Informasi Dokumen
- Penulis:
- Didah Nur Faridah
- Pengajar:
- Prof. Dr. Ir. Dedi Fardiaz, MSc
- Dr. Ir. Nuri Andarwulan, MS
- Dr. Ir. Titi Candra Sunarti, MS
- Sekolah: Institut Pertanian Bogor
- Mata Pelajaran: Ilmu Pangan
- Topik: Perubahan Karakteristik Kristalin Pati Garut Dalam Pengembangan Pati Resisten Tipe III
- Tipe: disertasi
- Tahun: 2011
- Kota: Bogor
Ringkasan Dokumen
I. PENDAHULUAN
Bagian ini menjelaskan latar belakang penelitian mengenai pati garut dan pentingnya pati resisten tipe III (RS3) dalam konteks pangan fungsional. Pati resisten memiliki banyak manfaat kesehatan, termasuk sebagai prebiotik dan dalam pengelolaan berat badan. Penelitian ini bertujuan untuk memodifikasi pati garut agar meningkatkan kadar RS3, dengan fokus pada proses autoclaving-cooling, hidrolisis asam, dan debranching. Hal ini relevan dengan tujuan pendidikan untuk memahami pengolahan pangan dan aplikasi dalam industri makanan.
1.1. Latar Belakang
Latar belakang penelitian menekankan pentingnya pangan fungsional dalam diet modern. Pati resisten, terutama RS3, berperan dalam kesehatan pencernaan dan pengendalian berat badan. Penelitian ini mengkaji potensi pati garut sebagai sumber RS3 yang dapat dimanfaatkan untuk meningkatkan nilai gizi dan kesehatan masyarakat.
1.2. Perumusan Masalah
Permasalahan yang dihadapi adalah rendahnya kadar pati resisten dalam pati garut secara alami. Penelitian ini berfokus pada bagaimana proses modifikasi dapat meningkatkan kadar RS3, sehingga memberikan solusi untuk meningkatkan kualitas pangan fungsional yang berbasis sumber lokal.
1.3. Tujuan Penelitian
Tujuan utama dari penelitian ini adalah untuk mengeksplorasi berbagai metode modifikasi pati garut yang dapat meningkatkan kadar RS3. Ini mencakup autoclaving-cooling, hidrolisis asam, dan debranching, serta analisis dampaknya terhadap sifat fisik dan kimia pati.
1.4. Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan kontribusi signifikan dalam pengembangan pangan fungsional berbasis pati garut. Hasilnya dapat digunakan sebagai dasar untuk penelitian lebih lanjut dalam aplikasi RS3 di berbagai produk makanan, mendukung tujuan pendidikan dalam inovasi pangan.
II. TINJAUAN PUSTAKA
Bagian ini membahas literatur yang relevan mengenai struktur pati, jenis-jenis pati resisten, serta proses modifikasi yang dapat dilakukan. Tinjauan pustaka ini penting untuk memberikan dasar teoritis bagi penelitian dan mendukung pemahaman mahasiswa mengenai interaksi antara struktur kimia dan sifat fungsional pati.
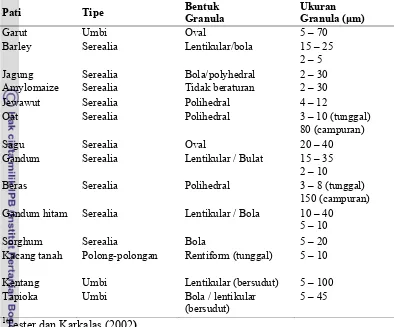
2.1. Umbi Garut
Deskripsi mengenai tanaman garut dan karakteristik umbinya. Pati garut memiliki potensi sebagai bahan baku pangan fungsional. Informasi ini penting bagi mahasiswa untuk memahami sumber daya lokal dan aplikasinya dalam industri pangan.
