Dr. Fathurrahman, SP., M.Sc
Kultur Jaringan dan Transformasi
Genetika Kelapa
Sawit
Kultur Jaringan dan Transformasi Genetika Kelapa Sawit
Dr. Fathurrahman, SP., M.Sc
Hak Cipta © Dr. Fathurrahman, SP., M.Sc, 2023
Sanksi Pidana Pasal 113 ayat (3) UUHC No. 28 Tahun 2014:
Setiap orang yang dengan tanpa hak dan atau tanpa izin pencipta atau pemegang hak melakukan pelanggaran hak ekonomi pencipta sebagaimana dimaksud dalam pasal 9 ayat (1) huruf a, huruf b, huruf e, dan/atau huruf g, untuk penggunaan secara komersial dipidana dengan pidana penjara paling lama 4 (empat) tahun dan/atau pidana denda paling banyak Rp 1.000.000.000,00 (satu miliar rupiah).
Editor :
Prof. Dr. Ir. Hasan Basri Jumin, M.Sc Dr. Mardaleni, SP., M.Sc
Heriyanto, SP, M.Si
Layout & Cover : KALYSTA
ISBN : 978-623-6598-68-9 Penerbit : UIR PRESS- Pekanbaru
Anggota IKAPI : 015/Anggota Luar Biasa/RA/2022 Dicetak oleh : UIR PRESS
Gedung Serbaguna Universitas Islam Riau
Jl. Kaharuddin Nasution No. 113 Perhentian Marpoyan Pekanbaru, 28284
Email: [email protected]
Dr. Fathurrahman, SP., M.Sc
iv
KATA SAMBUTAN
Assalamu'alaikum warahmatullahi wabarakatuh
Keberadaan buku yang baru terbit dengan judul “Kultur Jaringan dan Transformasi Genetika Kelapa Sawit” yang ditulis oleh Dr. Fathurrahman, SP., M.Sc sangat bermanfaat untuk menjadi bahan bacaan dan rujukan bagi para dosen, mahasiswa dan peminat bidang pertanian khususnya Pemuliaan Tanaman. Penulis menyajikannya dalam bentuk paparan yang lebih dalam tentang Teknik kultur jaringan kelapa sawit yang dilakukan penembakan partikel emas yang dilumuri oleh gen desaturase dalam aturan antisense ke jaringan sasaran yaitu embrio belum matang. Inisiasi ekplan menjadi dua sasaran yaitu regenerasi langsung dan inisiasi embriogenik kalus. Analisis konfirmasi sampel tertransformasi juga mendalam karena melibatkan analisis PCR dan gas kromatografi untuk konfirmasi keberhasilan dalam transformasi genetik.
Dengan membaca buku ini ide untuk mengembangkan tanaman transgenik dengan beberapa metode terus digali sehingga pengembangannya pemuliaan tanaman secara modern dapat dilakukan berdampingan dengan pemuliaan tanaman konvensional. Hal penting yang harus diperhatikan agar dapat bermanfaat dan pertanian Indonesia akan lebih maju ke depannya. Saya sebagai Dekan Fakultas Pertania
Universitas Islam Riau mengucapkan selamat atas terbitnya buku ini, semoga kelak akan terbit buku yang lain yang berkaitan dengan pemuliaan tanaman untuk memperkaya khasanah sumber keragaman genetik tanaman. Sekali lagi selamat dan semoga dapat terus berkarya untuk kemajuan bangsa dan negara. Wassalamualaikum warahmatullahi wabarakatuh.
Dekan
Dr. Siti Zahrah, M.P.
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah subhanahu wa ta’ala atas segala karunia dan inayah-Nya sehingga buku referensi ini berhasil diselesaikan. Judul buku referensi ini adalah “Kultur Jaringan dan Transformasi Genetika Kelapa Sawit”. Buku ini diharapkan dapat memberikan sumbangan pemikiran bagi peningkatan ilmu pengetahuan dan teknologi (IPTEK) sekarang dan dimasa yang akan datang.
Buku ini berisi tentang bioteknologi, yaitu bagaimana aplikasi kultur jaringan yang berupakan dasar dari proses menghasilkan tanaman transgenik. Media kultur jaringan berpengaruh penting dalam menyediaan eksplan sebagai jaringan sasaran dalam transfer gen. Oleh karena itu perlu dilakukan beberapa jenis media untuk untuk ditentukan yang terbaik untuk mendukung pertumbuhan eksplan bagi menjadi planlet langsung maupun melalui kalus dan embrio somatis.
Selain dari itu dalam buku ini dijelaskan penggunaan bahan kimia tertentu sebagai penanda seleksi juga penting untuk menyeleksi eksplan mana saja yang tertransformasi gen asing.
Dalam bab ini penggunaan teknologi gen gun dengan menggunakan alat biolistik dapat meningkatkan efektifitas dalam transfer gen ke dalam nukleus eksplan. Penggunaan
Dr. Fathurrahman, SP., M.Sc
vi
plasmid pADST 35 yang mengandung gen desaturase dalam aturan antisense berfungsi untuk meningkatkan biosintesis asam lemak jenuh. Sehingga dengan peningkatan persentase asam lemak jenuh berbanding asam lemak jenuh dapat digunakan untuk keperluan penggunaan senyawa skunder minyak kelapa sawit dijadikan berbagai produk untuk kepentingan manusia. Hasil tanaman trangenik yang diperoleh menunjukkan bahwa setelah dianalisis secara molekuler terbukti gen desaturase telah terintegrasi ke dalam nukleus kelapa sawit dura. Selanjutnya analisis menggunakan HPLC juga menunjukkan peningkatan kandungan asam lemak jenuh dan penurunan kandungan asam lemak tak jenuh pada sampel tertransformasi dan tanaman ini disebut tranaman transgenik.
Buku ini diharapkan menjadi referensi bagi para Dosen, peneliti dan mahasiswa dalam melakukan penelitian di laboratorium. Selain itu untuk memahami tentang teknologi aplikasi teknik kultur jaringan kelapa sawit dan transfomasi genetika.
Ucapan terima kasih penulis sampaikan kepada Universitas Islam Riau Pekanbaru sebagai institusi yang telah mewadahi para dosen untuk melaksanakan Catur Dharma Perguruan Tinggi khususnya pada Dekan Fakultas Pertanian dan pada Program Studi Agroteknologi. Ucapan terima kasih juga disampaikan kepada semua pihak yang terlibat dalam baik secara langsung maupun tidak langsung dalam penerbitan buku ini.
Pekanbaru, 2 Oktober 2023 Penulis
Dr. Fathurrahman, SP., M.Sc
viii
BAB II
KELAPA SAWIT DAN KEPENTINGAN NYA ... 21
2.1 Kandungan Minyak Tumbuhan ... 21
2.2 Manfaat Minyak Nabati ... 23
2.3 Biosintesis Asam Lemak Tumbuhan ... 26
2.3.1 Jalur biosintesis asam lemak ... 26
2.3.2 Pentingnya Enzim ∆-9-stearoyl-ACP desaturase ... 30
2.3.3 Enzim asil-ACP tioesterase ... 32
2.3.4 Triasilgliserol Sintetik ... 33
2.3.5 Pemanjangan Asam Lemak (FAE) ... 36
2.4 Kultur Jaringan Tanaman Kelapa Sawit ... 38
2.4.1 Regenerasi Embrio Belum Matang (EBM) menjadi planlet ... 38
2.4.2 Faktor hormonal dalam kultur jaringan kelapa sawit ... 40
2.5 Penggunaan Teknologi Antisense ... 41
2.6 Metode Transformasi Genetik dan Rekayasa Genetika ... 42
2.6.1 Transformasi Biolistik ... 44
2.6.2 Transformasi yang dimediasi oleh Agrobacterium ... 48
2.6.3 Transformasi Protoplas ... 51
2.6.4 Penyerapan DNA Langsung ... 53
2.6.5 Metode Transformasi Agrolistrik ... 54
2.7 Pembibitan dan Transformasi Genetik Tanaman Minyak ... 55
Daftar Pustaka ... 58
BAB III
PENGARUH MEDIA TUMBUH KULTUR JARINGAN
TERHADAP EKSPLAN KELAPA SAWIT………..73
3.1 Bahan, Media Tumbuh, EBM kelapa sawit dan Basta….. 73
3.2 Regenerasi langsung ke planlet dan kalus dan kalus embriogenik ... 74
3.3 Regenerasi langsung ke planlet dan kalus dan kalus untuk embriogenik jaringan yang gun (ditembak) ... 79
3.4 Uji resistensi Pospinotrichinaacetyl trans ferase (PPT)… 81 3.5 Pengaruh Media Terhadap Kultur Jaringan ... 81
3.5.1 Regenerasi Langsung ke Planlet ... 81
3.6 Pembentukan Kalus dan Kalus Embriogenik ... 87
3.7 Uji Resistensi Endogen Eksplan Kelapa Sawit Terhadap Pospinothricine Acetyl Transferase (PPT) ... 91
Daftar Pustaka... 93
BAB IV TRANSFORMASI GENETIK DENGAN METODE BIOLISTIK ... 97
4.1 Pendahuluan... 97
4.2 Plasmid ... 98
4.3 Bahan kimia, Kit dan Enzim ... 99
4.4 Isolasi Plasmid ... 101
4.5 Proses Biolistik ... 102
4.5.1 Persiapan pembawa mikro ... 102
4.5.2 Pelapisan Pembawa Mikro dengan DNA ... 103
4.5.3 Penembakan pembawa mikro ke jaringan target ... 103
4.6 Analisa Molekul ... 105
4.6.1 Isolasi DNA genomik dari daun kelapa sawit... 105
4.6.2 Reaksi rantai polimerase (PCR) ... 106
4.6.3 Pemindahan eksplan ke Greenhouse ... 111
Dr. Fathurrahman, SP., M.Sc
x
4.7 Analisis Molekular Pada Tanaman Yang Diubah ... 112
4.7.1 Pencernaan konstruk pADST35 ... 112
4.7.2 Isolasi DNA genom ... 112
4.7.3 Analisis reaksi berantai polimerase ... 114
Daftar Pustaka………121
BAB V PERUBAHAN KANDUNGAN ASAM LEMAK SAWIT MELALUI REKAYASA TRANSFORMASI GENETIK ... 123
5.1 Profil asam lemak ... 123
5.2 Instrumentasi Gas Kromatografi ... 124
5.3 Instrumentasi Spekstroskopi Massa ... 126
5.4 Pengaruh Integrasi Gen Desaturase Terhadap Profil Asam Lemak ... 129
Daftar Pustaka ... 136
BAB VI KESIMPULAN ... 139
Glosarium ... 143
Dr. Fathurrahman, SP., M.Sc
xii
3.3 Tangkai buah sawit dura yang telah dipisahkan dari tandannya. Buah pada 1/4 ujung tangkai
dan 1/4 tangkai tidak digunakan ... 82 3.4 Eksplan yang berasal dari embrio kelapa sawit
(Elaeis guineensis Jacq. dura) yang belum
matang dikultur secara in vitro selama 2 minggu
pada media Y3 ... 82 3.5 Respon embrio kelapa sawit (Elaeis guineensis
Jacq. dura) yang belum matang setelah 5 bulan dikulturkan pada media yang diberi hormon
IBA 2 mg l-1 ... 84 3.6 A. Pertumbuhan dan perkembangan kalus .. kelapa
sawit (Elaeis guineensis Jacq.dura) setelah 5 bulan ... 88 B. Diferensiasi kalus embriogenik setelah 5,5 bulan, (1)kalus embriogenik dikultur pada media MS,
(2)kalus embriogenik dikultur pada media Y3 ... 89 3.7 Uji kultur embrio belum matang (EBM) pada media
N6 bebas hormon dengan konsentrasi herbisida pos- pinothricin acetyl transferase (PPT) yang berbeda ... 92 4.1 Urutan lengkap gen desaturase ∆9-stearoyl-ACP
dari E. guineensis Jacq tenera ... 100 4.2 Sistem Gene Gun (Biolistic PDS-1000/He; USA).
