www.elsevier.com / locate / livprodsci
Use of digestible fibre in replacement to available
carbohydrates
Effect on digestion, rate of passage and caecal fermentation
pattern during the growth of the rabbit
*
Thierry Gidenne , Rachel Bellier
Institut National de la Recherche Agronomique, Station de Recherches Cunicoles, BP 27, 31326 Castanet-Tolosan, France Received 14 September 1998; received in revised form 20 April 1999; accepted 10 May 1999
Abstract
The dietary replacement of available carbohydrates (mainly starch) by digestible fibre was assessed on rabbit growth and digestion. Two diets supplying hemicellulose (diet HC) or a mix of hemicellulose and pectins (HC1P) were compared to a
control diet having a higher starch content, but a similar content in low digested fibre (lignocellulose ADF5155 g / kg). Three groups of 17 rabbits were allocated at weaning (28 d) to each diet. Nine animals of each group were cannulated to follow the caecal microbial activity from 45 to 70 d old. Compared to control, rabbits fed HC or HC1P increased feed intake (110%), but only HC1P resulted in a higher weight gain and in a similar feed conversion (mean 2.00), particularly
during the postweaning period (4–6 wk old). Transit of liquids was similar among diets, either in the caeco–colic segment or in the whole tract. HC1P diet gave short caeco–colic transit for particles (13.1 h), and also the highest caecal SCFA level
(62.2 mM / l) and pool (3.8 mM), while HC and control diet led to similar caecal fermentation (mean556.5 and 2.25, respectively for SCFA level and pool). SCFA concentration increased according to the mean intake of ‘‘hemicellulose1
pectins’’, suggesting that microbial activity and fibre degradation were more affected by the nature of the cell wall than by the caeco–colic retention time. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Rabbit digestion; Caecal fermentation; Starch; Hemicellulose; Pectins; Dietary fibre
1. Introduction hindgut flora and to ensure digestive (motility and transit) and metabolic functions of the host (Mathers,
´
For monogastric species and particularly for the 1991; Carre, 1992; Edwards and Parrett, 1996). growing rabbit, supplying dietary fibre corresponds Digestive consequences of fibre are determined as to two main roles: to provide nutrients for the well by the quantity of dietary fibre consumed as by the degradability of cell wall components in relation to their physico–chemical structure.
*Corresponding author Tel.: 133-5-6128-5103; fax: 1
33-5-Previous studies on growing rabbit have addressed 6128-5319.
E-mail address: [email protected] (T. Gidenne) the problem of increasing the intake of low digestible 0301-6226 / 00 / $ – see front matter 2000 Elsevier Science B.V. All rights reserved.
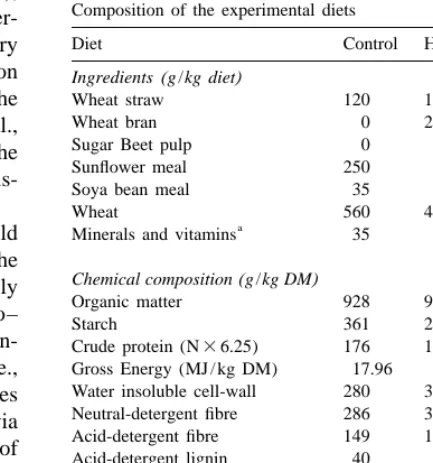
Table 1 fibre to prevent digestive troubles (mainly diarrhoea),
Composition of the experimental diets but associated to a negative effect on growth
per-Diet Control HC HC1P
ˆ
formance (Ma ı tre et al., 1990). Conversely, dietary
starch could also play a substantial role in interaction Ingredients(g /kg diet)
with fibre, as it is incompletely hydrolysed in the Wheat straw 120 150 140
Wheat bran 0 270 50
small intestine of the weanling rabbit (Blas et al.,
Sugar Beet pulp 0 0 200
1988; Blas et al., 1994) causing alteration of the
Sunflower meal 250 50 50
caeco–colic digestion associated with digestive dis- Soya bean meal 35 98 162
turbances. Wheat 560 400 367
a
Therefore, the reduction of starch supply should Minerals and vitamins 35 32 31 be explored without excessive reduction in the
Chemical composition(g /kg DM) digestible energy and without modifying the supply
Organic matter 928 925 924
in low digested fibre fraction. Because the caeco– Starch 361 290 256 colic fermentation is relatively short (7–12 h; Giden- Crude protein (N36.25) 176 167 179 ne, 1997), the use of rapidly fermented fibre (i.e., Gross Energy (MJ / kg DM) 17.96 18.13 17.87
Water insoluble cell-wall 280 341 338 digestible fibre) replacing available carbohydrates
Neutral-detergent fibre 286 339 327 could be of interest to supply energy to the host via
Acid-detergent fibre 149 150 163 caecal microbial activity, without alteration of rate of Acid-detergent lignin 40 30 27
b
passage. The level of degradation for hemicellulose Hemicellulose 137 189 164 fractions (30–45%) and for pectins (70%) in the Uronic acids 27 18 59
c
Pectins 30 19 82
rabbit classes these cell wall constituents in the
a
‘‘digestible fibre’’ compared to low-digested fibres Contained: (g / kg, respectively for C, HC; HC1P diets)
calcium carbonate and bicalcium phosphate (21.0, 17.5, 18.0), such as lignocellulose (Gidenne et al., 1998a). The
DL-methionine (0.5, 1.5, 1.5),L-lysine (1.5, 1.0, 0.0) salt (7.0, 7.0, present study assessed two ways of starch
replace-6.5); and 5.0 g / kg diet of vitamin premix (retinol 540 mg / kg, ment either mainly through a supply of hemicellulose
cholecalcipherol 5 mg / kg, a-tocopherol 3.3 mg / kg, robenidine
or with a mix of hemicellulose and pectins. Further- 13.2 mg / kg). b
more, we aimed to develop a new approach to obtain Hemicellulose fraction estimated by difference between NDF and ADF.
a more comprehensive information on caecal
diges-c
Water insoluble pectins: calculated from tabulated data on tion, that included simultaneous in-vivo
measure-ingredients (IO7, 1993). ment of rate of passage, nutrient digestion and caecal
microbial activity (CMA), over a substantial period of the rabbit growth (6–10 wk old).
