IDENTIFIKASI KERAGAMAN GEN
SIGNAL TRANSDUCER
AND ACTIVATOR OF TRANSCRIPTION
5A (STAT5A)
PROMOTER
PADA SAPI BALI
RIRI JUNIARTI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Identifikasi Keragaman Gen Signal Transducer and Activator of Transcription 5A (STAT5A) Promoter pada Sapi Bali adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2015
ABSTRAK
RIRI JUNIARTI. Identifikasi Keragaman Gen Signal Transducer and Activator of Transcription 5A (STAT5A) Promoter pada Sapi Bali. Dibimbing oleh ASEP GUNAWAN dan JAKARIA.
Gen signal transducer and activator of transcription 5A (STAT5A) merupakan gen yang mengontrol sifat reproduksi sapi dan memiliki hubungan signifikan terhadap fertilitas dan perkembangan embrio. Penelitian ini bertujuan untuk mengidentifikasi keragaman genetik gen STAT5A promoter pada sapi bali yang berasal dari BPTU Pulukan Bali, BPT-HMT Serading, dan VBC Kabupaten Barru. Sapi bali yang diidentifikasi sebanyak 42 sampel dengan menggunakan metode PCR-sekuensing. Amplifikasi gen STAT5A promoter menghasilkan fragmen dengan panjang 603 bp. Hasil sekuens gen STAT5A promoter ditemukan 11 SNPs. SNP yang teridentifikasi pada daerah CpG island I yaitu g.-1448C>A,
g.-1409C>A, g.-1405C>A, g.-1401C>A, g.-1450G>A, g.-1361G>A, g. -1384A>G, dan g.-1350G>T. SNP yang ditemukan pada daerah CpG island II yaitu g.-1167C>T, g.-1128G>A, dan g.-1093G>A. Selain itu, analisis SNP menunjukkan mutasi di gen STAT5A daerah promoter dapat menjadi kandidat awal marker seleksi potensial untuk sifat reproduksi. Studi validasi lebih lanjut diperlukan untuk membuktikan peran gen STAT5A untuk digunakan dalam seleksi genom.
Kata kunci: CpG island, gen STAT5A, sapi bali, sekuensing, SNP
ABSTRACT
RIRI JUNIARTI. Identification of Signal Transducer and Activator of Transcription 5A (STAT5A) Gene Promoter Polymorphisms in Bali Cattle. Supervised by ASEP GUNAWAN and JAKARIA.
Signal transducer and activator of transcription 5A (STAT5A) is gene that control the reproductive traits of cattle and had a significant relationship with fertility and embryonic development. The aim of this study was to identify the genetic variability of STAT5A genes promoter in Bali cattle from BPTU Pulukan Bali, BPT-HMT Serading, and VBC Kabupaten Barru. A total of 42 bali cattle were identified using PCR-sequencing method. STAT5A gene promoter amplifications resulted fragments with the lengths of 603 bp. The results sequences of STAT5A gene promoter were found 11 SNPs. SNPs identified in the CpG island I were g.-1448C>A, g.-1409C>A, g.-1405C>A, g.-1401C>A, g. -1450G>A, g.-1361G>A, g.-1384A>G, and g.-1350G>T. SNPs were found in CpG island II were g.-1167C>T, g.-1128G>A, and g.-1093G>A. Morever, SNPs analysis revealed mutation in promoter region STAT5A gene, could be potensial preliminary candidate marker selection for reproductive trait. Further validation studies are required for proving the roles of STAT5A gene to be use in genomic selection.
IDENTIFIKASI KERAGAMAN GEN
SIGNAL TRANSDUCER
AND ACTIVATOR OF TRANSCRIPTION
5A (STAT5A)
PROMOTER
PADA SAPI BALI
RIRI JUNIARTI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada
Departemen Ilmu Produksi dan Teknologi Peternakan
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala nikmat, berkah dan rahmat-Nya sehingga skripsi ini dapat diselesaikan. Shalawat dan salam senantiasa terlimpah kepada Rasulullah SAW, keluarga, para sahabat dan pengikutnya. Terima kasih sebesar-besarnya penulis ucapkan kepada Dr agr Asep Gunawan, SPt MSc selaku pembimbing akademik dan pembimbing skripsi, Dr Jakaria, SPt MSi selaku pembimbing skripsi atas segala ilmu, bimbingan dan motivasi yang telah diberikan. Terima kasih kepada Edit Lesa Aditia, SPt MSc selaku penguji sidang atas segala masukan yang telah diberikan.
