Productivity, microclimate and water use in
Grevillea robusta
-based
agroforestry systems on hillslopes in semi-arid Kenya
C.K. Ong
a, C.R. Black
b,∗, J.S. Wallace
c, A.A.H. Khan
a, J.E. Lott
b,1, N.A. Jackson
c,
S.B. Howard
b,2, D.M. Smith
caInternational Centre for Research in Agroforestry, PO Box 30677, Gigiri, Nairobi, Kenya
bSchool of Biological Sciences, University of Nottingham, Sutton Bonington Campus, Loughborough LE12 5RD, UK cInstitute of Hydrology, Crowmarsh Gifford, Wallingford OX10 8BB, UK
Received 12 March 1999; received in revised form 26 November 1999; accepted 25 January 2000
Abstract
This paper describes a multi-disciplinary project to examine the changing interactions between trees and crops as the trees in semi-arid agroforestry systems establish and mature; the project is one of the most detailed and highly instrumented long-term studies of tree and crop growth, system performance, resource capture, hydrology and microclimate ever carried out within an agroforestry context. Its primary objective was to compile a comprehensive experimental database to improve the mechanistic understanding of tree/crop interactions and support the development and validation of process-based simulation models describing resource capture and tree and crop growth in semi-arid agroforestry systems.
Grevillea robustaA. Cunn. (grevillea) trees were grown as mono-cultures or in mixtures with cowpea (Vigna unguiculata
L.) or maize (Zea maysL.) over a 68-month period. Allometric approaches were used to determine seasonal and annual
growth increments for leaf area and leaf, branch and trunk biomass in grevillea. Crop performance was examined during each growing season, while the spatial distribution of tree and crop roots was established during the latter stages of the experiment using coring and mini-rhizotron approaches. Detailed hydrological studies examined effects on the soil water balance and its components (precipitation, interception, runoff and soil moisture status); equivalent measurements of spatial and temporal variation in microclimatic conditions allowed the mechanistic basis for beneficial and detrimental effects on understorey crops and the influence of proximity to trees on crop performance to be examined. Transpiration by grevillea and water movement through lateral and tap roots were measured using sap flow methodology, and light interception by the tree and crop canopies was routinely determined.
This multi-disciplinary study has provided a detailed understanding of the changing patterns of resource capture by trees and crops as agroforestry systems mature. This paper provides an overview of the underlying rationale, experimental design and core measurements, outlines key results and conclusions, and draws the attention of readers to further papers providing more detailed consideration of specific aspects of the study. © 2000 Elsevier Science B.V. All rights reserved.
Keywords:Agroforestry;Grevillea robusta; Kenya; Maize; Microclimate; Productivity
∗Corresponding author. Tel.:+44-115-9516337; fax:+44-115-9516334.
E-mail address:[email protected] (C.R. Black)
1Present address: Department of Agriculture and Food Studies, University of Plymouth, Seale-Hayne Campus, Newton Abbot, Devon TQ12
6NQ, UK.
2Present address: British Waterways, Llanthory Warehouse, Gloucester Docks, Gloucester GL 1 2EJ, UK.
1. Introduction
Agroforestry has attracted considerable attention because of its potential to maintain or increase pro-ductivity in areas where high energy input, large-scale agriculture is impractical (Kidd and Pimentel, 1992). It is often assumed that appropriate agroforestry systems can provide the essential ecological functions needed to ensure sustainability and maintain microclimatic and other favourable influences, and that such benefits may outweigh their greater use of water in areas of limited water availability. Young (1989) suggested that trees and shrubs play an essential role in minimising erosion by reducing runoff, so improving water conservation. However, supporting evidence is either limited (Lal, 1989; Kiepe, 1995) or extrapolated from plantations or natural forests (Sanchez, 1987).
The hydrological and biological factors determining the success or failure of agroforestry systems are still poorly understood, although the existence of traditional agroforestry systems under arid and semi-arid condi-tions provides evidence that they have a useful role even in the water-limited environments of north-west India (Shankarnarayan, 1984) and the Sahel (Vandenbelt and Williams, 1992). Agroforestry research in dryland ar-eas falls into two broad categories: firstly, studies of microclimatic and soil amelioration around scattered trees in traditional parkland systems (Kater et al., 1992; Kessler, 1992; Vandenbelt and Williams, 1992; Belsky et al., 1993; Jonsson, 1995); and secondly, alley crop-ping systems, in which closely planted trees are pruned regularly to produce hedges (Singh et al., 1989; Rao et al., 1991; Ong et al., 1992). Several previous agro-forestry trials have been criticised on the grounds that interference resulting from the extension of tree roots into adjacent mono-crop control plots may have been underestimated; such effects would introduce a bias in favour of the agroforestry treatment (Rao et al., 1993). The present paper describes a trial involving Gre-villea robustaA. Cunn., maize (Zea mays) and cowpea (Vigna unguiculata) in which interference by tree roots was controlled by periodic trenching around the plots. Recent alley cropping studies at Machakos, Kenya in-volvingSenna spectabilisgrown in rotation with maize and cowpea (McIntyre et al., 1996, 1997) have shown that there was little difference in light interception or evapotranspiration between mono-crop and alley crop-ping treatments, and hence no improvement in
pro-ductivity during the cropping season. This observa-tion is consistent with other alley cropping trials at Machakos involving root trenching and species such asLeucaena leucocephala(Govindarajan et al., 1996) which demonstrated that alley cropping is detrimen-tal to crop productivity in semi-arid areas because soil fertility benefits resulting from the application of tree mulch or increased root turnover are outweighed by increased competition for water. Other studies indi-cate that regular pruning encourages the proliferation of fine tree roots near the soil surface, decreasing spa-tial niche separation between tree and crop roots and hence the potential for complementarity in the use of below-ground resources (Van Noordwijk and Purno-mosidhi, 1995). Grevillea was chosen for the present study owing to its popularity with farmers in Machakos District and because experimental studies of root distri-bution (Laycock and Wood, 1963; Jonsson et al., 1995) suggested it may extract substantial quantities of wa-ter from beneath the crop rooting zone. Grevillea is a fast-growing member of the Proteacae with potential as a multi-purpose cash crop as its timber is suitable for flooring and the manufacture of plywood and light fur-niture, and its flowers are suitable for honey production (Harwood and Getahun, 1990).
1.1. Rationale
runoff or deep drainage resulting from increased shad-ing, reduced soil temperature, the windbreak effect of the trees, or increased abstraction of water at depth or during the dry season would increase the proportion of rainfall used for transpiration. A possible disadvantage is that interception losses resulting from the evapora-tion of rainfall from the tree canopy may range from 10 to 30% in agroforestry systems (Ong and Black, 1994), although interception losses are lower when the tree canopy is sparse (5–10%; Wallace et al., 1995).
