdomen. Struktur ‘appendages’ seperti alat mulut dan tungkai telah terbentuk. Stadia hari berikutnya (tujuh dan delapan) merupakan pematangan embrio hingga saat penetasan telur terjadi.
Hal ini diduga karena telur pada stadia ini telah memiliki embrio dengan struktur yang lengkap. Pada lama penyimpanan tujuh hari, stadia telur umur tujuh hari tidak menetas. Diduga hal yang sama terjadi pada kematian embrio hari ke 8 pada penyimpanan tiga hari. Kerusakan embrio karena makin menipisnya cadangan makanan dari kuning telur.
Berdasarkan hasil uji anova, intensitas cahaya tidak memberikan respon yang nyata terhadap pemecahan dormansi pupa C. trifenestrata. Namun demikian terlihat bahwa ada kecenderungan pemberian intensitas yang berbeda akan memberikan waktu keluarnya imago yang berbeda. Intensitas cahaya yang memberikan respon tercepat terhadap keluarnya imago adalah 240 lux (lampu 18 watt). Kokon atau pupa yang mendapatkan perlakuan penyinaran dengan lampu ini cenderung lebih cepat keluar menjadi imago dibanding dengan penyinaran lampu yang lain (8 dan 23 watt). Pemecahan dormansi pupa juga dipengaruhi oleh kondisi pupa. Kondisi pupa yang tidak terbungkus kokon memberikan respon positif untuk pemecahan dormansi pupa. Pupa yang tidak terbungkus kokon lebih cepat keluar menjadi imago dibanding yang terbungkus kokon. Diduga pupa yang tidak terbungkus kokon lebih kuat menerima rangsangan dari lingkungan berupa cahaya dibanding yang pupa yang terbungkus kokon. Salah satu rangsangan untuk memecahkan dormansi pada pupa adalah cahaya. Cahaya akan diteruskan ke otak melalui kutikula pada pupa. Pada pupa yang terbungkus kokon, sebelum menuju kutikula pupa cahaya harus melewati kokon sebagai barier. Selanjutnya proses endokrin yang terjadi adalah proses pengaktifan hormone ekdison yang akan menyebabkan terjadinya pergantian kulit dari pupa menjadi imago. Pada perhitungan efektivitas perlakuan, terlihat perlakuan dengan 23 watt dengan kondisi pupa terbungkus kokon menunjukkan efektivitas perlakuan tertinggi (65%). Namun beberapa kokon/pupa ditemukan mati dan tidak menjadi imago karena parasitoid dan kering. Bila pupa yang mati karena parasitoid tidak dimasukkan dalam perhitungan maka efektivitas perlakuan tertinggi adalah pada perlakuan lampu 8 watt dengan kondisi pupa terbungkus kokon (100%). Namun bila dilihat jumlah ulangannya, maka perlakuan lampu 23 watt dengan kondisi pupa terbungkus kokon lebih banyak dari perlakuan 8 watt dengan kondisi pupa terbungkus kokon. Sehingga dapat disimpulkan perlakuan yang paling efektif adalah perlakuan lampu 23 watt dengan kondisi pupa terbungkus kokon. Parasitoid yang ditemukan menyerang pupa C. trifenestrata adalah dari famili Ichneumonidae dan Sarchopagidae.
©
Hak Cipta milik IPB, tahun 2009 Hak cipta dilindungi Undang-Undang1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
Cricula trifenestrata (Lepidoptera: Saturniidae):
PERKEMBANGAN EMBRIO, PENUNDAAN PENETASAN
TELUR, DAN PEMECAHAN DORMANSI PUPA
TEGUH LARASATI ANDRIANI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Bio Sains Hewan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul : Cricula trifenestrata (Lepidoptera: Saturniidae): Perkembanngan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa
Nama : Teguh Larasati Andriani NIM : G 352070241
Disetujui Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA Dr. drh. Damiana Rita E, MS Ketua Anggota
Diketahui
Koordinator Mayor Biosains Hewan Dekan Sekolah Pascasarjana
Dr. Bambang Suryobroto Prof. Dr. Ir. Khairil Anwar Notodiputro, MS
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah SWT karena atas segala berkat dan rahmat-Nya sehingga penulis dapat menyelesaikan penelitian dan penulisan tesis ini sebagaimana yang diharapkan. Tema yang dipilih dalam penelitian yang dilaksanakan mulai dari bulan Februari 2008 sampai April 2009 adalah Cricula trifenestrata (Lepidoptera: Saturniidae): Perkembangan Embrio, Penundaan Penetasan Telur, dan Pemecahan Dormansi Pupa. Penelitian ini dilaksanakan di laboratorium PPSHB.
Pelaksanaan penelitian dan penulisan tesis ini tidak terlepas dari bantuan berbagai pihak. Penghargaan dan terimakasih dari lubuk hati yang paling dalam penulis ucapkan kepada Dr. Ir. Dedy Duryadi Solihin, DEA, Dr. drh. Damiana Rita E. selaku komisi pembimbing atas bimbingan yang telah diberikan. Semoga jasa baik Bapak dan Ibu mendapatkan pahala yang setimpal dari Tuhan YME. Penulis juga menyampaikan ucapan terimakasih kepada Kepala MAN 1 Kudus yang telah memberi ijin dan dukungan selama melaksanakan studi di IPB. Kepada Pimpinan IPB dan Sekolah Pascasarjana IPB, Pimpinan dan staf pengajar Departemen Biologi FMIPA IPB, Ketua Mayor BSH IPB, Pimpinan dan staf Laboratorium Biologi Molekuler PPSHB.
Penelitian ini tidak akan terlaksana tanpa bantuan dana, untuk itu penulis mengucapkan terimakasih kepada Departemen Agama RI atas kesempatan yang diberikan sehingga penulis dapat mengikuti program pascasarjana ini. Ucapan terimakasih penulis sampaikan kepada Dr. Sutrisno Hari atas diskusi tentang Saturniidae, Bapak Dr. drh. Adiwinarto dan Mbak Tini atas masukannya dalam pembuatan preparat embrio, Dr. Kikin Muttaqin atas bantuan dokumentasi embrio, Bapak Roni atas bantuan kokon, Ibu Aisyah dan Bapak Heri atas kerjasama dan dukungannya.
Kepada keluarga terutama Bapak dan Ibu saya (Soejatmin dan Sumaryatun) yang senatiasa mendukung dan mendoakan keberhasilan saya. Kakak-kakak dan adik saya (Mas Guh, Mbak Tik, Mas Sulis, Mbak Heni, Indah) serta keponakan-keponakan tercinta (Niken, Ais, Aji, Melati, Eno, Ata). Teman-teman satu angkatan, teman-teman satu laboratorium, dan semua pihak yang mendukung penelitian ini. Mudah-mudahan Allah SWT membalas budi baik Bapak, Ibu dan saudara-saudara semuanya. Amin
RIWAYAT HIDUP
Penulis dilahirkan di Kudus, Jawa Tengah pada tanggal 29 September 1974
sebagai anak kelima dari enam bersaudara, dari pasangan H. Soejatmin dan Hj. Sumaryatun. Penulis menyelesaikan pendidikan Sekolah Dasar pada SD Negeri
Jember 2 Kudus pada tahun 1987, SMP Negeri 2 Kudus pada tahun 1990 dan SMA Negeri 1 Kudus pada tahun 1993. Lulus dari Jurusan Biologi Fakultas MIPA Universitas Diponegoro pada tahun 1999. Tahun 2007 memperoleh kesempatan melanjutkan program Pascasarjana IPB dengan beasiswa dari Departemen Agama.
