ABSTRAK
ISOLASI DAN KARAKTERISASI ENZIM PROTEASE DARI Actinomycetes ISOLAT LUMPUR HUTAN BAKAU
Oleh
Riki Eriyanti Andini
Pada penelitian ini, enzim protease diisolasi dari actinomycetes isolat lumpur hutan bakau. Produksi enzim ini dilakukan dalam media fermentasi Mineral Salt Medium yang mengandung skim milk sebanyak 2% selama 120 jam. Sentrifugasi pada suhu 4ºC dengan laju kecepatan 3500 rpm selama 30 menit menghasilkan ekstrak kasar enzim. Ekstrak kasar enzim dimurnikan dengan pengendapan terfraksi menggunakan garam ammonium sulfat dan didialisis dengan membran semipermeabel. Aktivitas protease di uji dengan menggunakan metode Kunitz termodifikasi dan kadar protein dengan metode Lowry.
Proses pemurnian meningkatkan aktivitas unit enzim sebesar 1,002 U/mL, kadar protein 0,723 mg/mL, aktivitas spesifik 1,386 U/mg pada ekstrak kasar enzim menjadi 8,78 U/mL, kadar protein 1,04 mg/mL, aktivitas spesifik 8,44 U/mg pada fraksi ammonium sulfat dengan tingkat kemurnian 6,08 kali.
II. TINJAUAN PUSTAKA
A. Hutan Bakau
Lautan Indonesia adalah bagian dari wilayah Indopasifik yang merupakan salah satu pusat keanekaragaman biota laut terbesar di dunia. Sumber biota laut tersebut merupakan aset potensial yang dapat didayagunakan menjadi aneka produk termasuk diantaranya farmasi. Jenis biota laut di daerah tropis Indonesia diperkirakan 2-3 kali lebih besar dibandingkan dengan biota laut di daerah subtropis dan daerah beriklim dingin (Van Soest, 1994).
Hutan bakau atau disebut juga hutan mangrove adalah hutan yang tumbuh di atas rawa-rawa berair payau yang terletak pada garis pantai dan dipengaruhi oleh pasang-surut air laut. Hutan ini tumbuh khususnya di tempat-tempat di mana ter-jadi pelumpuran dan akumulasi bahan organik. Baik di teluk-teluk yang terlin-dung dari gempuran ombak, maupun di sekitar muara sungai di mana air melam-bat dan mengendapkan lumpur yang dibawanya dari hulu (Irwanto, 2007).
nyakan bersifat khas hutan bakau karena telah melewati proses adaptasi dan evolusi. Hutan-hutan bakau menyebar luas di bagian yang cukup panas di dunia, terutama di sekeliling khatulistiwa di wilayah tropika dan sedikit di subtropika. Luas hutan bakau Indonesia antara 2,5 hingga 4,5 juta hektar, merupakan mangrove yang terluas di dunia melebihi Brazil (1,3 juta ha), Nigeria (1,1 juta ha) dan Australia (0,97 ha) (Spalding at al., 1997 dalam Noor at al., 1999).
B. Mikroorganisme
Mikroorganisme merupakan jasad hidup yang mempunyai ukuran sangat kecil (Kusnadi dkk., 2003). Setiap sel tunggal mikroorganisme memiliki kemampuan untuk melangsungkan aktivitas kehidupan antara lain dapat mengalami pertumbuhan, menghasilkan energi dan bereproduksi dengan sendirinya. Mikroorganisme memiliki fleksibilitas metabolisme yang tinggi karena mikroorganisme ini harus mempunyai kemampuan menyesuaikan diri yang besar sehingga apabila ada interaksi yang tinggi dengan lingkungan menyebabkan terjadinya konversi zat yang tinggi pula. Mikroorganisme ini juga tidak memerlukan tempat yang besar, mudah ditumbuhkan dalam media buatan, dan tingkat pembiakannya relatif cepat (Darkuni, 2001). Oleh karena aktivitasnya tersebut, maka setiap mikroorganisme memiliki peranan dalam kehidupan, baik yang merugikan maupun yang menguntungkan.
1. Sel
Reaksi-reaksi biokimia pada sel hidup berlangsung dalam ukuran yang sangat kecil dari sel hidup atau bagiannya, dengan dinding yang rapuh dan tebalnya hanya beberapa molekul (Michael et al., 2008). Terdapat dua kelas utama sel, yaitu sel prokariot dan sel eukariot (Albert et al., 1994). Sel prokariot memiliki struktur yang sederhana, pertumbuhan sel yang mudah dan cepat, serta mekanisme yang relatif sederhana dalam proses reproduksi. Sel prokariot bereproduksi dengan cara aseksual yang sangat sederhana. Organisme ini tumbuh hingga ukurannya berlipat ganda, lalu membelah diri menjadi sel anak yang identik (Lehninger, 1988). Dalam keadaan optimum, sel prokariot tunggal dapat membelah diri setiap 20 menit dan karena itu dapat mencapai jumlah 5 miliar sel dalam waktu kurang dari 11 jam(Albert et al., 1994). Sel eukariot merupakan divisi terbesar dari organisme yang hidup, yaitu seluruh organisme yang ada di bumi, termasuk hewan, tumbuhan, fungi, dan protozoa. Pada sel eukariot nukleus dan sitoplasma terdapat sebagai dua kompartemen yang terpisah oleh membran inti (Wolfe, 1993).
2. Bakteri
biasanya hanya berukuran 0,5-5 μm, meski ada jenis yang dapat menjangkau 0,3 mm dalam diameter (Thiomargarita). Bakteri umumnya memiliki dinding sel, seperti sel hewan dan jamur, tetapi dengan komposisi sangat berbeda (peptido-glikan). Bakteri berukuran sangat kecil dan dapat berkembang biak dengan cepat, cukup dengan cara membelah diri. Kemampuan bakteri untuk membelah dengan cepat memungkinkan populasi bakteri dapat segera menyesuaikan diri terhadap perubahan-perubahan dalam lingkungannya. Banyak bakteri yang bergerak menggunakan flagela, yang berbeda dalam strukturnya dari flagela kelompok lain. (Albert et. al., 1994).
Bakteri merupakan prokariot, dan berdasarkan bentuknya, bakteri dibagi menjadi tiga golongan besar, yaitu:
a. Kokus (Coccus) dalah bakteri yang berbentuk bulat seperti bola, dan mem-punyai beberapa variasi sebagai berikut:
o Mikrococcus, jika kecil dan tunggal o Diplococcus, jka bergandanya dua-dua
o Tetracoccus, jika bergandengan empat dan membentuk bujur sangkar o Sarcina, jika bergerombol membentuk kubus
o Staphylococcus, jika bergerombol
o Streptococcus, jika bergandengan membentuk rantai
b. Basil (Bacillus) adalah kelompok bakteri yang berbentuk batang atau silinder, dan mempunyai variasi sebagai berikut:
o Streptobacillus, jika bergandengan membentuk rantai
c. Spiril (Spirilum) adalah bakteri yang berbentuk lengkung dan mempunyai variasi sebagai berikut:
o Vibrio (bentuk koma), jika lengkung kurang dari setengah lingkaran o Spiral, jika lengkung lebih dari setengah lingkaran
Gambar 1. Bentuk-bentuk sel bakteri (Davidson, 2007)
Adaptasi bakteri pada lingkungan termal memiliki peranan yang sangat penting dalam penelitian dan pembelajaran teori adaptasi dalam biologi evolusioner (Crill et al, 1996). Temperatur merupakan kunci variabel lingkungan, karena temperatur dapat mempengaruhi pengaturan hampir seluruh laju proses fisiologis dan oleh karena itu mempengaruhi faktor-faktor biologis seperti pertumbuhan dan reproduksi (Bronikowski, 2001).