ADF), mainly provided by one source (wheat straw)
2. Materials and methods to avoid major change in the type of cellulose or lignin. The ratio digestible protein / digestible energy 2.1. Diets and feeding was maintained constant among the diets, by adjust-ing the incorporation of soya bean meal and wheat. Three experimental diets were prepared (Table 1)
in a pelleted form with raw material classically used 2.2. Animals, housing and experimental protocol in rabbit feed formulation, to obtain a variation in the
level of ‘‘digestible’’ fibre: a control diet, and two Three groups of 17 New Zealand white rabbits diets where a portion of starch was substituted either were allocated (after controlling the effect of litter by hemicellulose (evaluated as NDF-ADF) supplied origin) at weaning (28 d old) to one of the ex-mainly by wheat bran (HC diet), or by perimental diets. They were housed in individual ‘‘hemicellulose1pectins’’ through a supply in beet metabolism cages (55340 cm) and submitted to a
pulp (HC1P diet). In addition, the three diets have 12 h light (7:00 to 19:00) and 12 h dark schedule.
rabbits of each group were surgically fitted with whole tract was calculated according to MRT5M t /
i i T-cannula in the caecum as described by Bellier et M , where t was the time that has elapsed betweeni i al. (1995). Sampling of caecal digesta and diges- marker administration and the ith defecation and Mi tibility measurements were performed after a one was the quantity of marker excreted in faeces. MRT week recovery period. Apparent faecal digestibility includes the minimal transit time (TTm), which was coefficients were calculated from individual and the time that has elapsed between dosing and the first daily total collection during six consecutive days (42 marker appearance in the faeces. TTm reflects the to 48 d of age). retention time of digesta without a delay in the Samples of caecal digesta were individually col- mixing compartments, so it represents the rate of lected in vivo via the cannula at 45, 50, 60 and 70 passage in the tubular segment of the tract, i.e., days of age. Rabbits were placed in a special mainly in the small intestine and also in the distal hammock for 10 min, and digesta drained by gravity colon (Gidenne, 1994).
in sampling tubes (Gidenne and Bellier, 1992). Caecal volume was determined in-vivo according Sampling time was fixed at the end of the caecot- to the following marker dilution procedure, on six
51
rophy period (12:00) when inter-individual varia- rabbits per group. The concentration of chromium-tions are lower (Bellier et al., 1995). After sampling, EDTA was determined in samples of caecal digesta the caecal material was immediately subjected to the collected 1, 2, 4 and 6 h after the introduction of the determination of pH. Portions of caecal digesta marker dose (between 11:30 and 12:00). A linear sample (5 to 10 g fresh matter) were placed in tubes adjustment of the decrease in marker concentration containing (2%, v / v) H PO3 4 or H SO2 4 storage was performed. The caecal volume was then ob-solution (1 and 2 ml / tube), respectively for further tained by extrapolating to zero time the curve of analyses of short chain fatty acids (SCFAs) and tracer concentration against time (Leng and
¨
ammoniacal nitrogen (NH ), and stored at 2258C. Hornicke, 1976), and expressed in g of fresh matter.
3 51
A solution of chromium-EDTA and cell wall The caecal pool of SCFAs was calculated by
multip-141
particles labelled with cerium (Gidenne, 1994) lying the SCFA concentration (mM / l) and the liquid were used as liquid- and solid-phase markers. Cell content of the caecum (in ml, obtained from caecal wall particles were prepared from each diet, and volume and dry matter determination).
corresponded to the NDF residue after sieving on a
0.05 mm screen. Digesta mean retention time of 2.3. Analytical methods solid and liquid phase was measured on six
cannu-lated animals per group, by following the kinetics of Dry matter was determined on feeds and faeces by the faecal excretion of the two markers, using an heating for 24 h at 1038C. Organic matter (OM) was
automatic faecal sampler (Automatic Sampler; API, determined by ashing at 5508C for 5 h.
Measure-Castanet, France). First, rate of passage between ments of fibre fractions (neutral detergent fibre: caecum and rectum was recorded during a five-day NDF, acid detergent fibre: ADF, acid detergent period (from 48 to 52 d old), following the adminis- lignin: ADL) were made according to Van Soest et al. tration (at 11:30) of the two markers in the caecal (1991) using an amylolytic pre-treatment with a cannula. Then, rate of passage in the whole tract was thermostable amylase, and water insoluble cell walls
´
obtained on the same cannulated rabbits, during a were obtained according to Carre and Brillouet six-day period (from 53 to 57 d old) following the (1989). Nitrogen was determined by DUMAS com-oral administration (at 8:30) of markers. A dose of bustion method using Leco apparatus (Model FP-labelled particle corresponded to 100 mg DM 428, Leco Corp., St. Joseph, MI, USA), and con-(activity52 kBq) and was given orally using a verted to crude protein using the factor 6.25. Gross
by the m-phenylphenol method (Blumenkrantz and 3. Results
Asboe-Hansen, 1973). Starch was hydrolysed
en-zymatically and the resultant glucose was measured According to the objectives of the formulation, the by using the hexokinase (EC 2.7.1.1)-glucose-6- three experimental diets were similar in their levels phosphate dehydrogenase (NAD) (EC1.1.1.49) sys- of lignocellulose (meanly 155g ADF / kg, Table 1).