The hypothesis that agroforestry may increase pro-ductivity by capturing a larger proportion of the annual rainfall (Ong et al., 1992) was supported by the Hyder-abad studies, which demonstrated that improvements in annual rainfall utilisation from 40 to 80% were possi-ble in perennial pigeonpea (Cajanus cajan)/groundnut agroforestry systems, primarily because the use of off-season rainfall was increased (Marshall, 1995). These observations demonstrate the potential of agroforestry for temporal complementarity in areas where signifi-cant rainfall occurs outside the normal cropping season. However, the short-term nature of these experiments, often involving only one or 2 years of measurements, made it impossible to assess the long term implications. The presence of trees may also modify microclimatic conditions in ways that improve the water use effi-ciency of understorey crops, although regular pruning limits the extent of such effects in alley cropping sys-tems (Wallace, 1996). Several factors may be involved. Firstly, shading by the trees may increase the fraction of available water used for transpiration by decreasing soil evaporation, particularly when the crop canopy is sparse and rain is received as frequent, low intensity events. Under these circumstances, any reduction in the quantity of radiation reaching the soil decreases evaporation as this process is primarily energy-limited. Decreased windspeed at ground level may also limit evaporation. Secondly, agroforestry may confer mi-croclimatic benefits by decreasing the air temperature, windspeed and saturation deficit experienced by un-derstorey crops, thereby reducing evaporative demand (Monteith et al., 1991). In C3 crops, in which pho-tosynthesis becomes light saturated at relatively low irradiances, the reduced flux of photosynthetically ac-tive radiation (PAR) resulting from partial shading may have little effect on assimilation (Stirling et al., 1990), although this is less likely to apply to C4 species with their much higher light-saturated photosynthetic rates.
The potential benefits of shade are therefore likely to depend on tree spacing and age, canopy structure, in-cident radiation, shading intensity and the photosyn-thetic pathway of the understorey crop. Thirdly, shad-ing may alter the surface temperature of understorey crops in ways that benefit their phenology and produc-tivity (Monteith et al., 1991; Vandenbelt and Williams, 1992). In areas of high incident radiation and ambi-ent temperature, tissue temperatures frequambi-ently exceed optimal levels in unshaded crops, particularly during drought periods; under such conditions, partial shade may exert an ameliorative influence by bringing tem-peratures within the optimum range.
There are, therefore, several mechanisms whereby agroforestry may use available water more effectively than mono-crops and improve microclimatic condi-tions for understorey crops. The key question is whether the potential benefits outweigh the detrimental influ-ence of competition for water and nutrients between trees and crops. The present study, extending over a pe-riod of almost 6 years, is one of the most comprehensive and highly instrumented agroforestry experiments ever attempted. Its objectives were to quantify the changing influence of trees on the microclimatic and hydrolog-ical conditions experienced by understorey crops un-der semi-arid conditions as the system matured, and to establish the consequences for water use, light inter-ception and tree and crop performance. Fig. 1 shows the main categories of measurements, the periods when they were conducted, and seasonal rainfall. This paper describes the experimental design, instrumentation and measurement protocols, evaluates effects on the atmo-spheric and soil environment, and examines their influ-ence on tree and crop growth. More detailed consider-ation of the soil water balance, partitioning of light and water between the trees and crops, system productiv-ity and the modelling aspects of the work is presented elsewhere.
2. Materials and methods
2.1. Site description
Table 1
Seasonal and annual rainfall (mm) at Machakos: historical means and recorded values for CIRUS between planting and project end (October 1991 to June 1997)
Averagea 1991 1992 1993 1994 1995 1996 1997
Long growing season (1 March to 31 July) 359 229 261 112 199 302 235 245b
Dry season (1 August to 30 September) 9 16 2 4 8 11 8 n/a
1991–92 1992–93 1993–94 1994–95 1995–96 1996–97 Short growing season (1 October to 28/29 February) 414 404 773 381 628 337 168
Annual total 782 653 675 799 810 666 507 n/a
aData for 9-year period (1963–1971) from Machakos Maruba Dam station. bData available only until 7 July 1997.
above the Maruba river. Climate and soil were typical of the Kenyan uplands (Scott et al., 1971), where rainfall is bimodal with rainy seasons extending from March to June (long rains) and October to December (short rains); little rain is received between July and Septem-ber. Mean rainfall during the short and long growing seasons between 1963–1971 was, respectively, 414 and 359 mm, with an annual average of 782 mm (Table 1). Seasonal rainfall during the experimental period ranged from 112 to 773 mm, but the variation in annual rainfall was smaller (651–811 mm). Monthly potential evapo-ration varies between 95 and 165 mm, giving an annual total of 1450 mm (Huxley et al., 1989). The climate is relatively cool, with annual mean, minimum and max-imum temperatures of 20.1, 13.8 and 23.2◦C; monthly mean relative humidity ranged from 56% during the day to 96% at night.
2.2. Experimental design and management
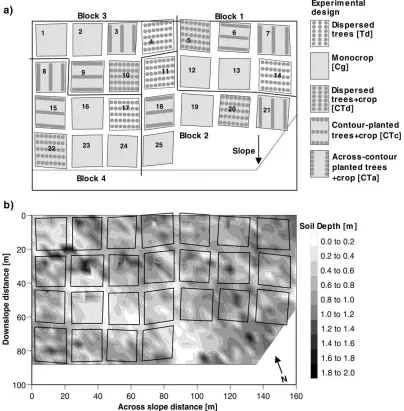
CIRUS examined the extent of complementarity be-tweenG. robusta(A. Cunn.; grevillea) and associated crops in the use of above and below-ground resources. The 0.6 ha site had no previous cropping history be-fore being cleared of scrubby vegetation dominated byAcaciaspecies in July 1991. The trial comprised a balanced incomplete block design containing five treatments replicated four times (Fig. 2a); the remain-ing five plots were mono-crop controls used for mea-surements not reported here. The five treatments were:
• Cg — mono-crop maize (Z. maysL, Katumani com-posite) or cowpea (V. unguiculataL. Walp cv.‘K80’) planted at intra- and inter-row spacings of 0.3 m×1 m and 0.15 m×0.5 m
• Td — dispersed mono-culture trees planted at 3 m×4 m spacing, 35 trees per plot (833 trees ha−1)
• CTd — dispersed trees with crops; trees planted at 3 m×4 m spacing, 35 trees per plot (833 trees ha−1)
• CTc — contour-planted tree rows with crops; tree
spacing 2 m×9 m, 30 trees per plot (640 trees ha−1) • CTa — across-contour tree rows with crops; tree
population as for CTc.
Three-month-old grevillea saplings were planted in October 1991. A rotation of maize and cowpea was initially adopted during the short and long growing seasons, but maize was grown continuously from the 1994 long rains onwards because infection by root-rot (Fusarium udum) caused substantial yield losses in cowpea. The mono-crop plots (Fig. 2a) had a maximum of two sides adjacent to agroforestry plots to minimise the risk of interference. The plots were separated by grass walkways, and a strip of vetiver grass (Vetiver zizanoidesL.) was contour-planted across the centre of each plot to control erosion. These were cut to a height of 10–15 cm at 7–14 day intervals to minimise compe-tition with the trees and crops and ensure they did not constitute a significant component of the water balance. No fertilisers or organic residues were applied.
Fig. 2. (a) Experimental design of CIRUS; the rhomboidal shape of some plots resulted from the contour planting of the tree rows; (b) soil depth measured at 4 m×4 m grid intervals across the site; depths between grid intersections were interpolated to produce the depth contours.
observations. Lateral extension of grevillea roots was examined before each rainy season to ensure they had not extended into adjacent plots; trenches were dug to a depth of 0.75 m to sever such roots.