Teknologi senjata gen juga dikenal sebagai
teknologi tembakan partikel ... 104 4.3 Siklus PCR terdiri atas tiga tahap untuk
pereplikaan DNA ... 106 4.4 Mesin Thermal Cycle untuk amplifikasi PCR ... 107 4.5 Vektor pADST 35 ditransformasikan ke dalam sistem
kelapa sawit ... 113 4.6 Hasil elektroforesis gel agarosa konstruk pADST 35
dengan enzim HindIII ... 113
Kultur Jaringan dan Transformasi Genetika Kelapa Sawit
4.7 Hasil elektroforesis gel agarosa DNA genom
tanpa pencernaan ... 115 4.8 Elektroforesis gel agarosa hasil amplifikasi
konstruk pADST35 dan amplifikasi DNA genom
tanaman non-transformasi ... 117 4.9 Hasil elektroforesis gel agarosa DNA genomik dari
TRD1 dan TRDL1 tanaman yang ditransformasi
putatif ... 117 4.10 Peta duplikasi gen desaturase endogen
menggunakan primer DSATF6-DSATR3 ... 118 4.11 Hasil elektroforesis gel agarosa pADST35
membangun digesti dengan kombinasi
primer DSATF6-DSATR1... 119 4.12 Hasil elektroforesis gel agarosa DNA genomik
dari tanaman yang ditransformasi diduga
TRD6 ... 120 5.1 Alur proses kromatografi gas dalam mentukan
persentase asam lemak ... 123 5.2 Komponen kromatografi gas komponen
pelengkapnya ... 124
xiv
Dr. Fathurrahman, SP., M.Sc
2
1.2 Status Luasan Kelapa Sawit
Melihat trennya, luas perkebunan kelapa sawit Indonesia cenderung meningkat. Dalam 10 tahun terakhir luas perkebunan kelapa sawit melonjak 60,11%, tepatnya sejak 2011 hingga 2021.
Status mengenai pengusahaannya, swasta mencatatkan kepemilikan terbesar terhadap perkebunan kelapa sawit pada tahun lalu. Luasnya mencapai 8,04 juta ha. Adapun perkebunan kelapa sawit milik rakyat tercatat sebanyak 6,03 juta ha dan seluas 550.333 ha sisanya merupakan perkebunan kelapa sawit milik negara. Berdasarkan wilayahnya, Riau memiliki perkebunan kelapa sawit terbesar di Indonesia mencapai 2,86 juta ha. Jumlah tersebut setara dengan 19,62% dari luas perkebunan kelapa sawit Indonesia sepanjang tahun lalu. Luas urutan nomor 2 adalah Kalimantan Barat kelapa sawit yang seluas 2,11 juta ha disusul Kalimantan Tengah sebesar 1,88 juta ha.
1.3 Ekspor Minyak Sawit
Pandemi Covid-19 yang terjadi sejak awal tahun 2020 diperkirakan menyebabkan penurunan produksi CPO sebesar 5,01 persen dibanding tahun 2019 menjadi 45,74 juta ton. Pada tahun 2021, produksi CPO kembali mengalami sedikit penurunan menjadi 45,12 juta ton. Produksi minyak sawit (CPO) terbesar tahun 2021 diperkirakan berasal dari Provinsi Riau dengan produksi sebesar 8,96 juta ton atau sekitar 19,55 persen dari total produksi Indonesia. Produksi terbesar selanjutnya berasal dari Provinsi Kalimantan Tengah dengan produksi sebesar 7,28 juta ton atau 12,47 persen. Produksi perkebunan kelapa sawit di Indonesia menurut provinsi tahun 2021 selengkapnya dapat dilihat pada Tabel 3.2 .Berdasarkan status pengusahaannya, pada tahun 2020 sebesar 61,07 persen dari produksi minyak sawit (CPO) atau 27,94 juta ton minyak
sawit (CPO) berasal dari perkebunan besar swasta, sebesar 33,88 persen atau 15,50 juta ton dari perkebunan rakyat dan sisanya 5,05 persen atau 2,31 juta ton berasal dari perkebunan besar negara. Meskipun total produksi pada tahun 2021 diperkirakan mengalami penurunan, struktur produksi menurut status pengusahaan tidak jauh berbeda dari tahun-tahun sebelumnya, yakni didominasi oleh produksi perkebunan swasta dengan perkiraan sebesar 27,36 juta ton CPO (60,64 persen); diikuti perkebunan rakyat dengan total produksi 15,50 juta ton (34,36 persen); serta sisanya sebesar 2,26 juta ton (5 persen) diproduksi oleh perkebunan besar negara (lihat Gambar 1.1).
Walaupun volume ekspor cenderung meningkat, peningkatan tersebut tidak sesuai nilai ekspor minyak sawit yang cenderung fluktuatif. Berdasarkan publikasi World Bank Commodities Price Data. Pada tahun 2017 terjadi peningkatan rata-rata harga minyak sawit di pasar dunia sebesar 751 US$/
mt, sehingga menyebabkan peningkatan nilai ekspor hingga 27,32 persen dibanding tahun 2016 (Gambar 1.2). Peningkatan harga tersebut tidak berlangsung lama sebab pada tahun 2018 dan 2019, harga minyak sawit kembali mengalami penurunan menjadi 639
Gambar 1.1 Perbandingan Produksi Minyak Sawit Indonesia menurut Status Pengusahaan, 2021 (Sumber : https://www.bps.go.id/
publication /2022/11/30/)
Dr. Fathurrahman, SP., M.Sc
4
US$/mt pada 2018 dan 601 US$/mt pada 2019. Pada tahun 2020, harga minyak sawit di pasar dunia kembali melambung menjadi 752 US$/mt. Kenaikan harga tersebut menyebabkan peningkatan nilai ekspor sebesar 16,94 persen dibanding 2019, meskipun dari sisi volume ekspor minyak sawit mengalami penurunan sebesar 8,55 persen. Selanjutnya pada tahun 2021, harga minyak sawit kembali melonjak 1,5 kali lebih besar menjadi 1.131 US$/mt, sehingga menyebabkan peningkatan nilai ekspor secara signifikan menjadi 28,68 miliar US$.
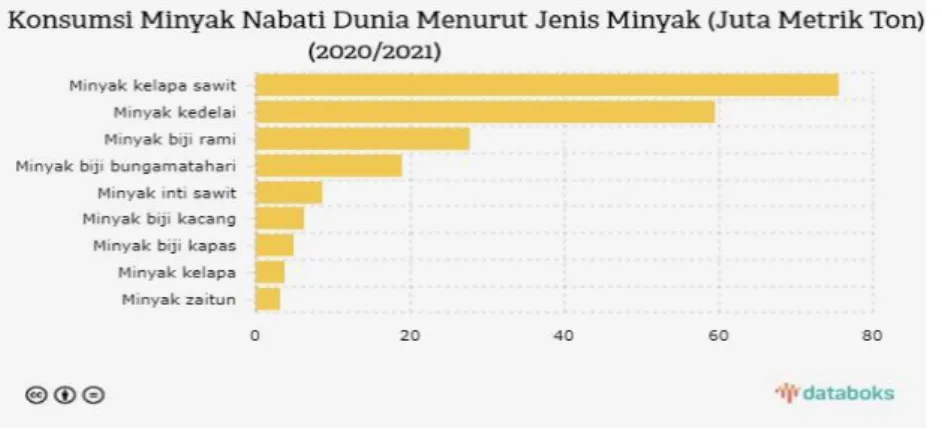
1.4 Persaingan Minyak Nabati Dunia
Kebutuhan konsumsi minyak dan lemak dunia semakin meningkat setiap tahunnya, sedangkan produksinya kurang dari permintaan. Hal ini merupakan peluang yang baik bagi komoditi kelapa sawit yang merupakan salah satu komoditi penghasil minyak dan lemak di dunia untuk terus meningkatkan produksi dan luas areal budidaya kelapa sawit untuk memenuhi permintaan konsumen. Hal ini menjadikan kelapa sawit sebagai komoditas pertanian terpenting dalam hal nilai ekspor. Namun, minyak
Gambar 1.2 Perkembangan Volume dan Nilai Ekspor Minyak Sawit, 2017-2021(Sumber : https://www.bps.go.id/ publication /2022/11/30/).
sawit menghadapi persaingan dari beberapa minyak dan lemak lainnya, yang terpenting adalah minyak kedelai (Glycine max), minyak bunga matahari (Helianthus annus), minyak biji rami ( ) minyak biji matahari, minyak inti sawit, minyak biji kacang, minyak biji kapas, minyak kelapa dan minyak seperti dapat dilihat pada Gambar 1.3.
