BEKASAM IKAN SEPAT RAWA (Trichogaster trichopterus)
YULIANTI SRI REJEKI
DEPARTEMEN TEKNOLOGI HASIL PERAIRAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR
BOGOR
YULIANTI SRI REJEKI. C34070089. Pengaruh Kondisi Kultivasi terhadap
Produksi Antibakteri dari Bakteri Asam Laktat Asal Bekasam Ikan Sepat Rawa (Trichogaster trichopterus). Dibimbing oleh DESNIAR dan KUSTIARIYAH.
Biota perairan merupakan komoditi yang mudah sekali mengalami kerusakan. Salah satu upaya pengawetan yang dilakukan adalah melakukan fermentasi bekasam. Proses fermentasi tersebut melibatkan bakteri asam laktat (BAL) yang memiliki sifat penting dalam menghasilkan metabolit antimikroba. Potensi BAL dalam menghasilkan antimikroba sangat besar sehingga perlu dilakukan penelitian karakterisasi isolat BP (8) serta uji aktivitas antibakteri yang dihasilkan dari isolat tersebut pada kondisi kultivasi yang berbeda.
Tujuan penelitian ini adalah untuk mengkarakterisasi isolat BP (8) sebagai BAL serta mempelajari pengaruh kondisi kultivasi yang berbeda terhadap aktivitas antibakteri yang dihasilkan. Penelitian ini dilakukan dalam dua tahapan, yaitu (1) karakterisasi isolat BP (8) sebagai BAL dan pendugaan genus dan (2) produksi antibakteri, yang meliputi a) penyegaran isolat; b) kultivasi isolat BP (8) menggunakan alat inkubasi yang berbeda, yaitu magnetic stirrer, shaker
waterbath dan inkubator, pada tahap ini parameter yang diamati adalah optical density (OD) dan pH; c) pemanenan, supernatan diberi tiga perlakuan, yaitu tidak
dinetralkan, dinetralkan, dan dinetralkan lalu diendapkan dengan amonium sulfat 50% saturasi; d) uji aktivitas antibakteri menggunakan metode agar well
diffusion, pada tahap ini yang diukur adalah diameter zona hambat.
Isolat BP (8) memiliki karakteristik selnya bulat, tetrad dan beberapa berpasangan, Gram-positif, tidak berspora, non-motil, katalase negatif, oksidase negatif dan mampu memfermentasi glukosa tanpa menghasilkan gas (homofermentatif). Isolat BP-8 adalah bakteri asam laktat yang tergolong homofermentatif dan diduga termasuk dalam genus Pediococcus.
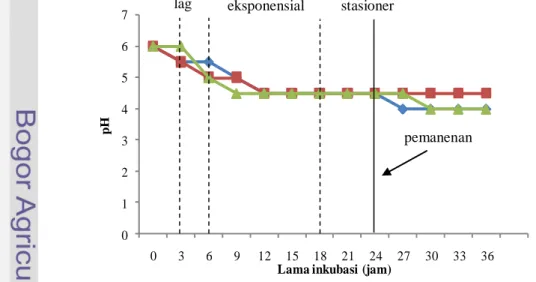
Kondisi kultivasi yang berbeda tidak memberikan pengaruh pada pertumbuhan isolat BP (8). Pemanenan yang dilakukan pada jam ke-24, dimana pada kurva pertumbuhan semuanya telah berada di pertengahan fase stasioner (OD660 4-4,5), begitu pula dengan kurva perubahan keasaman yang berada pada
titik pH yang sama, yaitu pada pH 4,5. Pemanenan dilakukan pada jam ke-24 (telah memasuki fase stasioner) karena diasumsikan asam laktat yang dihasilkan sudah stasioner dan bakteriosin sudah terbentuk.
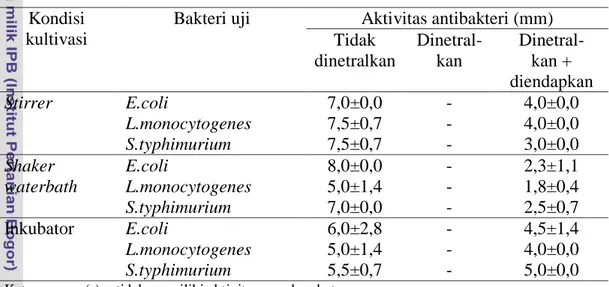
Hasil uji aktivitas terhadap bakteri uji pada kondisi asam dan pengendapan menunjukkan hasil positif, sedangkan pada kondisi netral menunjukkan hasil negatif. Pada kondisi asam, senyawa yang bekerja melawan bakteri uji adalah asam laktat, sedangkan pada pengendapan diduga terdapat bakteriosin. Pada kondisi netral, pengaruh asam organik telah hilang, sedangkan protein tidak terendapkan. Sel bakteri pada nilai OD dan nilai pH yang sama tidak selalu menghasilkan aktivitas antibakteri yang sama.
BEKASAM IKAN SEPAT RAWA (Trichogaster trichopterus)
YULIANTI SRI REJEKI C34070089
SKRIPSI
sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada Fakultas Perikanan dan Ilmu Kelautan
Institut Pertanian Bogor
DEPARTEMEN TEKNOLOGI HASIL PERAIRAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR
BOGOR
Nama : Yulianti Sri Rejeki NRP : C34070089
Departemen : Teknologi Hasil Perairan
Menyetujui: Pembimbing 1
Desniar, S.Pi, M.Si NIP. 19701224 199702 2 001
Pembimbing 2
Dr. Kustiariyah, S.Pi, M.Si NIP. 19750818 200501 2 001
Mengetahui:
Ketua Departemen Teknologi Hasil Perairan
Dr. Ir. Ruddy Suwandi, MS., MPhil. NIP: 19580511 198503 1 002
Dengan ini saya menyatakan bahwa skripsi yang berjudul ”Pengaruh Kondisi Kultivasi terhadap Produksi Antibakteri dari Bakteri Asam Laktat Asal Bekasam Ikan Sepat Rawa (Trichogaster trichopterus)” adalah karya saya sendiri dan belum diajukan dalam bentuk apapun pada perguruan tinggi manapun. Sumber informasi yang dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir skripsi ini.
Bogor, Desember 2011
Yulianti Sri Rejeki
© Hak cipta milik IPB, tahun 2011 Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhya dalam
Puji syukur penulis panjatkan kehadirat Allah SWT atas segenap limpahan karunia yang tak terhitung banyaknya. Shalawat serta salam semoga tetap tercurah kepada Rasulullah SAW.
Penyusunan skripsi yang berjudul: ”Pengaruh kondisi kultivasi terhadap produksi antibakteri dari bakteri asam laktat asal bekasam ikan sepat rawa (Trichogaster trichopterus)” merupakan salah satu syarat untuk memperoleh gelar sarjana pada Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
Penulis mengucapkan terima kasih kepada seluruh pihak yang telah membantu dalam penyelesaian skripsi ini, diantaranya kepada:
1. Desniar, S.Pi, M.Si dan Dr. Kustiariyah, S.Pi, M.Si. sebagai komisi pembimbing atas segala saran, kritik, arahan, perbaikan, dan motivasi, serta ilmu yang telah diberikan.
2. Dr. Tati Nurhayati, S.Pi, M.Si. sebagai dosen penguji yang telah memberikan masukan dan pengarahan kepada penulis.
3. Dr. Ir. Ruddy Suwandi, MS, M.Phil selaku Ketua Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
4. Dr. Ir. Agoes M. Jacoeb, Dipl. Biol selaku komisi pendidikan Departemen Teknologi Hasil Perairan.
5. Ayah dan ibu serta adik-adik tercinta (Vindy, Nabila dan Dzafira) atas segala doa restu, motivasi dan semangatnya.
6. Bu Ema, Mba Silvi, Mba Dini, Mba Lastri, Mas Ipul, Pak Wahyu, Mba Selin dan Pak Ranta, terimakasih atas bantuan dan bimbingan selama menjalankan penelitian.
7. Seluruh staf Dosen dan TU THP terimakasih atas bantuannya selama penyusunan skripsi.
8. Ikma Ratna Puspita dan Yoga Indra Purnama, teman seperjuangan dalam suka dan duka selama penelitian, teman bertukar pikiran dan berbagi canda tawa serta keluh kesah.
10. Yunko, Ratna dan Zahid atas bantuan dan kebersamaannya selama penelitian.
11. Gian, Dyhart, Rika, Tiza, Nisa, Salman dan Aristi yang selalu menyemangati, mendukung dan mendoakan.
12. Teman-teman THP 44 atas semangat, motivasi dan informasi, serta telah menjadi bagian dari hidup saya selama 3 tahun ini. Semoga tali silaturahim kita tidak putus.
13. Rekan THP 43, THP 45 dan THP 46 atas bantuan dan motivasinya.
14. Teman-teman kosan Mega 2 (Reni, Isna, Endah, Rika, Gian, Viviq, Sari, Tiah, Nibras dan Ratna) atas semangat, bantuan tenaga dan pikiran, tempat berbagi keluh kesah dan canda tawa dan tak lupa doa, dari penulis melakukan penelitian hingga penyelesaian skripsi ini.
Penulis menyadari bahwa dalam penulisan skripsi ini masih banyak kekurangannya. Penulis mengharapkan kritik dan saran yang bersifat membangun untuk perbaikan skripsi ini. Semoga skripsi ini bermanfaat bagi semua pihak yang memerlukannya.
Bogor, Desember 2011
Yulianti Sri Rejeki C34070089
Penulis dilahirkan di Jakarta pada tanggal 12 Juli 1989. Penulis merupakan anak pertama dari empat bersaudara dari pasangan Disun dan Muslikhatun. Pendidikan formal yang ditempuh penulis dimulai dari TK An-Ni’mah (tahun 1994-1995), kemudian melanjutkan pendidikan dasarnya ke SDN Pondok Labu 11 Pagi Pondok Labu (tahun 1995-2001). Penulis kemudian melanjutkan pendidikan ke SLTP Negeri 85 Jakarta Selatan (tahun 2001-2004) dan melanjutkan ke SMA Negeri 34 Jakarta Selatan (tahun 2004-2007). Pada tahun 2007, penulis diterima di Institut Pertanian Bogor melalui jalur Seleksi Penerimaan Mahasiswa Baru (SPMB), serta diterima di Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
Selama masa perkuliahan, Penulis aktif di organisasi kemahasiswaan seperti HIMASILKAN (Himpunan Mahasiswa Hasil Perikanan) periode 2008-2009 sebagai anggota divisi sosial kemasyarakatan dan periode 2009-2010 sebagai anggota divisi peduli pangan. Penulis juga mengikuti UKM Teater Air pada tahun 2008-2009. Penulis aktif sebagai Asisten mata kuliah Iktiologi pada tahun 2009-2010. Sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada Fakultas Perikanan dan Ilmu Kelautan, penulis melakukan penelitian dengan judul: Pengaruh kondisi kultivasi terhadap produksi antibakteri dari bakteri asam laktat asal bekasam ikan sepat rawa (Trichogaster trichopterus) dibimbing oleh Desniar, S.Pi, M.Si dan Dr. Kustiariyah, S.Pi, M.Si.