2.3. Soil characteristics
The soil was a well-drained, shallow to moderately deep (0–2.5 m) sandy clay loam characterised as a
2.62 g cm−3 in the eroded bedrock; all horizons had moisture retention (pF) characteristics typical of sandy or sandy clay soils. Further details are given by Jackson et al. (1997). Chemical analyses showed the soil was not nutrient-limiting for crop growth before CIRUS was established, and had an organic carbon content of 1% (Kiepe, 1995). Soil pH, organic carbon con-tent and available ammonium, nitrate, potassium, phos-phate, calcium and magnesium levels were determined in May 1996 to establish treatment effects on nutri-ent status. The only detectable effects were that NH4+ and NO3−levels were higher and phosphate content was lower in mono-crop plots than in the agroforestry treatments (p<0.01; Lott, 1998). None of the nutrient concentrations or other soil variables examined were limiting for crop growth.
A survey conducted at 4 m×4 m grid intervals in
February 1993 using a 5 cm diameter auger showed that soil depth above the bedrock varied greatly (Howard, 1997). Additional points were added during the instal-lation of neutron probe access tubes (June to October 1993) and root distribution studies carried out between October 1995 and August 1997. Fig. 2b shows that a band of relatively shallow soil (0.2–0.6 m) extended between the upper north-west and the lower south-east corner of the site. Soil depth was included as a variate in the ANCOVA analysis of crop yields described by Howard (1997).
Detailed studies of climatic and microclimatic ditions, resource capture and productivity were con-centrated on the Cg, CTd and Td treatments using the approaches outlined below.
2.4. Meteorology
Dry and aspirated wet-bulb air temperatures, wind speed and incident solar radiation were measured 2 m above the tree canopy in the centre of the site, and at adjacent positions under the tree canopy, 0.3 and 2.5 m from the base of trees. The height of the above-canopy instruments was adjusted before each rainy season to allow for tree growth. Rainfall was measured using an automatic tipping-bucket raingauge located 20 m uphill from the experimental site.
2.5. Radiation interception
Incident solar radiation, net radiation and photosyn-thetically active radiation (PAR) were determined.
In-terception by the trees and crops was measured using unscreened tube solarimeters (Delta-T Devices, Cam-bridge, UK) located beneath both canopies and refer-enced against incident values recorded above the tree canopy or at ground-level outside the experimental area. Groups of three solarimeters were used to integrate in-terception across adjacent crop rows or concentric rings around trees in the Cg, Td and CTd treatments during the four growing seasons between the 1994 long (L94) and 1995/96 short (S95/96) seasons. Detailed measure-ments of light interception by the trees were made us-ing six Kipp solarimeters mounted on a 3 m arm which moved over the ground area enclosed between four ad-jacent trees in the Td and CTd treatments. This sys-tem provided 48 measurements within the 12 m2 sam-pling area (hereafter defined as ‘cell’), thereby allowing spatial variation in the radiation environment beneath the trees to be quantified. Net radiation was measured 0.75 m above an area of bare soil and at the same height beneath the tree canopy 0.3 and 2.5 m from trees in the Td and CTd treatments.
2.6. Hydrological measurements
2.6.1. Rainfall interception and runoff
Throughfall beneath the tree and crop canopies in the Cg, Td and CTd treatments was measured using man-ually recorded raingauges (127 mm diameter) refer-enced against the tipping bucket gauge and an identical manual gauge sited on bare soil. Stemflow gauges were attached to trees in the Td and CTd treatments. These comprised a flexible plastic collar attached to the trunk 0.75 m above the ground using silicone sealant (RS Components, Corby, Northants, UK) which drained into plastic 35 l Jerrycans. 2.5 m×20 m runoff plots leading to 1 m3collection tanks were installed in three replicate Cg, Td and CTd plots. Sub-samples of the water/soil slurry mixture were evaporated to dryness to determine the quantities of water lost as runoff and soil eroded.
2.6.2. Soil evaporation
in the CTd treatment. The lysimeters were weighed twice daily (08:00–09:00 and 17:00–18:00 hours) following rainfall during cropping seasons. Potential soil evaporation (Eso) was calculated using the Penman–Monteith formula (Monteith, 1965), and hourly means were recorded for wet and dry-bulb air temperatures, wind speed and net radiation. HourlyEso values were summed for comparison with daily mea-surements ofEs.
2.6.3. Soil water storage
A TDR system (SoilMoisture, Trase I) was used; this comprised ‘triple-wire’ sensors multiplexed to a central signal processing and logging unit. The waveguides were inserted horizontally in groups of four, at depths of 0.05, 0.15, 0.25 and 0.35 m in the Cg plots and at distances of 0.3, 1.0, 1.2, 1.5, 2.0 and 2.5 m from trees in the Td and CTd treatments. A site-specific calibration was established, and the sensors were logged hourly from November 1993 onwards.
Aluminium access tubes (44.5 mm diameter) were installed to 0.6–1.8 m, depending on soil depth. Eight tubes were installed in three replicates of the Td and CTd treatments, and four in three replicate Cg plots. Tube locations in the Td and CTd plots were identical to the TDR sensor positions, and were chosen to allow the influence of proximity to trees to be examined. Soil water content was measured weekly at 0.2 m depth in-tervals using an IH neutron probe (Didcot Instruments, Abingdon, UK). Calibrations were established for each depth interval and tube by Centre d’Etudes Nucleaires de Cadarache (France) using thermal neutron proper-ties. Over 300 calibrations relating soil water content (θv) to count rate and bulk density were performed. Moisture deficits were calculated by subtractingθv val-ues for specific horizons from the field capacity at a water potential of−0.05 MPa determined by moisture release analysis.
2.7. Tree growth
Allometric approaches were used to determine leaf, branch, trunk and total above-ground biomass and leaf area. During the first 28 months, these were based on measurements of tree height (H), basal diameter (BD), diameter at breast height (DBH) and total leaf number. Trunk volume was calculated as (Howard, 1997):
V =BD×0.33π
BD
2 2
H (1)
Trunk biomass was calculated as the product of trunk volume and the density of grevillea wood (0.47 g cm−3) determined by destructive analysis. Leaf area was es-timated as the product of leaf number per tree, mean leaf dry weight (1.52 g per leaf), and specific leaf area (SLA; 50.2 cm2g−1); mean leaf weight and SLA were determined for 1000 leaves sampled from three to six trees in each plot.
This approach could not be used during the later stages of the trial because the allometric relationships based on tree height and basal diameter were altered by pruning (Lott, 1998). The pipe model theory (War-ing et al., 1982; Whitehead et al., 1984) was therefore modified to allow growth to be determined from mea-surements of trunk diameter immediately below the lowest branch. Allometric relationships based on lin-ear regression analyses of log-transformed data were developed to provide reliable estimates of leaf area, to-tal above-ground biomass and the biomass of leaves, branches and trunks; these enabled tree growth to be determined non-destructively throughout the experi-mental period (Lott et al., 2000a).
2.8. Crop growth
Growth analysis during the first two seasons was re-stricted to the fresh and dry weights of pods and haulms at final harvest in cowpea (S91/92) and the stover, cobs and grain of maize (L92). Maize failed completely during the 1993 long rains (L93) because of the very low (<25% of seasonal average) and poorly distributed rainfall (Table 1). Regular destructive growth analy-ses of cowpea were carried out during the S92/93 and S93/94 growing seasons, and row-wise measure-ments of pod and haulm yields were made at maturity.
seasons using linear regression analyses based on paired weekly destructive and non-destructive measurements of plants reflecting the full range of growing conditions and plant sizes present. The relationships were based on measurements of basal stem diameter, height to the youngest expanded leaf and the top of the canopy, and leaf number. Leaf, stem, cob and grain fresh and dry weights were determined at maturity, and land equiva-lent ratios (LER) and performance ratios (PR; Willey, 1985; Rao et al., 1990, 1991) were calculated to es-tablish treatment effects on system performance. Full details are given by Lott et al. (2000c).