DAFTAR ISI
Halaman
DAFTAR TABEL ... xiii DAFTAR GAMBAR ... xiv DAFTAR LAMPIRAN ... xv PENDAHULUAN
Latar Belakang ... 1 Permasalahan ... 4 Hipotesis ... 4 Tujuan Penelitian ... 4 Manfaat Penelitian ... 4 Kerangka Penelitian ... 5 TINJAUAN PUSTAKA
Klasifikasi Cricula trifenestrata ... 6 Distribusi C. trifenestrata ... 6 Biologi C. trifenestrata. ... 7 Telur ... 7 Larva ... 8 Pupa ... 8 Imago ... 9 Perkembangan Embrio Serangga ... 10 Faktor Yang Mempengaruhi Metamorfosis Serangga ... 14 Pengaruh Cahaya pada Metamorfosis ... 15 Strategi Serangga Untuk Bertahan di Musim Dingin ... 16 Hormon Pengendali Metamorfosis ... 16 METODE PENELITIAN
HASIL
Pengamatan siklus hidup C. trifenestrata ... 25 Studi Perkembangan Embrio C. trifenestrata ... 30 Penundaan Penetasan Telur C. trifenestrata dengan Penyimpanan dalam Suhu 8 0C ... 33 Pemecahan dormansi Pupa C. trifenestrata ... 35 PEMBAHASAN
Siklus Hidup C. trifenestrata ... 39 Studi Perkembangan Embrio C. trifenestrata ... 39 Penundaan Penetasan Telur C. trifenestrata dengan Penyimpanan dalam Suhu8 0C ... 41 Pemecahan dormansi Pupa C. trifenestrata ... 42 SIMPULAN DAN SARAN
DAFTAR TABEL
Halaman
DAFTAR GAMBAR
Halaman
DAFTAR LAMPIRAN
Halaman
PENDAHULUAN
Latar Belakang
Industri sutera di Indonesia telah berkembang selama kurang lebih 80 tahun (mulai dari tahun 1930-an sampai sekarang) (Atmosoedarjo et al. 2000). Selama perkembangan tersebut ulat sutera yang dibudidayakan adalah Bombyx mori yang tergolong dalam famili Bombycidae. Banyak kendala yang dihadapi dalam budidaya sutera ini, antara lain masih tergantungnya pasokan bibit ulat sutera berkualitas dari luar negeri dan adanya hambatan klimatik (karena Bombyx bukan ngengat asli Indonesia).
Selain Bombyx mori, ada jenis ngengat lain yang mampu menghasilkan sutera yaitu Antheraea yamamai (dikembangkan di Jepang), Antheraea polypemu (dikembangkan di Amerika Utara), Antheraea pernyi (dikembangkan di China), Philosamia ricini (sutera eri dikembangkan di India), Antheraea mylitta (sutera tasar dikembangkan di India), dan Attacus atlas. Indonesia memiliki potensi penghasil sutera dari ulat sutera liar. Beberapa jenis ulat sutera liar yang daerah penyebarannya ada di Indonesia adalah Cricula trifenestrata, Attacus atlas dan Antherea spp (Paukstadt U & Paukstadt LH 2004) yang selama ini dianggap sebagai hama, ternyata mampu menghasilkan benang sutera yang bernilai ekonomis. Dibandingkan dengan Bombyx, ulat sutera liar asli Indonesia ini memiliki keunggulan yaitu relatif bebas dari hambatan klimatik.
Purwanti 2005). C. trifenestrata dapat diunggulkan sebagai ulat sutera yang menghasilkan benang berwarna khas yaitu kuning keemasan (‘golden silk’) dan lebih berpori serta tidak mudah kusut dibandingkan dengan sutera biasa. Kokonnya berbentuk seperti jaring (Kalshoven 1981) dan tidak berbau. Oleh karena itu banyak penelitian yang harus dilakukan untuk meningkatkan kualitas dan kuantitas kokon dari ulat sutera jenis ini.
Selama ini para pengumpul C. trifenestrata mendapatkan kokon dengan cara mengumpulkannya dari alam karena memang belum ada upaya budidaya. Masalah yang dihadapi para pengumpul adalah bahwa kokon tidak ditemukan secara kontinyu sepanjang tahun. Dugaan sementara kelangkaan kokon di masa-masa tertentu disebabkan oleh strategi adaptasi dari serangga ini dengan melakukan aktivitas hidupnya hanya pada kondisi lingkungan yang kondusif (‘favorable’) bagi kehidupannya.
tinggi, hewan vertebrata, dan hewan arthropoda menggunakan lama pencahayaan atau fotoperiode untuk mensinkronisasikan pertumbuhan, perkembangan, reproduksi, dormansi, migrasi dengan perubahan musim. Selain itu kondisi iklim mikro terutama kelembaban dapat mempercepat keluarnya ngengat dari kepompong. Pada ngengat jenis lain, Rachman (2001) menyatakan lama periode pupa Attacus atlas di musim penghujan lebih pendek daripada di musim kemarau. Perlakuan fotoperiode dapat mempersingkat waktu periode pupa di musim kemarau dan sebaliknya dapat memperpanjang waktu periode pupa di musim penghujan. Demikian juga menurut William et al. (1964), diapause pupa Antherea pernyi dapat diperlambat pada suhu 25 0C dengan kondisi penyinaran pendek (fotofase 4 - 12 jam) dan dapat dipercepat dengan kondisi penyinaran panjang (fotofase 15 - 18 jam). Ostrinia nubilalis (Pyralidae) yang dipelihara dalam laboratorium, diapause fakultatif dapat terjadi saat diinduksi dengan penyinaran pendek (McLeod & Beck 1963). Wilayah tropis memiliki kondisi iklim yang berbeda dengan wilayah subtropis. Beberapa serangga pada daerah tropis melakukan penyesuaian kondisi iklim dengan melakukan dormansi. Denlinger (1986) menyatakan bahwa dormansi serangga tropis meliputi aestivasi (diapause di musim kering) dan pluviasi (diapause di musim basah).
Permasalahan utama dalam domestikasi ulat sutera untuk keperluan komersial adalah penyimpanan telur yang aman untuk menjamin ketersediaan telur. Di alam waktu penetasan berlangsung serempak sehingga perlu diatur waktu penetasannya agar usaha budidaya dapat optimal. Diperlukan informasi yang akurat mengenai perkembangan embrio dari serangga ini terutama pada saat setelah telur dikeluarkan dari induk betina hingga menetas berkembang menjadi larva. Informasi ini akan sangat berguna untuk menentukan fase perkembangan embrio mana yang dapat dihentikan secara sementara. Apabila informasi ini tepat maka sangat berguna bagi
usaha penyimpanan telur dalam berbagai kondisi seperti pada suhu rendah (5 - 10 0C). Sebagai contoh padaBombyx mori, telur dapat ditunda penetasannya
perkembangan embrio, penyimpanan telur yang tetap fertil dan ‘viable’ dan percepatan kemunculan imago dari fase pupa dengan berbagai cara.
Permasalahan
Permasalahan dari penelitian ini adalah bagaimana perkembangan embrio C. trifenestrata, pada tahap / umur embrio berapa hari dapat dilakukan penundaan penetasan telur dan bagaimana pengaruh intensitas cahaya dan kondisi pupa terhadap pemecahan dormansi pupa C. trifenestrata.
Hipotesis
Hipotesis dari penelitian ini adalah suhu 8 0C dapat digunakan untuk menunda penetasan telur C. trifenestrata; intensitas cahaya dan kondisi pupa memberikan pengaruh pada pemecahan dormansi pupa C. trifenestrata.
Tujuan
Tujuan dari penelitian ini adalah mengetahui perkembangan embrio C. trifenestrata, melakukan upaya penundaan penetasan telur dan mengetahui
pengaruh intensitas cahaya dan kondisi pupa pada pemecahan dormansi pupa C. trifenestrata.