Gambar 1.3 Konsumsi berbagai minyak nabati dunia (sumber: https //
google. com/search?q=kebutuhan+minyak+sawit.
Sebanyak 90% produksi minyak sawit digunakan dalam industri makanan (minyak masak tumbuhan) dan bahan bukan makanan (sabun, detergen dan surfaktan). Minyak tumbuhan telah menggantikan lemak hewan sebagai sumber utama lemak dalam bahan makanan dan kini menyumbang sebanyak 15-20% dari jumlah kalori dalam diet negara maju (Ohlrogge 1994). Salah satu faktor penyebab semakin populernya minyak nabati adalah kandungan asam lemak jenuhnya yang jauh lebih rendah dari lemak hewani, yaitu 40-50%. Karena peran asam lemak tak jenuh dalam kesehatan manusia belum ditentukan.
Penyelidikan tentang hubungan antara kadar kolesterol tinggi pada tikus yang diberi diet tinggi asam lemak jenuh dibandingkan dengan tikus yang diberi diet tinggi asam lemak tak jenuh (Bravo et al.1997). Minyak sawit olahan
yang memiliki nilai yodium lebih tinggi antara 60 sampai 65%
telah berhasil diproduksi, namun masih terdapat kekurangan pada hibrida yang mengikuti karakteristik E. oleifera dimana sebagian besar tandan matang sebelum mencapai kematangan (Sharma & Pau 1989).
1.5 Rekayasa Genetik Meningkatkan Daya Saing Minyak Nabati
Dengan meningkatnya persaingan minyak nabati dari sumber lain untuk minyak sawit, upaya perlu dilakukan untuk meningkatkan kualitas dan penggunaan minyak sawit. Metode pemuliaan telah digunakan dalam upaya meningkatkan karakteristik agronomis dan ekonomis tanaman kelapa sawit.
Hasil panen hibrida tenera (hibrida dura dan pisifera) menunjukkan peningkatan hasil 30% dibandingkan dengan varietas dura (Soh et al. 1994). Kerugian dari metode pemuliaan terbatas, karena kelapa sawit memiliki masa generasi yang panjang dan membutuhkan lahan yang luas untuk pembibitan (Rajanaidu et al. 1993).
Untuk saat ini dan masa akan datang, penggunaan metode rekayasa genetika merupakan pendekatan alternatif yang diharapkan dapat meningkatkan kinerja agronomi dan ekonomi melalui modifikasi komposisi asam lemak minyak sawit. Untuk mempertahankan persaingan kelapa sawit di pasar internasional, penggunaan metode rekayasa genetika untuk memperbaiki karakteristik pohon kelapa sawit menjadi penting. Hal ini disebabkan adanya persaingan dengan tanaman penghasil minyak lainnya seperti kedelai, jagung dan rapeseed.
Keberhasilan modifikasi komposisi asam lemak pada beberapa tanaman transgenik telah banyak dilaporkan. Diantaranya adalah peningkatan komposisi asam lemak pada biji rep transgenik melalui transfer antisense mRNA untuk stearoyl-
Dr. Fathurrahman, SP., M.Sc
8
ACP desaturase (Knutzon et al. 1992a) dan produksi asam laurat pada biji rep transgenik melalui transfer tiosterase lauroyl-ACP.
gen yang diperoleh dari California Bay (Voelker et al. 992). Pada dasarnya komposisi asam lemak suatu tanaman tergantung pada spesies, genus atau famili tanaman (Gunstone et al. 1986).
Sebagai contoh, minyak nabati yang diperoleh dari Brassicacea memiliki kadar asam erusat yang tinggi (20 – 50%), sedangkan minyak tumbuhan dari spesies Umbelliferae memiliki kadar asam petroselinat yang tinggi (60 – 85%) (Gunstone et al. 1986), kedua jenis ini asam lemak jarang ditemukan pada keluarga tumbuhan lain.
Metode rekayasa genetika selain dapat mempercepat proses peningkatan karakteristik agronomi dan ekonomi juga dapat mengatasi beberapa kendala utama yang dihadapi dalam metode pemuliaan tradisional seperti sumber gen yang akan ditransfer tidak terbatas pada spesies yang berkerabat.
Hal ini membuka peluang sumberdaya genetik yang sangat luas untuk dimanfaatkan. Terlepas dari kelebihan metode rekayasa genetika, dimungkinkan untuk melakukan modifikasi yang lebih spesifik dan dalam banyak kondisi dalam waktu yang lebih singkat (Gasser & Fraley 1991). Properti seperti ini memberikan peluang yang baik bagi minyak sawit untuk bersaing dengan pasar minyak nabati dunia. Sifat fisik minyak nabati tergantung pada komposisi asam lemaknya. Biasanya komposisi asam lemak suatu tanaman tidak dapat memenuhi kebutuhan semua industri yang luas. Jika ditinjau ada salah satu ciri yang menarik yaitu komposisi asam lemak pada mesocarp dan kernel berbeda secara kimiawi dan mencakup semua jenis asam lemak rantai pendek, sedang, dan panjang.
Asam lemak jenuh (asam palmitat dan asam stearat) dalam minyak sawit mendekati angka 50%. Oleh karena itu, rekayasa minyak sawit untuk menghasilkan minyak yang mengandung
komposisi asam lemak lebih tinggi akan meningkatkan nilai minyak sawit bagi industri makanan dan industri oleokimia.
1.5.1 Teknik kultur jaringan kelapa sawit
Dalam melakukan transformasi genetik suatu tanaman ada beberapa syarat yang harus dipenuhi. Pertama gen yang akan digunakan harus dimasukkan ke dalam sel melalui dinding dan membran sel plasma. Kedua, transgen kemudian harus ditranslokasikan ke dalam inti sel, di mana gen tersebut harus berintegrasi secara stabil ke dalam genom tanaman inang.
Ketiga, transgen harus diwariskan ke progeni dengan rasio yang sesuai dengan hukum Mendel untuk membuktikan bahwa transformasi berhasil secara stabil. Isolasi dan karakterisasi promotor spesifik jaringan harus dilakukan sehingga ekspresi transgen terbatas pada penyimpanan minyak tisu. Hal lain yang harus diperhatikan adalah bagaimana jaringan yang ditransformasi dapat secara efisien dan mudah dihasilkan menjadi planlet. Keberhasilan transformasi genetik kelapa sawit telah dilaporkan oleh Parveez (1997), Zainal (1998), Shahabuddin (2000), Rashdan (2000).
Kultur jaringan kelapa sawit pertama kali dilaporkan 46 tahun yang lalu. Namun demikian, terdapat beberapa klon yang dihasilkan memiliki karakteristik abnormal pada tingkat rumah kaca. Ini akan mencegah penggunaan lahan bertingkat yang dapat mengurangi hasil panen (Jones 1988). Untuk mengatasi masalah tersebut, metode teknologi DNA rekombinan untuk mendeteksi klon abnormal pada tahap pra- peternakan dapat digunakan (Shah et al. 1994).
Teknik kultur jaringan memiliki beberapa keunggulan dibandingkan dengan biji yaitu multiflikasi bahan tanaman lebih cepat dan unggul serta permanen (Kushairi et al., 2010).
Manfaat ini memungkinkan memperbaiki bahan tanaman
Dr. Fathurrahman, SP., M.Sc
10
yang ada sebelumnya dengan sifat unggul, misalnya produksi minyak yang tinggi, kualitas minyak bagus, pertumbuhan yang lambat tinggi dan tahan terhadap penyakit dengan sifat seragam sehingga produktivitas diharapkan akan tinggi.
Kultur jaringan juga membuka peluang untuk produksi bahan tanaman baru melalui rekayasa genetika (Sogeke, 1998), seleksi tumbuhan yang tahan terhadap cekaman abiotik seperti kekeringan dan salinitas (Perez-Clemente dan Gomez-Cadenas, 2011), produksi tanaman haploid menggunakan interkultur, pemilihan tanaman yang tahan penyakit (Bridgen et al., 2018) serta upaya konservasi sumberdaya genetik untuk tanaman tua dan sakit (Oseni, 2018).
Kultur jaringan kelapa sawit sebenarnya sudah diterapkan pertama pada tahun 1970 (Corley dan Tinker, 2015). Dari aspek botani, penerapan kultur jaringan pada tanaman kelapa sawit menguntungkan bagi perusahaan. Hal ini terkait kelapa sawit tergolong tanaman monokotiledon dengan memiliki meristem apikal. Perbanyakan dengan menggunakan benih adalah satu- satunya cara efisien, sebelum munculnya teknologi kultur jaringan (Soh et al., 2011). Beberapa bagian untuk teknik kultur jaringandapatdigunakansebagai eksplan, misalnya tunas (daun muda), akar, bunga (perbungaan), biji, dan embrio (Weckx et al., 2019). Eksplan memiliki kelemahan dan kepentingan sendiri jenis eksplan. Daun muda (janur) paling banyak digunakan.