C. Actinomycetes
Actinomycetes adalah istilah yang digunakan untuk menunjukkan organisme jenis actinomycetales, subdivison utama dari Prokariot, yang terdiri dari semua organisme dengan sel prokariotik. Berbeda dengan organisme yang lain (Protista, Fungi, Plantae dan Animalia) yang semua memiliki sel eukariotik.
Actinomycetes dibagi menjadi tiga yaitu:
a. Actinomyceteae terdiri dari dua genus: Actinomyces dan Nocardia. Beberapa spesies bersifat patogen. A. Israeli menyebabkan actinomycosis dalam manusia, A. bovis pada sapi.
b. Mycobacteriaceae memiliki satu genus, Mycobacterium, yang berisi beberapa spesies patogen menyebabkan penyakit-penyakit seperti kusta dan TBC. c. Streptomyceteae terdiri dari beberapa organisme yang ditemukan di dalam
Actinomycetes memiliki sifat antara bakteri dan fungi. Actinomycetes adalah bentuk fungi, seperti bakteri yang terbentuk panjang, benang filamen bercabang-cabang seperti yang terlihat seperti jaring laba-laba abu-abu. Actinomycetes adalah dekomposer utama dari bahan tanaman keras seperti kulit kayu, koran dan batang kayu.
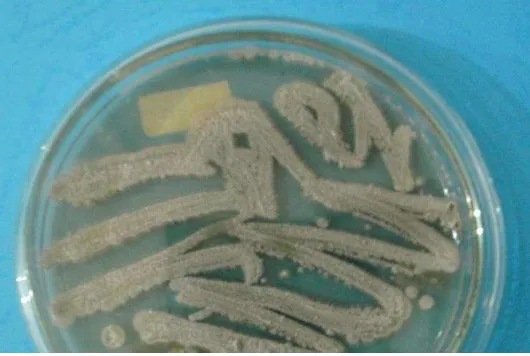
Gambar 2. Isolat Actinomycetes pada media ISP-2
D. Enzim
Enzim adalah biokatalis yang dihasilkan oleh jaringan yang meningkatkan laju reaksi kimia yang berlangsung di jaringan (Montegomery, 1993). Enzim merupakan unit fungsional dari metabolisme sel. Bekerja dengan urutan-urutan teratur, enzim mengkatalisis ratusan reaksi bertahap yang menguraikan molekul nutrien, reaksi yang mengubah kimiawi, dan yang membuat makromolekul sel (Lehninger, 1988). Berdasarkan strukturnya, enzim terdiri atas komponen yang disebut apoenzim yang berupa protein dan komponen lain yang disebut gugus prostetik yang berupa nonprotein. Gugus prostetik dibedakan menjadi koenzim dan kofaktor. Koenzim berupa gugus organik yang pada umumnya merupakan vitamin, seperti vitamin B1, B2, NAD+ (Nicotinamide Adenine Dinucleotide). Kofaktor berupa gugus anorganik yang biasanya berupa ion-ion logam, seperti Cu2+, Mg2+, dan Fe2+. Beberapa jenis vitamin seperti kelompok vitamin B merupakan koenzim. Jadi, enzim yang utuh tersusun atas bagian protein yang aktif yang disebut apoenzim dan koenzim, yang bersatu dan kemudian disebut holoenzim (Rahayu, 1990).
1. Penggolongan Enzim
1. Oksidoreduktase, enzim-enzim yang mengkatalisis berbagai macam reaksi oksidasi-reduksi.
2. Transferase, merupakan kelompok enzim yang mengkatalisis reaksi pe- mindahan berbagai gugus seperti amino, karboksil, karbonil, metil, asil, glikosil, atau fosforil.
3. Hidrolase, merupakan kelompok enzim yang mengkatalisis pemutusan ikatan antara karbon dengan atom lain sambil mengikat molekul air. 4. Liase, merupakan kelompok enzim yang mengkatalisis pemecahan ikatan
antara karbon dengan karbon, karbon dengan belerang serta beberapa jenis ikatan antara karbon dengan nitrogen (tidak termasuk ikatan peptida). 5. Isomerase, merupakan kelompok enzim yang mengkatalisis reaksi
rasemisasi isomer optik atau geometrik dan reaksi-reaksi oksidasi-reduksi intramolekular tertentu. Namun trivial dari enzim-enzim yang termasuk kelas ini antara lain adalah epimirase, rasemase, dan mutase.
6. Ligase, merupakan kelompok enzim yang mengkatalisis pembentukan ikatan antara karbon dengan oksigen, belerang, nitrogen, dan atom-atom lain.
2. Fungsi dan Cara Kerja Enzim
Seperti katalis lainnya enzim dapat menurunkan energi aktivasi suatu reaksi kimia.
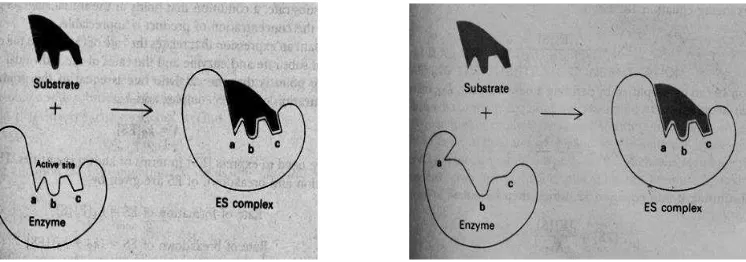
Saat berlangsungnya reaksi enzimatik terjadi ikatan sementara antara enzim dengan substrat (reaktan). Ikatan sementara ini bersifat labil dan hanya untuk waktu yang singkat saja. Selanjutnya ikatan enzim-substrat akan pecah menjadi enzim dan hasil akhir. Enzim yang terlepas kembali setelah reaksi dapat berfungsi lagi sebagai biokatalisator untuk reaksi yang sama. Enzim bekerja dengan dua cara, yaitu menurut Teori Kunci-Gembok (Lock and Key Theory) dan Teori Kecocokan Induksi (Induced Fit Theory).
Gambar 3. Pembentukan kompleks enzim-substrat berdasarkan Teori Kunci-Gembok (Lock and Key Theory) dan Teori Kecocokan Induksi (Induced Fit Theory).
konfigurasi semula. Berbeda dengan teori kunci gembok, menurut teori kecocokan induksi reaksi antara enzim dengan substrat berlangsung karena adanya induksi substrat terhadap situs aktif enzim sedemikian rupa sehingga keduanya merupakan struktur yang komplemen atau saling melengkapi. Menurut teori ini situs aktif tidak bersifat kaku, tetapi lebih fleksibel (Murray, 1997).