Manfaat Penelitian
Alur Kerangka Penelitian
Gambar 1 Alur kerangka penelitian
Industri sutera (Bombyx) di Indonesia tergantung pasokan bibit berkualitas dari luar negeri, namun budidayanya menghadapai hambatan klimatik (Bombyx bukan ngengat asli Indonesia).
Perlu eksplorasi mengenai sumber benang sutera yang asli Indonesia selain dari Bombyx sehingga relatif bebas hambatan klimatik.
C. trifenestrata merupakan salah satu penghasil sutera liar asli Indonesia yang belum dibudidaya tetapi jumlahnya melimpah di alam.
Informasi biologi ulat sutera C. trifenestrata terbatas.
1. Perkembangan embrio 2. Cara penundaan penetasan 3. Pemecahan dormansi pupa
1. Siklus hidup 2. Pakan larva 3. Pengolahan kokon
4. Pemanfaatan kokon untuk keperluan lain 5. Breeding
6. Domestikasi
TINJAUAN PUSTAKA
Klasifikasi Cricula trifenestrata
Cricula trifenestrata termasuk dalam kelompok ngengat yaitu sejenis kupu nokturnal (aktif di malam hari) yang memiliki ciri-ciri antara lain antena pada jantan berbentuk quadripektinate sedang pada betina berbentuk bipektinate. Dalam genus Cricula, di antara spesies, terdapat perbedaan dalam hal warna. Spot mata pada sayap telah bermodifikasi, ocelus pada sayap belakang mereduksi menjadi titik kecil kehitaman atau jendela hyaline kecil yang bulat. Pada sayap depan, ocelus juga mereduksi, tapi terdapat satu, dua atau tiga spot yang mirip membentuk seperti sebuah kepulauan pada sayap. Terdapat seksual dimorfisme, betina selalu lebih besar dengan sayap yang membulat dan biasanya berwarna lebih gelap dari jantan. Warna sayap juga bervariasi dari coklat, merah gelap-oranye hingga coklat kekuningan (Nassig et al.1996).
Dalam klasifikasi, C. trifenestrata termasuk dalam kingdom Animalia, phylum Arthropoda, kelas Insecta, ordo Lepidoptera, famili Saturniidae, genus Cricula (Triplehorn & Johnson 2005).
Distribusi C. trifenestrata (Lepidoptera: Saturniidae)
Di dunia ada sekitar 1200 - 1500 spesies dari famili Saturniidae dengan pusat keragaman ada di Neotropik, Asia Tenggara antara Peninsula Indomalaya dan New Guinea (70 spesies). Di Sumatra dan pulau kecil lain di sekitarnya terdapat sekitar 24 spesies (Nassig et al. 1996).
(tersebar di paparan Sunda), dan C. trifenestrata (tersebar di daerah oriental). Malaysia barat diketahui memiliki 2 spesies yaitu C. trifenestrata dan C. elaezia. Borneo diketahui memiliki 3 spesies yaitu C. bornea (spesies endemik), C. elaezia (tersebar di paparan Sunda) dan C. trifenestrata. Jawa dan Bali diketahui memiliki 2 spesies yaitu C. trifenestrata dan C. elaezia (Nassig et al. 1996).
C. trifenestrata merupakan spesies Cricula yang paling luas penyebarannya. Spesies ini dapat ditemukan sebagai subspecies yang berbeda di wilayah India selatan dan Asia Tenggara yang meliputi Philipina (C. t. treadawayi) dan Indonesia (Jawa
yaitu C. t. javana; Sumatra yaitu C. t. javana; Sulawesi yaitu C. t. kransi & C. t. banggaiensis; Sumba yaitu C. t. tenggarrensis; Maluku yaitu C.t. serama & C.
t. banggaiensis) (Nassig et al. 1996).
Biologi Cricula trifenestrata
C. trifenestrata disebut juga ulat kipat bersifat polifag dan sangat rakus. Selain jambu mete larva ulat sutera ini juga menyerang alpukat, kenari, jambu, kedondong, mangga, kakao, dan kayumanis (Kalshoven 1981).
C. trifenestrata termasuk serangga holometabola yaitu serangga yang mengalami metamorfosis sempurna. Siklus hidupnya terbagi menjadi empat tahap yaitu telur, larva atau ulat, pupa atau kepompong dan imago atau ngengat dewasa (Gullan & Cranston 2000). Lamanya siklus hidup C. trifenestrata dari telur hingga imago rata-rata 63 – 77 hari (Deptan 1995).
Telur
Larva
Menurut Rono et al. (2008) larva C. trifenestrata terdiri dari lima instar dengan pergantian kulit empat kali. Perubahan larva dari instar 1 ke instar 5 membutuhkan waktu masing-masing 5 hari, sehingga fase larva berlangsung kurang lebih 25 hari. Larva instar 1 berwarna kuning hingga kuning kecoklatan yang kemudian akan berubah menjadi kuning kemerahan dengan kepala hitam. Bagian thoraks terdiri dari 3 segmen dan tiap segmen terdapat sepasang kaki dan tubuhnya ditutupi rambut-rambut halus. Bagian abdomen memiliki 5 pasang proleg yang terdapat pada segmen abdomen ketiga hingga keenam dan segmen abdomen kesepuluh. Larva instar 2 berwarna kombinasi dari kuning, merah dan hitam dengan kepala coklat, tubuh ditumbuhi bulu-bulu halus. Larva instar 3, warna tubuhnya adalah kuning kemerahan dengan kepala coklat, tubuhnya ditumbuhi bulu-bulu halus berwarna putih dan bagian ventralnya berwarna merah. Larva ini bergerak lebih aktif, memakan lebih banyak dan ukuran bertambah dibandingkan 2 instar sebelumnya. Larva instar 4 berwarna mirip dengan instar 5 namun berbeda dalam ukuran. Larva instar 5 berwarna merah dengan kepala merah tubuhnya ditumbuhi bulu-bulu halus berwarna putih agak kasar dan terdapat garis hitam melingkar mulai dari kepala sampai abdomen.
Hasil penelitian Rono et al. (2008) mengenai ukuran larva instar 1 hingga
instar 5 menunjukkan peningkatan panjang rata-rata dari 4.08 ± 0.15 hingga 81.49 ± 0.23 mm dan lebar rata-rata dari 2.17 ± 0.18 hingga 11.22 ± 0.22 mm.
Pupa
Hasil penelitian Rono et al. (2008) mengenai ukuran pupa menunjukkan bahwa panjang dan lebar rata-rata pupa jantan adalah 31.64 ± 0.37 mm dan 10.61 ± 0.31 mm, sedangkan panjang dan lebar rata-rata pupa betina adalah 35.57 ± 0.40 mm dan 12.58 ± 0.31 mm.
Pupa C. trifenestrata terbungkus dalam kokon yang berbentuk jala rapat berwarna kuning emas . Dalam keadaan normal stadia pupa antara 21 - 26 hari, tetapi apabila keadaan tidak menguntungkan dapat terjadi sampai 3 bulan (Deptan 1995). Penundaan perkembangan dari stadia pupa menjadi imago dikenal sebagai diapause. Masa istirahat yang relatif panjang ini dapat dihentikan dengan cara memberikan rangsangan fisik/ lingkungan berupa perlakuan fotoperiode (Rachman 2001).
Hasil penelitian Rono et al. (2008) mengenai ukuran kokon menunjukkan bahwa panjang dan lebar rata-rata kokon jantan adalah 41.98 ± 0.55 mm dan 12.31 ± 0.29 mm, sedangkan panjang dan lebar rata-rata kokon betina adalah 50.89 ± 0.43 mm dan 16.22 ± 0.31 mm.