Halaman
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xiii
DAFTAR LAMPIRAN ... xiv
1 PENDAHULUAN ... 1 1.1 Latar Belakang ... 1 1.2 Tujuan ... 2 2 TINJAUAN PUSTAKA ... 3 2.1 Fermentasi ... 3 2.2 Bekasam ... 5
2.3 Bakteri Asam Laktat (BAL) ... 6
2.4 Faktor yang Mempengaruhi Pertumbuhan Bakteri Asam Laktat ... 9 2.5 Senyawa Antimikroba ... 10 2.5.1 Asam organik ... 10 2.5.2 Hidrogen peroksida (H2O2) ... 11 2.5.3 Karbondioksida (CO2) ... 12 2.5.4 Diasetil ... 12 2.5.5 Bakteriosin ... 12
2.6 Cara Kerja Zat Antimikrobial ... 14
2.7 Bakteri Uji ... 15
2.7.1 Escherichia coli ... 15
2.7.2 Salmonella typhimurium ... 15
2.7.3 Listeria monocytogenes ... 16
3 METODOLOGI ... 17
3.1 Waktu dan Tempat ... 17
3.2 Bahan dan Alat ... 17
3.3 Metodologi Penelitian ... 17
3.3.1 Karakterisasi isolat BP (8) ... 20
(1) Pewarnaan Gram (Fardiaz 1989) ... 20
(2) Pewarnaan spora (Fardiaz 1989) ... 20
(3) Uji motilitas (Fardiaz 1989) ... 21
(4) Uji katalase (Fardiaz 1989) ... 21
(5) Uji oksidase (Fardiaz 1989) ... 21
(6) Uji fermentasi glukosa (Hayward 1957) ... 21
(2) Kultivasi isolat BP (8) ... 23
(3) Pemanenan ... 24
(4) Uji aktivitas antibakteri ... 24
4 HASIL DAN PEMBAHASAN ... 26
4.1 Karakterisasi Isolat ... 26 4.1.1 Pewarnaan Gram ... 26 4.1.2 Pewarnaan spora ... 27 4.1.3 Uji motilitas ... 28 4.1.4 Uji katalase ... 29 4.1.5 Uji oksidase ... 30
4.1.6 Uji fermentasi glukosa ... 30
4.1.6 Total kadar asam laktat ... 31
4.2 Produksi Antibakteri ... 33
4.2.1 Kultivasi isolat BP (8) ... 34
(1) Pertumbuhan bakteri ... 34
(2) Perubahan pH ... 36
4.2.2 Aktivitas antibakteri ... 37
5 KESIMPULAN DAN SARAN ... 44
5.1 Kesimpulan ... 44
5.2 Saran ... 44
DAFTAR PUSTAKA ... 45
Nomor Halaman 1 Karakterisasi dan pendugaan genus isolat BP (8) ... 26 2 Aktivitas penghambatan antibakteri pada kondisi tidak
dinetralkan, dinetralkan dan pengendapan ... 38 3 Aktivitas penghambatan antibakteri dengan menggunakan asam
Nomor Halaman 1 Fermentasi homolaktat ... 7 2 Fermentasi heterolaktat ... 8 3 Escherichia coli ... 15 4 Salmonella typhimurium ... 16 5 Listeria monocytogenes ... 16
6 Diagram alir tahap karakterisasi ... 18
7 Diagram alir tahap produksi antibakteri... 19
8 Bentuk sel dan hasil pewarnaan Gram isolat BP (8) ... 27
9 Hasil pewarnaan spora isolat BP (8) ... 28
10 Hasil uji motilitas isolat BP (8) ... 29
11 Hasil uji fermentasi glukosa ... 31
12 Densitas optik isolat BP (8) dengan alat inkubasi magnetic stirrer, shaker waterbath dan inkubator ... 34
13 Perubahan pH selama pertumbuhan isolat BP (8) dengan alat inkubasi magnetic stirrer, shaker waterbath dan inkubator ... 36
14 Aktivitas penghambatan pada bakteri E. coli, S. typhimurium dan L. monocytogenes oleh supernatan yang tidak dinetralkan pada kondisi inkubasi dengan magnetic stirrer, shaker waterbath dan inkubator ... 39
15 Aktivitas penghambatan pada bakteri E. coli, S. typhimurium dan L. monocytogenes oleh supernatan yang dinetralkan dan diendapkan pada kondisi inkubasi dengan magnetic stirrer, shaker waterbath dan inkubator ... 41
Nomor Halaman 1 Dokumentasi alat-alat ... 50 2 Pengujian sifat pembedaan pada bakteri anaerobik fakultatif dari
genus bakteri Gram-positif kokusmenurun Bergey’s Manual (Holt et al. 1994) ... 51 3 Pembuatan buffer fosfat pH 7 ... 53 4 Nilai OD660 kultur dan pH selama inkubasi 36 jam dengan alat
inkubasi yang berbeda ... 53 5 Perhitungan total kadar asam laktat ... 54 6 Hasil uji aktivitas antibakteri pada perlakuan kode A, kode N
dan kode E pada kondisi kultivasi dengan inkubator, shaker
waterbath dan magnetic stirrer ... 54
7 Hasil uji aktivitas antibakteri dengan kontrol positif dan kontrol negatif ... 56
1. PENDAHULUAN
1.1 Latar Belakang
Hasil perairan merupakan komoditi yang mudah sekali mengalami kerusakan atau biasa dikenal dengan istilah perishable food. Berbagai upaya dilakukan untuk mencegah hal tersebut, yaitu dengan dilakukannya pengolahan agar bahan baku dapat lebih awet. Pengawetan yang dilakukan antara lain pengeringan, pengasapan, penggaraman, penggorengan, pembekuan, fermentasi dan lain-lain.
Fermentasi merupakan suatu cara pengolahan melalui proses memanfaatkan penguraian senyawa dari bahan-bahan protein kompleks. Protein kompleks tersebut terdapat dalam tubuh ikan yang diubah menjadi senyawa-senyawa lebih sederhana dengan bantuan enzim yang berasal dari tubuh ikan atau mikroorganisme serta berlangsung dalam keadaan yang terkontrol (Adawyah 2007).
Makanan hasil fermentasi laktat tanpa disadari telah lama menjadi bagian di dalam menu makanan sehari-hari di beberapa kawasan di Indonesia. Bekasam atau bekacem, yang berasal dari Sumatera Selatan, merupakan ikan awetan dengan cara fermentasi bakteri asam laktat. Bekasam bukan saja merupakan makanan tradisional yang digemari, tetapi juga menjadi contoh pengawetan secara biologis yang luas penggunaannya. Makanan sejenis juga ditemukan di Filipina,
yaitu burongisda, di Jepang, yaitu naresushi atau funasushi
(The National Academy 1992) dan di Thailand, yaitu pla-ra, pla-paeng-daeng,
pla-chao, pla-chom, pla-som, dan lain-lain. Pada penelitian sebelumnya, Tanasupawat et al. (1998) melaporkan bahwa dari produk pla-ra telah berhasil diisolasi strain Lactobacillus, Leuconostoc, Pediococcus, Enterococcus dan Staphylococcus.
Asam yang dihasilkan merupakan hasil dari perombakan karbohidrat (nasi) menjadi asam laktat oleh bakteri asam laktat (BAL). Sifat yang terpenting dari bakteri asam laktat adalah kemampuannya untuk merombak senyawa kompleks menjadi senyawa yang lebih sederhana sehingga dihasilkan asam laktat (Fardiaz 1992). Produksi asam laktat di dunia mencapai 80.000 ton dan sekitar 90% diantaranya dihasilkan oleh bakteri asam laktat melalui fermentasi
dan sisanya dihasilkan secara sintetis (Hidayat et al. 2006). Produksi asam oleh bakteri asam laktat berjalan secara cepat, hal ini dapat menyebabkan pertumbuhan mikroorganisme lain yang tidak diinginkan dapat terhambat (Fardiaz 1992).
Bakteri asam laktat juga mempunyai aktivitas terapeutik pada pangan fermentasi, antara lain sebagai serum kolesterol, anti tumor dan antibakteri (Biotol 1991). Salah satu karakteristik yang paling penting dari BAL adalah kemampuannya untuk memproduksi beragam metabolit dengan sifat antimikroba, antara lain asam laktat, asam asetat, etanol, diasetil, CO2, H2O2, reuterin dan
bakteriosin (Roller 2003). Sifat antibakteri serta senyawa aktif dalam BAL (bakteriosin) penting di dalam industri pangan, karena kemampuannya dapat menghambat kontaminasi mikroorganisme pembusuk dan mikroorganisme patogen, sehingga pangan menjadi lebih awet.
Beberapa penelitian yang telah dilakukan menunjukkan bahwa BAL asal pangan fermentasi mampu menghasilkan senyawa antibakteri yang menghambat pertumbuhan beberapa bakteri patogen pada pangan. Salah satu yang mempengaruhi pertumbuhan BAL adalah suhu dan kondisi lingkungan pertumbuhan. Bakteri asam laktat memiliki potensi dalam menghasilkan bioaktif yang berfungsi sebagai antibakteri, oleh karena itu perlu dilakukan karakterisasi isolat menggunakan Bergey’s Manual (Holt et al. 1994) untuk menduga genus bakteri yang akan menghasilkan aktivitas antibakteri, selanjutnya isolat dilakukan uji aktivitas antibakteri dengan kondisi kultivasi yang berbeda untuk mengetahui pengaruh kondisi kultivasi dalam memproduksi senyawa antibakteri.
1.2 Tujuan
Penelitian ini bertujuan untuk mengkarakterisasi isolat BP (8) sebagai BAL serta mempelajari pengaruh kondisi kultivasi yang berbeda terhadap aktivitas antibakteri yang dihasilkannya.