Tambahan penggunaan janur adalah berlimpah pada tanaman kelapa sawit yang merupakan eksplan relatif steril (Setiowati et al., 2013). Tetapi sisi negatifnya adalah waktu pembentukan embrio somatis yang relatif lama dapat mencapai dua tahun.
Produksi eksplan normal akan berlanjut. Penggunaan eksplan bunga memiliki keuntungan tidak menyebabkan kerusakan tanaman yang serius, karena sampel yang diambil jauh dari target tumbuh, dapat diambil dalam periode 6-8 bulan dan
tingkat perkembangan embrio langsung sudah cukup tinggi, sehingga intensitas penggunaan zat pengatur tumbuh lebih sedikit. Menurut Guzman dan Peralta, (2010) kelemahan penggunaan eksplan bunga ini menghasilkan sedikit kalus dan gangguan pertumbuhan diamati produksi vegetatif dan buah relatif tinggi rendah.
Untuk mendapatkan ortet dengan kualitas yang baik diperlukan area yang luas selama proses seleksinya. Selain itu, diketahui juga bahwa tanaman klon cenderung memiliki sex ratio lebih tinggi dibandingkan tanaman hasil persilangan, sehingga membutuhkan suplai polen lebih banyak (Ginting et al., 1996; Kushairi et al., 2010). Pengembangan klon di Amerika Latin dan Afrika terutama terfokus pada beberapa aspek, seperti seleksi ortet dan evaluasi lapangan, manajemen penentuan tingkat keseragaman melalui statistik dan alat lainnya, serta masalah penyakit tanaman seperti layu Fusarium, Ganoderma dan busuk pucuk (Durand-Gasselin et al., 2010).
Dari bervariasinya kendala yang dihadapi oleh produsen klon kelapa sawit, terdapat dua tantangan utama yakni abnormalitas dan efisiensi proses dalam kultur jaringan. Berikut uraian mengenai tantangan kultur jaringan kelapa sawit yang secara umum dihadapi dan usaha pemecahan masalahnya.
1.5.2 Pengubahan komposisi asam lemak melalui transfer gen
Untuk mengidentifikasi enzim target untuk rekayasa genetika jalur biosintesis asam lemak minyak sawit, perlu dipahami jalur biosintesis asam lemak secara detail. Namun pengetahuan di bidang genetika molekuler kelapa sawit, khususnya jalur sintesis asam lemak, masih terbatas (Shah 1994). Spesifisitas substrat 3-ketoacyl synthase I tidak jelas dan penemuan baru enzim 3-ketoacyl-ACP synthase III (Jaworski et
Dr. Fathurrahman, SP., M.Sc
12
al. 1989) dan 3-ketoacyl-ACP synthase IV (Dehesh et al. 1998), menunjukkan adanya gen sintase yang belum ditemukan sejauh ini. Beberapa laboratorium sedang menyelidiki lokasi jalur biosintesis ini dan beberapa gen yang terlibat dalam produksi asam lemak telah berhasil diisolasi. Isolasi dan karakterisasi gen yang terlibat dalam sintesis asam lemak diperlukan untuk konstruksi vektor ekspresi yang cocok untuk transformasi asam lemak minyak sawit. Beberapa gen yang terlibat dalam produksi asam lemak telah berhasil diisolasi.
Diantaranya adalah gen asetil–KoA karboksilase, ketoasil sintase (KAS), protein pembawa asil (ACP), stearoil ACP-desaturase, asil ACP thiosterase, hidroksilase, lipoksigenase, desaturase asam lemak (Budziszweski et al. 1996). Beberapa gen telah diisolasi dari E.
guineensis var. tenera dan E. oleifera. Untuk gen lengkap seperti
∆9-stearoil ACP desaturase (Shah & Rashid 1997), β-ketoasil ACP sintase I (Rashid 1996), dan oleil thiosterase (Asemoto & Shah 1998). Untuk gen tidak lengkap juga telah dipencilkan seperti palmitoil-ACP thiosterase (Cha 2001) dan 3-ketoasil- ACP sintase III (Shahabuddin 2000). Enzim ∆9-stearoil-ACP desaturase berperan sebagai faktor penting dalam menentukan persentase komposisi asam lemak jenuh dan asam lemak tak jenuh (Ohlogge & Browse 1995; Harwood 1988). Adapun jalur transfer elektron dalam desaturasi asam lemak oleh stearoyl- CoA desaturase dapat dilihat pada Gambar 1.4.
1.5.3Penentuan jenis media kultur jaringan kelapa sawit Penentuan media kultur jaringan yang sesuai untuk multiplikasi eksplan kelapa sawit varietas dura yang tumbuh dengan cepat secara in vitro sangat diperlukan. Diharapkan di dalam sistem kultur jaringan, media kultur tidak menjadi faktor pembatas di dalam regenerasi tumbuhan yang akan ditransformasi. Selanjutnya mengetahui peranan gen desaturase
yang berasal dari endogenus tenera dapat terintegrasi atau tidak jika ditransformasikan ke dalam kelapa sawit varietas dura. Varietas tenera adalah hasil persilangan dura dan pisifera.
Dura digunakan sebagai sumber induk betina dan pisifera digunakan sebagai sumber induk jantan. Vektor antisense untuk gen desaturase telah diisolasi (Shahabuddin 2000) adalah dikendali- kan oleh promoter konstitutif ubikuitin. Langkah selanjutnya adalah pemindahan gen ke jaringan sasaran embrio belum matang dan kalus Elaeis guineensis Jacq. dura dengan menggunakan teknik transformasi biolistik. Eksplant sasaran yang telah menjadi planlet dilakukan analisis molekul iaitu ekstrak DNA genom, PCR dan analisis profil asam lemak untuk menentukan sama ada gen desaturase diintegrasi dan diekspresikan ke dalam genom kelapa sawit. Secara ringkas objektif kajian adalah sebagai berikut, penentuan media yang terbaik, untuk mendapatkan tumbuhan transgenik dura dengan menggunakan teknik biolistik, menganalisis inkorporasi gen stearoil-ACP desaturase dalam aturan antisense.
Gambar 1.4 Jalur transfer elektron dalam desaturasi asam lemak oleh stearoyl-CoA desaturase (SCD) (Chad et al. 2009).
Dr. Fathurrahman, SP., M.Sc
14
DAFTAR PUSTAKA
Abass, M.H., S.D. Al-Utbi, and E.A.R.H. Al-Samir. 2017.
Genotocixity assessment of high concentrations of 2,4-D, NAA and Dicamba on date palm callus (Phoenix dactylifera L.) using protein profile and RAPD markers. Journal of Genetic Engineering and Biotechnology 15: 287-295.
Bridgen, M.P., W.V. Houten, and T. Eeckhaut. 2018. Plant tissue culture techniques for breeding mark. Springer International Publishing, USA. Constantin.
Budziszewski, G.J., Craft, K.P.C. and Hildebrand, D.F. 1996.
Uses of biotechnology in modifying plant lipids. Lipids 31 : 557-569.
Chad M. Paton and James M. Ntambi. 2009. Biochemical and physiological function of stearoyl-CoA desaturase American Journal of Physiology-Endocrinology and Metabolism. 297:
1, E28-E37.
Corley, R.H.V. and P.B. Tinker. 2016. The oil palm. Edisi kelima.
Wiley Blackwell.
Corley, R.H.V., C.H. Lee, I.H. Law, and C.Y. Wong. 1986.
Abnormal flower development in oil palm clones. The Planter 62(723): 233-240.
Dehesh, K., Edwards, P., Fillatti, J., Slabaugh, M. and Byrne, J.
1998. KAS IV : A 3-ketoacyl-ACP synthase from Cuphea sp. is a medium chain specific condensing enzyme. Plant J. 15 (3) : 383-390.
Ditjenbun (Direktorat Jendral Perkebunan). 2018. Statistik perkebunan Indonesia komoditas kelapa sawit 2016-2018.
[01 Februari 2019].
Durand-Gasselin, T., A. Labeyrie, P. Amblard, F. Potier, B.
Cochard, and de Franqueville, B. 2010. Strategies to develop oil palm clones for Latin America and Africa. Proceedings on Advances in Oil Palm Tissue Culture. Yogyakarta, Indonesia.
Durand-Gasselin, T., A. Labeyrie, P. Amblard, F. Potier, B.
Cochard, de Franqueville, and B. Nouy. 2010. Strategies to develop oil palm clones for Latin America and Africa.
Proceedings on Advances in Oil Palm Tissue Culture.
Yogyakarta, Indonesia
Ernayunita, dan Y. Yenni. 2016. Multiplikasi embrio somatik kelapa sawit (Elaeis guineensis Jacq,) menggunakan media padat dan cair. WARTA PPKS 21(2): 59-63.
Ernayunita, H. Rahmadi, Y. Yenni, R. D. Setiowati, dan I.Y.
Harahap. 2019. Vegetative characterization to identify oil palm (Elaeis guineensis Jacq.) planlet abnormalities. AIP Conference Proceedings 2099: 1-8.
Ernayunita, H. Y., Rahmadi, I.Y., Harahap, dan A.R. Purba.
2016. Peran NAA, GA, karbon aktif, dan sukrosa dalam kultur embrio zigotik klon OG Hybrid (Elaeis guineensis Jacq. x Elaeis oleifera) open pollinated. Jurnal Penelitian Kelapa Sawit. 24(3): 115-126.
Ernayunita, H.Y. Rahmadi, dan Y. Yenni. 2017. Perbanyakan bahan tanam unggul kelapa sawit melalui kultur jaringan di PPKS. WARTA PPKS 21(4): 8-14.
Fatmawati, dan G. Ginting. 2000. Pembentukan embrio somatik langsung dari eksplan daunkelapa sawit (Elaeis guineensis Jacq.) WARTA PPKS 8(2): 57-61.
GAPKI (Gabungan Pengusaha Kelapa Sawit Indonesia). 2019.
https://gapki.id/news/14413/ sawit-indonesia-menyong song- awal- tahunyang-lebih menjanjikan. (diakses pada 10 April 2019).