¨
tem (Boehringer Mannheim). SCFAs were analysed They differed mainly by their level of starch (high by gas–liquid chromatography (CP9000, Chrom- for control) in balance with the hemicellulose or pack, Middelburg, The Netherlands) on a semi-capil- pectin level (high for HC and HC1P), although the
lary column (Bellier, 1994). total fibre level (NDF or WICW) differed slightly (10%) among diets.
2.4. Statistical analyses
3.1. Effect of the cannulation and of the diet on Rabbits having an abnormally low feed intake (3
intake and growth
SD under mean) during the measurements were discarded from the statistical analyses. A two ways
Among the six groups of rabbits, only one animal analysis of variance was done using the general
receiving HC diet died because of an acute diarrhoea linear model ‘‘GLM’’ procedure (Statistical Analysis
(at 48 d of age). Following the implantation of the System, 1988), with the effect of cannulation and
cannula the rabbits recovered a normal feed intake diets and interaction between diet and cannulation as
within four or five days, so that from 28 to 45 d of main effects for the parameters of intake and growth.
age the daily feed intake was only 10% lower for Digestibility and rate of passage data were examined
cannulated animals. This resulted in a lower live by one-way analysis of variance (ANOVA).
weight gain (230%) for cannulated animals
com-The data on the caecal content composition (Table
pared to control (Table 2). From 45 to 70 days old, 6 and Fig. 1) were examined to evaluate the effect of
the daily feed intake and the weight gain did not the age, diet and age3diet interactions. The effect
differ significantly among cannulated or control of the individual was controlled in this model, to
group. In addition, interaction between the effect of take into account that the measurements (four
sam-the diet and sam-the group of rabbits was not significant ples at four successive ages) were repeated on the
whatever the parameter. same animals. The significance of the effect of the
Dietary treatments affected the intake as well as diet was thus calculated using the mean square of the
the weight gain, particularly during the two weeks intra-individual variations as an error term (split-plot
after weaning. The feed intake was significantly design). Least-square means comparisons were
per-higher (110%) for HC and ‘‘HC1P’’ groups from
formed using the PDIFF test of GLM. Data from
four to six weeks of age, but resulted in a sig-unhealthy animals or animals having an abnormal
nificantly higher weight gain only for ‘‘HC1P’’
SCFA pattern (i.e., with a propionate / butyrate ratio
group (Table 2). Consequently the feed conversion over 2) were not included in the analysis. The in
was not significantly different (P50.29) among the
vivo SCFA pattern from 6 to 10 weeks of age was
three groups for the period 4–6 wk (mean 2.00), but statistically analysed after pooling the results from
it was significantly lower (P50.031) for the control
the three diets, as no interaction between the diet and
group compared to HC1P during the 6–10 weeks
the age of the rabbit was found. In addition, the
old period (respectively, 2.73 vs. 3.06). effect of age on CMA was analysed according to the
GLM procedure (linear model). The partial
correla-tion coefficients between the parameters of the caecal 3.2. Nutrient intake and digestibility fermentation and the digestion of the nutrients (over
Table 2
Effect of caecal cannulation and of dietary treatment on feed intake and growth 1
Effect of the cannulation Effect of diets Statistical difference 2
n523 n524 n516 n515 n516 Cannulation
Control Cannulation Control HC HC1P RMSE Cannulation Diet 3Diet
Live weight(g)
Implantation of a caecal cannula at five weeks old. 2
Number of rabbits.
RMSE5Root mean square error.
a, b: Means, between diets, having a common superscript did not differ significantly at the level P50.05.
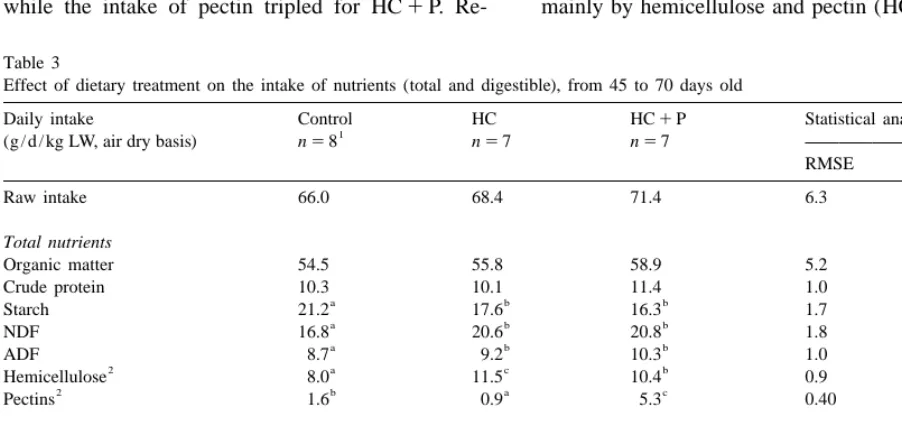
the dietary ADF level was similar, the ADF intake versely the starch intake is reduced by around 20% increased by 20% for HC1P group (Table 3). for these two groups.