2. TINJAUAN PUSTAKA
2.1 Fermentasi
Fermentasi merupakan suatu cara pengolahan melalui proses memanfaatkan penguraian senyawa dari bahan-bahan protein kompleks. Protein kompleks tersebut terdapat dalam tubuh ikan yang diubah menjadi senyawa-senyawa lebih sederhana dengan bantuan enzim yang berasal dari tubuh ikan atau mikroorganisme serta berlangsung dalam keadaan yang terkontrol (Adawyah 2007). Fermentasi secara teknik dapat didefinisikan sebagai suatu proses oksidasi anaerobik atau partial anaerobik karbohidrat yang menghasilkan alkohol serta beberapa asam, namun banyak proses fermentasi yang menggunakan substrat protein dan lemak (Muchtadi dan Ayustaningwarno 2010).
Fermentasi terbagi menjadi dua, yaitu fermentasi spontan dan tidak spontan (membutuhkan starter). Fermentasi spontan adalah fermentasi yang biasa dilakukan menggunakan media penyeleksi, seperti garam, asam organik, asam mineral, nasi atau pati. Media penyeleksi tersebut akan menyeleksi bakteri patogen dan menjadi media yang baik bagi tumbuh kembang bakteri selektif yang membantu jalannya fermentasi. Fermentasi tidak spontan adalah fermentasi yang dilakukan dengan penambahan kultur organisme bersama media penyeleksi sehingga proses fermentasi dapat berlangsung lebih cepat (Rahayu et al. 1992).
Hasil fermentasi diperoleh sebagai akibat metabolisme mikroba-mikroba pada suatu bahan pangan dalam keadaan anaerob. Mikroba yang melakukan fermentasi membutuhkan energi yang umumnya diperoleh dari glukosa. Dalam keadaan aerob, mikroba mengubah glukosa menjadi air, CO2 dan energi (ATP).
Beberapa mikroba hanya dapat melangsungkan metabolisme dalam keadaan anaerob dan hasilnya adalah substrat yang setengah terurai. Hasil penguraiannya adalah air, CO2, energi dan sejumlah asam organik lainnya, seperti asam laktat,
asam asetat, etanol serta bahan-bahan organik yang mudah menguap. Perkembangan mikroba-mikroba dalam keadaan anaerob biasanya dicirikan sebagai proses fermentasi (Muchtadi dan Ayustaningwarno 2010).
Fermentasi glukosa pada prinsipnya terdiri dari dua tahap, yaitu (1) pemecahan rantai karbon dari glukosa dan pelepasan paling sedikit dua pasang
atom hidrogen, menghasilkan senyawa karbon lainnya yang lebih teroksidasi daripada glukosa, (2) senyawa yang teroksidasi tersebut direduksi kembali oleh atom hidrogen yang dilepaskan dalam tahap pertama, membentuk senyawa-senyawa lain sebagai hasil fermentasi (Fardiaz 1989).
Tahap pertama fermentasi glukosa selalu menghasilkan asam piruvat. Jasad renik melakukan pemecahan glukosa menjadi asam piruvat melalui empat jalur, yaitu:
1. Jalur Embden-Meyerhof-Parnas (EMP) atau glikolisis, ditemukan pada fungi dan kebanyakan bakteri, serta hewan dan manusia.
2. Jalur Entner-Doudoroff (ED), hanya ditemukan pada beberapa bakteri. 3. Jalur Heksosamonofosfat (HMF), ditemukan pada berbagai organisme. 4. Jalur Fosfoketolase (FK), hanya ditemukan pada bakteri yang tergolong
laktobasili heterofermentatif .
Jalur EMP terdiri dari beberapa tahap, masing-masing dikatalis oleh enzim tertentu. Jalur tersebut ditandai dengan pembentukan fruktosa difosfat, kemudian pemecahan menjadi dua molekul gliseraldehida fosfat. Reaksi ini dikatalis oleh enzim aldolase. Kemudian terjadi reaksi dehidrogenasi gliseraldehida fosfat yang merupakan reaksi oksidasi yang menghasilkan energi dalam bentuk ATP. Reaksi ini dikatalis oleh enzim gliseraldehida fosfat dehidrogenase. Atom hidrogen yang terlepas akan ditangkap oleh NAD membentuk NADH2. Proses fermentasi dapat
berlangsung terus jika NADH2 dapat dioksidasi kembali pada tahap kedua
fermentasi sehingga melepaskan atom hidrogen kembali. Jadi NAD berfungsi sebagai pembawa hidrogen dalam proses fermentasi (Fardiaz 1989).
Jalur FK merupakan percabangan dari jalur HMF, karena bakteri ini tidak mempunyai enzim aldolase yang dapat memecah fruktosa 1,6-difosfat menjadi 2 triose-fosfat, dan tidak mempunyai enzim transaldolase dan transketolase yang penting dalam jalur HMF (Fardiaz 1989).
Pada tahap kedua fermentasi, asam piruvat akan diubah menjadi produk-produk akhir yang spesifik untuk berbagai proses fermentasi, menggunakan atom hidrogen yang diproduksi pada tahap pertama fermentasi (Fardiaz 1989).
2.2 Bekasam
Bekasam merupakan produk olahan ikan dengan cara fermentasi yang rasanya asam. Olahan tersebut banyak dikenal di daerah Jawa Tengah, Sumatera Selatan dan Kalimantan Selatan. Ikan yang dapat digunakan sebagai bekasam merupakan jenis ikan air tawar. Pengolahan bekasam di daerah Kalimantan Selatan umumnya dikenal dengan nama samu. Bahan baku berupa ikan gabus, betok, sepat siam dan sepat rawa dengan penambahan garam sekitar 15-20%, dan ditambahkan samu atau beras ginseng sebanyak 15%, kemudian difermentasi sekitar satu minggu sampai menghasilkan aroma dan rasa yang khas bekasam (Adawyah 2007).
Produk-produk fermentasi ikan yang menggunakan garam dan bahan yang berfungsi sebagai sumber karbohidrat banyak terdapat di negara-negara Asia Tenggara. Burongisda adalah produk sejenis bekasam yang berasal dari Filipina.
Burongisda ini dibuat dari campuran ikan air tawar, nasi, garam dan angkak
(beras merah sebagai pewarna). Proses fermentasi pada pembuatan burongisda berlangsung sampai daging ikan menjadi lembut serta rasa dan bau asam mulai berkembang. Bakteri asam laktat yang dominan pada burongisda adalah
Leuconostoc mesenteroides, Pediococcus cereviceae dan Lactobacilus plantarum (Rahayu et al. 1992).
Pla-ra merupakan produk sejenis bekasam yang berasal dari Thailand
dengan waktu fermentasi selama 6 bulan. Proses pembuatan pla-ra biasanya menggunakan udang sebagai tambahan bahan baku yang berfungsi sebagai pembentuk aroma. Produk pla-ra mengandung protein sebesar 11,61-23,32%, pH sekitar 4,10-6,90 dan asam laktat sebesar 0,17-1,94%. Mikroba yang ditemukan pada produk ini adalah Pediococcus halophilus, Staphylococcus epidermidis,
Micrococcus sp. dan Bacillus sp. (Rahayu et al. 1992).
Pengolahan bekasam dilakukan dengan menambahkan sumber karbohidrat dalam kondisi anaerobik. Karbohidrat didekomposisi melalui proses fermentasi menjadi gula-gula sederhana kemudian dikonversi menjadi alkohol dan asam yang berperan sebagai pengawet dan memberikan rasa dan bau spesifik pada bekasam (Irianto 2008). Selama proses fermentasi, protein ikan akan terhidrolisis menjadi asam-asam amino dan peptida, kemudian asam-asam amino akan terurai lebih
lanjut menjadi komponen-komponen lain yang berperan dalam membentuk citarasa produk (Adawyah 2007).
Kualitas produk-produk ikan fermentasi dijaga untuk waktu yang relatif lama karena adanya penghambatan pertumbuhan bakteri lain oleh hidrogen peroksida dan antibiotik yang dihasilkan oleh Lactobacilli (Irianto 2008).
2.3 Bakteri Asam Laktat (BAL)
Bakteri asam laktat adalah kelompok bakteri yang melakukan penguraian karbohidrat (glukosa) menjadi asam laktat yang akan menurunkan pH serta menimbulkan rasa asam (Muchtadi dan Ayutaningwarno 2010). Anggota bakteri asam laktat merupakan bakteri Gram-positif, batang atau kokus yang tunggal, berpasangan atau rantai tidak berspora, terkadang membentuk segi empat, katalase negatif, toleran terhadap asam dan anaerob fakultatif (Defigueredo dan Splittstoesser 1976; Mozzi et al. 2010). Bakteri yang termasuk kelompok bakteri asam laktat adalah Aerococcus,
Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Pediococcus (Ringo dan Gatesoupe 1998).
Bakteri asam laktat dibagi menjadi dua grup berdasarkan hasil akhir metabolisme glukosa. Bakteri asam laktat yang hanya menghasilkan asam laktat pada fermentasi glukosa termasuk dalam golongan homofermentatif. Bakteri asam laktat yang menghasilkan asam laktat, CO2 dan etanol dari heksosa termasuk
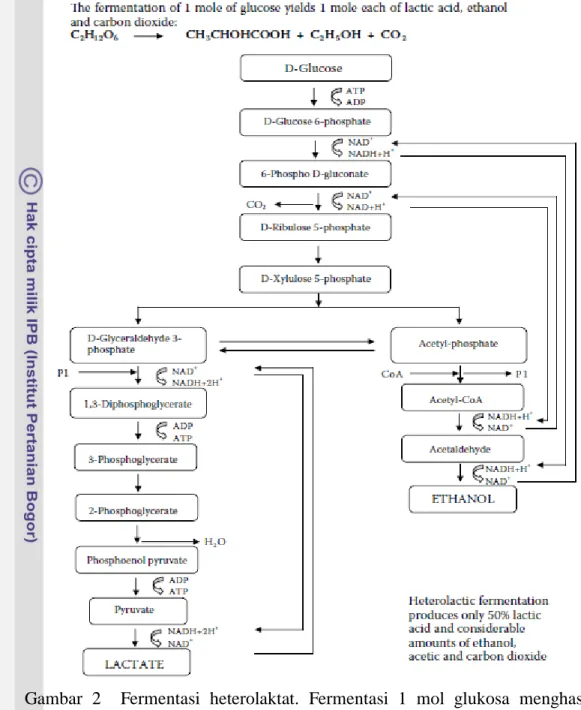
dalam golongan heterofermentatif (Jay et al. 2005). Golongan heterofermentatif memfermentasi glukosa melalui jalur fosfoketolase, sedangkan golongan homofermentatif melalui jalur EMP (Embden-Meyerhof-Parnas Pathway) (Hidayat et al. 2006). Bakteri asam laktat yang tergolong homofermentatif dapat mengubah 95% dari glukosa atau heksosa lainnya menjadi asam laktat. Karbondioksida dan asam-asam volatil lainnya juga dihasilkan, tapi jumlahnya sangat kecil (Adawyah 2007). Perbedaan fermentasi homolaktat dan heterolaktat dapat dilihat pada Gambar 1 dan Gambar 2.