Ginting, G., C. Mollers, dan K. Pamin. 1996. Embriogenesis somatik pada kelapa sawit untuk perbanyakan secara in vitro klon unggul. Jurnal Penelitian Kelapa Sawit 4(1): 1-16.
Ginting, G., C. Mollers, dan K. Pamin. 1996. Embriogenesis somatik pada kelapa sawit untuk perbanyakan secara in vitro klon unggul. Jurnal Penelitian Kelapa Sawit 4(1): 1-16.
Kushairi, A., A.H. Tarmizi, I. Zamzuri, M. OngAbdullah, R. Samsul Kamal, S.E. Ooi, and N. Rajanaidu. 2010.
Production, performance, and advances in oil palm tissue culture. International Seminar on Advances in Oil Palm Tissue Culture.Yogyakarta: Indonesia.
Latif, S. 2004. Keragaan dan produktifitas klon kelapa sawit asal kultur jaringan di Sumatera bagian utara. Jurnal Penelitian Kelapa Sawit 12(1): 11-24.
Leng, T.C., N.S. Ping, B.P. Lim, and C.L. Keng. 2011. Detection of bioactive compounds from Spilanthes acmella (L.) plants and its various in vitro culture products. Journal of Medicinal Plants Reseacrh 5(3): 371-378.
M., W.A. Nchu, N. Godswill, N.M.A. Wiendi, A. Wachjar, and N.E.G. Frank. 2015. Induction of oil palm (Elaeis guineensis Jacq. var. Tenera) callogenesis and somatic embryogenesis from young leaf explants. Journal of Applied Biology &
Biotechnology. 3(04): 004-010.
Manchanda, P., A. Kaur, and S.S. Gosal. 2018. Somaclonal variation for sugarcane improvement. In : Biotechnologies of Crop Improvement. Editor : Gosal, S.S., and S.H. Wani.
Springer.
Mohd Din, A., A. Kushairi, N. Rajanaidu, A. Noh, and Z.A.
Issa. 2005. Performance of various oil palm introgressed population at MPOB. Proceedings of Agricultural, Biotechnology, and Sustainability Conference. 111-143.
Monteiro, T. R., E. O. Freitas, G.F. Nogueira, and J.E. Scherwinski- Pereira. 2017. Assessing the influence of subcultures and liquid medium during somatic embryogenesis and plant regeneration in oil palm (Elaeis guineensis Jacq.).The Journal of Horticultural Science and Biotechnology. 93: 196–203.
Naher, L., U.K. Yusuf, A. Ismail, S.G. Tan and M.M.A. Mondal.
2013. Ecological status of Gamoderma and basal stem rot disease of oil palm (Elaeis guineensis Jacq.). AJCS 7:1723- 1727.
Dr. Fathurrahman, SP., M.Sc
18
Oseni, O.M., V. Pande, and T.K. Nailwal. 2018. A review on plant tissue culture, a technique for propagation and conservation of endangered plant species. International Journal of Current Microbiology and Applied Sciences 7(7):
3778 -3786.
Owoyele, B., and G.O. Owolabi. 2014. Traditional oil palm (Elaeis guineensis Jacq.) and its medicinal uses : A review.
TANG Humanitas Medicine 4(3): 1-8.
Perez-Clemente, R.M., and Gómez-Cadenas. 2012. A In vitro Tissue Culture, a Tool for the Study and Breeding of Plants Subjected to Abiotic Stress Conditions.
Purba, A.R., dan T. Hutomo. 1993. Pemilihan ortet untuk perbanyakan vegetatif kelapa sawit secara kultur jaringan.
Metode indeks seleksi. Buletin PPKS 1(2): 119-130.
Sambanthamurthi, R., R. Singh, A.P.G. Kadir, M.O. Abdullah and A. Kushairi. 2008. Opportunities for the oil palm via breeding and biotechnology. In : Jain, S.M., and P.M.
Priyadarshan. Breeding Plantation Tree Crops: Tropical Species. Springer Science & Business Media.
Scherwinski-Pereira, J.E., R.S. da Guedes, P. Cesar, Jr. Fermino, T.L. Silva, and F.H.S. Costa. 2010. Somatic embryogenesis and plant regeneration in oil palm using the thin cell layer technique. In Vitro Cellular and Developmental Biology. 46:
378-385.
Setiowati, R.D., Ernayunita,A.F. Simamora, E. Nazri, Fakhrullah, T.C. Hidayat, dan I.Y. Harahap. 2011. Keragaan klon kelapa sawit PPKS di beberapa kebun komersil. Jurnal Penelitian Kelapa Sawit. 19(3): 101-108.
Setiowati, R.D., Ernayunita, H.Y. Rahmadi, dan Y. Yenni. 2013.
Klon kelapa sawit mengenal bahan tanaman kelapa sawit hasil kultur jaringan. Pusat Penelitian Kelapa Sawit, Medan.
Sharma, M. 2006. Challenges facing the Malaysian palm oil industry-multi pronged strategies for raising oil yield, productivity and profitability. In : Kushairi, A., R.
Sambanthamurthi, A.M. Ong, and C.K. Choong. Proc.
Clonal & Qty. Rep. Material. Malaysian Palm Oil Board.
Sianipar, N.F., G.A. Wattimena, H. Aswidinnoor, M.
Thenawidjaya, N. Toruan-Mathius, dan G. Ginting. 2007.
Karakterisasi secara morfologi abnormalitas embrio somatik kelapa sawit (Elaeis guineensis Jacq.) dari eksplan daun. Jurnal AgroBio en 3(1): 32-39.
Sogeke, A.K. 1988. Stages in the vegetative propagation of oil palm, Elaeis guineensis Jacq. Through tissue culture. Journal of Oil Palm Research. 10(2): 1-9.
Soh, A.C., S. Mayes, and J.A. Roberts. 2017. Oil Palm Breeding Genetics and Genomics. CRC Press.
Steinmacher, D.A., N.G. Krohn, A.C.M. Dantas, V.M. Stefenon, C.R. Clement, and M.P. Guerra. 2007. Somatic embryogenesis in peach palm using the thin cell layer technique: Induction, morphohistological aspects and AFLP analysis of somaclonal variation. Annals Botany. 49:
1-11.
Sumaryono, I. Riyadi, R.T. Saptari, H.Y. Rahmadi, and Ernayunita. 2018. Embryogenic callus initiation from leaf explants of Elaeis oleifera x Elaeis guineensis (OxG) hybrids.
IOP Conference Series: Earth and Environmental Science. 183 012009: 1-6.
Tan, C.C., G. Wong, A.C. Soh, T.Y. Hor, S.P. Chong, and K.
Gopal. 2003. Experiences and lessons from oil palm clonal evaluation trials and commercial test plantings. PIPOC International Palm Oil Congress. MPOB, Bangi: 1093-1119.
Van Harten, A.M. 1998. Mutation breeding. Theory and Practikal Applications. Cambridge University Press.
Vasil, I.K., and V. Vasil. 1975. Totipotency and embryogenesis in plant cell and tissue cultures. In Vitro 8(3): 117-127.
Dr. Fathurrahman, SP., M.Sc
20
Weckx, S., D. Inze, L. Maene. 2019. Tissue Culture of Oil Palm:
Finding the Balance Between Mass Propagation and Somaclonal Variation. Frontier in Plant Science. 10(722): 1- 17.
BAB II
KELAPA SAWIT DAN KEPENTINGANNYA
2.1 Kandungan Minyak Tumbuhan
Kelapa sawit (Elaeis guineensis Jacq) merupakan salah satu tanaman komoditas utama di Indonesia. Kelapa sawit (Elaeis guineensis Jacq) diklasifikasikan ke dalam famili palmae, ordo plamales dan genus elaeis. Tumbuhan ini tergolong dalam tumbuhan monokotil. Secara fisiologis, pohon kelapa sawit melakukan fotosintesis sepanjang tahun dan sebagian besar hasil fotosintesis diubah menjadi bahan penyimpanan (Somerville & Browse 1991).
Kelapa sawit memiliki umur panjang hingga puluhan tahun dan juga dikenal sebagai tanaman tahunan. Namun, usia produktif adalah antara 20 hingga 35 tahun. Berbuah saat berumur tiga setengah tahun. Tumbuhan ini memiliki nilai ekonomi yang penting terutama bagi negara-negara dengan iklim tropis seperti Indonesia, Malaysia, Papua New Guinea, Nigeria, Brazil dan Panama (Hardon et al. 1985). Elaeis guineensis Jacq. Dari benua Afrika dan ditanam secara komersial di Afrika.
22
Dr. Fathurrahman, SP., M.Sc
Elaeis oleifera dikenal sebagai Elaeis melanococca yang banyak diperoleh di Amerika Selatan merupakan salah satu spesies dalam genusnya yang berkerabat dekat dengan Elaeis guineensis.
Selain itu, ada spesies lain yang kurang dikenal dari Amazon, Elaeis odora, yang merupakan spesies lain dalam genus Elaeis (Hardon 1976). Namun E. oleifera kurang menguntungkan jika ditanam secara komersial karena rendahnya produksi minyak dengan karakteristik morfologi dan agronomis yang kurang menarik seperti ukuran buah yang kecil dan jumlah tandan buah yang sedikit. Meskipun demikian, Elaeis oleifera memiliki beberapa ciri khusus jika dibandingkan dengan spesies Elaeis lainnya, seperti memiliki kandungan asam lemak tak jenuh tunggal asam oleat yang tinggi (C:18,1) yaitu sekitar 60%, nilai yodium tinggi lebih dari 80, karakteristik pohonnya rendah dan tahan terhadap penyakit Fusarium (Porim Bulletin 1983;
Rao et al. 1989). Sejak tahun 1960-an, pengembangan kelapa sawit telah berpindah dari benua Afrika ke negara-negara Asia seperti Indonesia, Malaysia, Filipina, Thailand, dan Papua Nugini.