3. Faktor-Faktor Yang Mempengaruhi Reaksi Enzimatik
Terdapat beberapa faktor yang mempengaruhi reaksi enzimatik, antara lain adalah:
1. Substrat (reaktan)
Kecepatan reaksi enzimatik umumnya dipengaruhi kadar substrat. Penambahan kadar substrat sampai jumlah tertentu dengan jumlah enzim yang tetap, akan mempercepat reaksi enzimatik sampai mencapai maksimum. Penambahan substrat selanjutnya tidak akan menambah kecepatan reaksi (Orten dan Neuhaus, 1970).
2. Suhu
3. Kemasaman (pH)
pH dapat mempengaruhi aktivitas enzim. Daya katalisis enzim menjadi rendah pada pH rendah maupun tinggi, karena terjadi denaturasi protein enzim. Enzim mempunyai gugus aktif yang bermuatan positif (+) dan negatif (-). Aktivitas enzim akan optimum kalau terdapat keseimbangan antara kedua muatan. Pada keadaan asam cenderung bermuatan positif, dan pada keadaan basa cenderung bermuatan negatif, sehingga aktivitas enzim menjadi berkurang atau bahkan menjadi tidak aktif. pH optimum untuk masing-masing enzim tidak selalu sama (Orten dan Neuhaus, 1970).
4. Penghambat enzim (inhibitor)
Inhibitor enzim adalah zat atau senyawa yang dapat menghambat enzim. Ada beberapa cara penghambatan enzim, seperti penghambat secara bersaing (kompetitif), penghambat tidak bersaing (non-kompetitif), penghambat umpan balik (feed back inhibitor), dan penghambat alosterik (Wolfe, 1993). 5. Waktu Inkubasi
Waktu yang diperlukan oleh enzim untuk bereaksi secara optimum dengan produk tidaklah seragam, ada beberapa yang membutuhkan waktu yang lebih lama untuk bereaksi (Orten dan Neuhaus, 1970).
E. Protein
nitrogen dan kadang kala sulfur serta fosfor (Wood. J. H et al, 1966). Protein berperan penting dalam struktur dan fungsi semua sel makhluk hidup dan virus.
Kebanyakan protein merupakan enzim atau subunit enzim. Jenis protein lain berperan dalam fungsi struktural atau mekanis, seperti misalnya protein yang membentuk batang dan sendi sitoskeleton. Protein terlibat dalam sistem kekebalan (imun) sebagai antibodi, sistem kendali dalam bentuk hormon, sebagai komponen penyimpanan (dalam biji) dan juga dalam transportasi hara. Sebagai salah satu sumber gizi, protein berperan sebagai sumber asam amino bagi orga-nisme yang tidak mampu membentuk asam amino tersebut (heterotrof).
Protein merupakan salah satu dari biomolekul raksasa, selain polisakarida, lipid, dan polinukleotida, yang merupakan penyusun utama makhluk hidup. Selain itu, protein merupakan salah satu molekul yang paling banyak diteliti dalam biokimia. Protein ditemukan oleh Jöns Jakob Berzelius pada tahun 1838.
Biosintesis protein alami sama dengan ekspresi genetik. Kode genetik yang dibawa DNA ditranskripsi menjadi RNA, yang berperan sebagai cetakan bagi translasi yang dilakukan ribosom. Sampai tahap ini, protein masih "mentah", hanya tersusun dari asam amino proteinogenik. Melalui mekanisme pascatranslasi, terbentuklah protein yang memiliki fungsi penuh secara biologi.
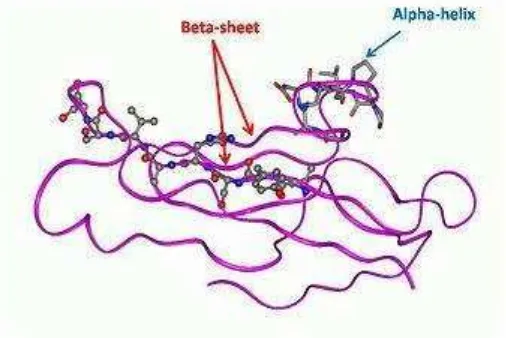
dimensi lokal dari berbagai rangkaian asam amino pada protein yang distabilkan oleh ikatan hidrogen. Gabungan dari aneka ragam dari struktur sekunder akan menghasilkan struktur tiga dimensi yang dinamakan struktur tersier. Struktur tersier biasanya berupa gumpalan. Beberapa molekul protein dapat berinteraksi secara fisik tanpa ikatan kovalen membentuk oligomer yang stabil (misalnya dimer, trimer, atau kuartomer) dan membentuk struktur kuartener.
Gambar 4. Struktur tersier protein. Protein ini memiliki banyak struktur sekunder beta-sheet dan alpha-helix yang sangat pendek
F. Protease
amino-ekso-peptidase dari gugus amino terminal, sedang endopeptidase memecah ikatan peptida dari dalam (Bergmann, 1942; Mubarik et al., 2000).
Menurut Harley (1960) dalam Winarno (1986), endopeptidase dapat dibedakan berdasarkan gugus reaktif pada sisi aktif yang terlibat dalam katalisis, menjadi:
a. Protease serin, artinya memiliki residu dalam lokasi aktifnya.
b. Protease Sulfhidril, artinya mempunyai residu sulfhidril pada lokasi aktifnya. Enzim ini dihambat oleh senyawa oksidator, senyawa yang menyebabkan suasana menjadi alkali, dan logam berat.
c. Protease metal, yaitu protease yang keaktifannya tergantung pada adanya metal per mol enzim. Metal tersebut dapat terdiri dari Mg, Zn, Co, Fe, Hg, Ni, dan sebagainya. Enzim tersebut dihambat oleh EDTA yang dapat mengkhelat metal sehingga keaktifan enzim hilang.
d. Protease asam, yaitu enzim yang pada lokasi aktifnya terdapat dua gugus karbonil. Keaktifannya dapat dihambat oleh p-bromofenasilbromida.
Penggunaan enzim protease dalam berbagai produk komersial semakin meluasnya
sejalan dengan kemajuan dalam bidang bioteknologi. Protease dimanfaatkan
dalam bidang industri antara lain dalam pengolahan pangan, penyamakan kulit,
kebutuhan ini masih tergantung pada produksi impor. Salah satu cara
mengantisipasi ketergantungan terhadap impor tersebut perlu ada usaha untuk
memproduksi enzim protease (Daniel, 1979; Suhartono, 2000; Thomas, 1984).
Enzim protease dapat diperoleh dari empat macam sumber, yaitu: a) Tumbuhan, misalnya dari pepaya.
b) Hewan, misalnya pankreas sapi. c) Bakteri, misalnya Bacillus subtilis.
d) Fungi, misalnya Aspergillus niger, Aspergilus flavus.