Gambar 1 Fermentasi homolaktat. Fermentasi 1 mol glukosa menghasilkan 2 mol asam laktat melalui jalur Embden-Meyerhof Parnas (Theron dan Lues 2011).
Gambar 2 Fermentasi heterolaktat. Fermentasi 1 mol glukosa menghasilkan masing-masing 1 mol asam laktat, etanol dan karbondioksida melalui jalur fosfoketolase (Theron dan Lues 2011).
Salah satu karakteristik yang paling penting dari BAL adalah kemampuannya untuk memproduksi beragam metabolit dengan sifat antimikroba. Bakteri asam laktat memproduksi asam laktat, asam asetat, etanol, diasetil, CO2
(sebagai asam karbonat), H2O2, reuterin, derivat asam laktat (hidroksi asam laktat)
dan peptida kecil tergantung pada tipe strain dan nutrisi, fisik, dan lingkungan pertumbuhan. Antimikroba ini dapat menghambat atau membunuh mikroorganisme target seperti kapang, ragi, bakteri vegetatif, spora bakteri, dan
bahkan virus. Spektrum antimikroba bervariasi tergantung pada spesifikasi metabolit (Roller 2003).
Bakteri asam laktat termasuk mikroorganisme yang aman jika ditambahkan dalam pangan karena sifatnya tidak toksik dan tidak menghasilkan toksin, maka disebut food grade microorganism atau dikenal sebagai mikroorganisme yang Generally Recognized As Safe (GRAS), yaitu mikroorganisme yang tidak beresiko terhadap kesehatan, bahkan beberapa jenis bakteri tersebut berguna bagi kesehatan. Bakteri asam laktat bermanfaat untuk peningkatan kualitas higiene dan keamanan pangan melalui penghambatan secara alami terhadap flora berbahaya yang bersifat patogen (Daeschel 1983 diacu dalam Kusmiati dan Malik 2002).
Efektivitas BAL dalam menghambat bakteri pembusuk dipengaruhi oleh kepadatan dan strainnya serta komposisi media (Jeppensen dan Huss 1993 diacu dalam Rostini 2007). Produki substansi penghambat dari BAL dipengaruhi oleh media pertumbuhan, pH, dan temperatur lingkungan (Ahn dan Stiles 1990 diacu dalam Rostini 2007).
2.4 Faktor yang Mempengaruhi Pertumbuhan Bakteri Asam Laktat
Proses fermentasi sangat dipengaruhi oleh adanya pertumbuhan bakteri asam laktat. Oleh karena itu, perlu dibuat kondisi yang ideal bagi pertumbuhan bakteri tersebut. Faktor-faktor lingkungan yang mempengaruhi pertumbuhan bakteri asam laktat antara lain adalah suhu, nilai pH, kadar garam, dan karbohidrat.
Suhu akan berpengaruh terhadap pertumbuhan sel dan juga pembentukan produk oleh mikroba. Hal ini berhubungan dengan jenis mikroba yang dominan selama fermentasi (Fardiaz 1988). Berdasarkan suhu (minimum, optimum dan maksimum) untuk pertumbuhannya mikroba dibedakan atas tiga grup, yakni psikrofilik, mesofilik dan termofilik. Nilai pH medium merupakan salah satu parameter penting yang dapat mempengaruhi pertumbuhan mikroba. Bakteri pada umumnya tumbuh dengan baik pada pH sekitar 6,5-7,5. Bakteri yang berperan dalam fermentasi silase adalah bakteri asam laktat. Asam laktat yang dihasilkan akan menurunkan nilai pH pada lingkungan pertumbuhannya.
Garam sering digunakan dalam proses fermentasi ikan. Jumlah garam yang ditambahkan dalam pembuatan bekasam berkisar antara 15-20% dari berat ikan segar (Murtini 1992). Kebutuhan garam untuk pertumbuhan optimum mikroorganisme bervariasi, tergantung dari sifat dinding sel dan tekanan osmotik internalnya (Fardiaz 1992). Karbohidrat merupakan sumber energi bagi bakteri asam laktat. Penambahan karbohidrat akan membuat lingkungan yang baik bagi pertumbuhan bakteri tersebut. Selama fermentasi, karbohidrat akan diuraikan menjadi senyawa-senyawa yang sederhana seperti asam laktat, asam asetat, asam propionat dan etil alkohol. Senyawa-senyawa ini yang meyebabkan rasa asam pada produk dan dapat berfungsi sebagai pengawet (Rahayu et al. 1992).
2.5 Senyawa Antimikroba
Bakteri asam laktat dapat berfungsi sebagai pengawet makanan karena mampu memproduksi asam organik, menurunkan pH lingkungannya dan mengekskresikan senyawa yang mampu menghambat mikroorganisme pathogen, seperti H2O2, diasetil, CO2, asetaldehida, d-isomer asam asam amino dan
bakteriosin (Hardy 1975 diacu dalam Kusmiati dan Malik 2002). Sifat menghambat dapat disebabkan oleh lepasnya komponen intraselular antimikroba, seperti asam organik, bakteriosin dan hidrogen peroksida, dari sel nonmetabolit bakteri asam laktat (Roller 2003).
2.5.1 Asam organik
Aksi antimikroba dari asam organik terutama berdasarkan pada kemampuannya dalam mereduksi pH pangan dalam fase air. Ketika nilai pH <4, asam menghambat pertumbuhan bakteri. Mekanisme antimikroba berdasarkan pada hambatan enzim, fungsi membran, transpor nutrien, dan keseluruhan aktivitas metabolik. Asam organik dalam pangan dapat berperan sebagai pengawet, sedangkan garam atau esternya efektif sebagai antimikroba pada pH yang mendekati netral (Roller 2003).
Asam laktat mampu melemahkan permeabilitas bakteri Gram-negatif dengan merusak membran luar bakteri Gram-negatif. Asam laktat merupakan molekul yang larut dalam air sehingga mampu menembus ke dalam periplasma bakteri Gram-negatif melalui protein porin pada membran luarnya. Pelindung dari permeabilitas membran luar berupa lapisan lipopolisakarida yang terletak pada
permukaan membran dirusak oleh asam laktat sehingga substrat antimikroba yang lain, yaitu diasetil, bakteriosin, hidrogen peroksida dan laktoperidase sistem dapat berpenetrasi ke dalam membran sitoplasma (Alokomi et al. 2000). Pertumbuhan bakteri Gram-positif dan negatif berkurang, diindikasikan oleh meningkatnya aksi bakteriosin. Asam laktat memiliki efek bakterisidal pada pH dibawah 5, khususnya pada bakteri Gram-negatif (Ray 2004).
Asam asetat digunakan pada jenis makanan berbeda-beda untuk menghambat pertumbuhan dan mengurangi kelangsungan hidup bakteri Gram-positif dan negatif, ragi dan kapang. Asam asetat umumnya bakteriostatik pada 0,2% tapi bakterisidal diatas 0,3% dan lebih efektif menghambat bakteri Gram-negatif (Ray 2004).
2.5.2 Hidrogen peroksida (H2O2)
Beberapa bakteri asam laktat memproduksi H2O2 dibawah kondisi
pertumbuhan aerobik dan, karena kurangnya katalase selular, pseudokatalase, atau peroksidase, mereka melepas H2O2 ke lingkungan untuk melindungi diri mereka
sendiri dari aksi antibakteri (Ray 2004). Hidrogen peroksida diproduksi oleh bakteri asam laktat sebagai hasil dari aksi flavoprotein oksidase atau nikotinamida adenine dinukleotida (NADH) peroksidase. Efek antimikroba dari H2O2 adalah
hasil dari oksidasi grup sulfihidril yang menyebabkan denaturasi sejumlah enzim, dan dari peroksidase membran lipid meningkatkan permeabilitas membran (Kong dan Davison 1980 diacu dalam Ammor et al. 2006). Beberapa strain dapat memproduksi, pada kondisi pertumbuhan yang cocok, H2O2 yang cukup
menyebabkan bakteriostatik (6-8 µg/ml) tapi jarang bersifat bakterisidal (30-40 µg/ml) (Ray 2004). Hidrogen peroksida dapat juga sebagai prekursor untuk produksi bakterisidal radikal bebas seperti superoksida (O2-) dan radikal hidroksil
(OH-) yang dapat merusak DNA (Byczkowski dan Gessner 1988 diacu dalam Ammor et al. 2006).
Hidrogen peroksidamerupakan agen pengoksidasi kuat dan dapat menjadi antimikroba terhadap bakteri, jamur dan virus (juga bakteriofage). Pada kondisi anaerob, sangat sedikit H2O2 yang dapat dihasilkan dari strain ini. Aksi antibakteri
ini dihasilkan dari sifat pengoksidasi kuat dan kemampuannya untuk merusak komponen selular, khususnya membran. Karena sifat oksidasinya, maka dapat
menyebabkan efek yang tidak diinginkan dalam mutu pangan, seperti diskolorasi pada daging yang diproses, sehingga penggunaannya terbatas dalam pengawetan pangan (Ray 2004).
2.5.3 Karbondioksida (CO2)
Karbondioksida diproduksi terutama oleh BAL heterofermentatif. Karbon
dioksida memainkan peranan penting dalam membuat lingkungan anaerobik yang menghambat enzimatik dekarboksilase, dan akumulasi CO2 membran lipid
bilayer dapat menyebabkan disfungsi permeabilitas (Eklund 1984 diacu dalam Ammor et al. 2006). Karbon dioksida secara efektif dapat menghambat banyak mikroorganisme perusak makanan, terutama bakteri psikrotropik Gram-negatif (Farber 1991 diacu dalam Ammor et al. 2006).
2.5.4 Diasetil
Diasetil diproduksi oleh strain dalam semua genera dari BAL oleh fermentasi sitrat (Ray 2004). Diasetil menghambat pertumbuhan bakteri Gram-negatif yang bereaksi dengan pemanfaatan arginin (Jay 1986 diacu dalam Ammor
et al. 2006). Antibakteri ini efektif terhadap bakteri Gram-positif dan negatif.
Bakteri Gram-negatif sebagian sensitif pada pH ≤5. Diasetil efektif pada konsentrasi 0,1-0,25%. Studi terkini menunjukkan bahwa diasetil lebih bakterisidal bila dikombinasi dengan panas. Aksi antibakteri kemungkinan diproduksi dengan deaktivasi beberapa enzim penting. Grup karbonil (-CO-CO-) bereaksi dengan arginin pada enzim dan memodifikasi situs katalitiknya (Ray 2004).