Penemuan gen pengatur ketebalan cangkang pada buah kelapa sawit pada tahun 1941 dan hibrida dura x pisifera mengakibatkan genotipe tenera menjadi bahan tanaman yang disukai karena karakteristiknya yang unggul (Zakri 1993).
Varietas dura merupakan induk betina yang memiliki mesocarp tipis, cangkang dan kernel tebal (2 – 8 mm) yang dikendalikan oleh sepasang alel (Sh+ Sh+). Pisifera ditentukan oleh alel homozigot resesif (Sh-Sh-) yang digunakan sebagai induk jantan yang memiliki mesokarp lebih tebal, cangkang tipis atau bahkan tanpa biji sama sekali. Tenera merupakan bentuk peralihan antara dura dan pisifera yang dikendalikan oleh alel heterozigot (Sh+Sh-) yang menghasilkan mesocarp tebal, cangkang tipis, dan kernel cukup tebal (Opeke 1982).
Varietas tenera memberikan rendemen minyak yang lebih tinggi dibandingkan dengan varietas dura yaitu 20 - 26%.
2.2 Manfaat Minyak Nabati
Tingkat produksi minyak sawit lebih tinggi jika dibandingkan dengan minyak nabati lainnya, yaitu menghasilkan 4 - 7 ton/hektar per tahun. Ia mampu menghasilkan minyak tiga kali lebih banyak dari kelapa, dan 10 kali lebih banyak dari kedelai (Rajanaidu dan Jalani 1995). Ada dua jenis minyak yang dihasilkan buah sawit, yaitu minyak mesokarpa dan minyak kernel (Opeke 1982). Kandungan minyak tertinggi terdapat pada mesokarpa yang mencapai 40- 60% dari berat basah buah. Bagian mesokarpa mengandung 44% asam palmitat jenuh, 39% oleat dan 10% asam linoleat.
Kandungan asam oleat memiliki berbagai aplikasi dalam produk industri (Bhat 1989). Bagian kernel mengandung asam laurat yang tinggi yaitu sekitar 50% dan asam meristik 16%.
Minyak kelapa sawit tidak mengandung asam linolenat, melainkan mengandung 500 ppm tokoferol yang merupakan antioksidan alami sehingga lebih cocok digunakan sebagai minyak goreng (Berger & Ong 1985). Tabel 2.1 menunjukkan komposisi asam lemak dari tumbuhan penghasil minyak dan lemak utama.
Kandungan utama minyak nabati adalah triasilgliserol yang mengandung asam lemak jenuh dan asam lemak tak jenuh yang memiliki rantai karbon mulai dari rantai ke-12 sampai ke- 22 yang diesterifikasi menjadi gliserol. Minyak kedelai, bunga matahari, dan biji kapas masing-masing mengandung 55%, 79%, dan 54% asam linoleat, sedangkan minyak sawit, rapeseed, dan minyak kacang tanah mengandung 41%, 60%, dan 50%
asam oleat (asam lemak tak jenuh tunggal). Secara luas, asam lemak tanaman biji lobak yang telah diesterifikasi digunakan
Dr. Fathurrahman, SP., M.Sc
24
sebagai bahan bakar mesin oleh beberapa negara (Hans &
Heldt 1997). Kandungan asam linolenat yang berasal dari biji rami di negara-negara Eropa digunakan untuk memproduksi cat dan pernis.
Minyak dari mesocarp dan kernel kelapa sawit berbeda dalam komposisi dan kegunaannya. Minyak mesokarp sering digunakan untuk industri minyak goreng, margarin, detergen, sabun, makanan berlemak dan es krim, sedangkan minyak inti sawit lebih sering digunakan sebagai bahan pembuatan sabun, shampo, margarin, metil ester, minyak goreng dan makanan berlemak. Bagian mesokarp buah lontar kaya akan asam palmitat yang menyebabkannya membentuk lemak semi padat yang stabil pada suhu kamar. Karena itu bagian ini banyak digunakan di sektor industri.
Penggunaan minyak goreng yang mengandung asam lemak jenuh tingkat tinggi tidak baik untuk kesehatan manusia.
Meskipun demikian, penelitian terbaru menunjukkan bahwa penggunaan minyak sawit bermanfaat bagi kesehatan manusia (Sundram & Koh 1999). Metabolisme tubuh manusia membutuhkanasamlemaktakjenuhdenganduaataulebihunsur
karbon ikatan rangkap, tetapi tidak dapat menggabungkan diri pada ikatan rangkap setelah rantai C-9. Asam linoleat dan asam linolenat merupakan asam lemak yang sangat penting bagi kesehatan manusia. Minyak inti sawit yang kaya asam lemak rantai sedang (C:12 – C:14) merupakan sumber penting dalam industri nutrisi, deterjen dan kosmetik (Luhs & Friedt 1994b). Baru-baru ini, minyak inti sawit menjadi lebih populer daripada minyak kelapa karena kandungan asam jenuhnya (C:12.0) lebih tinggi. Sebagian besar minyak nabati digunakan untuk produksi produk makanan, namun juga digunakan dalam industri. Asam erusat (C22:1) merupakan komponen yang berasal dari biji Brassica napus, dapat diubah menjadi asam erusida dan ini dapat digunakan sebagai agen slip dalam pembuatan film plastik (Ohlrogge 1994). Selain itu, dimungkinkan untuk digunakan sebagai prekursor dalam pembuatan nilon 13,13, yang merupakan jenis termoplastik suhu tinggi. Biaya pemurnian asam erusat dari Brassica napus terlalu tinggi. Namun hal ini dapat diatasi dengan menaikkan kadar asam menjadi 50 sampai 90%, sehingga biaya produksi asam erusat dari minyak tumbuhan ini akan jauh lebih murah dan berpotensi untuk menggantikan minyak bumi sebagai prekursor pembuatan nilon 13, 13 (Ohlrogge 1994).
Tabel 2.2 secara singkat menunjukkan penggunaan utama minyak nabati untuk nutrisi dan oleokimia. Metil ester dari minyak nabati telah digunakan untuk mesin. Sejak saat itu, demonstrasi penggunaan minyak dan lemak semakin berkembang. Malaysia telah berhasil memproduksi minyak sawit yang dapat dikonversi menjadi metil ester atau solar minyak sawit (POD). Oli ini telah diuji pada mesin diesel dan menunjukkan hasil yang memuaskan (Masjuki dan Sapuan 1995). Selain itu, metil ester dari minyak nabati berpotensi sebagai alternatif pengganti minyak solar karena lebih ekonomis dan lebih aman. Minyak biji rep dengan kandungan
15 atau 18 asam lemak karbon dari asetil-KoA dan malonil- KoA. Umumnya, kompleks enzim asetil-KoA karboksilase (ACCase) mengkatalisis reaksi jalur biosintesis asam lemak pertama, yaitu pembentukan malonil-KoA dari asetil-KoA dan CO2. Gugus malonil selanjutnya ditransfer dari koA ke protein pembawa asil (ACP), ACP ini sebagai kofaktor protein sebelum melewati lokasi jalur biosintesis. Protein pembawa asil (ACPs) mengandung gugus prostetik phosphopantetheine pada rantai asil yang mengalami pemanjangan untuk dilekatkan sebagai thioester. Setelah dipindahkan ke ACP, malonil-tioester akan memasuki serangkaian reaksi kondensasi dengan asil-ACP atau asetil-KoA sebagai akseptor. Reaksi ini menyebabkan pembentukan ikatan karbon dan pelepasan CO2. Dalam produksi asam lemak 18-karbon, tiga enzim terpisah berfungsi untuk mengkatalisasi reaksi kondensasi.
Jalur asam lemak de novo diprakarsai oleh reaksi kondensasi antara asetil-KoA danmalonil-ACP. Reaksi pertama, kondensasi asetil-KoA dan malonat-ACP akan membentuk asetoasetil-ACP. Reaksi ini membutuhkan enzim -ketoasil ACP sintase III. Reaksi kedua, kondensasi berikutnya, akan memperpanjang rantai asam lemak dengan menambahkan dua karbon dari setiap reaksi kondensasi, menghasilkan rantai asil 16-karbon. Enzim β-ketoacyl-ACP synthase I mengkatalisis reaksi ini dan malonyl-ACP digunakan sebagai sumber dua karbon. Reaksi kondensasi ketiga, pemanjangan rantai asam lemak C:16 menjadi C:18 dikatalisis oleh isoform asam lemak sintase yaitu -ketoasil-ACP sintase II. Pada setiap siklus reaksi kondensasi di atas, prosesnya melibatkan 4 tahap, yaitu dekarboksilasi untuk menghilangkan satu molekul karbon dioksida, reduksi, dehidrasi dan reduksi lagi untuk menghasilkan rantai asil-ACP yang akan digunakan pada siklus kondensasi berikutnya. Setelah rantai asam lemak C:18 yang
Dr. Fathurrahman, SP., M.Sc
28
dihasilkan akan didesaturasi menjadi oleyl-ACP (C:18.1) oleh enzim ∆9-stearoyl-ACP desaturase (Jones and Cassells 1995).
Gambar 2.1 Skema sederhana jalur sintesis asam lemak pada plastida tumbuhan. Angka yang tercantum mewakili enzim: 1, asetil-KoA karboksilase; 2, transilase ACP malonil-CoA;
3, β-ketoasil-ACP sintase III; 4, β-ketoasil-ACP sintase I ; 5, β-ketoasil-ACP sintase II ; 6 stearoyl-ACP desaturase; 7, tioesterase oleoyl-ACP; 8, asil-ACP thioesterase ; 9 asetil- KoA sintase ; 10 asil-CoA sintase. Sumber: dimodifikasi dari Ohlrogge (1994).