Compared to control group, the intake of hemicellul- Organic matter digestibility did not differ sig-ose increased by four units (140%) for HC group, nificantly from control when starch is replaced
while the intake of pectin tripled for HC1P. Re- mainly by hemicellulose and pectin (HC1P group),
Table 3
Effect of dietary treatment on the intake of nutrients (total and digestible), from 45 to 70 days old
Daily intake Control HC HC1P Statistical analysis
1
(g / d / kg LW, air dry basis) n58 n57 n57
RMSE P.F
Raw intake 66.0 68.4 71.4 6.3 0.25
Total nutrients
Organic matter 54.5 55.8 58.9 5.2 0.29
Crude protein 10.3 10.1 11.4 1.0 0.053
a b b
Starch 21.2 17.6 16.3 1.7 ,0.001
a b b
NDF 16.8 20.6 20.8 1.8 ,0.001
a b b
ADF 8.7 9.2 10.3 1.0 0.004
2 a c b
Hemicellulose 8.0 11.5 10.4 0.9 ,0.001
2 b a c
Pectins 1.6 0.9 5.3 0.40 ,0.001
Digestible nutrients
Organic matter 38.9 37.2 41.1 3.7 0.20
Crude protein 8.6 8.5 9.4 0.9 0.18
a a b
NDF 4.2 4.4 6.4 0.8 ,0.001
a a b
ADF 1.1 0.9 1.8 0.4 0.001
2 a a b
Hemicellulose 3.1 3.4 4.6 0.4 ,0.001
1
a, b: See Table 2. 2
Table 4
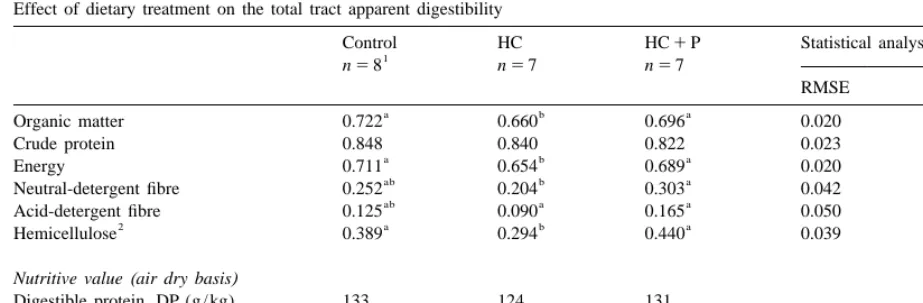
Effect of dietary treatment on the total tract apparent digestibility
Control HC HC1P Statistical analysis
1
n58 n57 n57
RMSE P.F
a b a
Organic matter 0.722 0.660 0.696 0.020 ,0.001
Crude protein 0.848 0.840 0.822 0.023 0.083
a b a
Energy 0.711 0.654 0.689 0.020 ,0.001
ab b a
Neutral-detergent fibre 0.252 0.204 0.303 0.042 ,0.001
ab a a
Acid-detergent fibre 0.125 0.090 0.165 0.050 0.019
2 a b a
Hemicellulose 0.389 0.294 0.440 0.039 ,0.001
Nutritive value(air dry basis)
Digestible protein, DP (g / kg) 133 124 131
Digestible energy, DE (MJ / kg) 11.35 10.48 10.98
DP/ DE (g / MJ) 11.7 11.8 11.9
1
a, b: See Table 2.
RMSE: Root mean square error. 2
See Table 1.
although there was a 2 unit decrease (Table 4). In The minimum transit time was identical between contrast, the replacement of starch by hemicellulose solid and liquid phase, thus only mean values for the led to a 6 unit decrease in OM digestibility. From 6 two phases were given in the Table 5. TTm for the to 10 wk old, the intake in digestible OM was similar whole tract was meanly 4.4 h from which 50% among diets (Table 3) and logically led to a similar accounted for caeco–rectal transit. TTm remained weight gain (Table 2). Crude protein digestibility or unaffected by the dietary treatments.
intake in digestible protein were not significantly Contrary to the liquid phase, the MRT of the solid affected by dietary treatments. In accordance with phase was affected by the diets, particularly in the the aims of the dietary formulation the ratio DP/ DE caeco–colic segment when it decreased (P50.008)
was similar among the diets. Whatever the diet, the by 50% for ‘‘HC1P’’ group compared to control.
hemicellulosic fraction was digested at a higher rate Although not statistically significant (high interin-compared to lignocellulose, reaching a level of 44% dividual variations) MRT for the total tract tended to for HC1P group. Compared to control diet, the be lower (P50.077) for HC1P group. TTm was
NDF digestibility was improved with the addition of not significantly different among the three groups, pectin, while it decreased with the addition of indicating therefore that the change in total and hemicellulose (mainly from wheat bran). This re- caeco–rectal MRT occurred in the mixing compart-sulted in a high quantity of fibre degraded (g / d) for ments, and mainly in the caecum and proximal HC1P group whatever the fibre fraction (Table 3), colon.
whereas fibre degradation was similar for control and
HC groups. 3.4. Caecal fermentation and microbial activity
3.3. Rate of passage Whatever the variable, no significant interactions were found between the effect of age and the effect The rate of passage in the whole tract of the of the diet (Table 6), so these two effects will be digesta solid phase was meanly 15.5 h, from which presented separately hereafter.
at least the two-thirds was attributable to the caeco–
colic segment (Table 5). The MRT of the liquid 3.4.1. Effect of age
Table 5
Mean retention time (MRT) of solid and liquid phase of digesta in the whole tract and caeco–colic segment of the growing rabbit, according to dietary treatment
Control HC HC1P Statistical analysis
1
n55 n54 n55
RMSE P.F
2 ab a b
Live weight (g) 1349 1234 1480 93 0.014
2 a a b
Feed intake (g / d) 95.2 94.5 116.7 10.7 0.013
MRT: whole tract (h)
Solid phase 18.0 15.7 13.1 2.6 0.077
Liquid phase 51.0 49.4 44.8 5.3 0.21
MRT: caecum–rectum (h)
a b b
Solid phase 14.7 9.6 7.8 2.0 0.008
Liquid phase 42.3 42.4 38.1 4.6 0.31
TTm: minimal transit time
Whole tract (h) 4.6 3.9 4.6 2.3 0.72
Caecum–rectum (h) 2.1 2.4 2.4 1.2 0.92
1
a, b: See Table 2. 2
Mean values for the period 48–57 days of age.