The timing of germination, floral initiation, anthesis, silking and physiological maturity was determined to establish treatment effects on the duration of the veg-etative, reproductive and grain-filling periods. Floral initiation was determined by dissecting the whorl to establish the presence of tassle primordia. Anthesis and silking were defined as occurring when spikelets emerged from the leaf whorl and silks (stigmata) ap-peared from the husk; maturity was defined either by the formation of a black layer at the pedicel of at least one grain per cob (Daynard and Duncan, 1969), or when 90% of the leaf area had senesced during sea-sons when CTd maize failed to set grain.
2.9. Root studies
Roots were sampled twice during each of the seasons indicated in Fig. 1, when the maize had 6–8 leaves and at anthesis. On each occasion, soil cores were taken to the bedrock in three replicate Cg, Td and CTd plots and separated into 0.2 m depth increments. These were sampled along a transect between a maize plant or grevillea tree and the mid-point between rows. The cores were collected using a 4.8 cm i.d. auger driven into the soil using a hydraulic hammer and extracted using a jack. Roots were washed from the soil, placed in vinegar and stored at 4◦C before being separated into maize and grevillea roots, which were distinguish-able on the basis of colour and morphology; dead roots were discarded. Root lengths were determined using a flat-bed scanner (ScanJet IIcx, Hewlett Packard, Palo Alto, CA, USA) and image-analysis software (DT-Scan, Delta-T Devices, Cambridge, UK). The lengths of proteoid roots were measured after shaving off the rootlets into water and collecting them by filtration be-fore scanning. Root dry weights were also determined.
Mini-rhizotrons were used to allow changes in root length to be estimated over a shorter time-scale than is possible using coring approaches. A new design based on innovations described by Gijsman et al. (1991) was used. Aluminium frames with open sides were installed in square access holes in the soil profile and lined with inflatable rubber bladders which eliminated air-filled gaps between the mini-rhizotron and the surrounding soil, minimising root proliferation along the walls of the access frames. To observe root growth, the bladders were deflated and removed before inserting an endo-scope. A 30 cm2circular area of soil was photographed at each sampling depth using a 35 mm SLR camera. Fifteen mini-rhizotrons were installed to the bedrock in December 1995, six each in randomly selected Td and CTd plots and three in the Cg treatment, using a sampling pattern similar to that used for the coring measurements. Photographs taken at 0.1 m depth in-crements at weekly intervals during the 1996 long and short rains and 1997 long rains were used to examine distribution of roots in maize and grevillea.
2.10. Sap flow
Sap flow through the trunks and lateral roots of gre-villea was measured using heat balance gauges; tech-nical details are given by Khan and Ong (1995), Lott et al. (1996) and Howard et al. (1997). Trunk gauges were calibrated as described by Khan and Ong (1995) and sap flow was corrected for variation in trunk di-ameter as reported by Lott (1998). These procedures enabled sap flow to be determined to within 7–10% for trunks and roots up to 10 cm in diameter (Khan and Ong, 1995; Lott et al., 1996; Howard, 1997), a level of accuracy consistent with that reported for other species (Baker and van Bavel, 1987; Ishida et al., 1991; Valan-cogne and Nasr, 1993). Measurements were made dur-ing all croppdur-ing seasons from the 1992/93 short rains onwards and periodically during the dry seasons (Fig. 1).
2.11. Meristem and soil temperatures
measurements of soil temperature at a depth of 2 cm immediately adjacent to maize plants until ca. 45 days after sowing (DAS), when stem elongation began; there-after thermocouples were inserted into the meristem. Soil temperature initially provides a good approxima-tion of meristem temperature in cereals as the meristem remains below the soil surface until stem elongation begins (Norman et al., 1984).
3. Results and discussion
3.1. Tree and crop growth
The trees were well established after 4.5 years, with a mean height of ca. 8 m and a basal diameter of ca. 0.15 m (Lott et al., 2000b). Regular pruning of the basal branches ensured that the trees produced single trunks and precluded excessive shading of the under-storey crops. The experimental period reported here encompassed nine cropping seasons, during which un-usually low (112 mm), high (777 mm) and average (ca. 350 mm) rainfall were experienced (Table 1); the vary-ing quantity and distribution of rainfall and increasvary-ing maturity of the trees resulted in substantial changes in tree/crop interactions.
3.1.1. Grevillea
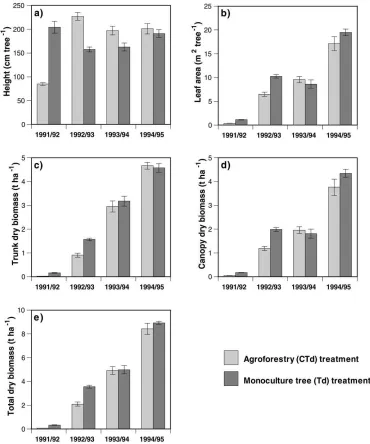
Development of suitable allometric procedures (Lott et al., 2000a) was essential if reliable estimates of tree growth were to be obtained and resource use efficiency calculated from concurrent measurements of water use and radiation interception. Growth was initially more rapid in mono-culture trees (Td) than in the agroforestry treatment (CTd), as is apparent from the annual in-crements for height, leaf area, trunk, canopy and total above-ground biomass (Fig. 3). The values for all pa-rameters except height were greater in Td than in CTd trees (p<0.05) during 1991/92 and 1992/93, suggesting that trees in the latter treatment were adversely affected by competition with associated crops for above and below-ground resources during the establishment pe-riod. However, CTd trees exhibited growth rates com-parable to the Td treatment from 1993/94 onwards. The annual height increments (Fig. 3) and daily mean growth rates (Lott et al., 2000b) were greater in Td trees during 1991/92 but were subsequently lower than in CTd grevillea (p<0.05). Tree height in the CTd
treatment, therefore, recovered to sole tree levels by January 1994 and thereafter was similar in both treat-ments. However, trunk biomass in CTd grevillea never recovered fully from the setback caused by competi-tion with associated crops during tree establishment, permanently reducing their economic value (Lott et al., 2000b).
3.1.2. Crop growth
Table 2 shows above-ground biomass and grain yield at final harvest for the nine cropping seasons between S91/92 and S95/96. Cowpea was grown during the S91/92, S92/93 and S93/94 seasons and maize in all other seasons. As the shade net treatments were first introduced in L94 season, data are not available for earlier seasons. Detailed analyses of crop and system performance are presented by Lott et al. (2000c).
The results clearly demonstrate the changing influ-ence of the trees on crop performance. Grain yield did not differ significantly between treatments during the first three cropping seasons, despite rapid tree growth and substantial inter-seasonal variation in rainfall (Table 1). Significant treatment effects were first ap-parent in S93/94, when biomass and grain yield were higher (p<0.001) in Cg than in CTd cowpea, although it is possible that the unusually high rainfall during the preceding S92/93 season (773 versus long-term aver-age of 414 mm) may have mitigated the competitive ef-fect of the trees in the CTd system. From L94 onwards, above-ground biomass and grain yield were greatly re-duced in CTd maize, with almost no yield being ob-tained in three of the final four seasons. Yield reached ca. 50% of that for unshaded Cg maize only when rain-fall was well above average (628 versus long-term av-erage of 414 mm) during S94/95, suggesting that water availability was the primary limitation for crop growth once the trees had established.