Produksi tioester asam oleat melalui jalur di atas akan terhidrolisis dan dikeluarkan dari plastida. Enzim oleoyl-ACP thioesterase berperan dalam reaksi ini. Asam lemak bebas dapat
diubah dari tioester asam lemak lainnya, khususnya rantai yang lebih panjang dari C:10. Hal ini menyebabkan asil ACP thioesterase biasanya memiliki lebih dari satu isoform pada suatu tumbuhan. Klon cDNA yang mengkode tioester asil-ACP dari biji safflower telah berhasil diisolasi dan dikarakterisasi (Knutzon et al. 1992a). Isolasi tioesterase asil-ACP spesifik untuk rantai asil panjang sedang telah berhasil dilakukan dari tanaman dengan komposisi asam lemak panjang sedang tinggi.
Sebagai contoh, klon acyl-ACP thioesterase cDNA spesifik untuk lauryl-ACP diisolasi dari biji teluk California, spesies yang mengakumulasi kaprat (C:10.0) dan laurat (C:12.0) tingkat tinggi selama pengembangan benih (Voelker et al. 1992 ).
Asam lemak yang ditemukan dalam lipid penyimpanan sangat bervariasi dan memiliki rantai karbon antara C:18 sampai C:24, sedangkan asam lemak yang ditemukan pada membran tumbuhan memiliki rantai karbon antara C:16 sampai C:18 saja.
Pada beberapa jenis sel tertentu, ada yang membutuhkan asam lemak lebih tinggi, antara lain kacang tanah, bunga matahari, rapeseed, plax, dan tanaman wijen. Kandungan minyaknya 45% dari berat biji (Somerville & Browse 1991). Sedangkan untuk tanaman lain memiliki karakteristik yang berbeda seperti pada tanaman biji-bijian yang kaya akan kandungan protein terutama yang berasal dari tanaman polong-polongan, kandungan lemaknya hanya mencapai 1-2% dari berat biji (Murphy 1993b).
Asam lemak nabati merupakan sumber energi yang penting karena dapat dipecah untuk mendapatkan energi efektif dua kali lebih banyak daripada glukosa. Pada tumbuhan, sintesis dan pengaturan asam lemak lebih kompleks daripada organisme lain. Asam lemak nabati diproduksi di plastida dan bukan di sitosol seperti pada organisme lain. Namun, fungsi asam lemak yang paling penting adalah untuk pembentukan
Dr. Fathurrahman, SP., M.Sc
30
membran sel (Albert et al. 1994). Sintesis asam lemak juga dapat memenuhi kebutuhan biologis seperti fiksasi struktural dan penyimpanan karbon. Seluruh sel ditutupi oleh membran sel hidrofobik. Membran sel juga melindungi organel didalamnya yang sebagian besar tersusun atas fosfolipid, yaitu molekul kecil yang menyerupai trigliserida karena sebagian besar terdiri dari asam lemak dan gliserol.
Sebagian besar enzim yang terlibat dalam jalur di atas telah diisolasi dalam bentuk klon cDNA yang diperoleh dari beberapa spesies. Klon merupakan sumber minyak dan tanaman yang sangat penting yang mengandung asam lemak spesifik. Gen parsial untuk enzim β-ketoacyl-ACP synthase II berhasil diisolasi dari E. guineensis, tenera (Cha 2001).
2.3.2 Pentingnya Enzim ∆-9-stearoyl-ACP desaturase
Kehadiran enzim ∆9-stearoyl-ACP desaturase dalam jaringan fotosintetik pertama kali dilaporkan oleh Nagai dan Bloch (1968), diikuti dengan isolasi biji safflower (Mc Keon
& Stumpf (1982).Klon cDNA untuk enzim ini telah berhasil diperoleh dari R communis, R. cucumis dan Carthamus tinctorius dengan menggunakan antibodi sebagai probe. Klon cDNA dari tanaman lain telah diisolasi, yaitu Oryza sativa (Akagi et al.
1995), Brassica napus (Slocombe et al. 1994), Spinacia oleracaea (Nishida et al. 1992), Linus usitatissimum (Singh et al. 1994), Brassica rapa (Knutzon et al. 1992a) dan Elaeis guineensis Jacq.
(Rashid 1997). Protein dari desaturase ∆-9stearoyl-ACP aktif juga telah diperoleh dari ekspresi klon cDNA safflower.
Dalam biosintesis asam lemak, asam oleat (C:18.1) merupakan zat metabolik utama yang berdasarkan posisinya dalam biosintesis. Asam ini merupakan produk akhir dari reaksi sintetase asam lemak yang terjadi di dalam plastid dan merupakan substrat awal untuk reaksi modifikasi selanjutnya
yang terjadi baik di dalam maupun di luar plastid. Asam oleat bebas dihasilkan dari reaksi sekuensial enzim ∆-9 stearoyl-ACP desaturase dan enzim oleyl-ACP thioesterase. Gen dalam regulasi antisense desaturase stearoyl-ACP telah diperkenalkan pada tanaman transgenik Brassica, Nicotiana dan Elaeis guineensis Jacq (Cahoon et al. 2010; Knutzon et al. 1992a; Shahabuddin 2000).
Penemuan ini menyebabkan penurunan rasio antara asam lemak jenuh dan tak jenuh. Hal ini membuktikan bahwa enzim ini berperan sebagai enzim utama dalam menentukan keseimbangan antara asam lemak jenuh dan tak jenuh bagi tumbuhan tingkat tinggi. Penggunaan teknologi rekayasa genetika pada ekspresi enzim stearoyl-ACP desaturase diyakini mampu mengubah komposisi asam lemak tanaman (Akagi et al. 1995).
Di sebagian besar jaringan tanaman, lebih dari 75% asam lemak adalah asam lemak tak jenuh. Stearoyl-ACP desaturase yang larut dalam enzim memperkenalkan ikatan rangkap pertama dengan stearoyl-ACP (C:18.0) antara karbon 9 dan karbon 10 untuk membentuk asil oleoyl-ACP tak jenuh tunggal (C:18.1). Enzim desaturase stearoyl-ACP unik karena enzim desaturase lainnya adalah protein yang terikat membran, sementara mereka ada secara bebas di sitosol. Selain itu, enzim ini menunjukkan spesifisitas yang lebih tinggi terhadap stearoil- ACP dibandingkan dengan palmitoyl-ACP sejalan dengan kandungan palmitat dan oleat yang lebih tinggi daripada stearat dalam komposisi asam lemak tumbuhan (Harwood 1988).
Penyaringan koleksi cDNA minyak sawit telah berhasil mengidentifikasi tiga klon putatif yang mengkode enzim desaturase ∆9-stearoyl-ACP minyak sawit, yaitu pTD7, pTD8, pTD9 dan analisis urutan lengkap yang diperoleh menunjukkan bahwa minyak sawit ∆9-stearoyl-ACP gen desaturase berukuran
Dr. Fathurrahman, SP., M.Sc
32
1709 bp. Perbandingan urutan gen desaturase ∆-9 stearoyl-ACP minyak sawit menunjukkan tingkat homologi yang tinggi berkisar antara 70 hingga 78% pada level DNA dan 75 hingga 84% pada level asam amino, dengan ∆9-stearoyl-ACP desaturase urutan klon cDNA dari beberapa tumbuhan lain seperti R.
communis, T. alata, Brassica spp dan C. tinctorius (Rasyid 1997).
Sekuensing DNA dari klon cDNA beberapa spesies tanaman telah diperoleh. Setelah dilakukan perbandingan pada urutan DNA dan homologi antara 78 dan 84% dan ini menunjukkan bahwa urutan enzim ini diabadikan pada tumbuhan. Berat molekul protein desaturase ∆9-stearoyl-ACP dewasa (polipeptida) adalah sekitar 40 Kda, termasuk peptida transit yang biasanya antara 30 dan 35 asam amino. Fungsi polipeptida transit adalah untuk memastikan lokasi target enzim dalam plastida.
2.3.3 Enzim asil-ACP tioesterase
Enzim ini berperan penting dalam penghentian pemanjangan rantai asam lemak. Perannya adalah melepaskan asam lemak dari ACP sehingga dapat dikeluarkan dari plastid ke sitoplasma dan akan bergabung menjadi trigliserida.
Kandungan asam palmitat dan asam oleat yang tinggi pada minyak sawit disebabkan oleh enzim asil-ACP thioesterase dengan spesifisitas yang tinggi terhadap palmitoyl-ACP dan oleoyl-ACP pada mesokarp sawit menunjukkan kecenderungan menuju palmitoyl-ACP dan selanjutnya menjadi oleoyl-ACP.
substrat. Penjelasan selanjutnya adalah bahwa asam palmitat dilepaskan dari palmitoyl-ACP dalam pemanjangan rantai asam lemak. Selanjutnya, asam palmitat bergerak keluar dari plastid dan modifikasi gugus ester menjadi trigliserida melalui reaksi asil transferase di sitoplasma. Mekanisme ini menyebabkan peningkatan kadar asam palmitat dan trigliserida. Respon yang
tinggi terhadap oleoyl-ACP mempengaruhi tingkat rendemen kandungan asam oleoyl-ACP pada mesokarpa sawit. Rekayasa genetika dapat mengubah komposisi asam lemak yang berbeda dan meningkatkan kandungan oleat dengan mengurangi asam palmitat.
Acyl-ACP thioesterase telah berhasil dimurnikan dari jaringan biji Brassica napus. Selain itu, klon cDNA parsial thioesterase palmitoyl-ACP juga telah berhasil diisolasi dari E. guineensis tenera dengan metode RT-PCR (Cha 2001). Dari pengamatan heterogenitas yang konsisten, ditemukan bahwa Brassica napus mengandung gen thioesterase yang dikodekan oleh keluarga multigen kecil. Gen thioesterase juga berhasil dimurnikan dari safflower dan rapeseed (Loader et al. 1993).