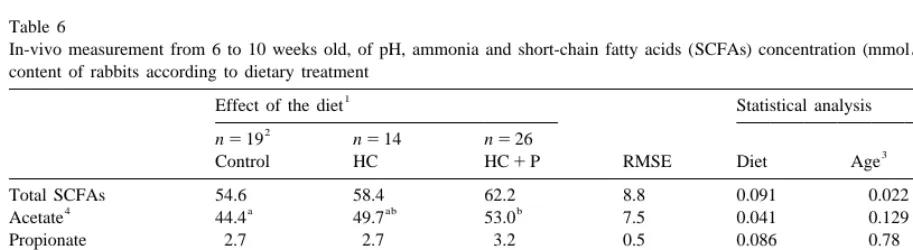
Table 6
In-vivo measurement from 6 to 10 weeks old, of pH, ammonia and short-chain fatty acids (SCFAs) concentration (mmol / l) in the caecal content of rabbits according to dietary treatment
1
Effect of the diet Statistical analysis
2
n519 n514 n526 Diet
3
Control HC HC1P RMSE Diet Age 3age
Total SCFAs 54.6 58.4 62.2 8.8 0.091 0.022 0.83
4 a ab b
Acetate 44.4 49.7 53.0 7.5 0.041 0.129 0.85
Propionate 2.7 2.7 3.2 0.5 0.086 0.78 0.95
Butyrate 6.8 5.4 5.5 1.5 0.35 ,0.001 0.75
5
Minor SCFAs 0.8 0.7 0.7 0.1 0.80 0.41 0.37
Propionate:butyrate 0.46 0.57 0.67 0.18 0.31 0.002 0.80
Ammonia 4.0 3.6 3.8 1.7 0.98 0.54 0.59
pH 6.50 6.57 6.35 0.23 0.090 0.67 0.84
1
Means from samples covering the period 45 to 70 days old. 2
a, b: See Table 2. 3
Comparison of in vivo sampling according to age (at 45, 50, 60 and 70 days old). 4
Linear effect of the age (P,0.05) 5
Isobutyrate1valerate1isovalerate.
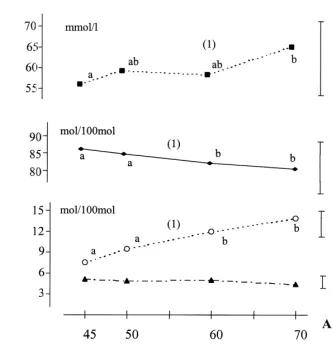
45 d to 70 d of age (Fig. 1), because of a significant 0.001) with age from 0.72 (at 45 d) to 0.36 (at 70 d). linear increase (P50.03) in acetate (from 48.4 to Ammonia or pH were not significantly affected by
52.5 mmol / l) and butyrate (from 4.3 to 8.9 mmol / l) the age (Table 6). levels. From 45 to 70 d of age, we registered a
two-fold increase of the butyrate molar proportion 3.4.2. Effect of the diet
(Fig. 1), associated with a slight decrease of the Compared to control, the total SCFA levels in acetate molar proportion (25%). Consequently the HC1P group were 8 units higher (P50.09) as the
Fig. 1. In vivo caecal SCFA pattern according to the age of the rabbits. Values are means of the three diets. a, b: Means having a common superscript did not differ significantly at the level P50.05. (1) Linear effect of the age (P,0.05).
other hand, levels of butyrate propionate or ammonia parameters (SCFA levels or molar proportion), ex-remained unaffected by dietary treatments. In return, cept the level of ammonia that was positively related the acetate molar proportion was significantly lower to crude protein digestibility (r50.59, P50.02).
(P50.034) for control compared to HC and HC1P A relatively low number of animals was involved
group (respectively, 81.6% – 85.1 – 85.1%), while in the in-vivo determination of the caecal volume the butyrate molar proportion tended to be higher and SCFA pool (Table 7), because the marker (P50.07) for control (12.1%) compared to the two dilution procedure is relatively difficult to manage
other groups (mean 9.1%). and time consuming. Therefore, only large differ-The relationship between caecal fermentation and ences among treatments could appeared as signifi-the digestion of signifi-the main nutrients (OM, CP, NDF, cant. At 70 d of age, the digesta content of the ADF and NDF-ADF) has been assessed by calculat- caecum corresponds meanly to 55g of fresh matter ing the partial correlation coefficients (r) between that represented 5% of the live weight. The caecal pH, NH or SCFA levels and the digestibility content reached 71g for HC1P that was 80% over
3
coefficients or the daily quantity of nutrients di- the value for HC group. Therefore the SCFA pool gested. Digestibility or nutrients intake were not was significantly higher for HC1P (165%)
Table 7
In-vivo measurement of the digesta content and of SCFA pool in the caecum of growing rabbits (7 wk old) according to dietary treatment
Control HC HC1P Statistical analysis
1
n54 n53 n55
RMSE P.F
Live weight (g) 1120 1013 1204 99 0.075
ab a b
Caecal digesta content (g) 59.2 39.3 71.4 12.4 0.020
SCFAs (mM / l) 59.8 65.0 68.5 10.0 0.52
2 a a b
SCFA pool (mM) 2.3 2.2 3.8 0.66 0.013
1
a, b: See Table 2. 2
SCFAs (mM / l)3liquid caecal content (ml).