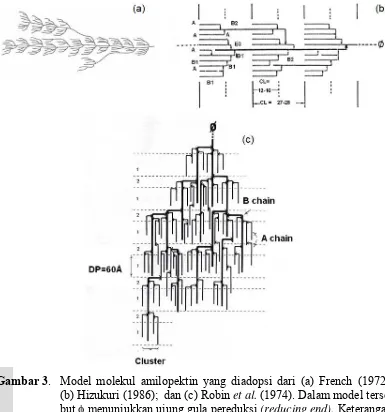
2.2. Struktur Amilosa dan Amilopektin
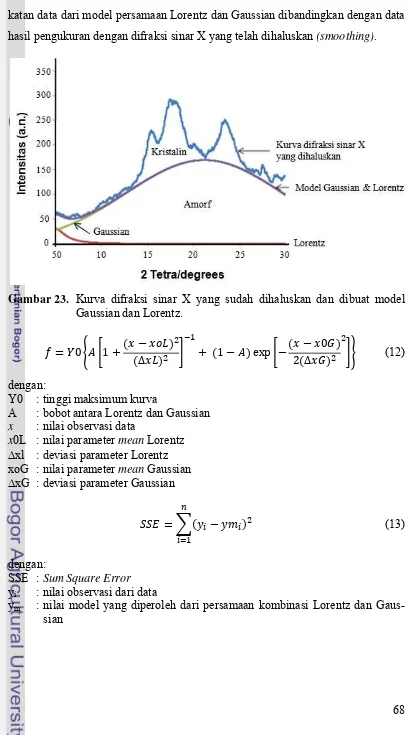
Penjelasan mengenai perbedaan struktur amilosa dan amilopektin, serta bagaimana struktur ini mempengaruhi sifat gelatinisasi dan retrogradasi pati. Pemahaman tentang struktur ini sangat penting dalam konteks modifikasi pati untuk meningkatkan RS3.
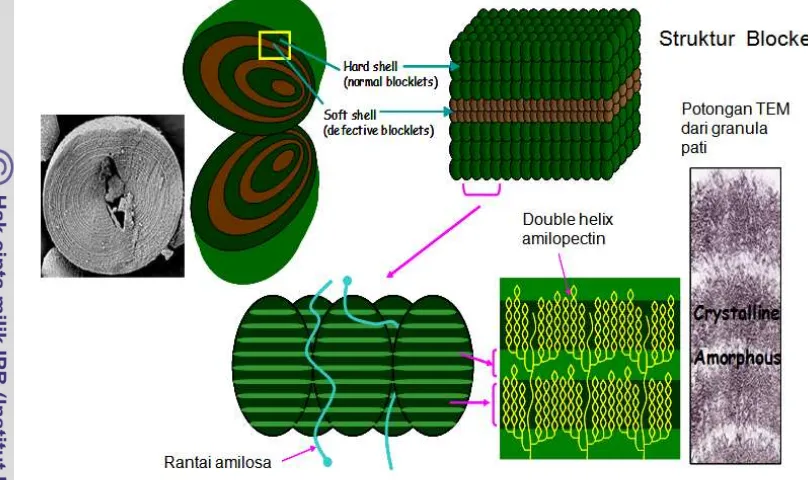
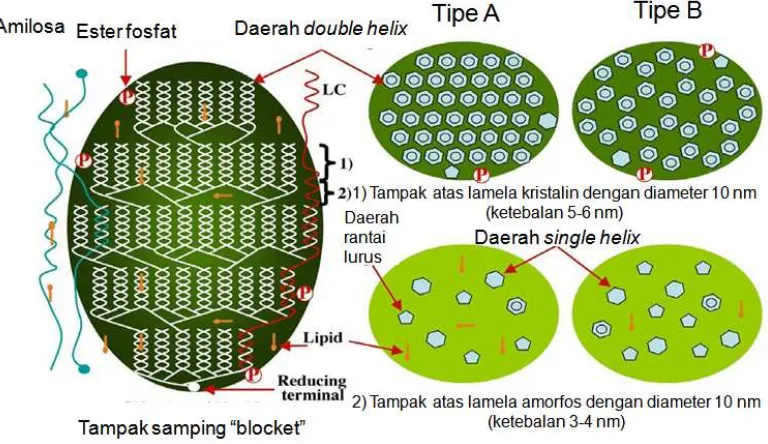
2.3. Model Struktur Granula Pati
Model struktur granula pati memberikan wawasan tentang bagaimana pati disimpan dan dimodifikasi. Ini relevan untuk pengembangan metode ekstraksi dan modifikasi dalam penelitian.