Fig. 3. Annual mean growth increments for: (a) height, (b) leaf area, (c) trunk dry biomass, (d) canopy dry biomass and (e) total above-ground dry biomass in the monoculture (Td) and dispersed agroforestry (CTd) treatments of grevillea. Double standard errors of the mean are shown.
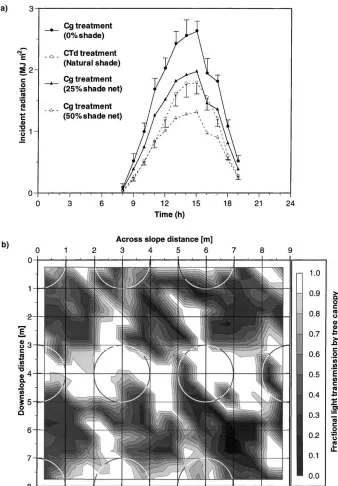
seasons (L94 and S95/96), but were lower during the seasons of greatest soil water availability (S94/95 and L95; p<0.05). These results indicate that shade was not solely responsible for the marked reduction in pro-ductivity in CTd maize, as the mean intensity of shade imposed by grevillea was intermediate between the Cg 25% and Cg 50% shade net treatments (cf. Fig. 4a).
3.2. Solar radiation
established. Incident radiation decreased with increas-ing shadincreas-ing intensity in the shade net treatments. The values for CTd maize, calculated on the assumption that shading was uniform throughout the cell bounded by four neighbouring trees, were intermediate between the Cg 25% and Cg 50% treatments; however, in reality, the discontinuous nature of the tree canopy caused substan-tial local variation in shading depending on proximity to the trees and solar angle. Mean daily incident radi-ation values for understorey crops in the CTd system were influenced by time since last pruning and hence the intensity of shade provided by the tree canopy (Lott, 1998; Lott et al., 2000b).
Fig. 4b illustrates the shading pattern under the tree canopy in the CTd treatment determined using the mov-able solarimeter array before the trees were pruned on 21 May 1996. As the site had a slope of between 18
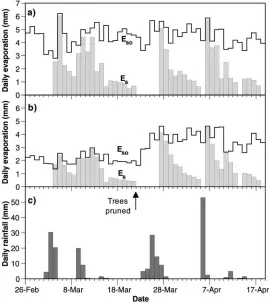
Fig. 5. Comparison of soil evaporation (Eso, solid bars), measured using micro-lysimeters, and potential soil evaporation (Eso, stepped line)
calculated using the Penman–Monteith formula for: (a) bare soil and (b) soil 0.3 m from the base of the trees in the CTd treatment; data are for the 1995 long growing season; (c) timing and quantity of rainfall.
and 22%, the shadows cast by the trees were signifi-cantly larger than their projected canopy areas (shown as circles), even when measurements were made at so-lar noon, as in Fig. 4b. Contrary to expectation, the area of soil beneath the trees was in direct sunlight at mid-day, implying that localised soil evaporation would be high at this time. Mean radiation interception by the trees bounding the 3 m×4 m cell was 40%. This and other information will enable processes such as soil evaporation to be scaled up from point measurements to obtain values expressed on an area average basis (cf. Wallace et al., 1998).
3.3. Soil evaporation
0.3 m from trees in the CTd treatment. Penman– Monteith (Monteith, 1965) potential evaporation (Eso) values for each sampling position and daily rainfall are also shown.Es andEsowere higher for bare soil than close to the trees, although the calculatedEsoand mea-suredEsvalues agreed closely at both locations on the first day after rainfall.EsandEsoincreased sharply after trees in the CTd treatment were pruned. These results suggest that Stage 1 evaporation, during which evap-oration rate is dependent on the quantity of radiation reaching the ground, lasted for no more than 1 day fol-lowing rainfall in the bare soil treatment. Evaporation rates for soil close to trees were lower during the first day after rain, with the result that Stage 1 lasted for ap-proximately 2 days. These observations suggest that the presence of trees may affect the fraction of rainfall lost as soil evaporation depending on the frequency of rain-fall: where rainfall occurs as a series of small rainfall events separated by 1 or 2 days, trees may significantly reduceEs; conversely, where rainfall occurs as a lim-ited number of larger storms separated by a week or more, trees may have less effect on seasonalEsvalues. More detailed analysis of these measurements is given elsewhere (Wallace et al., 1998; Jackson and Wallace, 1999).
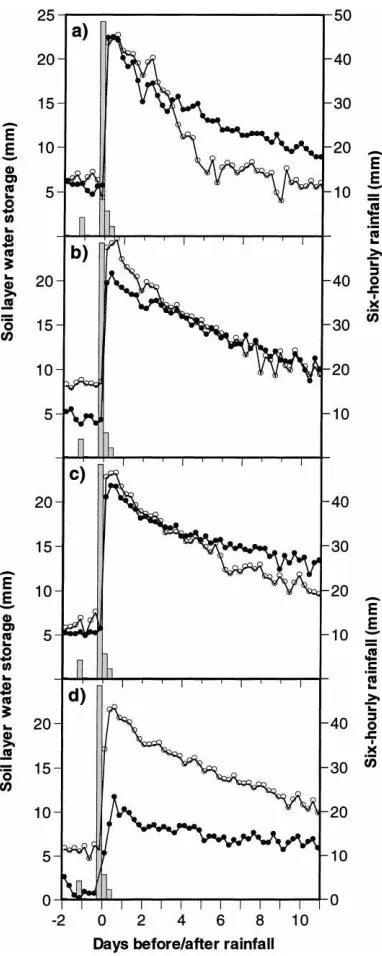
3.4. Soil water content
Fig. 6 shows the changes in surface soil water con-tent following an isolated rainfall event in the Td treat-ment during the S94/95 season. Overall, there was a greater increase in soil water storage 2.5 m from trees than at 0.3 m, particularly in the 0.3–0.4 m horizon. Although this effect may have resulted from differ-ences in infiltration rate, it is more likely that a com-bination of changes in rainfall interception, stemflow and micro-runoff affected net rainfall at the two po-sitions. The rate at which surface soil water content decreased after rain was greater 2.5 m from trees than at 0.3 m, as shown by the changes in the 0–0.1 m layer. It may be possible to correlate the differing rates of decrease in soil water content with differences inEs and/or the patterns of water uptake by tree roots at var-ious radial distances from trees and at different soil depths.