Terima kasih penulis ucapkan kepada ayahanda dan ibunda tercinta, Zaenal Arifin dan Erius Erina yang senantiasa memberikan do’a, kasih sayang, motivasi serta dukungan moril maupun materil. Ungkapan terima kasih juga disampaikan kepada kakak-kakak dan adik tersayang, Erni Findriawati SE, Eries Hermawandi SE dan Dodi Rindiansyah SKom, Sinta Septiani serta adik Fifi Melati Putri atas segala do’a, motivasi, dan senyumannya. Terima kasih kepada Prof Dr Ir Cece Sumantri MAgrSc yang senantiasa memberikan motivasi, masukan, dan inspirasi selama penulis berada di Laboratorium Genetika Molekuler Ternak. Di samping itu, penulis sampaikan terima kasih kepada Kak Alit, Kak Selvi, Kak Isyana, Kak Olin, Kak Ferdy, Kak Furqon, Kak Eryk, Alwiyah, Mustika, Pramujo, Hadi, Rindang, Ninin dan teman-teman laboratorium genetika molekuler ternak atas segala ilmu, motivasi, dan kebersamaannya selama ini. Terima kasih kepada teman seperjuangan alih jenis Ahmad, Endah, Yuninda, Adita, dan Ichi serta keluarga besar Domilion IPTP 48 atas segala semangat dan dukungannya. Kepada Mute, Isti, Kasita, Ilma, Rani, dan Andin terima kasih atas do’a, ukhuwah dan kebersamaannya. Penulis juga sampaikan terima kasih kepada Septiani DA SHut, drh Maya Shofa dan Rahmawati NF SSi atas segala do’a, dukungan dan diskusinya selama ini. Penulis berharap semoga Allah SWT berkenan membalas segala kebaikan semua pihak yang telah membantu. Semoga karya ilmiah ini bermanfaat.
Bogor, September 2015
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan 2
Ruang Lingkup Penelitian 2
METODE 2
Lokasi dan Waktu Penelitian 2
Alat 3
Bahan 3
Prosedur 3
Ekstraksi DNA 3
Amplifikasi DNA 4
Elektroforesis 4
Sekuensing Fragmen Gen STAT5A Promoter 5
Analisis Data 5
HASIL DAN PEMBAHASAN 6
Amplifikasi Gen STAT5A Promoter 6
Struktur Gen STAT5A dan CpG Islands 6
SNPs (Single Nucleotide Polymorpihsms) Gen STAT5A 8
SIMPULAN 14
SARAN 14
DAFTAR TABEL
1 Kriteria penentuan CpG island 8
2 Nilai frekuensi genotipe dan alel SNPs di CpG islands gen STAT5A pada
sapi bali 11
3 SNPs gen STAT5A serta asosiasinya pada beberapa bangsa sapi 13
DAFTAR GAMBAR
1 Posisi penempelan primer pada fragmen gen STAT5A promoter 4 2 Hasil amplifikasi gen STAT5A promoter pada sapi bali 6
3 Rekonstruksi gen STAT5A pada sapi 6
4 CpG island gen STAT5A promoter pada sapi bali 7
5 SNPs pada hasil sekuen gen STAT5A promoter pada sapi bali 9
DAFTAR LAMPIRAN
1 Sekuens gen STAT5A yang diakses di GenBank kode akses AY280369.1 18 2 Sekuens gen STAT5A yang diakses di GenBank kode akses AJ242522.1 19 3 Sekuens gen STAT5A yang diakses di GenBank kode akses AJ237937.1 23 4 Homologi gen STAT5A promoter pada sapi bali dengan Bos taurus
(GenBank AJ242522.1) 30
5 Komposisi basa sekuen gen STAT5A promoter dan CpG islands pada sapi
PENDAHULUAN
Latar Belakang
Sapi bali (Bos javanicus) merupakan salah satu sapi asli Indonesia yang berasal dari domestikasi banteng (Bibos banteng) (Payne dan Rollison 1973). Populasi sapi bali cukup besar dengan wilayah penyebarannya yang luas di Indonesia dan sebagian besar berada di kepulauan Indonesia bagian timur (Handiwirawan dan Subandriyo 2004; Talib et al. 2003). Sapi bali memiliki keunggulan dibandingkan sapi lokal lainnya yaitu fertilitas dan persentase karkas yang tinggi, kadar lemak daging rendah dan mampu