4. Discussion higher than in hard faeces, Luick et al. 1992) that are ingested via caecotrophy practice. Consequently a 4.1. In-vivo approach of the digestive processes of portion of the liquid marker was recycled and
the growing rabbit contributed to increase liquid retention time, as
observed previously in caeco–colic segment by Although caecal cannulation led to a temporary Gidenne and Jehl (1996) using the same method. loss in feed intake and live weight, no interaction Leng et al. (1977) mentioned a shorter MRT for with the effect of the diets was detected for intake or liquids (mean 16.4 h) between caecum and rectum,
14
growth, as observed in previous experiments using but they used C -PEG (polyethylene glycol) that is the same methodology (Bellier and Gidenne, 1996; incompletely recovered in excreta. Rate of passage Gidenne et al., 1998b). Such a technique appeared of particles in the caeco–colic segment (MRT2
particularly convenient to assess in-vivo the hindgut TTm) was meanly 8.4 h that was in the range of digestive processes, as various measurements could previous results obtained by mathematical modelisa-be performed over a complete growth period: digesta tion (Gidenne, 1994; Gidenne and Perez, 1996). composition, caecal volume (using marker dilution As expected, particles and liquids have strictly the technique) or caeco–rectal rate of passage. Further- same retention time in the distal colon (corre-more, our study demonstrated that it was possible to sponding to the caeco–rectal TTm, Gidenne, 1994) combine SCFA analysis in digesta and digestive as digesta are already aggregated into faecal pellets volume measurements, to evaluate in-vivo the pro- (Gidenne and Jehl, 1996). Similar conclusions could duction of fermentation end-products. also be drawn for digesta retention in the small The association of a double labelling technique intestine, which was assessed by the difference (simultaneous measurements for liquid and solid between total and caeco–rectal TTm (meanly 2.1 h). phase) to that of caecal cannulation let us to provide The oro–caecal retention time was estimated, for valuable new informations about digesta rate of liquid and particles, by difference between total and passage in different segment of the rabbit tract, such caeco–rectal MRT measured consecutively on the as oro–caecal transit of liquids. In the whole tract, same animals. A differential transit seemed to occur liquid retention time reached 48 h, in agreement with between liquid and solid phase, as we found a Luick et al. (1992). The caeco–colic segments shorter rate of passage for particles (mean 4.9 h) than (caecum1proximal colon) were the main site of for liquid (mean 7.5 h). This differential transit could
liquid retention (meanly 39 h, obtained by difference be attributed to the retention in the stomach, since between caeco–rectal MRT and TTm). The retro- liquids and particles moved without differential grade transport of fluid from the proximal colon to transit in the small intestine. This higher gastric the caecum (occurring when hard faeces are pro- retention time for liquids would be related to the duced) is mainly responsible of this phenomenon retention in the fundus of soft faeces having a high
¨
(Bjornhag, 1981). It also explains the high con- concentration in liquid marker.
¨
volume was first assayed by Leng and Hornicke HC1P). However, a slight effect of the overall
(1976) using the marker dilution method and de- dietary fibre level could not be excluded on feed scribed diurnal variations according to caecotrophy intake, as NDF varied by 10% among diets. In practice. Therefore we applied this procedure after return, only a supply in hemicellulose and pectin the caecotrophy period (at 11:30) to reduce potential resulted in higher weight gain, corresponding also to variations. Our data indicated that the caecal digesta a digestibility similar to that of high starch diet. This content ranged from 4 to 6% of the live weight. confirmed the previous study of Gidenne and Jehl Similar values were reported for seven-week-old (1996) reporting the high availability for the growing rabbits in the review of Gidenne (1997), thus rabbit of a mix of hemicellulose and pectins in diets validating this procedure. with higher content of lignocellulose. The growing Very few studies have assessed in-vivo caecal rabbit compensated the replacement of starch by a fermentative activity (CFA) over complete growth higher fibre digestion (over 40% for hemicellulosic period, because of difficulties to obtain individual fraction) combined with a higher fibre intake, thus consecutive measurements over several weeks. Dur- leading to an increased quantity of fibre degraded ing the growth, the fermentation pattern evolved to a and to a high caecal fermentative activity. In con-pattern similar to the adult, corresponding mainly to trast, the hemicellulose rich diet was of low di-a linedi-ar incredi-ase in butyrdi-ate di-and di-a decredi-ase in di-acetdi-ate gestibility, even for the fibre fractions, but it was not molar proportion, as previously described (Bellier et associated with a reduction of the caecal SCFA al., 1995; Gidenne et al., 1998b). Our approach was levels. Although there was no significant individual also of interest to control that dietary effects re- relationship between the main nutrients intake and mained constant according to age. We confirmed our the caecal fermentation activity, we would point out previous results (Gidenne et al., 1998b), indicating that the mean SCFA caecal concentration increased any significant interaction between age and diet, according to the mean intake of digestible fibre from 6 to 10 wk of age. During this period, the (hemicellulose1pectins). In parallel, the reduction
caecal SCFA concentration increased linearly by of the caeco–colic retention time did not affect the 20% (reaching 65 mM / l). A similar increase was fermentation processes. Contrary to ruminants, mi-also observed by Gidenne et al. (1998b) with diets crobial activity and fibre degradation in the rabbit having a similar NDF / starch ratio (ranging from 0.9 seemed not largely affected by the caeco–colic rate to 1.2). With diets having a higher NDF / starch ratio of passage, but would be mainly controlled by the (over 1.3) Jehl and Gidenne (1996) did not mention nature of fibre. The caecal retention time is too short such a rise in caecal SCFAs with age, probably to allow bacterial degradation of cell wall that are because the SCFA concentrations (over 65 mM / l) not easily accessible (such as cellulose), but is already levelled off at six weeks of age. For instance, sufficiently long to allow a substantial degradation of earlier stabilisation of caecal SCFA level (at five the more easily digestible fibre fractions (e.g., pectins weeks old) was observed by Candau et al. (1978) or even hemicellulose).