During the 1994 long rains, the soil and saprolitic rock below 1.2 m were much drier in the CTd than in
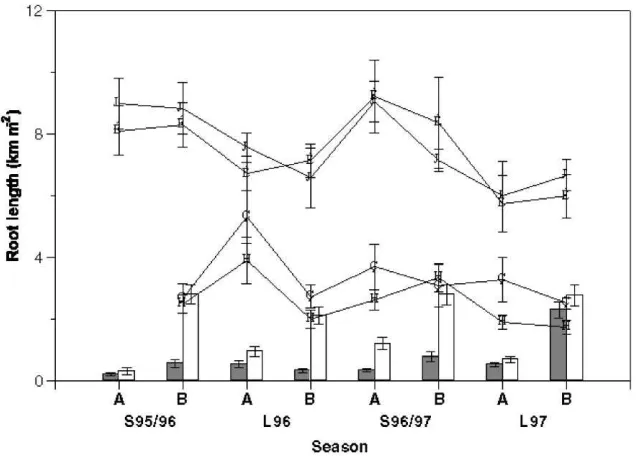
Fig. 7. Mean root lengths per unit land area for ordinary (solid lines, circles) and proteoid roots (dotted lines, triangles) of grevillea in the Td (open symbols) and CTd (closed symbols) treatments; histograms show root lengths for maize in the Cg (closed bars) and CTd (open bars) treatments at the 6–8 leaf stage (A) and anthesis (B). Double standard errors of the mean are shown.
the Td or Cg treatments. Abstraction of water from this layer had previously occurred during an extended dry period (ca. 8 months) between February and Octo-ber 1993 (Wallace et al., 1995) but subsequent seasonal rainfall was insufficient to recharge the soil profile, sug-gesting that little or no water was abstracted from the rocky substratum at this stage of the trial. However, the trees were able to abstract water from greater depths during the preceding prolonged drought at a time when available moisture was still present.
3.5. Root length distribution
The vertical and horizontal distributions of root length density were determined in the Td and CTd treatments on eight occasions during the four cropping seasons between L94 and S95/96; over 10 000 samples were collected, washed and scanned. Fig. 7 summarises the dataset and shows mean root lengths per unit ground area for grevillea and maize in the Cg and CTd treat-ments. Root lengths were always much greater in gre-villea than in maize, but decreased between the short and long rains; the reasons for this are unclear, although the reduction prior to the 1997 long rains may have
resulted at least in part from the pruning of the trees in October 1996. During the four seasons examined, the presence of well-established trees invariably sup-pressed both the growth of maize roots (Fig. 7) and yields at final harvest (Table 2); this effect was severe in all except the final season, when above-average rainfall permitted good maize growth beneath the trees. More detailed analysis of these studies is given by Smith et al. (1999).
3.6. Sap flow
Fig. 8. Diurnal time courses on three consecutive days during the 1995/96 short growing season for (a) saturation deficit and net radiation and (b) sap flow in trees in the Td and CTd agroforestry treatments. Double standard errors of the mean are shown.
an effect which Hatton and Vertessy (1990) attributed to time lags between sap flow at the measurement height (breast height or 1.5 m) and transpiration by the canopy. However, as the sap flow gauges were installed imme-diately below the canopy in the present study, the poten-tial for lags between sap flow at the measurement point
for Td and CTd grevillea, in a manner similar to that adopted previously for the tree component of other agroforestry systems (Marshall et al., 1994; Ong et al., 1996). Detailed analysis of water use by the trees and crops is presented elsewhere (Lott, 1998; Lott et al., 2000d).
3.7. Meristem temperature
Season-long measurements of meristem temperature in unshaded Cg maize and maize grown under natu-ral or artificial shade (Lott, 1998) revealed a substan-tial moderating influence of shade on the thermal en-vironment. The mean diurnal range experienced by Cg 0% maize varied from 10◦C in S94/95 to 20◦C in S95/96, whereas the corresponding values for CTd maize ranged from 9 to 13◦C. Mean maximum meris-tem meris-temperature in CTd maize was also reduced by up to 6◦C relative to Cg maize. Similar temperature mod-eration resulting from the presence of tree canopies has been reported by Ovalle and Avendano (1987), who recorded 3–10◦C differences in maximum soil temper-ature in an Acacia woodland relative to unshaded con-trol plots. In the present study, mean meristem temper-atures were greater throughout the diurnal time-course under 25 and 50% artificial shade than in CTd maize during S94/95 (p>0.05), emphasising the substantial moderating influence of the tree canopy. However, this effect disappeared during the later stages of the trial, with the result that mean meristem temperatures were higher in CTd maize than in the shade net treatments (p<0.001) during S95/96. This alteration in tempera-ture moderation resulted from continued tree growth and the periodic removal of the basal branches, which progressively increased the distance between ground level and the base of the tree canopy to over 2 m, thereby increasing airflow through the understorey environment. Thus, shading by overstorey trees may either increase or decrease the temperatures experienced by under-storey crops and hence the rates of thermal time ac-cumulation and plant development. As temperature is an important rate-modifier for growth and development (Monteith, 1977; Garcia-Huidobro et al., 1982), the in-fluence of shade on understorey crops may, therefore, be beneficial or detrimental depending on whether the temperatures experienced by unshaded crops are above, below or close to the optimum.
3.8. Concluding remarks
The present study has provided a detailed mecha-nistic understanding of the changing influence ofG. robustaon the microclimatic and hydrological condi-tions experienced by understorey crops as the trees in semi-arid agroforestry systems establish and mature, and has established the consequences for water use, light interception and tree, crop and system perfor-mance. Trees in the agroforestry treatment (CTd) grew more slowly than those in sole stands (Td) during the first 600 days after planting (Lott et al., 2000b) because of competition with associated crops for available re-sources; the above-ground biomass of CTd trees never recovered fully, reducing their economic value. Crop biomass and grain yield in the CTd system were not sig-nificantly affected during tree establishment, but were negligible during three of the final four seasons, attain-ing 50% of the values for unshaded mono-crop maize only when rainfall was much above average (Lott et al., 2000c). The conclusion that water availability was the primary limitation for crop growth once the trees had established was supported by the performance of maize grown under nets which provided shade in the absence of competition for water and nutrients. Crop growth was consistently greater than in CTd maize and often comparable to unshaded mono-crop maize (Lott et al., 2000c), demonstrating that shade alone was not respon-sible for the substantial yield losses in CTd maize.
Proximity to trees significantly altered understorey microclimatic conditions and water availability, par-ticularly during the later stages of the trial; thus shad-ing decreased the mean diurnal temperature range and maximum meristem temperature by up to 7◦C relative to mono-crop maize. The non-uniform shade provided by the trees introduced substantial spatial variation in the rate of thermal time accumulation, thereby inducing systematic local variation in the development rate and yield of the understorey crops. Similar spatial variation in incident radiation, saturation deficit and soil water availability also contributed to the systematic variation in the performance of understorey crops (Lott, 1998).
sea-son except during tree establishment, when the rapidly growing roots exploited deep moisture reserves within the soil profile (Jackson et al., 1998). Measurements of sap flow through lateral roots demonstrated that gre-villea trees may extract substantial quantities of water over 2 m from the trunk, increasing their competitive influence on associated crops (Lott et al., 1996).
A central objective was to provide core information to support the development and validation of process-based simulation models describing resource capture, tree and crop growth and the soil water balance of agroforestry systems in semi-arid regions. The dataset obtained provides detailed functional information con-cerning tree/crop interactions in an overstorey agro-forestry system over an extended period and at a level of detail that will be important in achieving this goal. Data for cropping seasons providing contrasting grow-ing conditions have already been used by Lott et al. (1998) to test output from the HyPAR model (Law-son et al., 1996), and there is considerable scope for using the database to support the future development of robust models capable of simulating tree and crop performance in semi-arid environments.