memanfaatkan pakan berkualitas rendah (Handiwirawan dan Subandriyo 2004). Selain itu, Talib (2002) menegaskan bahwa sapi bali memiliki kemampuan reproduksi yang baik dan daya adaptasi lingkungan yang tinggi. Kemampuan reproduksi yang baik ini merupakan potensi besar untuk meningkatan populasi sapi nasional dalam rangka swasembada daging nasional. Namun, dibanding kelebihan tersebut sapi bali memiliki kelemahan diantaranya tingkat kematian pra sapih yang tinggi disebabkan kapasitas produksi air susu induk yang rendah dan cekaman lingkungan yang tidak menunjang (Talib 2002). Sapi bali memiliki tingkat kematian pedet yang tinggi sekitar 8%-48% dan produksi susu 164-274.5 kg/6 bulan (Talib et al. 2003). Upaya perbaikan sifat reproduksi pada sapi Bali dapat dilakukan melalui perbaikan mutu genetik melalui seleksi baik secara konvensional maupun molekuler. Seleksi secara molekuler akan lebih efektif dilakukan untuk sifat kuantitatif dengan nilai heritabilitas sedang hingga tinggi seperti sifat reproduksi pada sapi bali (Gunawan et al. 2011). Sifat reproduksi sapi bali diantaranya umur pertama beranak, jarak kelahiran, laju kebuntingan memiliki nilai heritabilitas masing-masing yaitu 0.21, 0.41 dan 0.40 (Gunawan et al. 2011).
dilakukan pada sapi Dutch Holstein-Frisian dan berhubungan signifikan terhadap komposisi lemak susu (Schennink et al. 2009). Alel G pada ekson 8 posisi 12195 berkorelasi dengan penurunan protein dan persentase lemak susu (Khatib et al. 2008). Sebagian besar penelitian gen STAT5A yang telah dilakukan umumnya adalah bagian ekson dan intron, namun penelitian pada bagian promoter belum banyak dilakukan. Promoter merupakan sekuen DNA pada gen yang bertanggung jawab dalam ekspresi gen tersebut dan tempat melakukan transkripsi dalam proses perakitan protein (Brandenberg et al. 2011). Pada promoter sebagian gen tertentu memiliki CpG island yang akan berpengaruh terhadap terjadinya metilasi DNA dan berakibat pada ekspresi gen tersebut. Oleh sebab itu penelitian ini dilakukan untuk mengetahui keragaman gen STAT5A promoter pada sapi bali.
Tujuan
Penelitian ini bertujuan mengidentifikasi keragaman gen signal transducer and activator of transcription 5A (STAT5A) promoter pada sapi bali di pusat pembibitan sapi bali yaitu BPTU Pulukan Bali, BPT-HMT Serading NTB, dan Village Breeding Centre (VBC) Kabupaten Barru di Sulawesi Selatan dengan menggunakan metode polymerase chain reaction dan sekuensing. Selain itu, penelitian ini bertujuan untuk mengetahui SNPs pada CpG islands yang ditemukan serta keragaman genetik pada tiga daerah tersebut.
Ruang Lingkup Penelitian
Penelitian ini meliputi identifikasi SNPs dan keragaman gen signal transducer and activator of transcription 5A (STAT5A) promoter pada 42 ekor sapi bali yang terdapat di tiga pusat pembibitan sapi bali yaitu BPTU Pulukan Bali (16 ekor), BPT-HMT Serading (10 ekor), dan VBC Kabupaten Barru (16 ekor). Keragaman gen STAT5A promoter dianalisis menggunakan teknik Polymerase Chain Reaction dan sekuensing. Analisis data yang dilakukan yaitu identifikasi SNPs, CpG island, frekuensi alel dan genotipe. Analisis hasil sekuensing dilakukan dengan mensejajarkan (alignment) sekuen nukleotida, Basic Local Alignment Search Tools (BLAST) nukleotida dan identifikasi CpG island melalui MethPrimer.