2.5.5 Bakteriosin
Kata bakteriosin biasanya digunakan pada peptida bioaktif yang diproduksi oleh banyak strain bakteri dari grup Gram-negatif dan positif. Bakteriosin yang diproduksi oleh banyak strain bakteri asam laktat dan beberapa bakteri asam propionat merupakan bagian khusus dalam mikrobiologi pangan karena efek bakterisidalnya biasanya untuk membedakan kerusakan akibat bakteri Gram-positif dan patogen, dan pada kondisi stress bakteri Gram-negatif yang penting dalam pangan (Ray 2004).
Kondisi optimum produksi bakteriosin dipengaruhi oleh fase pertumbuhan, pH media, suhu inkubasi, jenis sumber karbon, jenis sumber
nitrogen, dan konsentrasi NaCl (Kim dan Ahn 2000 diacu dalam Usmiati dan Marwati 2007). Peptida bakteriosin disintesis di ribosom, kationik, amfipatik, mempunyai struktur α-helical atau β-sheet, atau keduanya, dan dapat memiliki thioether, jembatan disulfida atau grup thiol bebas. Keberadaan struktur α-helical amfipatik dengan sisi polar dan nonpolar yang berlawanan sepanjang poros yang panjang memungkinkan bakteriosin untuk berinteraksi, baik dengan fase air dan lemak, ketika mengikat permukaan membran sel bakteri yang sensitif, penting untuk destabilisasi fungsional dan kematian sel (Ray 2004).
Mekanisme aktivitas bakterisidal bakteriosin adalah sebagai berikut: (1) molekul bakteriosin kontak langsung dengan membran sel, (2) proses kontak ini mampu mengganggu potensial membran berupa destabilitas membran sitoplasma sehingga sel menjadi tidak kuat, dan (3) ketidakstabilan membran mampu memberikan dampak pembentukan lubang atau pori pada membran sel melalui proses gangguan terhadap PMF (Proton Motive Force) (Gonzalez et al. 1996 diacu dalam Usmiati 2007).
Bakteriosin dapat diproduksi oleh Lactococcus, Lactobacillus dan
Pediococcus yang berasal dari berbagai bahan makanan. Beberapa kelebihan
bakteriosin sehingga potensial digunakan sebagai biopreservatif, yaitu (i) bukan bahan toksik dan mudah mengalami degradasi oleh enzim proteolitik karena merupakan senyawa protein; (ii) tidak membahayakan mikroflora usus karena mudah dicerna oleh enzim saluran pencernaan; (iii) dapat mengurangi penggunaan bahan kimia sebagai pengawet pangan; (iv) penggunaannya fleksibel; dan (v) stabil terhadap pH dan suhu yang cukup luas sehingga tahan terhadap proses pengolahan yang melibatkan asam dan basa, serta kondisi panas dan dingin (Cleveland et al. 2001 diacu dalam Usmiati dan Marwati 2007).
Beberapa studi menunjukkan bahwa strain yang berbeda dalam spesies atau bahkan spesies yang berbeda dapat memproduksi bakteriosin yang sama. Bakteriosin yang diproduksi oleh spesies/strain BAL yang berbeda memiliki karakteristik fisika dan kimia yang sangat berbeda. Bakteriosin dikelompokkan berdasarkan keberadaan asam amino yang tidak biasa, lanthionin dan β-lanthionin (Roller 2003).
2.6 Cara Kerja Zat Antimikrobial
Secara umum, kemungkinan situs serangan suatu zat antimikrobial dapat diduga dengan meninjau struktur serta komposisi sel mikroba. Sel hidup yang normal memiliki sejumlah besar enzim yang melangsungkan proses-proses metabolik dan juga protein lainnya, asam nukleat serta senyawa-senyawa lain. Kerusakan pada salah satu dari situs ini dapat mengawali terjadinya perubahan-perubahan yang menuju pada matinya sel tersebut. Perubahan tersebut antara lain (Pelczar dan Chan 2005):
(a) Kerusakan pada dinding sel
Struktur dinding sel dapat dirusak dengan cara menghambat pembentukannya atau mengubahnya setelah selesai terbentuk.
(b) Perubahan permeabilitas sel
Membran sitoplasma mempertahankan bahan-bahan tertentu di dalam sel serta mengatur aliran keluar masuknya bahan-bahan lain. Membran memelihara integritas komponen-komponen selular. Kerusakan pada membran ini akan mengakibatkan terhambatnya pertumbuhan sel atau matinya sel.
(c) Perubahan molekul protein dan asam nukleat
Hidupnya suatu sel bergantung pada terpeliharanya molekul-molekul protein dan asam nukleat dalam keadaan alamiahnya. Suatu kondisi atau substansi yang mengubah keadaan ini, yaitu mendenaturasikan protein dan asam-asam nukleat dapat merusak sel tanpa dapat diperbaiki kembali. Suhu tinggi dan konsentrasi pekat beberapa zat kimia dapat mengakibatkan koagulasi (denaturasi) ireversibel komponen-komponen selular yang vital ini.
(d) Penghambatan kerja enzim
Setiap enzim yang ada di dalam sel merupakan sasaran potensial bagi bekerjanya suatu penghambat. Penghambatan ini dapat mengakibatkan terganggunya metabolisme atau matinya sel.
(e) Penghambatan sintesis asam nukleat dan protein
DNA, RNA dan protein memegang peranan amat penting di dalam proses kehidupan normal sel. Gangguan apapun yang terjadi pada pembentukan atau pada fungsi zat-zat tersebut dapat mengakibatkan kerusakan total pada sel.
2.7 Bakteri Uji
Bakteri uji merupakan bakteri yang bersifat patogen dan digunakan sebagai tolak ukur untuk mengetahui besarnya tingkat aktivitas antibakteri. Bakteri yang digunakan pada penelitian ini, yaitu Escherichia coli, Salmonella
typhimurium, dan Listeria monocytogenes.
2.7.1 Escherichia coli
Escherichia coli merupakan bakteri Gram-negatif, motil, tidak berspora,
berbentuk batang dan anaerobik fakultatif. Escherichia coli menyebabkan penyakit diare. Escherichia coli juga penyebab utama infeksi urin dan infeksi nosokomial termasuk septisemia dan meningitis (Holt et al. 1994). Escherichia
coli secara umum terdapat pada usus hewan ruminansia. Sumber makanan yang
berasosiasi dengan E.coli adalah daging mentah, susu, air (Wallace et al. 2011).
Gambar 3 Escherichia coli (Bio Research Laboratories, Inc. 2010). 2.7.2 Salmonella typhimurium
Salmonella typhimurium merupakan bakteri Gram-negatif, tidak berspora,
fakultatif anaerobik, motil, tidak membentuk spora. Salmonella menghasilkan
gas ketika ditumbuhkan pada media yang mengandung glukosa (Pelczar dan Chan 2005). Salmonella mesofilik, suhu pertumbuhan optimum
35-37 oC, tapi umumnya memiliki range pertumbuhan 5-46 oC. Salmonella mati pada suhu dan waktu pasteurisasi, sensitif pada pH rendah (Ray 2004).
Salmonella dapat menyebabkan gastroenteritis, diare, nausea dan muntah
(Pelczar dan Chan 2005). Salmonella secara alami terdapat pada usus hewan, banyak ditemukan pada daging mentah, daging unggas dan telur (Wallace et al. 2011). Daging ayam dan olahannya dilaporkan sebagai media penyebaran penyakit salmonellosis (Usmiati 2007).
Gambar 2 Salmonella typhimurium (Bio Research Laboratories, Inc. 2010). 2.7.3 Listeria monocytogenes
Listeria monocytogenes merupakan bakteri Gram-positif, psikrotropik,
fakultatif anaerobik, tidak berspora, motil, batang pendek. Pada kultur segar, selnya terkadang membentuk rantai pendek. Listeria monocytogenes tumbuh pada kisaran 1-44 oC, dengan suhu pertumbuhan optimum 35-37 oC. Bakteri ini memfermentasi glukosa tanpa menghasilkan gas. Sel ini cukup resisten terhadap pembekuan, pengeringan, kadar garam tinggi, dan pH ≥5. Listeria monocytogenes sensitif terhadap suhu pasteurisasi (Ray 2004). Bakteri ini secara alami terdapat pada tanah, usus hewan dan area pengolahan yang lembab (Wallace et al. 2011).
3. METODOLOGI
3.1. Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Februari-Agustus 2011, bertempat di Laboratorium Mikrobiologi Hasil Perairan dan Laboratorium Bioteknologi Hasil Perairan, Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, serta Laboratorium Terpadu, Fakultas Kedokteran Hewan, Institut Pertanian Bogor.
3.2. Bahan dan Alat
Isolat BP (8) diisolasi dari produk bekasam ikan sepat rawa (lama fermentasi 4 hari) asal kecamatan Indralaya, Kabupaten Ogan Ilir, Sumatera Selatan. Bakteri uji yang digunakan adalah Escherichia coli, Salmonella
typhimurium dan Listeria monocytogenes. Media yang digunakan adalah nutrient agar/broth (Oxoid CM 0003), media de Man Rogosa Sharpe agar (MRSA)
(Oxoid CM 0361), Lactobacillus MRSB HIMEDIA, Mueller Hinton Agar (MHA), Sulfid Indol Motility (SIM).
Bahan yang digunakan adalah malachite green (Merck), minyak immersi (Merck), H2O2 3% (Merck), fenolftalein (Merck), p-aminodimetilanilin oksalat
1% (Merck), ammonium sulfat (Merck), NaH2PO4 (Merck), Na2HPO4 (Merck),
NaCl (Merck), NaOH 0,1 N (Merck), NaOH 1 N (Merck), kristal violet (Merck), lugol (Merck), safranin (Merck), kertas saring Whattman 42, kertas pH dan millipore.
Alat yang digunakan untuk penelitian ini adalah Spectronic 20, Thermolyne type 42000 incubator, Yamato shaker waterbath, Yamato SM52
autoclave, Yamato type MH61 magnetic stirrer, Hitachi Himac CR 21G high-speed refrigerated centrifuge dan Olympus Optical mikroskop (Lampiran 1).