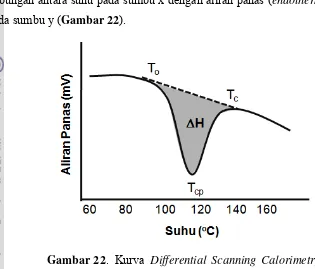
2.4. Pengaruh Proses Pengolahan terhadap Gelatinisasi dan Retrogradasi Pati
Membahas bagaimana proses pengolahan mempengaruhi sifat gelatinisasi dan retrogradasi pati. Ini penting untuk memahami bagaimana modifikasi dapat meningkatkan kadar RS3.
2.5. Pati Resisten
Diskusi mengenai berbagai jenis pati resisten dan mekanisme pembentukannya. Pengetahuan ini penting untuk mendukung pengembangan produk pangan fungsional yang lebih sehat.
III. METODE PENELITIAN
Metode penelitian yang digunakan dalam studi ini mencakup tahapan ekstraksi, modifikasi, dan analisis karakteristik pati garut. Penjelasan metodologi ini memberikan pemahaman praktis kepada mahasiswa tentang teknik-teknik yang digunakan dalam penelitian pangan.
3.1. Waktu dan Tempat Penelitian
Informasi tentang lokasi dan durasi penelitian yang memberikan konteks untuk pengumpulan data dan hasil yang diperoleh. Ini penting untuk memahami variabel lingkungan dalam penelitian.
3.2. Bahan dan Alat
Daftar bahan dan alat yang digunakan dalam penelitian, memberikan mahasiswa wawasan tentang peralatan laboratorium yang relevan dalam penelitian pangan.
3.3. Tahapan Penelitian
Rincian langkah-langkah yang diambil dalam penelitian, termasuk proses ekstraksi dan karakterisasi pati. Ini memberikan panduan praktis bagi mahasiswa untuk menerapkan metode serupa dalam penelitian mereka.
3.4. Prosedur Analisis
Metode analisis yang digunakan untuk mengukur perubahan dalam karakteristik pati garut setelah modifikasi. Pengetahuan ini penting untuk evaluasi hasil penelitian dan validasi data.
IV. HASIL DAN PEMBAHASAN
Bagian ini menyajikan hasil penelitian dan analisis terhadap data yang diperoleh. Diskusi mengenai hasil memberikan wawasan tentang implikasi dari modifikasi pati garut dalam pengembangan pangan fungsional.
4.1. Ekstraksi dan Karakterisasi Pati Garut
Hasil dari proses ekstraksi pati garut dan karakteristik kimianya. Ini penting untuk menunjukkan efektivitas metode yang digunakan dalam penelitian.
4.2. Penentuan Kondisi Proses Modifikasi Pati Garut
Analisis kondisi optimal untuk modifikasi pati garut, memberikan panduan bagi penelitian lebih lanjut dalam aplikasi RS3.
4.3. Pengaruh Perlakuan Modifikasi terhadap Karakteristik Pati Garut
Diskusi mengenai bagaimana berbagai perlakuan modifikasi mempengaruhi karakteristik pati, relevan untuk pengembangan produk pangan yang lebih sehat.
V. SIMPULAN DAN SARAN
Bagian ini menyimpulkan temuan penelitian dan memberikan rekomendasi untuk penelitian lebih lanjut. Ini penting untuk mengarahkan mahasiswa dalam memahami dampak dari penelitian mereka dan langkah selanjutnya dalam pengembangan ilmu pangan.
5.1. Simpulan
Simpulan dari penelitian ini menekankan keberhasilan dalam meningkatkan kadar RS3 dari pati garut melalui modifikasi. Ini memberikan kontribusi pada pengembangan pangan fungsional.
5.2. Saran
Rekomendasi untuk penelitian di masa depan, termasuk eksplorasi lebih lanjut tentang aplikasi RS3 dalam berbagai produk pangan. Ini penting untuk mendorong inovasi dalam bidang pangan.
Referensi Dokumen
- Study on outer chains from amylopectin between immobilized and free debranching enzymes ( Sunarti TC, Nunome N, Yashio, Hisamatsu M )
- Starch and enzyme-resistant starch from high-amylose barley ( Szczodrak J, Pomeranz Y )
- Starch as Ingredients: Manufacture and Applications ( Taggart P )
- Branching of amylase by the branching iso-enzymes of maize endosperm ( Takeda Y, Guan HP, Preiss J )
- Examination of the structure of amylase and amylopectin by fluorescent labeling terminal ( Takeda Y, Hanashiro M )