METODE
Lokasi dan Waktu Penelitian
Fakultas Peternakan, Institut Pertanian Bogor. Penelitian berlangsung pada bulan Februari sampai Mei 2015.
Alat
Alat yang digunakan pada ekstraksi DNA terdiri dari tabung eppendorf 1.5 ml, rak tabung, satu set pipet mikro dan tip, vortex, inkubator, microsentrifuge, dan tilter. Alat yang digunakan pada PCR dan elektroforesis terdiri dari mesin PCR thermocycler, satu set pipet mikro dan tip, tabung eppendorf, rak tabung, vortex mixer, microsentrifuge, pinset, refrigerator, stirer, magnet stearer, satu set alat pencetak gel, timbangan, gelas ukur, microwave, power supply 100 volt, dan UV transiluminator.
Bahan
Sampel darah yang digunakan adalah sampel darah sapi bali dengan total 42 sampel yang berasal dari Balai Pembibitan Ternak Unggul (BPTU) Pulukan Bali sebanyak 16 ekor, BPT-HMT Serading NTB sebanyak 10 ekor, dan Village Breeding Center (VBC) Kabupaten Barru sebanyak 16 ekor. Bahan yang digunakan dalam ekstraksi DNA meliputi Destilation water (DW), larutan EtOH absolute 70%, NaCl 0.2%, SDS (sodium dodesil sulfat), enzim Proteinase-K, 1x STE (sodium tris EDTA), CIAA (kloroform iso amil alkohol), etanol 70%, etanol absolut, fenol, dan TE (tris EDTA) 80%. Bahan yang digunakan dalam PCR adalah sampel DNA, DW, 5x Kappa 2G Buffer A, DMSO (dimethyl sulfoxide), MgCl2, dNTPs, Taq Polymerase serta pasangan primer forward dan reverse. Primer yang digunakan adalah forward: 5’-GCAGGGGAGTGCACATGAAA-3’ dan reverse: 5’-GCAGCGATTTCCTCCTCAAAG-3’. Primer dibuat menggunakan Primer Designing Tool dan primer dicek pada Primer Stats. Bahan yang digunakan dalam elektroforesis adalah produk PCR, 0.5x TBE (tris borat-EDTA), agarose, EtBr (Etidium Bromida), dan marker 100 bp. Bahan yang digunakan untuk sekuensing yaitu produk PCR gen STAT5A promoter dan primer forward dan reverse.
Prosedur
Ekstraksi DNA
disentrifugasi menggunakan kecepatan 12 000 rpm selama lima menit untuk memisahkan fenol dari molekul DNA dalam air. Fase DNA akan terbentuk dan dipindahkan sebanyak 40 μl ke tabung 1.5 ml yang baru. Selanjutnya, sampel ditambahkan 40 μl NaCl 5 M dan 800 μl etanol absolut. Tahap selanjutnya sampel didiamkan semalaman (overnight) dengan suhu -20oC. Molekul DNA disentrifugasi dengan kecepatan 12 000 rpm selama lima menit dan supernatan dibuang. Hasil endapan DNA didiamkan hingga kering, kemudian ditambahkan pelarut 100 μl buffer TE 80%.