3.3. Metode Penelitian
Penelitian ini terdiri dari dua tahapan, yaitu (1) karakterisasi isolat BP (8) sebagai BAL dan pendugaan genus dan (2) produksi antibakteri, yang meliputi a) penyegaran isolat; b) kultivasi isolat BP (8) dengan menggunakan alat inkubasi yang berbeda, yaitu magnetic stirrer, shaker waterbath dan inkubator,
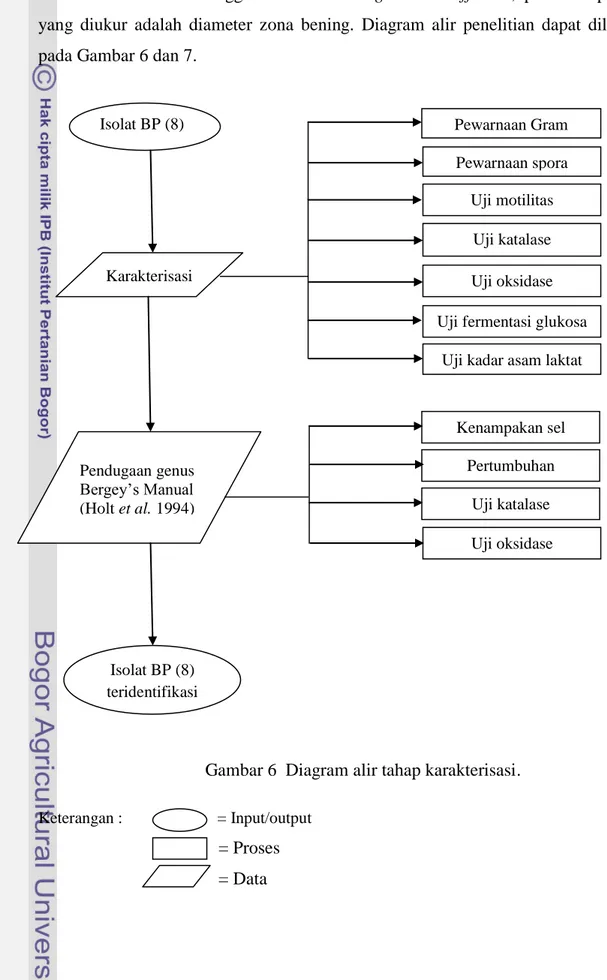
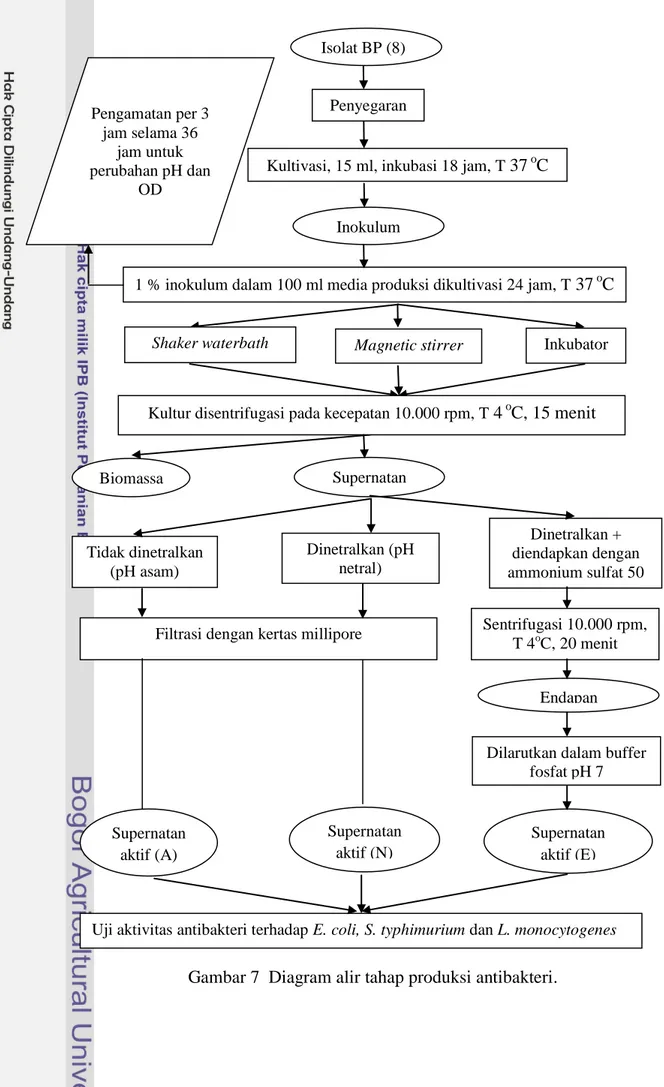
parameter yang diukur adalah optical density (OD) dan pH; c) pemanenan, supernatan diberi tiga perlakuan yaitu tidak dinetralkan, dinetralkan dan dinetralkan lalu diendapkan dengan ammonium sulfat 50% saturasi; d) uji aktivitas antibakteri menggunakan metode agar well diffusion, pada tahap ini yang diukur adalah diameter zona bening. Diagram alir penelitian dapat dilihat pada Gambar 6 dan 7.
Gambar 6 Diagram alir tahap karakterisasi.
Keterangan : = Input/output
= Proses = Data
Isolat BP (8) Pewarnaan Gram
Karakterisasi Pendugaan genus Bergey’s Manual (Holt et al. 1994) Isolat BP (8) teridentifikasi
Uji kadar asam laktat Pewarnaan spora
Uji motilitas Uji katalase Uji oksidase Uji fermentasi glukosa
Kenampakan sel Pertumbuhan
Uji katalase Uji oksidase
Gambar 7 Diagram alir tahap produksi antibakteri.
Uji aktivitas antibakteri terhadap E. coli, S. typhimurium dan L. monocytogenes Dinetralkan (pH netral) Dinetralkan + diendapkan dengan ammonium sulfat 50 % Sentrifugasi 10.000 rpm, T 4oC, 20 menit Endapan
Dilarutkan dalam buffer fosfat pH 7 Filtrasi dengan kertas millipore
Penyegaran
Kultivasi, 15 ml, inkubasi 18 jam, T 37 oC
1 % inokulum dalam 100 ml media produksi dikultivasi 24 jam, T 37 oC
Shaker waterbath Magnetic stirrer Inkubator
Kultur disentrifugasi pada kecepatan 10.000 rpm, T 4 oC, 15 menit
Biomassa Supernatan Tidak dinetralkan (pH asam) Supernatan aktif (E) Supernatan aktif (N) Supernatan aktif (A) Pengamatan per 3 jam selama 36 jam untuk perubahan pH dan OD Isolat BP (8) Inokulum
3.3.1 Karakterisasi isolat
Isolat BP (8) diperoleh dari hasil isolasi bakteri asal bekasam ikan sepat rawa yang diperoleh dari kecamatan Indralaya, Kabupaten Ogan Ilir, Sumatera Selatan. Tujuan karakterisasi adalah untuk verifikasi isolat BP (8) sebagai kelompok BAL dan menentukan pendugaan genus. Bakteri asam laktat merupakan bakteri Gram positif, batang atau kokus tunggal, berpasangan atau rantai tidak berspora, terkadang membentuk segiempat, katalase negatif, toleran terhadap asam dan anaerob fakultatif (Defigueredo dan Splittstoesser 1976; Mozzi et al. 2010). Karakterisasi isolat BP (8) untuk verifikasi meliputi pewarnaan Gram, pewarnaan spora, uji motilitas, uji katalase, oksidase, kadar asam laktat dan fermentasi glukosa. Pendugaan genus dilakukan mengikuti tahap karakterisasi menurut Bergey’s Manual (Holt et al. 1994) (Lampiran 2).
(1) Pewarnaan Gram (Fardiaz 1989)
Lapisan tipis dibuat secara aseptis dari suspensi bakteri di atas gelas objek dan dilakukan fiksasi pada udara terbuka. Lapisan tipis ini ditetesi zat warna kristal violet dan dibiarkan selama 1 menit, kemudian dibilas dengan air kran dengan cara memegang gelas objek pada posisi miring. Sisa air yang tertinggal pada gelas objek dibuang dan ditetesi dengan lugol serta dibiarkan selama 1 menit. Gelas objek dibilas kembali, kemudian dihilangkan warnanya dengan menggunakan alkohol 96% dan dibiarkan selama 10-20 detik, lalu dibilas. Olesan pada gelas objek kemudian diwarnai dengan safranin dan dibiarkan selama 10-20 detik. Objek gelas selanjutnya dibilas dengan air dan dikeringkan dengan kertas serap (tissue). Preparat ini diamati dibawah mikroskop menggunakan lensa objektif yang telah diolesi minyak immersi. Bakteri-Gram positif akan ditunjukkan dengan warna ungu, sedangkan bakteri Gram-negatif akan ditandai dengan warna merah atau merah muda.
(2) Pewarnaan spora (Fardiaz 1989)
Lapisan tipis dibuat secara aseptis dari suspensi bakteri di atas gelas objek dan difiksasi. Lapisan tipis ini ditetesi pewarna malachite green dan dibiarkan selama 20 menit tanpa pemanasan. Preparat kemudian dibilas dengan air dengan cara memegang gelas objek pada posisi miring dan dikeringkan dengan kertas serap (tissue). Olesan pada gelas objek kemudian ditambahkan beberapa tetes zat
warna safranin dan dibiarkan selama 30 detik dan dicuci dengan air mengalir serta dikeringkan. Preparat ini diamati di bawah mikroskop menggunakan lensa objektif yang telah diolesi minyak immersi. Endospora yang masih terdapat dalam sel vegetatif maupun spora bebas akan berwarna hijau biru, sedangkan sel vegetatif akan berwarna merah sampai merah muda.
(3) Uji motilitas (Fardiaz 1989)
Pengujian motilitas bakteri dilakukan dengan cara sebagai berikut: secara aseptis dengan menggunakan jarum ose yang lurus bagian ujungnya, isolat bakteri ditusukkan ke dalam media SIM. Inkubasi dilakukan pada suhu 37 °C selama 1hari. Pertumbuhan bakteri yang menyebar menunjukkan bahwa bakteri tersebut bersifat motil dan bila pertumbuhan bakteri tidak menyebar, hanya berupa garis saja, maka bakteri tersebut bersifat non motil.
(4) Uji katalase (Fardiaz 1989)
Isolat bakteri sebanyak 1 lup diambil secara aseptis dan dipindahkan pada gelas objek. Preparat tersebut ditetesi dengan larutan 3% H2O2. Adanya enzim
katalase ditandai dengan terbentuknya gelembung-gelembung seperti busa sabun. (5) Uji oksidase (Fardiaz 1989)
Sebanyak 1 gram p-aminodimetilanilin oksalat dilarutkan dalam 100 cc akuades kemudian dipanaskan. Larutan tersebut diteteskan ke kertas Whatman 42 hingga warna kertas berubah menjadi ungu. Isolat bakteri yang akan diujikan dioleskan ke kertas Whatman 42. Kertas yang tetap berwarna ungu menunjukkan hasil positif, sedangkan bila berubah menjadi merah maka negatif.