with a diet rich in digestible fibre (such as hemicel- Studies on digestive transit recover for the rabbit lulose or pectins) and with low starch content. an additional interest in terms of preventing exces-sive retention time that favours dysfunction or 4.2. Replacement of readily available alteration of the caecal microbial activity and thus
carbohydrates by digestible fibre digestive troubles (Gidenne, 1997). Compared to less
digestible fibre fraction, pectins are reputed to slower The replacement of available carbohydrates (es- the transit in monogastric (Hillmann et al., 1983; sentially starch) by fibre fractions of high digestibili- Potkins et al., 1991). In rabbit diets, pectins are often ty (such as pectins or hemicellulose), without supplied through incorporation of beet pulp, that are changes in the supply (level and nature) of lignocel- also suspected to slower transit (predisposing to lulose (low digested), led to a similar increase development of a pathogenic flora), since Auvergne (110%) in the voluntary feed intake either for a low et al. (1987) observed that lucerne replacement with
of liquids. Inversely, our results pointed out that carbohydrates, but without changes in lignocellulose incorporation of 20% of beet pulp did not modify the content, favoured caecal microbial activity without transit of liquids either between proximal or distal slowing the digestive transit. This appeared par-part of the tract. This agrees with the study of Fraga ticularly beneficial to the rabbit growth during the et al. (1991) reporting that beet pulp, in substitution postweaning period.
to lucerne, did not alter the transit of liquids in the whole tract. Compared to a starchy diet, the rate of
passage of particles was similarly stimulated with Acknowledgements hemicellulose or with a mix of hemicellulose and
pectins. The effect was logically pronounced in the The authors thank Andre Lapanouse and Muriel´ caeco–colic segment responsible for the two-thirds Segura for their technical assistance (Station de´ of the total retention time, and it was related with Recherches Cunicoles, INRA, Toulouse, France). NDF intake. However, Gidenne and Jehl (1996) did This study was partly supported by a fellowship not mention a stimulation of the caeco–colic transit grant from Ralston Purina France (Saint Quentin En for rabbits having a high fibre intake (.30 g NDF /
Yvelines, France). d), probably because caeco–colic transit was already
at a minimal level (mean 7 h).
Animals fed pectin-rich diet presented high caecal
References
volume, as already described for rabbit pig or rat (Garcia et al., 1993; Kesting et al., 1990; Brunsgaard
Auvergne, A., Bouyssou, T., Pairet, M., Bouillier-Oudot, M., et al., 1995). This was here associated to high level Ruckebusch, Y., Candau, M., 1987. Nature de l’aliment, finesse of SCFA, therefore increasing the caecal SCFA pool. de mouture et donnees anatomo – fonctionnelles du tube´ This increment in caecal fermentative activity would digestif proximal. Reprod. Nutr. Devel. 27, 755–768.
ˆ ´
Bellier, R., 1994. Controle nutritionnel de l’activite fermentaire contribute to explain that pectin-rich feeds were as
`
caecale. These de Doctorat, Institut National Polytechnique, efficiently utilised as a starchy feed for rabbit growth
´
Ecole Nationale Superieure d’Agronomie de Toulouse, (similar feed conversion on the overall growth Toulouse.
period). This effect was particularly pronounced Bellier, R., Gidenne, T., 1996. Consequences of reduced fibre during the postweaning period when young rabbit intake on digestion, rate of passage and caecal microbial
activity in the young rabbit. Br. J. Nutr. 75, 353–363. had a higher growth rate with feeds containing a mix
Bellier, R., Gidenne, T., Vernay, M., Colin, M., 1995. In vivo of hemicellulose and pectin. Such a favourable effect
study of circadian variations of the cecal fermentation pattern of starch replacement by digestible fibre for the in the post-weaned and adult rabbits. J. Anim. Sci. 73, 128– postweaned rabbit was also reported by Jehl and 135.
¨
Gidenne (1996) and was associated with a better Bjornhag, G., 1981. The retrograde transport of fluid in the proximal colon of rabbits. Swed. J. Agric. Res. 11, 63–69. health status in relation with a stimulation of the
´
Blas, E., Fernandez Carmona, J., Cervera, C., 1988. Effect of caecal microbial activity (increase of the caecal
digestive activity and starch intake on amylase activity and biomass and SCFA level). In contrast, the supply of pancreatic juice of rabbit. In: Proceedings of the 4th Congress hemicellulose (HC diet) with a feed containing of the World Rabbit Science Association, WRSA, Budapest, pp. ingredients of almost one origin (the wheat) gave 68–73.
´
Blas, E., Cervera, C., Fernandez Carmona, J., 1994. Effect of two similar growth and caecal fermentative activity than
diets with varied starch and fibre levels on the performances of a starchy diet. The lack of diversity in the botanical
4–7 weeks old rabbits. W. Rabbit Sci. 2, 117–121.
origin of fibre might be unfavourable to the CMA, as Blumenkrantz, N., Asboe-Hansen, G., 1973. New method for already observed recently (Gidenne et al., 1998b). quantitative determination of uronic acids. Anal. Biochem. 54,
In conclusion, the dietary balance between low 484–489.
