IDENTIFIKASI KERAGAMAN GEN FOLLICLE STIMULATING HORMONE RECEPTOR (FSHR Alu-1) PADA SAPI LOKAL INDONESIA DENGAN TEKNIK PCR-RFLP
Teks penuh
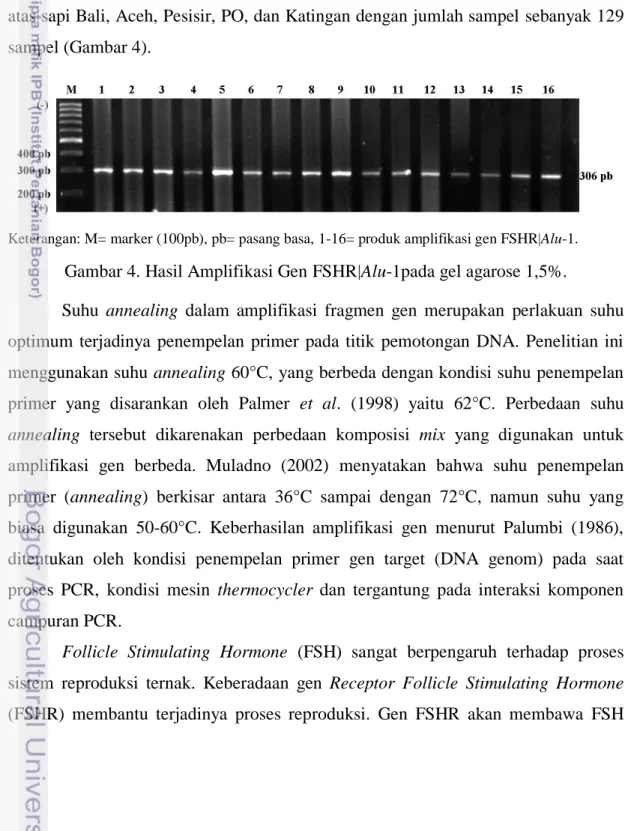
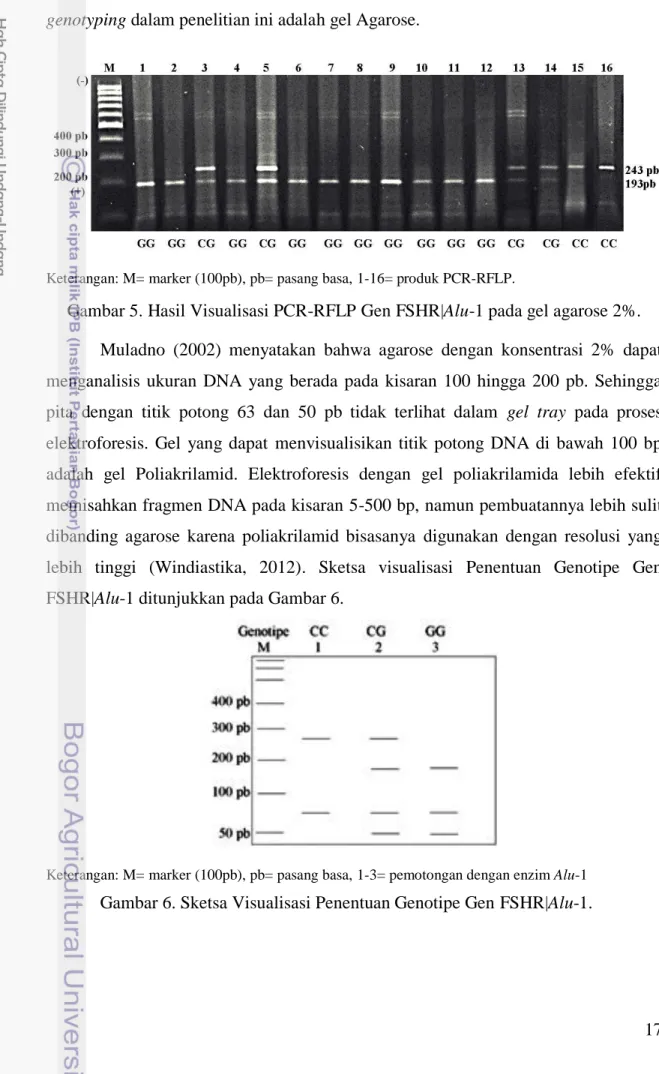
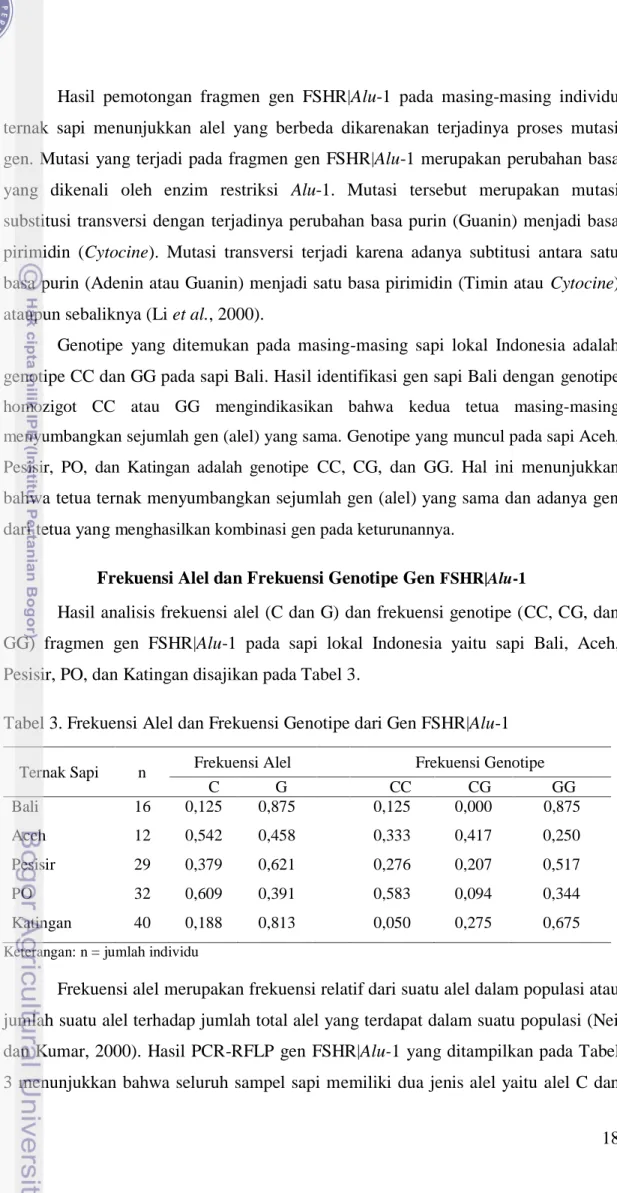
Gambar




Garis besar
Dokumen terkait
Tujuan dari penelitian ini adalah untuk mengidentifikasi keragaman gen DGAT1 dan SCD pada sapi bali di Balai Pembibitan Ternak Unggul Sapi Bali dengan menggunakan metode
Berdasarkan hasil analisis tersebut, dapat dikatakan bahwa gen calpastatin (CAST| Msp I) bersifat monomorfik pada populasi bangsa sapi Indonesia yaitu pada sapi
Hasil sekuens fragmen gen GHR|AluI sapi bali, limousin, simmental, dan pesisir dianalisa kesamaannya (homology) dengan sekuen yang terdapat di GenBank menggunakan perangkat lunak
SNP g.125550A>T gen FTO bersifat polimorfik pada bangsa sapi potong Indonesia yang terdiri atas: sapi madura, sapi pesisir, sapi katingan, sapi PO, sapi pasundan,
Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Desember 2015 ini ialah ayam lokal, dengan judul Identifikasi Kegaraman Gen Growth Hormone
Meskipun dalam penelitian ini tidak ada pola yang khusus mengenai fragmen DNA hasil pemotongan, namun sudah dapat diketahui bahwa sapi bali yang ada di Pulau Bali menunjukkan
Tujuan dari penelitian ini adalah mengidentifikasi keragaman gen Diacylglycerol O-acyltransferase 1 (DGAT1) menggunakan enzim restriksi Eae I dengan metode PCR-RFLP pada sapi
Manfaat penelitian ini adalah agar dapat menjadi informasi dasar tentang keragaman gen Follicle Stimulating Hormone FSH pada itik Pitalah Sumatera Barat dan dapat dimanfaatkan sebagai