Amplifikasi DNA
Sampel DNA sebanyak 2 μl dimasukkan ke dalam tabung 0.2 ml. Pereaksi amplifikasi DNA terdiri dari 17.9 μl DW, 6 μl 5x Kapa, 2 μl DMSO, 0.5 μl MgCl2, 0.5 μl dNTPs, 0.5 μl masing-masing primer dan 0.1 μl Taq Polymerase. Seluruh pereaksi dimasukkan ke dalam tabung 1.5 ml lalu dihomogenkan. Pereaksi tersebut sebanyak 28 μl dimasukkan ke dalam tabung 0.2 ml yang berisi sampel DNA. Sampel DNA yang telah ditambahkan pereaksi di spin down selama beberapa detik, kemudian dimasukkan ke dalam mesin PCR ESCO. Kondisi amplifikasi DNA di dalam mesin PCR pada tahap awal yaitu predenaturasi 95°C selama 5 menit. Pada tahap kedua 35 siklus terdiri dari denaturasi pada suhu 95°C selama 10 detik, annealing pada suhu 64°C selama 20 detik dan ekstensi pada suhu 72°C selama 30 detik. Tahap ketiga yaitu ekstensi akhir pada suhu 72°C selama 5 menit. Posisi penempelan primer gen STAT5A promoter pada amplifikasi DNA berdasarkan sekuen GenBank dengan kode akses AY280369.1 disajikan pada Gambar 1.
Gambar 1 Posisi penempelan primer pada fragmen gen STAT5A promoter
Elektroforesis
menit. Larutan agarose gel dihomogenisasi menggunakan magnetic stirer kemudian ditambahkan EtBr sebanyak 2.5 μl. Larutan agarose tersebut dimasukkan ke dalam alat pencetak gel dan didiamkan hingga mengeras. Produk PCR sebanyak 5 μl dimasukkan ke dalam sumur-sumur gel dan marker DNA 100 bp dimasukkan ke dalam sumur paling kiri. Proses selanjutnya gel dialiri listrik pada tegangan 100 volt selama 30-45 menit. Proses elektroforesis selanjutnya yaitu gel divisualisaikan menggunakan bantuan sinar UV pada mesin UV transiluminator. Hasil elektroforesis akan menunjukkan pita-pita dengan panjang fragmen DNA tersebut.
Sekuensing Fragmen Gen STAT5A Promoter
Sekuensing gen STAT5A promoter digunakan untuk mendapatkan sekuen DNA berupa untaian lambang nukleotida penyusun DNA yang akan dianalisis. Sekuensing dilakukan menggunakan mesin sekuenser (ABI Prims 3100-Avant Genetic Analyzer) melalui jasa perusahaan sekuensing 1st Base di Selangor, Malaysia.
Analisis Data
Data yang digunakan pada penelitian ini merupakan data sekunder. Analisis dilakukan pada hasil sekuen dengan program BioEdit (Hall 1999), serta dianalisis menggunakan metode BLAST (www.ncbi.nhl.nih.gov./BLAST) untuk mengetahui kesamaan dengan gen STAT5A promoter GenBank. Hasil sekuen dianalisis
keberadaan CpG island menggunakan MethPrimer
(http://www.urogene.org/methprimer/). Keberadaan mutasi atau SNP (Single Nucleotide Polymorphism) pada sekuen fragmen gen STAT5A promoter dianalisis menggunakan program Molecular Evolutionary Genetic Analysis 5 (MEGA5) (Tamura et al. 2011). Frekuensi genotipe gen STAT5A promoter dihitung berdasarkan rumus (Nei dan Kumar 2000) :
Keterangan :
xii = frekuensi genotipe ke-ii nii = jumlah individu bergenotipe ii N = jumlah individu sampel
Frekuensi alel gen STAT5A promoter dihitung berdasarkan rumus (Nei dan Kumar 2000) :
Keterangan :
xi = frekuensi alel ke-i
HASIL DAN PEMBAHASAN
Amplifikasi Gen STAT5A Promoter
Amplifikasi fragmen gen STAT5A promoter menghasilkan panjang produk PCR yaitu 603 bp (Gambar 2). Amplifikasi dilakukan dengan menggunakan suhu annealing 64°C selama 20 detik menggunakan mesin thermocycler ESCO. Suhu annealing merupakan suhu yang optimal untuk primer menempel pada DNA komplementer selama proses amplifikasi. Suhu annealing berkisar antara 50-65°C (Brandenberg et al. 2011). Waktu annealing dipengaruhi oleh kapasitas pemanasan mesin thermocycler, volume campuran PCR, konsentrasi primer dan gen target (Pelt-Verkuil et al. 2008).