Brunsgaard, G., Eggum, B.O., Sandstrom, B., 1995. Gastrointesti-and highly digestible fibre needs to be determined
nal growth in rats as influenced by indigestible polysaccharides with precision for the growing rabbit, to favour
Proceed-´
ings of the 2nde Journees de la Recherche Cunicole, INRA, effects of pectin, cellulose and lignin on stool PH, transit-time Toulouse, pp. 1.1–1.4. and weight. Br. J. Nutr. 50, 189–195.
´ ´
Carre, B., Brillouet, J.M., 1989. Determination of water insoluble IO7, 1993. La Banque de Donnees de l’Alimentation Animale, cell-walls in feeds: interlaboratory study. J. Assoc. Off. Agric. V2.5, Association Franc¸aise de Zootechnie, Paris.
Chem. 72, 463–467. Jehl, N., Gidenne, T., 1996. Replacement of starch by digestible ´
Carre, B., 1992. Factors affecting the digestibility of non-starch fibre in the feed for the growing rabbit. 2. Consequences for carbohydrates in monogastric animals. Poultry Sci. 61, 1257– microbial activity in the caecum and on incidence of digestive
1269. disorders. Anim. Feed Sci. Technol. 61, 193–204.
Edwards, C.A., Parrett, A.M., 1996. Plant cell wall polysac- Kesting, U., Schnabel, E., Bolduan, G., 1990. Degree and effects charides, gums and hydrocolloids: nutritional aspects. In: of fermentations in the hind gut of the pig. 1. Ileal flow-rates, Eliasson, A.-C. (Ed.), Carbohydrates in Food, pp. 319–345. fermentability and colon capacity. Arch. Anim. Nutr. 10, 933–
˜ ´
Fraga, M.J., Perez De Ayala, P., Carabano, R., 1991. Effect of 942. ¨
type of fiber on the rate of passage and on the contribution of Leng, E., Hornicke, H., 1976. Diurnal variations in the rabbits soft feces to the nutrient intake of finishing rabbits. J. Anim. cecal volume in vivo with C14-PEG. Z. Vet. Med. 23, 827–835.
¨
Sci. 69, 1566–1574. Leng, E., Clauss, W., Hornicke, H., 1977. Colon passage time in Garcia, G., Galvez, J.F., De Blas, J.C., 1993. Effect of substitution rabbits in relation to the formation of caecotrophes. Z. Vet.
of sugarbeet pulp for barley in diets for finishing rabbits on Med. 24, 324–332.
growth performance and on energy and nitrogen efficiency. J. Luick, B.R., Ayers, A., Cheeke, P.R., 1992. Rate of passage of Anim. Sci. 71, 1823–1830. Black Locust leaf and alfalfa meal in the rabbit gut. J. Appl.
´
Gidenne, T., Bellier, R., 1992. Etude in vivo de l’activite Rabbit Res. 15, 914–921, Proceedings of the 5th congress of fermentaire caecale chez le lapin. Mise au point et validation the World Rabbit Science Association, 25–30 July, Corvallis, d’une nouvelle technique de canulation caecale. Reprod. Nutr. OR.
ˆ
Dev. 32, 365–376. Maıtre, I., Lebas, F., Arveux, P., Bourdillon, A., Duperray, J., ´
Gidenne, T., 1994. Effets d’une reduction de la teneur en fibres Saint Cast, Y., 1990. Taux de lignocellulose, ADF de Van-alimentaires sur le transit digestif du lapin. Comparaison et Soest, et performances de croissance du lapin de chair. In:
` ´ ´ ´
validation de modeles d’ajustement des cinetiques d’excretion Proceedings of the 5th Journees de la Recherche Cunicole, Vol. ´
fecale des marqueurs. Reprod. Nutr. Dev. 34, 295–306. 2, ITAVI, Paris, pp. 56.1–56.11.
Gidenne, T., Jehl, N., 1996. Replacement of starch by digestible Mathers, J.C., 1991. Digestion of non-starch polysaccharides by fibre in the feed for the growing rabbit. 1. Consequences for non-ruminant omnivores. Proc. Nutr. Soc. 50, 161–172. digestibility and rate of passage. Anim. Feed Sci. Technol. 61, Potkins, Z.V., Lawrence, T.L.J., Thomlinson, J.R., 1991. Effects of 183–192. structural and non-structural polysaccharides in the diet of the Gidenne, T., Perez, J.M., 1996. Dietary cellulose in the growing growing pig on gastric emptying rate of passage of digesta to rabbit. 1. Consequences on digestion and rate of passage. Ann. the terminal ileum and through the total gastrointestinal tract.
Zootech. 45, 289–298. Br. J. Nutr. 65, 391–413.
Gidenne, T., 1997. Caeco–colic digestion in the growing rabbit: Statistical Analysis System, 1988. SAS / STAT Guide For Personal impact of nutritional factors and related disturbances. Livest. Computers, Version 6.03, SAS Institute, Cary, NC.
Prod. Sci. 51, 73–88. Van Soest, P.J., Robertson, J.B., Lewis, B.A., 1991. Methods for Gidenne, T., Carabano, R., Garcia, J., De Blas, C., 1998a. Fibre dietary fiber, neutral detergent fiber, and non starch polysac-digestion in the rabbit. In: De Blas, C., Wiseman, J. (Eds.), The charides in relation to animal nutrition. J. Dairy Sci. 74, Nutrition of the Rabbit, CABI Publishing, pp. 69–88, Ch. 5. 3583–3597.
Gidenne, T., Bellier, R., Van Eys, J., 1998b. Effect of the dietary Verdow, H., Van Echteld, C.J.A., Dekkers, E.M.J., 1977. Ammonia fibre origin on the digestion and on the caecal fermentation determination based on indophenol formation with sodium pattern of the growing rabbit. Anim. Sci. 66, 509–517. salicylate. Water Res. 12, 399–402.