G. Isolasi dan Pemurnian Enzim
Proses isolasi enzim haruslah memperhatikan letak enzim tersebut. Ada dua macam letak enzim, yaitu enzim yang berada di dalam sel yang disebut dengan intraseluler dan enzim yang berada diluar sel yang disebut ekstraseluler. Proses isolasi enzim yang berada diluar sel lebih mudah dilakukan dibandingkan dengan isolasi enzim yang berada di dalam sel, hal ini dikarenakan tidak perlu memecah dinding sel untuk mengeluarkan enzimnya dan sel lebih mudah dipisahkan dari pengotornya (Gumbira, 1987). Tahap – tahap proses isolasi dan pemurniaan enzim yang paling dasar menurut Darwis dan Sukara (1990) serta Judoamijojo et al (1989) adalah sebagai berikut:
1. Lisis Dinding Sel
dengan beberapa cara, antara lain dengan perlakuan senyawa anorganik seperti asam atau basa kuat yang mampu melarutkan komponen dinding sel, dengan menggunakan enzim lisozim. Pemecahan dinding sel secara mekanik atau fisik dapat dilakukan dengan cara pembekuan atau pencairan, ultrasonifikasi, dan homogenasi.
2. Sentrifugasi (Pemusingan)
Molekul – molekul dengan berat molekul yang besar dapat mengendap dengan cepat bila disentrifugasi dengan kecepatan tinggi. Kecepatan pengendapan molekul bergantung pada beberapa faktor seperti besar molekul, bentuk molekul, dan viskositas larutan. Tetapi kecepatan pengendapan molekul tersebut sangat bergantung pada berat molekulnya.
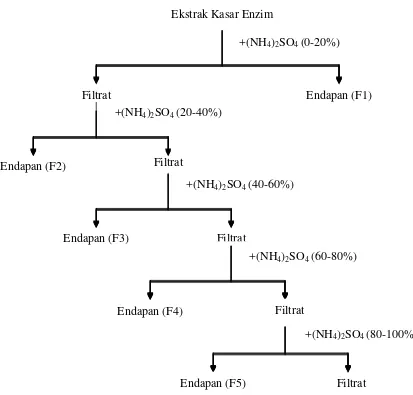
3. Fraksinasi
Cara pemurnian enzim yang umum dilakukan adalah dengan proses pengendapan bertahap atau biasa disebut fraksinasi. Pengendapan protein yang sering dilakukan adalah dengan senyawa elektrolit menggunakan garam amonium sulfat, (NH4)2 SO 4. Penambahan senyawa ini menyebabkan menurunnya kelarutan
enzim tersebut, sehingga terbentuk endapan protein.
(20-40)% jenuh, (40-60)% jenuh, (60-80)% jenuh, (80-100)% jenuh. Pengendapan ini dikenal sebagai salting out.
4. Dialisis
Dialisis adalah suatu proses pemisahan molekul – molekul besar dari ion – ion sederhana dengan menggunakan membran berdasarkan difusi. Proses ini terjadi karena adanya perbedaan tekanan osmosa antara cairan yang ada di dalam membran dengan cairan yang ada di luar membran. Membran yang digunakan adalah kantung selofan berbentuk selang.
III. METODE PENELITIAN
A. Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan November 2009 sampai dengan bulan Mei 2010 di Laboratorium Biomassa dan Laboratorium Biokimia Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Lampung.
B. Alat dan Bahan
Dalam penelitian ini alat-alat yang digunakan adalah microplate reader Hospitex Diagnostix, inkubator CO2 Memmert-Germany/INC-02, orbital shaker Wiggen
Hauser/OS150, autoclave Kleinfeld-Germany/HV-L25, centrifuge Hitachi/CF-46RX, laminar air flow ESCO/AVC4A1, water Bath Wiggen Hauser, jarum ose, pinset, lampu spiritus, mikropipet, magnetic stirer, timbangan, penangas air, lemari pendingin, kertas saring, termometer, cawan petri, dan peralatan gelas lainnya seperti labu erlenmeyer, gelas beker, labu takar, tabung reaksi, spatula, pipet tetes dan lain-lain.
selofan, buffer pospat pH 7. Sumber enzim adalah isolat actinomycetes yang diperoleh dari lumpur hutan bakau Pantai Ringgung, Perairan Teluk Lampung.
C. Prosedur Penelitian
1. Pembuatan Media dan Larutan Pereaksi
a. Pembuatan Media Pertumbuhan dan Peremajaan actinomycetes
1). Media ISP-2 untuk Peremajaan actinomycetes
Media ISP-2 terdiri dari 0,4% yeast extract; 1% malt extract; 0,4% dextrose dan 2,4% agar dilarutkan dalam air laut steril, lalu di sterilisasi selama 15 menit, pada 121ºC, 1 atm (Margavey, N. A., et al., 2004).
2). Pembuatan Mineral Salt Medium untuk Produksi Enzim
Mineral salt medium dibuat dengan cara (0,6% NaCl; 0,01% CaCl2; 0,01%
MgSO4·7H2O; 0,1% FeSO4·7H2O; 0,1% KH2PO4), ditambahkan dengan 2% skim
milk, lalu dilarutkan ke dalam air laut steril sampai pH 7,0. Kemudian di sterilisasi selama 15 menit pada 121ºC, 1 atm.
b. Pembuatan Media Uji Biokimia Protease
c. Pembuatan Pereaksi Untuk Pengukuran aktivitas Protease Metode Kunitz Termodifikasi
1) 0,65% larutan kasein
Sebanyak 0,6 gram kasein dilarutkan dalam buffer Phosfat pH 7,5 2) 110mM Larutan Asam Trikloro Asetat (TCA)
Sebanyak 1,7985 gram TCA dilarutkan kedalam labu takar 100 mL dengan aguades lalu encerkan hingga garis batas.
3) Pereaksi Lowry
Pereaksi A (Na2CO3), reagen Folin – Ciocelteau 1 N.
4) Larutan Standar L-Tirosin
Larutan tirosin dengan konsentrasi 1,1 mM.
d. Pembuatan Pereaksi Untuk Pengukuran Kadar Protein Dengan Metode Lowry
1). Pereaksi A: 2 g Na2CO3 dilarutkan dalam 100 mL NaOH 0,1 N.
2). Pereaksi B: 2 mL larutan CuSO4· 5H2O 1% ditambahkan ke dalam larutan
NaK-tartarat 1%.
3). Pereaksi C: 2 mL pereaksi B ditambahkan 100 mL pereaksi A. 4). Pereaksi D: Reagen Folin-Ciocelteau 1 N.
2. Peremajaan dan Pengamatan Actynomicetes
3. Uji Proteolitik
Media yang digunakan untuk uji biokimia biakan actinomycetes dalam mendegradasi protein adalah Mineral Salt Medium yang mengandung 10% gelatin. Sebanyak 10 mL media dimasukkan ke dalam tabung reaksi, kemudian dimasukkan biakan actinomycetes dan diinkubasi selama 4 hari. Setelah 4 hari lalu dipindahkan ke dalam lemari es dan di simpan selama 6 jam pada suhu 4ºC. Selanjutnya, biakan actinomycetes dikeluarkan dari dalam lemari es dan didiamkan selama 10 menit pada suhu ruang, lalu diamati tabung reaksi mana yang mencair.