Imago
Serangga dewasa C. trifenestrata adalah serangga nokturnal, berwarna kekuningan hingga kemerahan. Jantan memiliki dua spot gelap pada sayap depan, sedangkan betinanya memiliki tiga spot transparan yang tidak teratur pada sayap depan dan satu spot pada sayap belakang. Pada bagian dekat dasar sayap depan nampak garis hitam berombak. Kepala, torak, abdomen dan appendiks ditutup oleh sisik yang berwarna coklat kekuningan. Antena bipektin panjang dengan jumlah segmen yang tidak terdefinisi. Antena jantan lebih lebar dari betina. Segmen abdomen terakhir betina lebih lebar dari jantan. Ukuran tubuh betina lebih besar dari jantan.
Ngengat betina mulai bertelur satu hari setelah kawin. Ngengat betina yang tidak kawin juga bertelur pada hari kedua setelah keluar dari pupa namun telur yang dihasilkan adalah telur yang infertil. Hasil penelitian Rono et al. (2008) mengenai lama hidup imago menunjukkan bahwa lama hidup imago jantan rata-rata adalah 2.90 ± 0.38 hari, sedangkan betinanya adalah 4.20 ± 0.42 hari.
Perkembangan Embrio Serangga
Telur serangga adalah sebuah sel dengan dua lapis selaput bagian luar, satu selaput vitelin yang tipis yang mengelilingi sitoplasma dan korion bagian luar. Korion berupa kulit bagian luar dari telur tersebut yang keras. Telur mempunyai satu lubang yang kecil atau beberapa lubang (mikrofil) pada satu ujung, yang menjadi tempat masuknya sperma ke dalam telur. Di sebelah dalam selaput vitelin terdapat satu lapisan korteks, dimana di dalamnya terdapat kuning telur (‘egg yolk’) (Gambar 2 a) (Triplehorn & Johnson 2005).
Gambar 2 Pembelahan nukleus Gambar 3 Embrio dengan germ band
Setelah terbentuk sel-sel kutub, membran plasma dari telur akan melipat ke dalam di antara nuklei, sehingga tiap-tiap sel mempunyai satu nukleus yang disebut sel-sel blastoderm (Gambar 2 d) yang menyusun diri membungkus yolk dari embrio. Pada bagian ventral dari embrio, blastoderm akan membentuk penebalan yang merupakan bakal embrio nantinya (‘germ band’) yang kemudian akan memanjang dan berkembang menjadi embrio (Gambar 3), sedangkan sel blastoderm lainnya akan berkembang membentuk selaput ekstraembrionik. Daerah ventral yang akan menjadi embrio disebut ‘germ anlagen’ (Gilbert 1985).
Gambar 4 Perkembangan ‘amnion cavity’ (Chapman 1998)
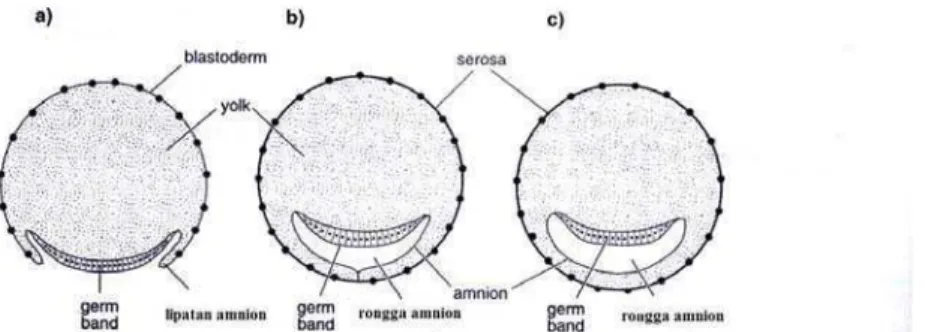
Pada waktu pembentukan amnion dan serosa, terjadi juga proses gastrulasi, yang dimulai dengan invaginasi (melekuk ke dalam) bagian bawah (venter) ‘germ band’. Sel-sel blastoderm yang merupakan hasil dari fase pembelahan, pada gastrulasi akan melakukan perpindahan-perpindahan untuk membentuk lapisan ektoderm, mesoderm dan endoderm. Pergeseran-pergeseran yang terjadi pada gastrulasi ini meliputi seluruh embrio. Migrasi dari sel pada suatu bagian embrio harus terkoordinasi dengan pergerakan-pergerakan yang terjadi pada waktu yang sama (Gilbert 1985). Invaginasi pada proses ini akan mendatar ke arah luar dan pinggir-pinggir luarnya bertemu dan bersatu membentuk pita longitudinal dari sel-sel (lapis dalam) yang dikelilingi oleh lapis luar yaitu ektoderm. Tipe lain pembentukan lapisan dalam adalah mengendapnya pita longitudinal bawah ke dalam kuning telur, yang kemudian tumbuh membentuk sel-sel ‘germ band’ yang tertinggal. Tipe yang lain lagi, lapisan dalam berkembang dari proliferasi ‘germ band’. Kemudian lapisan dalam berkembang menjadi dua pita longitudinal lateral (mesoderm) dan ‘median strands’ dengan masa sel pada ujung anterior dan posterior yang akan menjadi endoderm (Triplehorn & Johnson 2005).
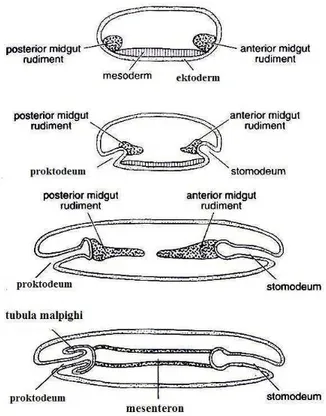
Gambar 5 Pembentukan saluran pencernaan (Chapman 1998)
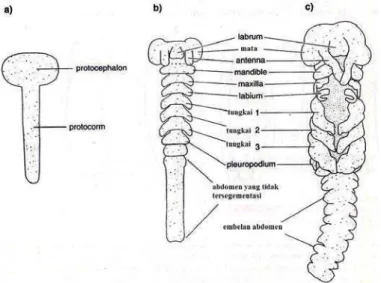
Gambar 6 Perkembangan appendages serangga (Chapman 1998)
Faktor Yang Mempengaruhi Metamorfosis Serangga
Metamorfosis merupakan perubahan bentuk, struktur dan fungsi (morfologi, anatomi, dan fisiologi) organisme dari larva menjadi dewasa. Perubahan tersebut diawali dan diatur oleh signal hormon. Namun demikian tanggapan jaringan atau organ terhadap signal hormon tidak sama, tergantung pada stadium perkembangannya (Muller 1997). Faktor dalam (intrinsik) maupun faktor luar (ekstrinsik) seringkali berinteraksi mempengaruhi proses metamorfosis. Faktor dalam antara lain genetis dan hormonal, sedangkan faktor luar yang umum adalah pakan, suhu, kelembaban, dan cahaya (Gullan & Cranston 2000).
Banyak serangga memiliki lebih dari satu generasi dalam satu tahun. Serangga seperti ini melanjutkan reproduksinya sepanjang musim selama kondisi lingkungan sesuai dan pakan tersedia. Pada beberapa serangga, perkembangan dapat terhenti selama satu tahapan khusus dalam siklus tahunan. Periode ini secara genetik telah diprogram dan disebut diapause. Periode istirahat pada musim dingin disebut hibernasi dan periode istirahat selama suhu tinggi disebut aestivasi. Diapause pada serangga dikendalikan secara genetik, namun pada awal maupun akhir dari fase ini dirangsang oleh faktor-faktor lingkungan seperti faktor fisik (suhu dan fotoperiode) dan faktor ketersediaan makanan (Hunter & McNeil 1997).