(6) Uji fermentasi glukosa (Hayward 1957)
Pendeteksian produksi gas dari isolat BP (8) dilakukan dengan metode fermentasi glukosa dalam tabung Durham. Isolat BP (8) diinokulasikan secara aseptik ke dalam media MRSB + glukosa 10% dan diinkubasi selama 24 jam pada suhu 37 oC. Hasil uji heterofermentatif ditunjukkan dengan adanya gas yang terbentuk dalam tabung Durham.
(7) Total kadar asam laktat (Moore et al. 2011)
Pengukuran kadar asam laktat dilakukan dengan metode titrasi menggunakan larutan NaOH (N=0,1091). Supernatan sebanyak 1 ml dilarutkan dengan pewarna fenolftalein, kemudian dititrasi oleh NaOH hingga warna larutan
supernatan berubah kemerahan. Persentase (%) asam laktat dihitung menggunakan rumus:
% Asam laktat = V NaOH x N NaOH x 90 x FP x 100% Bobot Sampel
Keterangan:
V NaOH = Volume NaOH yang terpakai
N NaOH = Normalitas NaOH yang terukur (0,1091) FP = Faktor Pengencer (1)
Bobot sampel = bobot supernatan (mg)
90 = BM Asam laktat
(8) Pendugaan genus (Holt et al. 1994)
Isolat BP (8) yang telah diverifikasi kemudian dilakukan pengujian ke tahap selanjutnya hingga tingkat genus dengan menggunakan buku identifikasi
Bergey’s Manual (Holt et al. 1994). Karakteristik yang diamati untuk
membedakan sifat dari genus bakteri asam laktat menurut buku Bergey’s Manual adalah sebagai berikut:
1. Kenampakan sel: sel diamati apakah berpasangan, 4 sel membentuk kelompok (tetrad), membentuk rantai (panjang, pendek, sangat panjang), atau membentuk kluster.
2. Pertumbuhan, bakteri ditumbuhkan pada:
(a) media produksi yang diinkubasi pada suhu 45 °C (b) media produksi dengan pH 9,6
(c) media produksi dengan penambahan 6,5 % NaCl 3. Reaksi katalase (uji katalase)
4. Keberadaan sitokrom (uji oksidase) 3.3.2 Produksi antibakteri
Isolat dari bakteri yang telah teridentifikasi dikultur untuk memproduksi supernatan aktif sebagai antibakteri. Tahapan ini meliputi penyegaran isolat, kultivasi BAL, pemanenan dan uji aktivitas antibakteri.
(1) Penyegaran isolat
Media yang digunakan dalam tahap ini adalah MRSA dan MRSB. Media MRSA yang sudah ditimbang dilarutkan dalam akuades kemudian diaduk hingga homogen di atas kompor listrik pada suhu 100 °C, begitu pula dengan MRSB.
Media yang telah siap kemudian disterilisasi menggunakan autoklaf pada suhu 121 °C dan tekanan 1 atm selama 15 menit.
Penyegaran bakteri dan persiapan inokulum dilakukan untuk mendapatkan biakan bakteri yang segar dan siap pakai. Proses penyegaran dan persiapan inokulum adalah sebagai berikut: isolat bakteri asam laktat dalam gliserol disegarkan pada media MRSA miring, disimpan pada kaleng bekas biskuit dengan dimensi kaleng 15x15x25 (cm) dengan kondisi anaerob kemudian diinkubasi pada suhu 37 oC selama 48 jam. Bakteri di MRSA miring diinokulasikan 1 lup dalam medium inokulum MRSB dengan volume kerja 15 ml. Setelah itu diinkubasi dengan shaker waterbath selama 18 jam pada suhu 37 oC kemudian diukur optical
density dengan λ=660 nm (OD660 0,6-0,8).
(2) Kultivasi isolat BP (8)
Kultivasi sel bakteri merupakan proses peningkatan konsentrasi beberapa atau semua komponen suatu populasi dan biasanya secara mutlak ditentukan oleh macam pengukuran yang digunakan untuk memantau proses tersebut. Pengukuran sering digunakan untuk mencerminkan pertambahan jumlah atau massa sel (Hadiutomo 1988).
Isolat BP (8) dikultivasi dengan menggunakan tiga alat yang memiliki kondisi yang berbeda, yaitu menggunakan magnetic stirrer, shaker waterbath dan inkubator. Magnetic stirrer pada kondisi suhu ruang dan agitasi 150 rpm, shaker
waterbath dengan suhu 37 °C dengan agitasi 150 rpm dan inkubator dengan
kondisi suhu 37 °C dan berada dalam kaleng yang hampa udara serta tanpa agitasi.
Sebanyak 1% inokulum dimasukkan dalam medium produksi (MRSB) dengan volume kerja 100 ml menggunakan botol Schott®. Media yang telah diinokulasi diinkubasi dengan tiga perlakuan, yaitu inkubasi dengan magnetic
stirrer pada kondisi suhu ruang dan agitasi 150 rpm, inkubator dengan kondisi
suhu 37 °C dan berada dalam kaleng yang hampa udara serta tanpa agitasi dan
shaker waterbath dengan suhu 37 °C dengan agitasi 150 rpm. Pengamatan
dilakukan setiap 3 jam selama 36 jam. Parameter yang diamati adalah perubahan pH dan pertumbuhan dengan mengukur OD pada λ =660 nm.
(3) Pemanenan
Pemanenan dilakukan terhadap kultur yang telah berumur 24 jam. Kultur disentrifugasi dengan kecepatan 10.000 rpm dengan suhu 4 oC selama 15 menit (Nurhasanah 2004 diacu dalam Nurmalis 2008), kemudian supernatan dipisahkan dari endapan (biomassa sel).
Supernatan diberi tiga perlakuan, yaitu (1) tidak dinetralkan/pH asam (kode A), (2) dinetralkan dengan NaOH 1N/pH netral (kode N), (3) dinetralkan lalu diendapkan dengan (NH4)2SO4 (ammonium sulfat) pada konsentrasi 50%
saturasi (kode E). Ammonium sulfat 50% kejenuhan adalah 341 g ammonium sulfat dilarutkan dalam 1 liter.
Supernatan yang tidak dinetralkan dan yang dinetralkan difiltrasi menggunakan millipore dengan ukuran pori 0,2 µm sehingga diperoleh supernatan bebas sel (kode A dan kode N).
Ammonium sulfat dimasukkan ke dalam supernatan netral secara perlahan-lahan sambil diaduk dengan magnetic srirrer sampai larut dan homogen, diendapkan dan disimpan dalam refrigerator selama 24 jam, kemudian disentrifugasi dengan kecepatan 10.000 rpm selama 20 menit pada suhu 4 oC (Purwanti 2003). Endapan yang diperoleh dilarutkan dalam buffer fosfat 0,1 M pH 7 (kode E). Pembuatan larutan buffer fosfat pH 7 dapat dilihat pada Lampiran 3.
(4) Uji aktivitas antibakteri
Uji aktivitas antibakteri dilakukan untuk mengetahui kemampuan isolat BAL dalam menghasilkan zat antibakteri. Uji aktivitas antibakteri dilakukan terhadap bakteri uji seperti Escherichia coli, Salmonella typhimurium dan Listeria
monocytogenes menggunakan metode difusi sumur agar (well diffusion agar)
(Hilmi dan Gokalp 2000). Bakteri uji disegarkan pada media NA miring secara aseptik, dan diinkubasi pada suhu 37 oC selama 24 jam. Sebanyak 1 lup bakteri uji diinokulasikan dalam 10 ml NB dan diinkubasi dalam shaker waterbath selama 18 jam setelah itu diukur OD pada λ=600 sebesar 0,6-0,8.
Sebanyak 20 µl bakteri uji berumur 18 jam (OD600 sebesar 0,6-0,8)
dicampurkan dalam 20 ml media MHA (Mueller Hinton Agar) pada suhu media 50 oC. Media yang telah diinokulasikan tersebut kemudian dituangkan ke dalam
cawan petri. Media agar yang telah padat dibuat sumur-sumur dengan pipet Pasteur steril dengan diameter 6 mm. Supernatan aktif (asam, netral dan yang diendapkan) diambil sebanyak 50 µl dan dimasukkan ke dalam sumur pada media agar, lalu diinkubasi pada suhu 37 oC selama 1 hari dan diamati aktivitasnya. Zona bening di sekeliling sumur menunjukkan adanya daya hambat. Areal penghambatan diukur berdasarkan diameter areal bening yang terbentuk di sekitar sumur (Hilmi dan Gokalp 2000).
4. HASIL DAN PEMBAHASAN
4.1 Karakterisasi Isolat
Karakterisasi isolat BP (8) untuk verifikasi meliputi pewarnaan Gram, pewarnaan spora, uji motilitas, uji katalase, uji oksidase, uji fermentasi glukosa, penentuan kadar asam laktat. Pendugaan genus dilakukan dengan menumbuhkan bakteri pada media produksi dengan pH 9,6, media produksi yang diinkubasi pada suhu 45 °C dan media produksi yang ditambahkan NaCl 6,5% (Holt et al. 1994). Hasil karakterisasi dapat dilihat pada Tabel 1.
Tabel 1 Karakterisasi dan pendugaan genus isolat BP (8) Karakteristik Hasil Verifikasi
Gram Positif
Bentuk tetrad, beberapa berpasangan
Spora -
Motilitas -
Katalase -
Oksidase -
Fermentasi glukosa Tidak menghasilkan gas (homofermentatif) Total kadar asam laktat
- Magnetic stirrer 4,76 % - Skaker waterbath 4,57 %
- Inkubator 4,96 %
Kemampuan untuk tumbuh pada:
pH 9,6 -
Suhu 45 °C +
NaCl 6,5 % +
Hasil verifikasi menunjukkan bahwa isolat BP (8) termasuk dalam anggota kelompok BAL dan pada pendugaan genus, isolat ini termasuk dalam genus
Pediococcus sp.