4. Penentuan Kondisi Optimum Pertumbuhan Isolat Actinomycetes
Media yang digunakan untuk menentukan kondisi optimum actinomycetes untuk menghasilkan enzim protease adalah Mineral Salt Medium yang mengandung (0,6% NaCl; 0,01% CaCl2; 0,01% MgSO4·7H2O; 0,1% FeSO4·7H2O; 0,1%
KH2PO4). Pada masing-masing media diberikan kondisi lingkungan yang
bervariasi yaitu variasi pH dan waktu inkubasi. Variasi pH yg dilakukan adalah 6, 6.5, 7, 7.5 dan pH 8. Variasi waktu inkubasi yang dilakukan adalah 24, 48, 72, 96, 120, 144, 168, 192, 216, 240 jam.
Setelah waktu pengamatan selesai, dilakukan pengukuran jumlah sel dengan melakukan pembacaan optical density (OD) pada panjang gelombang 600 nm menggunakan mikroplate rider. Dari data hasil pengukuran OD600 dapat diketahui
5. Penyiapan Inokulum
Isolat bakteri yang telah ditumbuhkan pada media ISP-2 selama 7 hari diinokulasikan ke dalam Erlenmeyer 500 mL yang berisi 100 mL media produksi dengan pH optimum dan kemudian diinkubasi pada temperatur optimum dan waktu inkubasi optimum. Biakan ini disebut starter atau inokulum.
6. Isolasi dan Pemurnian Enzim
Isolasi protein enzim dilakukan dengan menumbuhkan starter pada media produksi dengan pH optimum dan waktu inkubasi maksimum, agar diperoleh jumlah maksimal enzim yang diproduksi actinomycetes tersebut. Untuk memisahkan larutan enzim dari konstituen seluler lainnya dilakukan sentrifugasi pada 3500 rpm selama 30 menit. Filtrat yang diperoleh disebut ekstrak kasar enzim, terhadap ekstrak kasar enzim tersebut dilakukan penentuan kadar protein dengan metode Lowry dan uji aktifitas enzim menggunakan metode Kunitz termodifikasi.
a. Fraksinasi dengan Amonium Sulfat
Gambar 5. Skema proses fraksinasi enzim dengan penambahan amonium sulfat
Setelah itu endapan protein enzim dipisahkan dari filtratnya dengan sentrifugasi pada kecepatan 6000 rpm selama 15 menit. Kemudian endapan yang diperoleh dilarutkan dalam buffer pospat pH 7 .
aktifitas enzim metode Kunitz termodifikasi pada panjang gelombang 660 nm dan ditentukan kadar protein menggunakan metode Lowry pada panjang gelombang 600 nm.
7. Karakterisasi Enzim
a. Penentuan Temperatur Optimum
Untuk mengetahui suhu optimum dari enzim hasil isolasi, dilakukan pengukuran aktifitas enzim dengan metode Kunitz termodifikasi, dengan variasi temperatur yang digunakan adalah 30, 40, 50, 60, 70 dan 80ºC.
b. Penentuan pH Optimum
Untuk mengetahui pH optimum dari enzim hasil isolasi, dilakukan pengukuran aktifitas enzim menggunakan metode Kunitz termodifikasi dengan variasi pH yang digunakan adalah 6; 6,5; 7; 7,5; dan 8.
c. Penentuan Waktu Inkubasi Maksimum
Untuk mengetahui waktu inkubasi optimum dari enzim hasil isolasi, dilakukan pengukuran aktifitas enzim menggunakan metode Kunitz termodifikasi dengan variasi waktu inkubasi 30, 40, 50, 60, dan 70 menit.
d. Penentuan Nilai Km dan Vm
optimum. Nilai Km dan Vm ditentukan dengan menggunakan kurva Lineweaver-Burk.
8. Uji Aktifitas Protease dan Penentuan Kadar Protein
Uji aktifitas protease dilakukan pada tahap isolasi, tiap tahap pemurnian dan pada saat karakterisasi enzim hasil isolasi dan pemurnian. Penentuan kadar protein di- lakukan pada tahap isolasi dan pada tiap tahap pemurnian.
a. Pengujian Aktifitas Protease Metode Kunitz Termodifikasi
Analisis aktifitas dilakukan menurut metode Kunitz menggunakan substrat kasein (Soedigdo, 1988). Sedangkan pada penelitian ini digunakan medote Kunitz Termodifikasi (Anson, M.L., 1938). Pengukuran didasarkan pada satu unit enzim akan menghidrolisis kasein untuk menghasilkan warna dari tirosin per menit, dimana warna yang dihasilkan berasal dari reagen Folin-Ciocelteau. Prosedur pengujian adalah sebagai berikut: 5 ml kasein dimasukkan ke dalam tabung reaksi kemudian ditambahkan1 ml larutan enzim. Kemudian diinkubasi pada suhu 37ºC selama 10 menit. Setelah itu masukkan 5mL reagen TCA, lalu diinkubasi pada suhu 37ºC selama 30 menit menit, tabung reaksi dikeluarkan lalu disaring dan didapatkan filtratnya. Kemudian Filtrat yang didapatkan, diambil sebanyak 2 mL dan ditambahkan 5 mL reagen Na2CO3 dan 1 mL reagen Folin Ciucelteau.
berdasarkan jumlah asam amino (peptida sederhana) yang terbentuk dengan menggunakan kurva standar tirosin. Aktivitas 1 unit enzim ditetapkan sebagai jumlah enzim yang dibutuhkan untuk menguraikan 1µg tirosin dari kasein di dalam 1 ml volume reaksi per menit .
b. Penentuan Kadar Protein ( Metode Lowry)
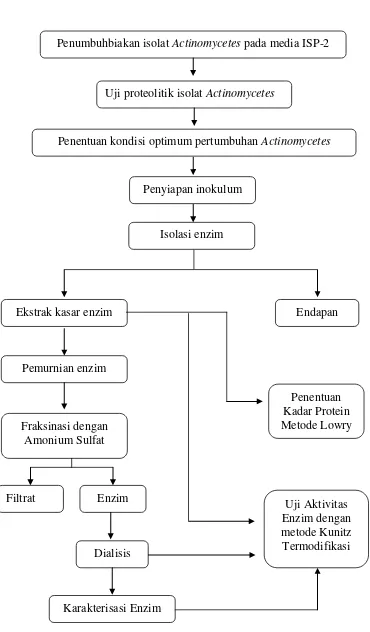
Gambar 6. Diagram alir penelitian
Penentuan Penumbuhbiakan isolat Actinomycetes pada media ISP-2
V. SIMPULAN DAN SARAN
1. Simpulan
Berdasarkan hasil penelitian yang telah dilakukan, maka dapat diambil beberapa kesimpulan sebagai berikut:
a. Dari ketiga isolat yang telah diremajakan, yaitu 4, 12 dan ANL-2b-3, yang memiliki aktivitas proteolitik adalah ANL-2b-3.
b. Fase eksponensial pertumbuhan actinomycetes tercapai pada waktu 144 jam dengan aktivitas enzim protease tertinggi pada waktu 120 jam.
c. Aktivitas spesifik enzim pada ekstrak kasar enzim sebesar 1,386 U/mL. Aktivitas spesifik enzim setelah tahap pemurnian sebesar 8,44 U/mL terjadi peningkatan kemurnian enzim sebesar 6,08 kali.