Pengaruh Cahaya pada Metamorfosis
Fotoperiode merupakan istilah untuk pengukuran waktu selama 24 jam yang terdiri dari waktu terang dan waktu gelap. Proporsi masing-masing bervariasi tergantung musim dan posisi daerah menurut garis lintang. Periode terang (‘day length’) juga disebut sebagai fotofase yaitu periode bumi menerima cahaya matahari sejak matahari terbit sampai tenggelam. Periode gelap (‘night length’), disebut scotofase yaitu periode sejak matahari tenggelam sampai terbit (Beck 1980). Fotoperiode berfungsi dalam adaptasi biologi dengan menyediakan informasi lingkungan dalam bentuk signal ruang sementara. Pada serangga pengaruh fotoperiode antara lain pada aktivitas lokomotor, perilaku makan, dan perilaku kawin. Aktivitas motorik biasanya berakibat pada perilaku makan, kawin, dan bertelur. Puncak aktivitas hewan diurnal bervariasi tergantung spesiesnya, namun tetap pada fase terang. Pada hewan nokturnal, gelap merupakan rangsangan untuk aktivitasnya (Beck 1980).
tersebut. Sekresi feromon pada ngengat terjadi pada separo waktu akhir scotofase, pada spesies lain dapat terjadi sebaliknya atau dengan pola lain (Gullan & Cranston 2000).
Strategi Serangga Untuk Bertahan di Musim Dingin
Pada saat musim dingin banyak serangga mati. Namun beberapa serangga dapat memiliki toleransi dengan temperatur yang bisa membekukan dengan mengontrolnya menjadi kristal es atau menggunakan anti beku (‘anti freezing’). Suhu musim dingin dapat membekukan sel. Ketika hal ini terjadi partikel dalam cairan tubuh mengelilingi nukleus membentuk kristal es. Jika kristal es terus terbentuk tanpa intervensi, maka membran sel akan terpotong, sehingga merusak sel dan jaringan. Kebanyakan organisme yang bertahan hidup pada musim dingin memproduksi molekul antifreezing dan mengeliminasi sampah dari tubuh (untuk mengurangi sejumlah nukleus yang ada untuk menginisiasi formasi es) (Danks 2006). Molekul antifreezing yang paling umum adalah glukosa dan gliserol. Ketika ditemukan dalam tingkat konsentrasi yang tinggi, glukosa dapat mengurangi titik beku hingga 2 derajat centigrade. Kebanyakan serangga memproduksi sejumlah gliserol (Chapman 1998). Kebanyakan organisme juga memproduksi protein antifreezing (Antifreezing Proteins/AFPs) pada temperatur rendah.
Hormon Pengendali Metamorfosis
Dalam menanggapi stimuli lingkungan, sel-sel neurosekretori dalam otak serangga mensekresi hormon peptida yang disebut prothoracicotropic hormone (PTTH). Melalui sistem yang kompleks, PTTH kemudian merangsang kelenjar prothoraks untuk mensintesis hormon steroid yang disebut ekdison. Ekdison merupakan hormon yang belum aktif (20-hydroxyecdysone). Untuk menyederhanakan, dua-duanya kemudian disebut sama sebagai ekdison. Setiap pergantian kulit dipicu oleh peningkatan kadar ekdison dalam hemolimfe (Kalthoff 1996; Muller 1997).
berhubungan dengan otak sehingga sekresi JH juga dikendalikan oleh otak. Hormon juvenile secara esensial menghambat metamorfosis. Apabila konsentrasi JH dalam hemolimfe tinggi pada saat pergantian kulit, terbentuklah larva. Penurunan konsentrasi JH dalam hemolimfe pada saat pergantian akan menghasilkan pupa, dan ketika hormone juvenile dalam hemolimfe tidak ada sama sekali akan diperoleh imago (Kalthoff 1996).
Keberadaan JH pada konsentrasi tinggi berperan dalam memelihara stadium larva dan mempengaruhi pertumbuhannya sehingga larva tumbuh menjadi semakin besar. Selama pertumbuhan larva, kutikula mengalami pengerasan sehingga suatu saat tidak mampu mengikuti pertumbuhan larva. Akibatnya, kutikula tersebut ditanggalkan dan diganti dengan kutikula baru yang telah disintesis sebelum pergantian kulit. Kutikula baru yang masih bersifat elastis selanjutnya juga akan mengalami pengerasan lagi seiring dengan pertumbuhan larva (Muller 1997). Menurut Wiggleworth (1997), gelapnya warna dan pengerasan kutikula dipengaruhi oleh bursicon yang dihasilkan ganglia thoracic. Proses penanggalan kutikula (ecdysis) tersebut terjadi beberapa kali di bawah pengaruh hormon ekdison.
proses ekdisis, ekdison juga bertanggung jawab terhadap proses metamorfosis. Menurut Muller (1997), metamorfosis sempurna meliputi dua tahap utama yaitu sebagai berikut: tahap pertama adalah perubahan dari larva instar terakhir menjadi pupa yang tidak aktif dan tahap kedua keluarnya imago dari pupa. Selama stadium pupa terjadi beberapa organogenesis dan organolisis.
Ketiadaan faktor lingkungan yang tepat dan sesuai, dapat menyebabkan hewan atau tumbuhan mengalami ‘tidur’. Keadaan demikian pada tumbuhan disebut dormansi, sedangkan pada hewan disebut hibernasi atau diapause. Diapause merupakan istilah yang sering digunakan untuk kelompok serangga.
Diapause dapat terjadi baik pada stadium telur, larva, maupun pupa. Diapause pupa ditandai dengan penurunan laju metabolisme, penghentian differensiasi menuju ke kedewasaan, dan resistensi terhadap kehilangan air melalui transpirasi. Laju metabolism yang diukur dengan laju konsumsi oksigen selama diapause turun sampai 3% (Beck 1980).
Rendahnya laju pengambilan O2 pada diapause pupa dikaitkan dengan aktivitas enzim oksidatif mitokondria. Pada kajian klasik terhadap diapause Hyalophora cecropia (Saturniidae), ditunjukkan bahwa status diapause diatur secara humoral. Tidak adanya prothoracicotropic hormon (PTTH), kelenjar prothoraks tidak menghasilkan hormon ecdyson sehingga perkembangan serangga menjadi stadium dewasa terhenti. Diapause pupa yang terjadi dan dipertahankan akibat tidak adanya stimulus PPTH pada kelenjar prothoraks, mengakibatkan diapause segera berakhir bila mendapat ekdison eksternal (Beck 1980).
METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian dilakukan di laboratorium PPSHB (Pusat Penelitian dan Sumber Daya Hayati) Institut Pertanian Bogor, berlangsung dari bulan Pebruari 2008 hingga Maret 2009.
Alat dan Bahan Penelitian
Bahan utama yang digunakan dalam penelitian ini adalah kokon dan telur C. trifenestrata, larutan kimia: HCl 37%, NaOH 5 M, laktofenol, dan asam asetat glacial, formalin 4%, dan alkohol 70%. Alat-alat yang digunakan adalah cawan petri berdiameter 11 cm, kandang imago dan kandang perlakuan berupa kotak berukuran panjang x lebar x tinggi = 40 cm x 40 cm x 100 cm, lampu Philips SL 8, 18, 23 watt, lemari pendingin, pengatur waktu (‘timer’), hygrometer, lux meter dan termometer maksimum-minimum.
Cara Kerja
.
Pengamatan siklus hidup C. trifenestrata
Pengamatan siklus hidup C. trifenestrata meliputi pengamatan telur, larva, pupa dan imago. Pengamatan dilakukan di lapangan dan di laboratorium.
Pengamatan Embrio C. trifenestrata
Studi embrio dilakukan pada telur C. trifenestrata mulai hari pertama hingga hari kedelapan setelah oviposisi (telur C. trifenestrata menetas pada hari ke 8 - 10 setelah oviposisi). Setiap stadia umur (n = 5) dari perkembangan telur (embrio) dibuat preparatnya dengan cara merendam telur dalam larutan NaOH 5 M selama tiga hari pada suhu ruangan yaitu 28 0C. Larutan NaOH 5 M berfungsi untuk menghancurkan cangkang telur dan membuat embrio menjadi transparan. Embrio yang telah dikeluarkan dari cangkangnya kemudian disimpan dalam alkohol 70% yang berfungsi sebagai pengawet preparat sebelum difoto. Pengamatan embrio dilakukan pada preparat embrio mulai dari hari pertama hingga hari ke delapan pada stadia telur. Pemotretan dilakukan dengan menggunakan mikroskop stereo Nikon WD 70 Plan Apo dan kamera Nikon D100 dengan adapter BR2.