Acknowledgements
This publication is an output from a research project funded by the Department for International Develop-ment of the UK. However, the DepartDevelop-ment for Interna-tional Development can accept no responsibility for any information provided or views expressed. This work was funded from projects R4853, R5810, R6363 and R6364 of the Forestry Research Programme. Addi-tional technical and material support was provided by ICRAF, The University of Nottingham, The Royal So-ciety, the UK Natural Environment Research Council and the Swedish International Development Agency. The authors thank Raphael Maweu and field staff at Machakos Research Station for their support and Jackie Humphreys for excellent secretarial assistance.
References
Baker, J.M., van Bavel, C.H.M., 1987. Measurement of mass flow in the stems of herbaceous plants. Plant, Cell Environ. 10, 777–782. Belsky, A.J., Mwonga, S.M., Amundson, R.G., Duxbury, J.M., Ali, A.R., 1993. Comparative effects of isolated trees on their
undercanopy environments in high- and low-rainfall savannas. J. App. Ecol. 30, 143–155.
Cannell, M.G.R., Van Noordwijk, M., Ong, C.K., 1996. The central agroforestry hypothesis: the trees must acquire resources that the crop would not otherwise acquire. Agrofor. Syst. 34, 27–31. Cooper, P.J.M., Keatinge, I.D.H., Hughes, G., 1983. Crop
evapotranspiration — a technique for calculation of its components by field measurements. Field Crops Res. 7, 299–312. Daynard, T.B., Duncan, W.G., 1969. The black layer and maturity in
corn. Crop Sci. 9, 473–476.
Garcia-Huidobro, J., Monteith, J.L., Squire, G.R., 1982. Time, temperature and germination of pearl millet (Pennisetum typhoidesS. & H.). 2. Constant temperature. J. Exp. Bot. 33, 288–296.
Gijsman, A.J., Floris, J., Van Noordwijk, M., Brouwer, G., 1991. An inflatable minirhizotron system for root observations with improved soil/tube contact. Plant and Soil 134, 261–269. Govindarajan, M., Rao, M.R., Mathuva, M.N., Nair, P.K.R., 1996.
Soil-water and root dynamics under hedgerow intercropping in semiarid Kenya. Agron. J. 88, 513–520.
Green, S.R., Clothier, B.E., 1988. Water use of kiwifruit vines and apple trees by the heat-pulse technique. J. Exp. Bot. 39, 115–123. Harwood, C.E., Getahun, A., 1990. Australian tree finds success in
Africa. Agrofor. Today 2, 5–8.
Hatton, T.J., Vertessy, R.A., 1990. Transpiration of plantationPinus radiataestimated by the heat pulse method and the Bowen ratio. Hydrol. Proc. 4, 289–298.
Howard, S.B., 1997. Resource Capture and Productivity of Agroforestry Systems in Kenya. Ph.D. thesis. University of Nottingham, UK, 259 pp.
Howard, S.B., Ong, C.K., Black, C.R., Khan, A.A.H., 1997. Using sap flow gauges to quantify water uptake by tree roots from beneath the crop rooting zone in agroforestry systems. Agrofor. Syst. 35, 15–29.
Huxley, P.A., Pinney, A., Gatuma, D., 1989. Development of Agroforestry Research Methodology Aimed at Simplifying the Study of Potential Tree/crop Mixtures. Final Report, Number 1-432-60005613. April 1989, ICRAF, Nairobi, Kenya, 109 pp. Ishida, T., Campbell, G.S., Calissendorff, C., 1991. Improved heat
balance method for determining sap flow rate. Agric. For. Meteorol. 56, 36–48.
Jackson, N.A., Wallace, J.S., 1999. Soil evaporation measurements in an agroforestry system in Kenya. Agric. For. Met. 94, 203–215. Jackson, N.A., Wallace, J.S., Smith, D.M., Roberts, J.M., 1997. Water Balance of Agroforestry Systems on Hillslopes — Phase II. Annual Report to DFID Forestry Research Programme. Institute of Hydrology, Wallingford, UK, 11 pp.
Jackson, N.A., Smith, D.M., Roberts, J.M., Wallace, J.S., Ong, C.K., 1998. Water Balance of Agroforestry Systems on Hillslopes. Final Report to the Forestry Research Programme. UK Department For International Development, Institute of Hydrology Report IH/DFID 98/3, 65 pp.
Jonsson, K., Fidgeland, L., Maghembe, J.A., Högberg, P., 1995. The vertical distribution of fine roots of five species and maize in Morogoro, Tanzania. Agrofor. Syst. 6, 63–69.
Kater, L.J.M., Kante, S., Budelman, A., 1992. Karite (Vitellaria papradoxa) and nere (Parkia biglobosa) associated with crops in south Mali. Agrofor. Syst. 18, 89–105.
Kessler, J.J., 1992. The influence of karite (Vitellaria paradoxa) and nere (Parkia biglobosa) trees on sorghum production in Burkina Faso. Agrofor. Syst. 17, 97–118.
Khan, A.A.H., Ong, C.K., 1995. Correction of systematic errors in estimates of transpiration obtained using a constant temperature heat balance technique. Exp. Agric. 31, 461–472.
Kidd, C.V., Pimentel, D., 1992. Integrated resource management. In: Agroforestry for Development. Academic Press, San Diego, USA, 233 pp.
Kiepe, P., 1995. No Runoff, No Soil Loss: Soil and Water Conservation in Hedgerow Barrier Systems. Ph.D. thesis. Wageningen Agricultural University, The Netherlands, 156 pp. Lal, R., 1989. Agroforestry systems and soil surface management of
a tropical alfisol: 2. Water runoff, soil erosion and nutrient loss. Agrofor. Syst. 8, 97–111.
Lawson, G.J., Cannell, M.G.R., Mobbs, D.C., Crout, N.M.J., Muetzelfeld, R., Wallace J.S., Gregory, P.J., Thomas, T.H., Jagtap, S., Ludlow, A.R., Cobbina, J., Arah, J., MacDonald, K.J., Taylor, J., Sharp, L., Wiggins, G., Flynn, L., Livesley, S., Willis, R.W., Friend, A.D., 1996. Agroforestry Modelling and Research Coordination Annual Report, 1995–1996. Institute of Terrestrial Ecology, Penicuik, UK, 178 pp.
Laycock, D.H., Wood, R.A., 1963. Some observations of soil moisture under tea in Nyasaland II. The effect of shade trees. Trop. Agric. (Trinidad) 40, 42–48.
Lott, J.E., 1998. Resource Capture and Use in Semi-arid Overstorey Agroforestry Systems. Ph.D. thesis. University of Nottingham, UK, 259 pp.
Lott, J.E., Ong, C.K., Black C.R., 1998. Comparison of output from HyPAR with observed maize yields in Kenya. Report prepared for the UK Department of International Development, Mounted on Website http:/www.ac.uk/ite/edin/agro/ampnews7.pdf, 45 pp. Lott, J.E., Black, C.R., Ong, C.K., 2000a. Allometric estimation of above-ground biomass and leaf area in managedGrevillea robusta agroforestry systems. Agrofor. Syst., in press.
Lott, J.E., Ong, C.K., Black, C.R., 2000b. Long term productivity of aGrevillea robusta-based agroforesty system in semi-arid Kenya I. Tree growth. For. Ecol. & Man., in press.
Lott, J.E., Ong, C.K., Black, C.R., 2000c. Long term productivity of aGrevillea robusta-based agroforestry system in semi-arid Kenya II. Crop growth and system productivity. For. Ecol. & Man., in press.