2. Saran
I. PENDAHULUAN
A. Latar Belakang Masalah
Lautan Indonesia merupakan bagian dari Indopasifik yang merupakan salah satu pusat keanekaragaman biota laut terbesar di dunia (Van Soest, 1994). Pada garis pantai lautan Indonesia terdapat hutan bakau atau disebut hutan mangrove. Hutan bakau ini merupakan hutan yang hidup digaris pantai dan dipengaruhi oleh pasang surut air laut (Irwanto, 2007).
Hutan bakau ini tumbuh khususnya di tempat-tempat dimana terjadi pelumpuran dan akumulasi bahan organik. Baik di teluk-teluk yang terlindung dari gempuran ombak, maupun di sekitar muara sungai dimana air melambat dan mengendapkan lumpur yang dibawanya dari hulu. Lumpur yang mengendap ini mengandung sejumlah mikroorganisme. Mikroorganisme yang ada di sekitar bakau dapat mendegradasi daun dan batang bakau, rumput laut serta mineral lain yang dapat digunakan sebagai sumber makanan (Anonim, 2006). Salah satu mikroorganisme yang terdapat di lingkungan hutan bakau ini adalah actinomycetes (Jensen et al., 1991; Das et al., 2006).
dan pada bidang pertanian. Selain itu, actinomycetes laut dapat bersimbiosis dengan berbagai invertebrata laut, terutama sponga. Actinomycetes menarik perhatian karena mereka dapat melakukan metabolisme yang unik dan kemampuan fisiologis yang menghasilkan produk alami (Salas, 2009). Actinomycetes mempunyai kemampuan sitotoksik dengan aktivitas farmakologi yang menarik dan karena itu mereka menyediakan sumber produk alami dengan aplikasi potensial dalam waktu dekat (Sanchez, 2009). Selain itu, Actinomycetes berpotensi menghasilkan senyawa metabolit sekunder, enzim selulase, enzim
protease dan enzim kitinase (Magarvey et al., 2004; Suryanto dan Yurnaliza,
2005; Rifaat et al., 2007).
Di Indonesia kebutuhan akan enzim protease juga semakin meningkat namun kebutuhan ini masih tergantung pada produksi impor. Salah satu cara mengantisipasi ketergantungan terhadap impor tersebut perlu ada usaha untuk memproduksi enzim protease dari mikroorganisme (Daniel, 1979; Suhartono, 1989; Thomas, 1984) . Selain itu, untuk melakukan peningkatan produksi enzim oleh mikroorganisme dapat dilakukan dengan memanipulasi kondisi lingkungan. Penggunaan mikroorganisme untuk produksi enzim mempunyai beberapa kelebihan, diantaranya mudah diproduksi dalam skala besar, waktu produksi relatif pendek serta dapat diproduksi secara berkesinambungan dengan biaya yang relatif rendah (Basuki, 1997).
Ringgung Teluk Lampung (Lina, 2009). Enzim protease dari bakteri mulai diperkenalkan tahun1960-an oleh Gebruder Schyder dari Swiss dan Novo Industri A/S dari Denmark, dan sampai sekarang penggunaan bakteri sebagai penghasil protease mempunyai peluang yang besar (Basuki, 1997).
Dalam penelitian ini, enzim protease yang akan diisolasi dan dikarakterisasi. Hal ini dikarenakan enzim protease yang dihasilkan dari actinomycetes belum banyak dimanfaatkan dan dikelola secara optimal. Ini tentunya merupakan aset besar yang berpeluang untuk dimanfaatkan dalam pengembangan bioteknologi. Dari hasil penelitian yang dilakukan oleh Vonothini et al (2008), protease dari Actinomycetes telaga India mempunyai temperatur optimum 40ºC, dan pH optimumnya adalah 7. Hasil dari penelitian tersebut dapat dipakai sebagai acuan untuk penelitian yang akan penulis lakukan. Pada penelitian ini penulis mencoba mengisolasi dan mengkarakterisasi enzim protease dari actinomycetes lumpur hutan bakau.
B. Tujuan
Tujuan penelitian ini adalah
1. Mempelajari kondisi pH dan waktu inkubasi optimum bagi pertumbuhan sel actinomycetes Isolat Lumpur Hutan Bakau dalam memproduksi enzim protease
C. Manfaat
Manfaat dari penelitian ini adalah
1. Dapat dimanfaatkannya actinomycetes Isolat Lumpur Hutan Bakau sebagai sumber alternatif penghasil enzim protease
DAFTAR PUSTAKA
Albert, B., D. Bray, J. Lewis, M. Raff, K. Robert, and J. D.Watson. 1994. Biologi Molekular Sel, edisi kedua, Jilid I. Alih Bahasa oleh Alex Tri Kantjono W. PT. Gramedia Pustaka Utama. Jakarta. Halaman 278-281. Alina, A. 2003. Isolasi Bakteri Penghasil Enzim Protease Alkalin Termostabil.
Bulletin Flasma Nutfah, Vol 9, No.2.
Anonim. 2006. Profil Mangrove Center Lampung.
Http://Mangrove.Unila.Ac.Id/Index/Php. Dibuka pada 17 September 2009, pukul 11.00 WIB
Anson, M.L., (1938) J. Gen. Physiol. 22, 79-89
Basuki, W. 1997. Enzim dalam industri deterjen. Proceeding the first
Conference on Industrial Enzyme and Biotechnology; Technology, Strategy and Finance, September 10–11, Jakarta.
Bergmann M, 1942. A classification of proteolytic enzymes. Adv. Enzymol. Beynom, R. J., J. S. Bond. 1989. Proteolytic enzymes: A Practical Approach.
Academic Press. Oxford. Halaman 128-131.
Bronikowski, A. M. 2001. Evolutionary Adaptation to Temperature. VIII. Effects of Temperature on Growth Rate in Natural Isolates of Escheria coli and Salmonella enterica From Different thermal Environments. Evolution. Volume 55, Nomor 1, Halaman 33-40.
Crill, W. D., R B. Huey., G. W.Gilchrist. 1996. Within and
Between-Generation Effects of Temperature on The Morphology and Physiology of Drosophila melanogaster. Evolution. Volume 50, Halaman 1205-1218. Daniel ICW, 1979. Fermentation and Enzyme Technology. John Willey and Sons,
New York.
Darwis, A. A. dan Sukara. 1990. Teknologi Mikrobial. Pusat Antar Universitas. IPB. Bogor.
Davidson, Michael. 2007. "Bacteria Cell Structure." Molecular Expressions: Cell Biology and Microscopy--Structure and Function of Cells and Viruses. Florida State University. Florida.
Gumbira, S. E. 1989. Biokonversi. Departemen Pendidikan dan Kebudayaan. Dirjen Dikti, PAU Bioteknologi IPB. 124-126.
Irwanto, 2007. Suksesi Hutan Mangrove Pulau Marsegu.
Http://Irwantoshut.Com/ .Dibuka pada 14 Februari 2008, pukul 11.00 WIB Judoamijojo, R. M., S. E. Gumbira, L. Hartato. 1989. Biokonversi.
Departemen Pendidikan dan kebudayaan. Dirjen Dikti, PAU Bioteknologi IPB. Bogor.
Lehninger, A. L. 1988. Dasar-Dasar Biokimia. Alih Bahasa oleh Maggy Thenawijaya. Erlangga. Jakarta.