Penundaan Penetasan Telur
Telur yang digunakan dalam percobaan ini adalah telur fertil hasil dari perkawinan imago jantan dan betina. Selanjutnya telur tersebut disucihamakan dengan formalin 4% selama lima detik, kemudian dibilas dengan air mengalir. Telur yang telah disucihamakan diletakkan di dalam cawan petri berdiameter 11 cm yang telah disterilkan. Jumlah telur setiap cawan petri adalah 30 butir. Selanjutnya telur inilah yang akan diberikan perlakuan penundaan penetasan. Telur tersebut disimpan dalam lemari pendingin pada suhu 8 0C.
Faktor Lama penyimpanan
H3 H7 H14 H21 H28
Umur telur
U1 U1H3 U1H7 U1H14 U1H21 U1H3 U2 U2H3 U2H7 U2H14 U2H21 U2H3 U3 U3H3 U3H7 U3H14 U3H21 U3H3 U4 U4H3 U4H7 U4H14 U4H21 U4H3 U5 U5H3 U5H7 U5H14 U5H3 U5H3 U6 U6H3 U6H7 U6H14 U6H3 U6H3 U7 U7H3 U7H7 U7H14 U7H3 U7H3 U8 U8H3 U8H7 U8H14 U8H3 U8H3 Dimana : U1: Stadia umur telur 1 hari
U2: Stadia umur telur 2 hari U3: Stadia umur telur 3 hari U4: Stadia umur telur 4 hari U5: Stadia umur telur 5 hari U6: Stadia umur telur 6 hari U7: Stadia umur telur 7 hari U8: Stadia umur telur 8 hari H3: Lama penyimpanan 3 hari H7: Lama penyimpanan 7 hari H14: Lama penyimpanan 14 hari H21: Lama penyimpanan 21hari H28: Lama penyimpanan 28 hari
Keempat puluh perlakuan tersebut diulang tiga kali dengan masing-masing ulangan sebanyak 30 telur sehingga total telur yang dipakai adalah 40 x 3 x 30 (3600 butir). Selanjutnya dilakukan penghitungan mengenai jumlah dan persentase telur yang menetas setelah diberikan perlakuan.
Pemecahan Dormansi Pupa
100 cm (Gambar 7). Pada percobaan ini digunakan tiga kotak inkubasi dengan penyinaran lampu yang berbeda yaitu 8, 18, dan 23 watt. Lama penyinaran setiap harinya adalah selama 12 jam.
Gambar 7 Kotak inkubasi
Rancangan percobaan yang digunakan adalah faktorial dengan dua faktor. Faktor pertama yaitu pemberian penyinaran lampu yang terdiri dari tiga taraf yaitu 8 watt, 18 watt, 23 watt; dan faktor kedua adalah kondisi pupa yang terdiri dari dua level yaitu pupa terbungkus kokon dan pupa tidak terbungkus kokon sehingga total semua perlakuan adalah enam perlakuan. Setiap perlakuan diulang 20 kali sehingga diperlukan 120 kokon.
Faktor Kondisi pupa B1 B2
Lampu
A1 A1B1 A1B2 A2 A2B1 A2B2 A3 A2B1 A2B2 Dimana : A1 penyinaran lampu 8W
A2 penyinaran lampu 18W A3 penyinaran lampu 23 W
B1 kondisi pupa terbungkus kokon B2 Kondisi pupa tidak terbungkus kokon
intensitas cahaya. Kelembaban udara di dalam kandang diatur menjadi 80%. Semua hasil pengamatan dicatat di dalam tabel pengamatan.
Efektivitas perlakuan dihitung dengan cara membandingkan jumlah imago yang keluar dari kokon/pupa setelah diberikan perlakuan dengan jumlah keseluruhan kokon/pupa pada tiap perlakuan.
Efektivitas perlakuan =
Jumlah imago yang keluar dari kokon/pupa pada tiap perlakuan
x 100% Jumlah keseluruhan kokon/imago pada tiap
perlakuan
Analisis Data
HASIL
Pengamatan Siklus Hidup C. trifenestrata
Telur C. trifenestrata pada saat pertama kali diletakkkan oleh betina berwarna putih dan biasanya diletakkan berderet rapi dengan arah yang teratur pada tepi daun (permukaan bawah daun) (Gambar 8). Bintik transparan (pada salah satu ujung yang merupakan tempat keluarnya larva) seluruhnya menghadap ke bagian pinggir daun. (Gambar 8). Bila telur tersebut fertil maka akan terjadi perubahan warna dari putih menjadi putih kelabu. Perubahan warna yang jelas mulai nampak pada hari ketujuh pengamatan sejak telur tersebut diletakkan oleh induknya. Bila telur tersebut steril maka akan terlihat perubahan bentuk menjadi cekung pada bagian tengahnya, sedang warnanya tetap dan tidak berubaha menjadi putih kelabu.
Gambar 8 Peletakkan telur C. trifenestrata
Telur C. trifenestrata yang fertil akan keluar pada hari ke 8 sampai 10 setelah diletakkan. Larva instar pertama akan keluar dari telur melalui bintik transparan yang ada di salah satu ujung telur dengan memakan kulit telur (Gambar 9). Larva biasanya berada di bawah permukaan daun, sendiri atau seringnya bergerombol.
S i y L b p
Setelah 5 ha instar 2 berw yang mulai 1mm.
Larva instar berwarna ku panjang di s
ari larva inst warna kunin terlihat den
r 2 akan berg uning kemer eluruh tubuh
Gamb tar pertama ng dengan k ngan jelas (G
Gamba ganti kulit m
rahan denga h (Gambar 1
bar 9 Larva akan bergan kepala cokla
Gambar 10)
ar 10 Larva menjadi larva an kepala c 1). Panjang
a instar 1 nti kulit men at, dan ramb
. Panjang la
a instar 2 a instar 3 set
oklat dan r larva 10 mm
njadi larva i but-rambut b arva 5 – 6 m
telah 5 hari. ambut-ramb m dan lebar 1
instar 2. Lar berwarna put
mm dan leb
Larva instar but putih ya
1 – 1.5 mm. rva
tih bar
S b d ( L m d ( k l
Setelah 5 ha berwarna m diseluruh tu (Gambar 12) Larva instar memiliki cir dan rambut-(Gambar 13 karena lebih larva 20 mm
ari larva inst merah denga
ubuh. Terda ). Panjang la
r 4 akan berg rri yang sam -rambut putih
) berbeda da h rakus, sehi m dan lebarny
Gamba tar 3 akan be an kepala m apat garis h arva 12 – 15
Gamba ganti kulit m ma dengan in
h panjang di alam hal uku ingga dapat ya 5 - 7 mm
ar 11 Larva erganti kulit merah dan hitam melin mm dan leb
ar 12 Larva menjadi larva star 4 yaitu b i seluruh tub uran. Larva i menyebabk .
a instar 3 menjadi lar rambut-ram ngkar hitam bar 2 mm.
a instar 4 a instar 5 set
berwarna m buh. Instar 4 instar 5 relat kan tanaman
rva instar 4. mbut putih m di sepanj
telah 5 hari. erah dengan 4 (Gambar 12
tif lebih besa menjadi gu
Larva instar yang panja ang abdom
Larva instar n kepala mer
t b m 5 m d h Pada tempat untu bukan poho masih dapat 5 - 6 hari. K mulutnya (G daun tanam hingga 80 ko
a pengamatan uk membentu on inang un
terbentuk m Kokon dibuat Gambar 14). man atau pad
okon.