Keterangan : (+) = memiliki aktivitas/tumbuh, (-) = tidak memiliki aktivitas/tidak tumbuh
4.1.1 Pewarnaan Gram
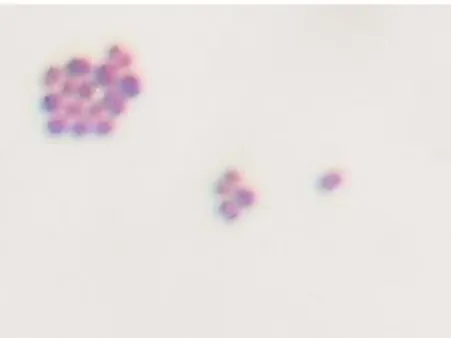
Pengamatan secara mikroskopik terhadap bentuk dan struktur sel merupakan tahap yang paling penting dalam karakterisasi bakteri. Isolat BP (8) memiliki bentuk bulat, selnya tetrad dan beberapa berpasangan. Pewarnaan Gram menunjukkan reaksi Gram positif pada isolat ini. Hasil pewarnaan Gram dapat dilihat pada Gambar 8.
Gambar 8 Bentuk sel dan hasil pewarnaan Gram isolat BP (8)
Ciri-ciri bakteri Gram-positif ditandai dengan terbentuknya warna ungu pada sel bakteri. Hal tersebut disebabkan karena bakteri ini mempunyai kandungan lipid yang lebih rendah, sehingga dinding sel bakteri akan lebih mudah terdehidrasi akibat perlakuan dengan alkohol. Dinding sel yang terdehidrasi menyebabkan ukuran pori-pori sel menjadi kecil dan daya permeabilitasnya berkurang sehingga zat warna ungu kristal yang merupakan zat warna utama tidak dapat keluar dari sel dan sel akan tetap berwarna ungu. Bakteri Gram-negatif terlihat berwarna merah karena bakteri ini kehilangan pewarna kristal violet pada waktu pembilasan dengan alkohol namun mampu menyerap pewarna tandingan yaitu safranin. Bakteri Gram-negatif mengandung lipid, lemak atau substansi seperti lemak dalam persentase lebih tinggi daripada yang dikandung bakteri Gram-positif. Dinding sel bakteri Gram-negatif juga lebih tipis daripada dinding sel bakteri Gram-positif (Pelczar dan Chan 2005).
4.1.2 Pewarnaan spora
Pewarnaan spora menunjukkan bahwa isolat BP (8) tidak membentuk spora. Spora bersifat tahan terhadap kondisi lingkungan ekstrim dan adanya bahan kimia beracun. Spora dibentuk oleh spesies bakteri yang termasuk dalam genera
Clostridium dan Bacillus untuk mengatasi lingkungan yang tidak menguntungkan
bagi bakteri. Spora terbentuk dalam sel sehingga seringkali disebut sebagai endospora dan dalam sel bakteri hanya terdapat satu spora. Sel yang semakin tua menyebabkan sel vegetatif akan pecah sehingga endospora akan terlepas dari sel dan membentuk spora bebas. Zat warna yang paling sering digunakan untuk mewarnai spora adalah malachite green yang akan tetap diikat oleh spora bakteri setelah pencucian dengan air dan sebagai counterstain digunakan safranin. Endospora yang masih terdapat di dalam sel vegetatif maupun spora bebas akan
berwarna hijau-biru, sedangkan sel vegetatif akan berwarna merah sampai merah muda. Spora juga lebih tahan terhadap pewarnaan, akan tetapi sulit untuk melepaskan zat warna yang telah terserap ke dalamnya, sehingga tidak dapat mengikat zat warna lain yang diberikan berikutnya (counterstain). Prinsip pewarnaan ini digunakan untuk membedakan spora dari sel vegetatif (Fardiaz 1987). Hasil pewarnaan spora dapat dilihat pada Gambar 9.
Gambar 9 Hasil pewarnaan spora isolat BP (8). 4.1.3 Uji motilitas
Pengujian motilitas menunjukkan bahwa isolat BP (8) bersifat non motil. Hal ini dapat dilihat dari pertumbuhannya yang tidak menyebar pada media SIM. Oleh karena isolat tersebut bersifat non motil, maka dapat dinyatakan bahwa bakteri tersebut tidak mempunyai flagella sebagai organ untuk bergerak. Flagella merupakan salah satu struktur utama di luar sel bakteri yang menyebabkan terjadinya pergerakan (motilitas) pada sel bakteri. Flagella terbuat dari sub unit - sub unit protein yang disebut dengan flagelin. Sebagian besar spesies bakteri yang termasuk ke dalam kelompok Bacillus dan Spirilum mempunyai flagella sebagai alat geraknya, tetapi jarang ditemukan pada kelompok bakteri yang berbentuk coccus (Pelczar dan Chan 2005). Hasil pengujian motilitas bakteri dapat dilihat pada Gambar 10.
Gambar 10 Hasil uji motilitas isolat BP (8). 4.1.4 Uji katalase
Uji katalase digunakan untuk mengetahui adanya enzim katalase pada isolat bakteri. Katalase adalah enzim yang dapat mengkatalisasi penguraian hidrogen peroksida (H2O2) menjadi air dan O2. Hidrogen peroksida bersifat
toksik terhadap sel karena bahan ini dapat menginaktivasikan enzim dalam sel. Uji ini penting dilakukan untuk mengetahui sifat bakteri terhadap kebutuhan akan oksigen (Lay 1994 diacu dalam Candra 2006).
Mikroba dapat dibedakan atas tiga grup berdasarkan kebutuhannya akan oksigen, yaitu mikroba yang bersifat aerobik, anaerobik dan anaerobik fakultatif. Setiap bakteri mempunyai suatu enzim yang tergolong flavoprotein yang dapat bereaksi dengan oksigen membentuk senyawa-senyawa beracun yaitu H2O2 dan
suatu radikal bebas yaitu O2*, dengan reaksi sebagai berikut: Flavoprotein oksigen H2O2 + O2*
Bakteri yang bersifat anaerobik fakultatif juga mempunyai enzim superoksida dismutase, tetapi tidak mempunyai enzim katalase, melainkan mempunyai enzim peroksidase. Enzim tersebut dapat mengkatalis reaksi antara H2O2 dengan senyawa organik, menghasilkan senyawa yang tidak beracun
(Fardiaz 1989). Reaksinya adalah sebagai berikut: H2O2 + Senyawa organik
peroksidase
Senyawa organik teroksidasi + H2O
Bakteri yang bersifat aerobik mempunyai enzim superoksida dismutase yang dapat memecah radikal bebas dan enzim katalase yang dapat memecah H2O2
sehingga menghasilkan senyawa-senyawa akhir yang tidak beracun. Reaksinya dapat dituliskan sebagai berikut:
2 O2*+ 2 H+ superoksida dismutase H2O2 + O2 2 H2O2 katalase 2 H2O2 + O2
Berbeda halnya dengan bakteri anaerobik obligat, bakteri ini tidak mempunyai enzim superoksida dismutase maupun katalase. Oleh karena itu oksigen merupakan racun bagi bakteri tersebut karena terbentuknya H2O2 dan O2*
(Fardiaz 1989).
Penentuan adanya enzim katalase diuji menggunakan larutan 3 % H2O2
pada koloni terpisah. Isolat bakteri yang bersifat katalase positif akan terlihat pembentukan gelembung udara di sekitar koloni. Isolat BP (8) tidak menghasilkan gelembung (tidak memiliki enzim katalase), sehingga dapat disimpulkan bahwa bakteri tersebut bersifat anaerobik fakultatif.
4.1.5 Uji oksidase
Uji oksidase berfungsi untuk menentukan adanya sitokrom oksidase yang ditemukan pada mikroorganisme tertentu. Sitokrom oksidase merupakan enzim yang berperan sebagai katalisator dalam transfer atom hidrogen dari sitokrom yang terakhir ke molekul oksigen. Sitokrom merupakan senyawa organik yang terdapat dalam sel hidup dan berperan dalam transfer atom hidrogen dari substrat ke molekul oksigen dengan katalis enzim sehingga dapat terbentuk air (Winarno dan Fardiaz 1984).
Berdasarkan hasil uji, dapat diketahui bahwa isolat BP (8) tidak menghasilkan enzim sitokrom oksidase yang mengkatalis transfer hidrogen dari sitokrom ke molekul oksigen. Hal ini menunjukkan bahwa bakteri tersebut tidak melakukan metabolisme energi melalui proses respirasi, melainkan melalui proses fermentasi. Hal tersebut terjadi karena dalam produk bekasam terdapat senyawa organik yang mampu digunakan oleh bakteri tersebut sebagai donor dan aseptor elektron untuk menghasilkan energi (Candra 2006). Berdasarkan uji katalase dan oksidase, dapat disimpulkan bahwa bakteri yang diisolasi dari produk bekasam mempunyai sifat anaerobik fakultatif.
4.1.6 Uji fermentasi glukosa
Uji fermentasi glukosa bertujuan untuk mengetahui apakah bakteri tersebut tergolong dalam bakteri homofermentatif atau heterofermentatif. Tabung
durham yang terdapat gelembung udara menunjukkan bahwa bakteri tersebut menghasilkan CO2, sehingga tergolong dalam BAL heterofermentatif. Tabung
durham yang tidak terdapat gelembung udara menunjukkan bahwa bakteri tersebut termasuk dalam golongan homofermentatif. Bakteri asam laktat yang hanya menghasilkan asam laktat pada fermentasi glukosa termasuk dalam golongan homofermentatif. Bakteri asam laktat yang menghasilkan asam laktat, CO2 dan etanol dari heksosa termasuk dalam golongan heterofermentatif
(Jay et al. 2005). Isolat BP (8) tidak menghasilkan gelembung udara pada tabung Durham, maka isolat BP (8) tergolong dalam BAL homofermentatif. Hasil uji fermentasi glukosa dapat dilihat pada Gambar 11.
Gambar 11 Hasil uji fermentasi glukosa isolat BP (8). 4.1.7 Total kadar asam laktat
Pengujian kadar asam laktat berfungsi untuk mengetahui total asam laktat yang terbentuk selama pertumbuhan isolat BP (8). Asam laktat yang dihasilkan ini akan menurunkan pH. Total kadar asam laktat yang dihasilkan oleh isolat BP (8) pada perlakuan kultivasi magnetic stirrer sebesar 4,76%, shaker waterbath sebesar 4,57% dan inkubator sebesar 4,96% (Lampiran 4).
Asam laktat pada produk fermentasi terbentuk akibat adanya pemecahan glukosa oleh bakteri asam laktat. Bakteri ini dapat dibedakan menjadi dua kelompok, yaitu bakteri homofermentatif dan heterofermentatif. Bakteri asam laktat yang menghasilkan asam laktat, CO2 dan etanol dari heksosa
termasuk dalam golongan heterofermentatif (Jay et al. 2005). Golongan heterofermentatif memfermentasi glukosa melalui jalur fosfoketolase, sedangkan golongan homofermentatif melalui jalur EMP (Embden-Meyerhof-Parnas) (Hidayat et al. 2006).