Lina, F. G. 2009. Isolasi dan Seleksi Actinomycetes Penghasil Enzim Selulase dari Lumpur Hutan Bakau Pantai Ringgung Perairan Teluk Lampung. [Skripsi]. Universitas Lampung. Bandar Lampung.
Lowry, O.H., Rosenbrough, N.J., Farr, A.L. & Randall, R.J., 1951, Protein Measurement with the Folin Phenol Reagent, J. Biol. Chem, 193: 265-275 Magarvey, N.A., J.M. Keller, V. Bernan, M. Dworkin, and D.H. Sherman. 2004. Isolation and Characterization of Novel Marine-Derived Actinomycete Taxa
Rich in Bioactive Metabolites†. Applied and Environmental Microbiology, Dec. 2004, P. 7520–7529 Vol. 70, No. 12
Mangunwidjaja.D,Suryani.A. 1994. Teknologi Bioproses. Penebar Swadaya. Jakarta. Halaman 72-79.
Michael, J., Pelczar, Jr., E. C. S. Chan. 2008. Dasar-Dasar Mikrobiologi 1. Alih Bahasa oleh Ratna Siri Hadioetomo. UI-Press. Jakarta.
Mubarik NR., A. Suwanto dan MT. Suhartono. 2000. Isolasi dan karakterisasi protease ektraseluler dari isolat bakteri termofilik ekstrim. Prosiding Seminar Nasional Industri Enzim dan Bioteknologi II. Mikrobiologi, Enzim dan Bioteknologi Dalam Perspektif Ekonomi dan Industri. Badan Pengkajian dan Penerapan Teknologi, Jakarta, 151–158.
Murray, R. K. 1997. Biokimia Harper. Alih Bahasa oleh dr. Andri Hartono. Penerbit Buku Kedokteran EGC. Jakarta.
Noor, R. M., M. Khazali., N. Suryadiputra. 1999. Panduan Pengenalan Mangrove di Indonesia. Direktur Jendral PKA / Wetlands International Indonesia Programme. Bogor.
Orten, J. M., O. W Neuhaus. 1970. Biochemistry. C. V. Mosby Company. Saint Louis.
Rahayu, K. 1990. Enzim Mikroba. Universitas Gajah Mada. Yogyakarta. Rao, N. 1994. Mikrooganisme Tanah dan Pertumbuhan Tanaman. UI press.
Jakarta. Halaman 17-18.
Rifaat, H.M., O.H. El-Said, S.M. Hassanein and M.S. M. Selim. 2007. Protease Activity of Some Mesophilic Streptomycetes Isolated from Egyptian Habitats. Journal of Culture Collections Vol. 5, 2006-2007, Pp. 16-24 Salas, J. A. 2009. Marine Actinomycetes: A New of Natural Product. Fakultas
Kedokteran. Universitas Oviedo. Spanyol. [Paper]
Sanchez, MC. 2009. Peran Kelautan Actinomycetes dalam Drug Discovery pada PharmaMan. Mikrobiologi R & D Dpt. PharmaMan. Spanyol. [Paper] Soedigdo. 1988. Metode Penelitian Biokimia. PAU Bioteknologi Institut
Teknologi Bandung. Bandung.
Soemitro, S. 2005. Pengaruh Modifikasi Kimiawi Selektif Terhadap Kestabilan Enzim Amilase dari Saccharomycopsis fibuligera. J. Bionatura. Vol. 7 (3): 259-273
Spalding, M. D., F. Blasco and C. D. Field (Eds). 1997. World Bakau Atlas. The International Society of Bakau Ecosystem. Okinawa, Japan. 179 pp. Stefani, Dwi. 2005. Isolasi dan Karakterisasi Enzim Protease dari Bakteri Isolat
Lokal. [Skripsi]. Universitas Lampung. BandarLampung.
Suhartono MT, 2000. Eksplorasi protease bakteri asal Indonesia untuk aplikasi industri dan riset bioteknologi. Prosiding Seminar Nasional IndustriEnzim dan Bioteknologi II. 125–133.
Suryanto, D., E.M. dan Yurnaliza. 2005. Eksplorasi Bakteri Kitinolitik : Keragaman Genetik Gen Penyandi Kitinase pada Berbagai Jenis Bakteri dan Pemanfaatannya. USU
Van Soest, R.W.M. 1994. “Demosponge distribution pattern, In: Van Kompen,
T.M.G, Braekman, J.C. (Eds), Sponge in Time and Space”, A.A. Balkema. Rotterdam.
Vonothini, G et al. 2008. Optimization of protease production by an actinomycete Strain, PS-18A isolated from an estuarine shrimp pond. African Journal of Biotechnology.Vol. 7 (18), pp. 3225-3230.
Winarno, F.G. 1986. Enzim Pangan. PT. Gramedia. Jakarta. Halaman 79-82. Wolfe, Stephen L. 1993. Molecular and Cellular Biology. Wadsworth
DAFTAR ISI
Faktor-Faktor Yang Mempengaruhi Reaksi Enzimatik ... 15
Protein ... 16
Protease ... 18
Isolasi dan Pemurnian Enzim ... 20
Lisis Dinding Sel ... 20
Sentrifugasi (Pemusingan) ... 21
Pembuatan Media dan Larutan Pereaksi ... 24
Peremajaan dan Pengamatan Actinomycetes………... ... 25
Uji Proteolitik ... 26
Penentuan Kondisi Optimum Pertumbuhan Isolat Actinomycetes ... 26
Penyiapan Inokulum ... 27
Isolasi dan Pemurnian Enzim ... 27
Fraksinasi dengan Amonium Sulfat ... 27
Dialisis ... 28
Karakterisasi Enzim ... 29
Uji Aktivitas Protease dan Penentuan Kadar Protein ... 30
HASIL PENGAMATAN DAN PEMBAHASAN ... 33
Peremajaan Isolat Actinomycetes dan Uji Proteolitik ... 33
Penentuan Kondisi Optimum Pertumbuhan Isolat Actinomycetes ... 34
Isolasi dan Pemurnian Enzim ... 37
Fraksinasi dengan Ammonium Sulfat ... 38
Dialisis………... ... 49
Karakterisasi Enzim ... 41
Penentuan Waktu Inkubasi Optimum Enzim Protease ... 41
Penentuan pH Optimum Enzim Protease ... 42
Penentuan Temperatur Optimum Enzim Protease... 44
Konstanta Michaelis-Menten ... 45
SIMPULAN ... 47
Simpulan ... 47
Saran ... 48
DAFTAR PUSTAKA ... 49
DAFTAR TABEL
Tabel Halaman
1. Hasil pemurnian enzim protease ... 40
2. Pengaruh waktu inkubasi terhadap pertumbuhan isolat Actinomycetes ... 56
3. Pengaruh variasi pH terhadap pertumbuhan isolat Actinomycetes. ... 56
4. Hasil dialisis setelah fraksinasi ... 56
5. Penentuan waktu inkubasi optimum enzim protease ... 57
6. Penentuan pH optimum enzim protease ... 57
7. Penentuan temperatur optimum enzim ... 57
8. Penentuan nilai Km dan Vmax ... 58
9. Data standar Tirosin ... 59
DAFTAR GAMBAR
Gambar Halaman
1. Bentuk-bentuk sel bakteri ... 9
2. Isolat Actinomycetes pada media ISP-2 ... 11