Gamba n di alam, m uk pupa. Te ntuk larvany meskipun tida
t dari serat-s Biasanya ko da ranting t
Gambar
ar 13 Larva menjelang m empat memb ya. Pupasi b ak dalam ko serat halus b okon berada tanaman. Ju
r 14 Kokon
a instar 5 masa prapupa
bentuk pupa berlangsung okon. Masa p
erwarna kun secara send umlah kokon
n dan pupa
a, larva men a bisa berup di dalam k pupasi berlan ning yang di diri atau berk n yang berk
k i m m B t m r t b B c b p b s Imag kokon pada imago. Sete mengemban melakukan Betina yang telur per in meletakkan rata-rata ada tubuh, bentu bentuk ante Bentuk abdo coklat keme buah bintik pada sayap buah bintik sedangkan a
go akan kelu malam hari elah keluar ngkan sayapn
kopulasi. K g telah dikaw
duk rata-rat telur, namu alah lima h uk abdomen nna (Gamba omen betina erahan. Saya transparan p belakang. S k transparan antena betina
uar dari koko i. Salah satu dari kokon, nya hingga s Kopulasi bisa wini jantan ta 200 butir n telurnya t hari. Imago
n, warna sa ar 15). Uku a lebih lebar ap betina be
pada sayap d Sayap jantan n pada saya a bipektinate
Gambar 15
on setelah 14 u ujung kok
, imago aka sempurna. B a tetap berla
akan meleta r. Betina ya idak fertil. M
jantan dan ayap, jumla uran tubuh b
dari jantan. rwarna cokl depan dan s n berwarna l
ap depan. e.
5 Imago jant
4 - 16 hari. B on berluban an berdiam Biasanya jant
angsung hin akkan telur ang tidak di Masa hidup
betina dapa h bintik tra betina relati
Tubuh imag lat terang hi satu bintik tr lebih terang Tipe antena
tan dan betin
Biasanya ima ng sebagai ja diri dan se tan dan betin ngga pagi d
sehari setela ikawini jant
imago janta at dibedakan ansparan pa if lebih besa
go ditutupi s ingga gelap. ransparan ya g dari betina a jantan ku
na
ago keluar d alan keluarn ecara perlah na akan sege dan siang ha ahnya. Juml tan juga dap
an dana beti n dari ukur ada sayap d ar dari janta sisik berwar
Gambar 16 C.trifenestrata yang sedang kopulasi
Jumlah C.trifenestrata di alam sangat dibatasi oleh pengendali biologinya seperti predator dan parasitoid. Dalam masa pengamatan di alam, predator larva yang teramati adalah burung. Terdapat parasitoid bagi telurnya yaitu Schelio sp (Hymenoptera: Scelionidae) dan parasitoid pupa yaitu Xanthopimpla sp (Hymenoptera: Ichneumonidae) dan (Diptera: Tachinidae).
Studi Perkembangan Embrio C. trifenestrata
Dalam pengamatan embrio C. trifenestrata, dilakukan uji pendahuluan mengenai pembuatan preparat embrio dengan menggunakan berbagai larutan kimia: HCl 37%, NaOH 5 M, laktofenol, dan asam asetat memperlihatkan bahwa larutan kimia yang paling bagus membuat preparat embrio ini adalah larutan NaOH 5 M dengan lama perendaman tiga hari. Larutan HCl 37% membuat telur menjadi berwarna lebih gelap dan tidak terlihat transparan. Laktofenol dan asam asetat dapat membuat telur menjadi transparan, namun larutan NaOH 5 M lebih cepat membuat telur menjadi transparan.
Hari pertama Hari kedua
Hari ketiga Hari keempat
Hari kelima Hari keenam (embrio dalam telur)
Hari ke-6 (dibuka dari kulit telur) Hari ketujuh (dibuka dari kulit telur)
1 1
1 mm 1 mm
H G H a b b n h m e a t m 2 s p Hari kedelap Gambar 17
Pada Hal ini terli adalah pros blastoderm’ blastoderm’ ‘Nuclei’ yan nantinya aka
Pada hari pertam menghasilka ektoderm, m atau masih t
Pada terlihat jelas menyebabka 2007). Bagia Selan segmentasi proses organ pan (dibuka Perkemban a pengamatan ihat dari ben ses pembela
yaitu kump akan bermi ng berada p an berkemba a embrio har a. Pada sta an bakal sal mesoderm da erjadi perger a embrio ha
s. Diduga p an posisi ata
an anterior d njutnya pad
dan pembe nogenesis jug
dari kulit tel ngan embrio
n embrio ha ntuk kuning ahan nukleu pulan masa n
igrasi ke ba pada bagian ang menjadi ri kedua ter dia ini didu luran pence an endoderm
rakan (blasto ari ketiga m proses blasto au kedudukan dan posterio
da embrio entukan stru
ga telah terja
lur)
C. trifenestr
ari pertama, d g telur dan us zigot. Ha
nukleus di d agian perifer n posterior a
‘germ band rlihat ‘germ uga mulai t ernaan setela m. Posisi em
okinesis). mulai terjadi
okinesis tela n embrio me or embrio me hari keem uktur ‘appen adi pada stad
rata pada ha
diduga telah perluasan ‘ asil dari ‘cl dalam sitopla r dan memb
akan memb d’.
band’ yang terjadi prose ah terbentuk mbrio pada s
proses seg ah selesai b enjadi tetap enjadi stabil mpat, mulai ndages’ pad
dia ini.
ari 1 - 8
h terjadi pros ‘germ band leavage’ ad asma yang sa
entuk lapisa entuk sel-se
g lebih luas es diferensia k tiga ‘germ saat ini masi
gmentasi me berlangsung. (Dow et al.
kedudukann nampak da tubuh em
ses ‘cleavag ’. ‘Cleavag dalah ‘sinsit
ama. ‘Sinsit an blastoder el kutub ya
dibandingk asi yang ak m layer’ ya ih belum tet
Pada embrio hari kelima, organ-organ dalam diduga mulai nampak terbentuk sebagai kelanjutan dari organogenesis. Bagian tungkai juga mulai bisa diamati, begitu juga dengan bagian kepala yang mulai jelas dapat diamati.
Pada embrio hari keenam, bentuk embrio semakin lengkap, bahkan struktur scoli mulai terlihat dengan jelas. Pada bagian kepala berwarna lebih gelap. Struktur larva telah lengkap.
Pada embrio hari ketujuh dan kedelapan merupakan proses penyempurnaan bentuk larva, antara lain terlihat pada bagian kepala, pada embrio hari ketujuh terjadi perubahan warna kepala menjadi lebih gelap dan semakin berwarna gelap pada embrio hari kedelapan. Terlihat bagian kepala semakin mengeras berkaitan dengan fungsinya untuk mendorong kulit telur saat proses penetasan terjadi. Struktur kepala yang kuat diperlukan untuk membantu keluarnya larva dari telur.
Penundaan Penetasan Telur C. trifenestrata dengan Penyimpanan dalam Suhu 8 0C
Pada percobaan ini dilakukan penyimpanan telur pada suhu 8 0C dengan lama penyimpanan 3, 7, 14, 21, dan 28 hari. Hasil percobaan menyatakan bahwa telur yang dapat menetas adalah telur pada lama penyimpanan tiga dan tujuh hari. Sedangkan telur pada lama penyimpanan 14, 21 dan 28 hari, tidak ada yang menetas. Oleh karena itu data yang dianalisis adalah data penyimpanan tiga dan tujuh hari saja. Hasil percobaan dapat dilihat pada Tabel 1.
penurunan angka prosentase penetasan telur seiring dengan penambahan waktu penyimpanan.