2.3.4 Triasilgliserol Sintetik
Produksi rantai asam lemak dari reaksi sintetase asam lemak akan digunakan dalam sel tumbuhan melalui dua jalur yang berbeda (Ohlrogge & Browse 1995; Murphy 1993a;
Somerville & Browse 1991). Untuk sintesis prokariotik, rantai asil-ACP digunakan untuk menghasilkan lipid plastid dan tidak diubah menjadi asam lemak bebas. Langkah selanjutnya adalah enzim asiltransferase yang mengubah rantai asil-ACP menjadi asam fosfatidat. Asam fosfatidat kemudian akan membentuk fosfatidil gliserol (PG) dan diasilgliserol (DAG).
DAG digunakan untuk menghasilkan galaktolipid yang merupakan komponen utama membran plastid. Untuk jalur eukariotik, rantai asil ACP pertama kali dihidrolisis menjadi asam lemak bebas oleh beberapa isoform enzim thioesterase seperti yang disebutkan di atas. Kemudian asam keluar melalui plastida untuk dimodifikasi pada membran retikulum endoplasma. Triacylglycerol (TAG) adalah penyimpanan lipid untuk sebagian besar tanaman.
Dr. Fathurrahman, SP., M.Sc
34
Roadmap Kennedy atau roadmap gliserol-3 fosfat adalah jalur akumulasi TAG (Ohlrogge & Browse 1995). Jalur reaksi produksi rantai asam lemak yang dilepaskan dari plastid untuk dipindahkan ke gliserol-3 fosfat melalui dua cara. Cara pertama melibatkan produksi asam lisofosfatidat dan cara kedua menghasilkan asam fosfatidat. Pemindahan rantai asam lemak dari plastida ke gliserol-3 fosfat melibatkan enzim asil transferase. Dari asam fosfatidat kemudian diubah menjadi diasilgliserol (DAG) oleh enzim fosfatase fosfatidat. Hasil akhir dari reaksi ini adalah DAG akan menghasilkan TAG. Reaksi ini menyebabkan akumulasi TAG berupa bintik-bintik minyak di antara lapisan membran ganda lemak retikulum endoplasma.
Proses selanjutnya adalah perluasan membran retikulum endoplasma. Selanjutnya bercak minyak tersebut akan dikeluarkan ke dalam sitoplasma. Bintik-bintik minyak kecil akan bergabung membentuk massa minyak yang lebih besar di sitoplasma sel.
Gambar 2.2 menunjukkan diagram skematik sintesis triasilgliserol tanaman. Di sekitar massa minyak akan terbentuk lapisan protein khusus yang disebut oleosin.
Gambar 2.2. Jalur sintesis triasilgliserol pada sel tumbuhan.
Protein ini berfungsi untuk melindungi benih selama proses dehidrasi dan melindungi sel tanaman dari komponen TAG yang berpotensi beracun dan deterjen karena TAG dengan rantai asam lemak sedang-panjang biasanya memiliki sifat deterjen (Murphy 1993b; Murphy 1994). Jalur gliserol-3 fosfat melalui mana oleyl-CoA pertama kali dimodifikasi. Proses ini akan menghasilkan beberapa turunan asam lemak yang biasa terdapat pada penyimpanan triasilgliserol, misalnya asam erusat, asam linoleat, asam linolenat dan asam risinoleat.
Produksi asam lemak tak jenuh ganda dikatalisis oleh dua enzim desaturase, yaitu 6-desaturase yang mengkatalisis reaksi untuk menghasilkan asam linoleat dan β-desaturase yang mengkatalisis reaksi untuk menghasilkan asam linolenat.
sekunder, keton, aldehida dan ester asil yang biasanya terdapat pada lilin (Post Beittenmiller 1996). Akumulasi VLCFA juga terjadi pada minyak biji serta triasilgliserol (TAG).
FAE dalam siklus panjangnya melibatkan empat reaksi yaitu pertama, kondensasi C:18.1-CoA dengan malonyl-CoA membentuk β-Ketoacyl-CoA; kedua, reduksi β-ketoacyl-CoA menjadi β-hydrocysyl-CoA ; ketiga, dehidrasi β-hydrocysyl-CoA menjadi trans 2,3-enoyl-CoA; keempat, pengurangan kembali trans-2,3-enoyl-CoA untuk menghasilkan asil CoA yang telah diperpanjang oleh dua karbon (Fehling & Mukherjee 1991).
Mekanisme FAE berbeda dari FAS di mana substrat FAE menggunakan malonil KoA dan asil KoA, sedangkan FAS menggunakan kofaktor ACP untuk siklus pemanjangannya.
Kandungan VLCFA pada biji arabidopsis mencapai hampir 28% yang sebagian besar terdiri dari asam esikosenoat (C:20.1).
Hal ini dibuktikan dengan isolasi gen pemanjangan asam lemak I (FAE1) dan cDNA keton yang mengkode enzim kondensasi β- ketoacyl-CoA synthase (KAS) dari Arabidopsis (Kunst et al. 1992;
James et al. 1995). Gen FAE1 mampu merangsang produksi VLCFA di jaringan dan merupakan enzim pembatas laju untuk sistem FAE. Untuk enzim reduktase dan dehidrase FAE ada di mana-mana dan diekspresikan dalam tanaman dan ragi.
Ditemukannya gen arabidopsis cut1 yang mengkode enzim kondensasi sistem FAE yang diperlukan untuk menghasilkan lilin kutikula (Millar et al. 1999). Spesifisitas gen ini belum teridentifikasi, tetapi diperkirakan bahwa gen cut1 berperan dalam pemanjangan VLCFA C:24. Peran lilin kutikula adalah untuk melindungi tanaman dari serangan bakteri dan patogen tunggal, sinar UV dan pengendalian kehilangan air. Sebaliknya bila kandungan VLCFA terlalu tinggi pada membran tanaman akan menyebabkan perubahan yang buruk pada sifat morfologi tanaman (Millar et al. 1998).
Dr. Fathurrahman, SP., M.Sc
38
2.4 Kultur Jaringan Tanaman Kelapa Sawit
2.4.1 Regenerasi Embrio Belum Matang (EBM) menjadi planlet
Teknik kultur jaringan embrio belum matang muerupakan salah satu cara digunakan sebagai sumber jaringan yang diwariskan ke beberapa jenis tumbuhan lain. Cutter dan Wilson (1954) mengkulturkan embrio dari buah kelapa secara in vitro pada berbagai media kultur jaringan, hingga embrio tersebut menjadi embriogenik, perkembangan tunas dan akar terlihat sangat lambat. Selain itu, embrio belum matang juga banyak digunakan dalam kultur jaringan dari gandum dan tanaman biji-bijian lainnya (Green & Phillips 1975; Shilito et al. 1989;
Prioli & Sondahl 1989). Pembentukan kalus dari EMB yang menunjukkan pertumbuhan lambat telah dilaporkan (Smith &
Thomas 1973). Embrio kelapa sawit yang belum matang juga telah digunakan dalam kultur jaringan untuk regenerasi kalus dan kalus embriogenik (Texeira et al. 1993).
Pada kultur jaringan secara in vitro, media Y3 berhasil mempengaruhi berat dan ukuran embrio kelapa dengan hasil yang lebih tinggi dibandingkan media MS dan White (Rillo dan Paloma 1990). Kultur jaringan kelapa sawit memiliki produksi kalus yang relatif lambat dan persentase produksi embriogenik dibandingkan dengan kultur jaringan jenis padi. Hal ini dikarenakan kelapa sawit termasuk tanaman tingkat tinggi yang proses fisiologisnya lebih kompleks. Kultur jaringan kelapa sawit menggunakan varietas yang berbeda seperti tenera, dura dan pisifera telah dilakukan. Sumber jaringan yang digunakan adalah jaringan ortotik untuk regenerasi eksplan dan produksi embriogenik. Setelah 2 tahun dikulturkan dalam ruang kultur, diketahui bahwa varietas tenera menghasilkan kalus embriogenik yang lebih tinggi yaitu 55,1% dibandingkan dura dan pisifera masing-masing 25,0% dan 23,1% (Wooi
1990).
Akhir-akhir ini, embrio yang belum matang (EBM) telah digunakan sebagai salah satu sistem model yang digunakan untuk berbagai transformasi genetik termasuk melalui transfer gen langsung, media Agrobacterium dan teknik transfer gen lainnya.Seringkalidalamprosestransformasigenetik,bagiandari EBM yang digunakan sebagai jaringan target akan mengalami cedera, sehingga jaringan tersebut mengalami kesulitan untuk tumbuh dan berkembang secara normal, misalnya kerusakan jaringan akibat proses biolistrik atau jaringan yang terinfeksi oleh Agrobacterium-mediated. transformasi. Selain itu, bisa juga terjadi gangguan pertumbuhan yang disebabkan oleh faktor genotip dan fenotipik. Faktor genotipik seperti keadaan fisiologis jaringan yang bersangkutan, yang menentukan bagus atau tidaknya jaringan tersebut dalam pertumbuhan dan perkembangannya. Faktor genotipe sangat penting dalam menentukan kemampuan suatu spesies untuk beregenerasi secara in vitro (Liu et al. 1990).
Sedangkan faktor fenotipe disebabkan oleh ketidakcocokan media kultur jaringan dengan berbagai komponen kimia (unsur makro, unsur mikro, vitamin, hormon pertumbuhan, antioksidan, asam amino dan lain-lain) untuk regenerasi (Ochoa-Aleja & Ireta 1990). Regenerasi dipengaruhi oleh peningkatan perkembangan sumber daya tanaman seperti kultur embrio, kalus, kalus embriogenik, eksplan dan planlet (Morrison et al. 1986), lokasi eksplan (Fari & Zako 1981) dan faktor lingkungan seperti suhu ruang kultur dan kekuatan cahaya. (Philips & Hubstenberger 1985). Kelapa sawit tidak dapat menghasilkan pucuk primer dan mikropropagasi tetapi tanaman ini menghasilkan embriogenesis somatik dari kultur daun atau akar muda (Jones 1984 ; Pannetier et al. 1981). Embrio dan planlet somatik telah berhasil diproduksi menggunakan