Lott, J.E., Ong, C.K., Black, C.R., 2000d. Long term water use of a Grevillea robusta-based agroforestry system in semi-arid Kenya. For. Ecol. & Man., submitted for publication.
Lott, J.E., Khan, A.A.H., Ong, C.K., Black, C.R., 1996. Sap flow measurements of lateral tree roots in agroforestry systems. Tree Physiol. 16, 995–1001.
Marshall, F.M., 1995. Resource Partitioning and Productivity in Perennial Pigeonpea/Groundnut Agroforestry Systems in India. Ph.D. thesis. University of Nottingham, UK, 324 pp.
Marshall, F.M., Black, C.R., Ong, C.K., 1994. Heat balance measurements of transpiration in perennial pigeonpea-groundnut agroforestry systems. In: Monteith, J.L., Scott R.K., Unsworth, M.H. (Eds.), Resource Capture by Crops. Nottingham University Press, Loughborough, UK, pp. 426–429.
McIntyre, B.D., Riha, S.J., Ong, C.K., 1996. Light interception and evapotranspiration in hedgerow agroforestry systems. Agric. For. Meteorol. 81, 31–40.
McIntyre, B.D., Riha, S.J., Ong, C.K., 1997. Competition for water in a hedgerow-intercrop system. Field Crops Res. 52, 151–160. Monteith, J.L., 1965. Evaporation and the environment. In: State
and Movement of Water in Living Organisms. Symposium of the Society for Experimental Biology, 1964. Cambridge University Press, Cambridge, pp. 205–234.
Monteith, J.L., 1977. Climate. In: de Alvim, P.T., Kozlowski, T.T. (Eds.), Ecophysiology of Tropical Crops. Academic Press, London, pp. 1–27.
Monteith, J.L., Ong, C.K., Corlett, J.E., 1991. Microclimatic interactions in agroforestry systems. For. Ecol. Manage. 45, 31– 44.
Norman, M.J.T., Pearson, C.J., Searle, P.G.E., 1984.The Ecology of Tropical Food Crops. Cambridge University Press, Cambridge, pp. 103–109.
Ong, C.K., Black, C.R., 1994. Complementarity in resource use in intercropping and agroforestry systems. In: Monteith, J.L., Scott, R.K., Unsworth, M.H. (Eds.), Resource Capture by Crops. Nottingham University Press, Loughborough, UK, pp. 255–278. Ong, C.K., Odongo, J.C.W., Marshall, F., Black, C.R., 1992. Water use of agroforestry systems in semi-arid India. In: Calder, I.R., Hall, R.L., Adlard, P.G. (Eds.), Growth and Water Use of Plantations. Wiley, Chichester, pp. 347–358.
Ong, C.K., Black, C.R., Marshall, F.M., Corlett, J.E., 1996. Principles of resource capture and utilisation of light and water. In: Ong, C.K., Huxley, P. (Eds.), Tree-Crop Interactions: a Physiological Approach. CAB International, Wallingford, UK and ICRAF, Nairobi, Kenya, pp. 73–158
Ovalle, C., Avendano, J., 1987. Interactions of the tree layer with the herbaceous understorey layer in the plant-communities ofAcacia cavenin Chile 1. Tree influence on the botanical composition, production and phenology of the herbaceous stratum. Acta Oecol. 8, 385–404.
Rao, M.R., Muraya, P., Huxley, P.A., 1993. Observations of some tree root systems in agroforestry intercrop situations and their graphical representations. Exp. Agric. 29, 183–194.
Rao, M.R., Ong, C.K., Pathak, P., Sharma, M.M., 1991. Productivity of annual cropping systems on a shallow alfisol in semi-arid India. Agrofor. Syst. 15, 51–63.
Rao, M.R., Sharma, M.M., Ong, C.K., 1990. A study of the potential of hedgerow intercropping in semi-arid India using a two-way systematic design. Agrofor. Syst 11, 243–258.
Sanchez, P., 1987. Soil productivity and sustainability in agroforestry systems. In: Steppler, H., Nair, P.K.R. (Eds.), Agroforestry: a Decade of Development. ICRAF, Nairobi, Kenya, pp. 205–226. Schulze, E.-D., Cermak, J., Mtyssek, R., Penka, M., Zimmermann,
Scott, R.M., Webster, R., Lawrence, C.J., 1971. A Land System Atlas of Western Kenya. Christchurch, Hampshire, UK.
Shankarnarayan, K.A., 1984. Agroforestry in Arid and Semi-arid India. Central Arid Zone Institute, Publication 24, Jodphur, India, 295 pp.
Singh, R.P., Ong, C.K., Saharan, N., 1989. Above and below-ground interactions in alley cropping in semiarid India. Agrofor. Syst. 9, 259–274.
Smith, D.M., Jackson, N.A., Roberts, J.M., Ong, C.K., 1999. Root distributions in aGrevillea robusta-maize agroforestry system in semi-arid Kenya. Plant and Soil 211, 191–205.
Stirling, C.M., Williams, J.H., Black, C.R., Ong, C.K., 1990. The effect of timing of shade on developments dry matter production and light use efficiency in groundnut (Arachis hypogaeaL.) under field conditions. Aust. J. Agric. Res. 41, 633–644.
Valancogne, C., Nasr, Z., 1993. Une methode de mesure du debit de seve brute dans de petits arbres par bilan de chaleur. Agronomie 9, 609–617.
Vandenbelt, R., Williams, J.H., 1992. The effect of soil surface temperature on the growth of millet in relation to the effect of Faidherbia albida trees. Agric. For. Meteorol. 60, 93– 100.
Van Noordwijk, M., Purnomosidhi, P., 1995. Root architecture in relation to tree-crop-soil interactions and shoot pruning in agroforestry. Agrofor. Syst. 30, 161–173.
Wallace, J.S., 1991. The measurement and modelling of evaporation from semiarid land. In: Sivakumar, M.V.K., Wallace, J.S., Renard,
C., Giroux, C. (Eds.), Soil Water Balance in the Sudano-Sahelian Zone. Proceedings of International Workshop, Niamey, Niger, February 1991, IAHS Publ. No. 199, IAHS Press, Institute of Hydrology, Wallingford, UK, pp. 131–148.
Wallace, J.S., 1996. The water balance of mixed tree-crop systems. In: Ong, C.K., Huxley, P.A. (Eds.), Tree-Crop Interactions: a Physiological Approach. CAB International, Wallingford, UK, pp. 189–233.
Wallace, J.S., Jackson, N.A., Ong, C.K., 1995. Water Balance of Agroforestry Systems on Hill Slopes. Final Report to the Forestry Research Programme, UK Overseas Development Administration, Institute of Hydrology, Wallingford, ODA 95/10, 39 pp.
Wallace, J.S., Jackson, N.A., Ong, C.K., 1998. Modelling soil evaporation in an agroforestry system in Kenya. Agric. For. Met. 94, 31–44.
Waring, R.H., Schroeder, P.E., Oern, R., 1982. Application of the pipe model theory to predict canopy leaf area. Can. J. For. Res. 12, 556–560.
Whitehead, D., Edwards, W.R.N., Jarvis, P.G., 1984. Conducting sapwood area, foliage area and permeability in mature trees of Pinus sitchensisandPinus contorta. Can. J. For. Res. 13, 405– 409.
Willey, R.W., 1985. Evaluation and presentation of intercropping advantages. Exper. Agric. 21, 119–133.