3. Pembentukan kompleks enzim-substrat berdasarkan teori kunci gembok (Lock and Key Theory) dan teori kecocokan induksi (Induced Fit Theory). ... 14
4. Struktur tersier protein ... 18
5. Skema proses fraksinasi enzim dengan penambahan amonium sulfat ... 28
6. Diagram alir penelitian ... 32
7. Isolat actinomycetes ... 33
8. Uji proteolitik dari isolat actinomycetes ... 34
9. Hubungan antara jumlah sel (OD600) dan aktivitas unit (U/mL) dengan waktu inkubasi pertumbuhan Actinomycetes ... 35
10. Kurva Hubungan pH dengan jumlah sel pada OD600 ... 37
11 Kurva Hubungan antara AUP dengan tiap fraksi setelah dialisis. ... 40
12. Kurva Hubungan antara waktu inkubasi optimum dengan AUP ... 42
13. Kurva Hubungan antara pH optimum dengan AUP ... 43
ISOLASI DAN KARAKTERISASI ENZIM PROTEASE DARI
Actinomycetes ISOLAT LUMPUR HUTAN BAKAU (Skripsi)
Oleh
RIKI ERIYANTI ANDINI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
ISOLASI DAN KARAKTERISASI ENZIM PROTEASE DARI
Actinomycetes ISOLAT LUMPUR HUTAN BAKAU
Oleh
RIKI ERIYANTI ANDINI
Skripsi
Sebagai salah satu syarat untuk mencapai gelar Sarjana Sains
pada Jurusan Kimia
Fakultas Matematika dan Ilmu Pengetahuan Alam
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
Judul Penelitian : Isolasi dan Karakterisasi Enzim Protease dari Actinomycetes Isolat Lumpur Hutan
Bakau
Nama Mahasiswa : Riki Eriyanti Andini
Nomor Pokok Mahasiswa : 0517011051
Jurusan : Kimia
Fakultas : Matematika dan Ilmu Pengetahuan Alam
MENYETUJUI
1. Komisi pembimbing
Dian Herasari, M.Si Dr. Eng. Suripto Dwi Yuwono, M.T. NIP. 19710806200003.2.001 NIP. 19740705200003.1.001
2. Ketua Jurusan
MENSAHKAN
1. Tim Penguji
Ketua : Dian Herasari, M.Si ...
Sekretaris : Dr. Eng. Suripto Dwi Yuwono, M.T. ...
Penguji
Bukan Pembimbing : Dra. Aspita Laila, M.S ...
2. Dekan Fakultas MIPA
Dr. Sutyarso, M.Biomed. NIP 195704241987031001
PERSEMBAHAN
Sebuah karya kecilku ini penulis persembahkan teruntuk:
Papa dan mama tercintaYang selalu memberikan kasih
sayangnya, tak pernah lelah bermandi peluh dan selalu
mendoakanku dalam sujudnya
Adik-adikku Tersayang (adek Tiwi dan adek Geo)
dan
Seseorang yang akan menemani hidupku kelak
Almamater Tercinta
Menjadi kuat
bukan berarti mampu melakukan segala hal
tapi menjadi kuat adalah
mampu berdiri tegak dan
mampu melangkah disaat yang lain tak
RIWAYAT HIDUP
Penulis dilahirkan di Kotabumi, Lampung Utara pada tanggal 9 Juni 1987 dan
merupakan anak pertama dari tiga bersaudara, putri dari pasangan Bapak Ir.
Misnandar Kaswan dan Ibu Suryati Asnawi, A.Md.
Penulis memulai pendidikan di Taman Kanak-Kanak Darma Wanita Ogan Lima,
dan diselesaikan pada tahun 1993. Kemudian melanjutkan pendidikan ke
Sekolah Dasar di SDN 1 Kotagapura, Kotabumi, Lampung Utara pada tahun 1999,
kemudian melanjutkan ke Sekolah Menengah Pertama di SLTPN 1 Kotabumi dan
menamatkan pendidikan pada tahun 2002. Selanjutnya penulis menyelesaikan
pendidikan Sekolah Menengah Umum di SMAN 3 Kotabumi pada tahun 2005.
Pada tahun yang sama penulis diterima sebagai mahasiswa Jurusan Kimia
Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Lampung melalui
jalur ujian Seleksi Penerimaan Mahasiswa Baru (SPMB).
Selama menjadi mahasiswa, penulis pernah menjadi anggota HIMAKI (Himpunan
Mahasiswa Kimia) pada tahun 2005. Pada tahun 2006 penulis pernah mengikuti
kegiatan KUILMA (Kunjungan Ilmiah Mahasiswa) di daerah Serang, Banten. Pada
tahun 2007 penulis mengikuti kegiatan KWI (Kunjungan Wisata Ilmiah) di daerah
menjadi mahasiswa, penulis pernah menjadi asisten praktikum untuk mata
Puji syukur penulis ucapkan atas kehadirat ALLAH SWT karena rahmat dan
hidayah-Nya sehingga penulis dapat menyelesaikan penulisan skripsi yang
berjudul :
ISOLASI DAN KARAKTERISASI ENZIM PROTEASE DARI ACTINOMYCETES ISOLAT LUMPUR HUTAN BAKAU
Skripsi ini disusun berdasarkan penelitian yang telah dilakukan sebagai syarat
untuk mencapai gelar Sarjana Sains pada Jurusan Kimia Fakultas Matematika dan
Ilmu Pengetahuan Alam Universitas Lampung.
Dalam kesempatan ini saya mengucapkan terima kasih kepada :
1. Ibu Dian Herasari, M.Si., selaku pembimbing utama dan pembimbing
akademis, atas kesediaannya meluangkan watu, memberi bimbingan, dan
pengarahan selama penyusunan skripsi ini.
2. Ibu Nurhasanah, M.Si., selaku pembimbing II, atas bantuan, saran dan bimbingannya.
3. ibu Dra. Aspita Laila, M.S., selaku penguji yang telah memberikan saran dan kritik untuk perbaikan skripsi ini.
6. Staf dosen dan karyawan di Lingkungan Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Lampung.
7. Papa, mama, adik-adikku (Pertiwi Surya Sari dan Geo Surya Andika) serta seluruh keluargaku atas dukungan dan doa kalian semua.
8. Seseorang yang kutunggu kedatangannya untuk menemani sisa umurku kelak. 9. Sahabat terbaikku, Pita Rini(Cacing) yang selama bertahun-tahun ini telah
memberikan support, doa dan perhatiannya, terima kasih untuk persahabatan yang indah ini.
10. Sahabat-sahabat terbaikku tice, tika sapi, ony, helm, liya.
11. Fera Yuliantina, S.Si. teman satu tim dan satu pembimbing yang telah berjuang bersamaku.
12. Teman-temanku satu angkatan dan teman-teman kimia 06 dan Tim Laboratorium Biomassa, Reni, Slamet, Peni, Pi2t, Sana, Kak bowo.
Akhirnya, penulis menyadari bahwa informasi yang penulis sajikan pada skripsi ini masih jauh dari sempurna, namun semoga skripsi ini dapat bermanfaat bagi semua.