Tabel 1 Keberhasilan perkembangan embrio dan penetasan telur Umur telur
(hari)
Prosentase telur
Lama penyimpanan Kontrol (menetas setelah hari ke-8) 3 hari 7 hari
1 0 0 71.63 ± 9.73
2 0 0 3 0 0 4 0 0 5 8.77 a ± 3.54 1.64 a ± 0.48
6 24.86 b ± 8.42 18.40 b ± 9.62 7 16.52 a ± 5.36 0
8 0 0
Angka yang diikuti oleh huruf yang tidak sama berbeda nyata pada taraf uji 5% (Duncan)
Pada telur yang tidak disimpan dalam lemari pendingin atau dalam hal ini sebagai kontrol, menetas setelah hari ke- 8 dengan prosentase penetasan sebesar 71.63%. Prosentase kontrol ini lebih tinggi dibandingkan dengan prosentase telur yang diberikan perlakuan.
Hasil uji anova yang dilakukan pada data tersebut di atas menunjukkan bahwa terdapat perbedaan yang nyata pada faktor umur telur (p = 0 < 0.05), lama penyimpanan (p = 0.01 < 0.05) dan interaksi antara kedua faktor (p = 0.02 < 0.05). Kombinasi yang memberikan respon terbaik yaitu stadia umur telur enam hari dengan lama penyimpanan dalam ruang pendingin (8 0C) selama tiga hari.
Pemecahan Dormansi Pupa C. trifenestrata
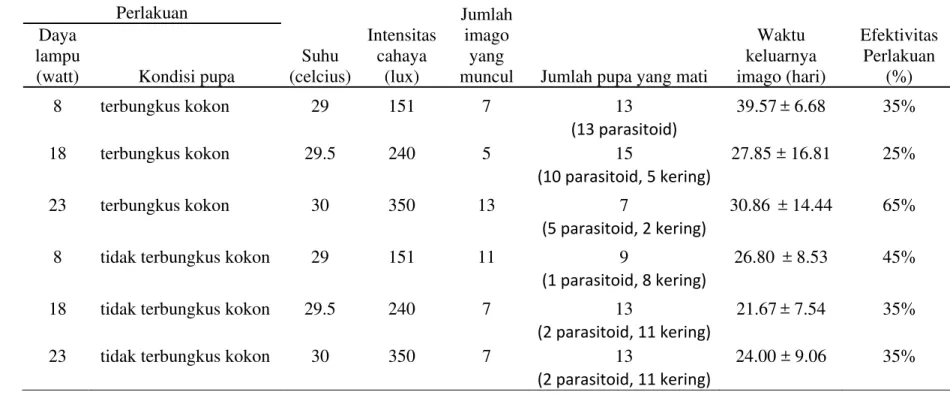
Dari percobaan didapatkan hasil bahwa dari enam perlakuan, perlakuan pencahayaan dengan lampu 23 watt dan dengan kondisi pupa tidak terbungkus kokon memberikan keberhasilan pemecahan dormansi pupa sebesar 65% sedangkan perlakuan yang lainnya di bawah 50% (Tabel 2). Sejumlah kokon/pupa tidak keluar menjadi imago karena ditemukan mati oleh parasitoid (Ichneumonidae dan Sarchopagidae) (Gambar 19) dan kekeringan (Gambar 20).
Gambar 18 Parasitoid pada pupa: A. Ichneumonidae; B. Sarchopagidae
Gambar 19 Pupa yang mati kekeringan
Bila kokon atau pupa yang mati karena parasitoid tidak dimasukkan dalam perhitungan maka efektivitas perlakuan menjadi seperti pada Tabel 3. Penyinaran lampu 8 watt dengan kondisi pupa terbungkus kokon adalah pelakuan yang paling
efektif (100%), karena dari semua kokon yang diberi perlakuan semuanya keluarnya imago.
Perlakuan yang memberikan rangsangan tercepat untuk keluarnya imago adalah perlakuan A2B2 yaitu penyinaran dengan lampu 18 watt terhadap pupa yang tidak terbungkus kokon dengan rata-rata waktu keluarnya imago 21.67 hari.
Intensitas cahaya tertinggi adalah pada kotak inkubasi dengan lampu 23 watt, sedang terendah pada lampu 8 watt. Demikian juga pada pengukuran suhu, nilai tertinggi pada kotak inkubasi dengan lampu 23 watt dan terendah pada lampu 8 watt. Terjadi peningkatan intensitas cahaya dan suhu seiring dengan meningkatnya ukuran watt lampu.
Tabel 2 Efektivitas perlakuan pada pemecahan dormansi pupa Perlakuan
Suhu (celcius)
Intensitas cahaya
(lux)
Jumlah imago
yang
muncul Jumlah pupa yang mati
Waktu keluarnya imago (hari)
Efektivitas Perlakuan
(%) Daya
lampu
(watt) Kondisi pupa
8 terbungkus kokon 29 151 7 13 39.57± 6.68 35% (13 parasitoid)
18 terbungkus kokon 29.5 240 5 15 27.85 ± 16.81 25% (10 parasitoid, 5 kering)
23 terbungkus kokon 30 350 13 7 30.86 ± 14.44 65% (5 parasitoid, 2 kering)
8 tidak terbungkus kokon 29 151 11 9 26.80 ± 8.53 45% (1 parasitoid, 8 kering)
18 tidak terbungkus kokon 29.5 240 7 13 21.67± 7.54 35% (2 parasitoid, 11 kering)
23 tidak terbungkus kokon 30 350 7 13 24.00 ± 9.06 35%
Tabel 3 Efektivitas perlakuan pada pemecahan dormansi pupa bila pupa yang mati karena parasitoid tidak masuk dalam perhitungan
Perlakuan
Suhu (celcius)
Intensitas cahaya
(lux)
Jumlah imago
yang muncul
Jumlah pupa yang
kering
Waktu keluarnya imago (hari)
Efektivitas Perlakuan
(%) Daya
lampu (watt)
Kondisi pupa
PEMBAHASAN
Siklus Hidup C. trifenestrata
Tahapan hidup C. trifenestrata terdiri dari telur, larva, pupa, dan imago. Telur yang fertil akan menetas setelah hari kedelapan, sedang larva terdiri dari lima instar dengan lama masing – masing instar berlangsung selama kurang lebih lima hari. Masa pupa berlangsung sekitar 14 – 16 hari. Imago C. trifenestrata mengalami rudimenter pada alat pencernaannya. Oleh karena itu imagonya hanya bertahan kurang lebih lima hari.
Keberadaan C. trifenestrata di alam dibatasi oleh predator dan parasitoid. Pemeliharaan di dalam ruangan akan mengurangi resiko serangan predator atau parasitoid. Keberhasilan pemeliharaan dalam ruangan ditentukan oleh ketepatan kondisi pemeliharaan meliputi suhu dan kelembaban ruangan. Suhu dan kelembaban ruang pemeliharaan adalah 28 - 29 0C dan 80 %.
Studi Perkembangan Embrio C. trifenestrata
yang lain akan membentuk serosa yang selanjutnya akan menjadi bagian dari ‘yolk sac’. Sel-sel serosa akan mengelilingi ‘germ band’ dan membungkus embrio membentuk membran amnion.
Tahapan berikut yang terjadi adalah tahap diferensiasi yaitu meliputi bentuk awal tubuh, penentuan anterior posterior, pembentukan segmen tubuh hingga spesialisasi struktur dan ‘appendages’. Studi mengenai penentuan longitudinal axis yang dilakukan pada Drosophila (Drosophilidae) menyatakan bahwa penentuan ini terkait dengan maternal (Chapman 1998). Saat oocyte masih berada di dalam ovarium, troposit memproduksi spesifik mRNA yang ditransfer ke oocyte dan terakumulasi pad