Gambar 2 Hasil amplifikasi gen STAT5A promoter pada sapi bali
Struktur Gen STAT5A dan CpG Islands
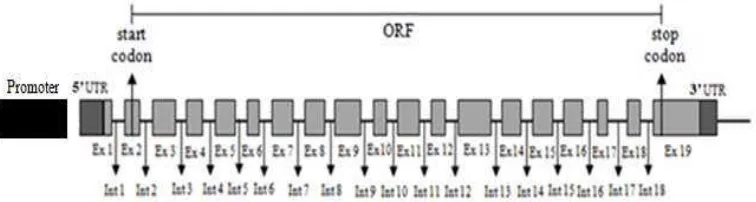
Posisi gen STAT5A berada pada kromosom 19 yang terdiri atas 19 ekson yang mengkode 794 rantai asam amino (Seyfert et al. 2000). Rekonstruksi gen STAT5A disajikan pada Gambar 3.
Signal transducers and activators of transcription (STATs) merupakan 7 anggota famili dari faktor transkripsi yang memediasi aksi beberapa hormon
peptida dan cytokines dalam sel target (Darnell et al. 1994; Schindler dan Darnell 1995). STAT5 dikenal sebagai mamary gland factor (MGF) karena berperan dalam faktor transkripsi induksi prolaktin dan juga dapat mengaktifkan transkripsi gen protein susu sebagai respon terhadap prolaktin (Wakao et al. 1994). Mediator utama dalam aksi hormon pertumbuhan pada gen target juga diperankan oleh STAT5 (Argetsinger dan Carter-Su 1996) serta berinteraksi dan bersinergi untuk glucocorticoid dan insulin (Lechner et al. 1997). STAT5 berperan penting dalam ekspresi gen sexually dimorphic di hati yang ditentukan oleh pola sekresi GH (growth hormon) dan pertumbuhan tubuh (Herrington et al. 2000). Selain itu, laju fertilitas dan perkembangan embrio memiliki hubungan yang signifikan dengan gen STAT5A (Khatib et al. 2009).
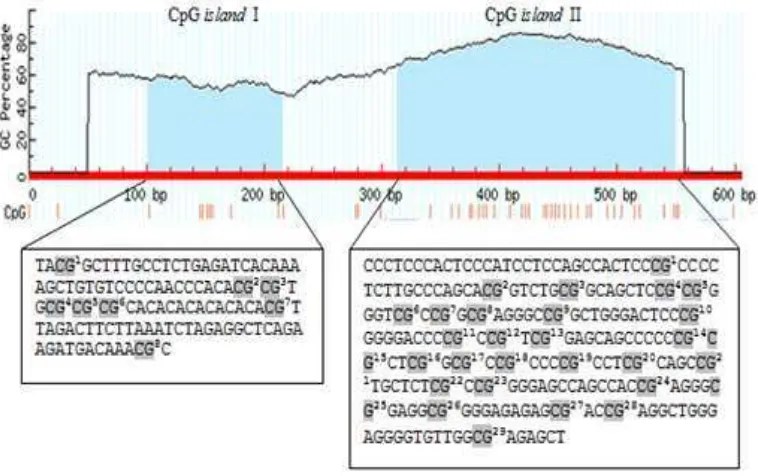
Keberadaan CpG island dapat diidentifikasi menggunakan MethPrimer dengan cara memasukkan sekuen target. Sekuen gen STAT5A promoter pada sapi bali yang telah dimasukkan ke dalam MethPrimer menunjukkan adanya 2 daerah CpG island yang dapat dilihat pada Gambar 4.
Gambar 4 CpG islands gen STAT5A promoter pada sapi bali
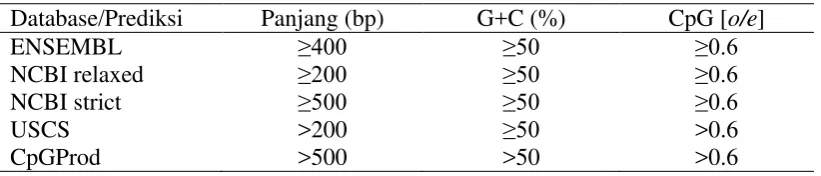
4) menunjukkan bahwa panjang CpG island I yaitu 115 bp dan CpG island II yaitu 236 bp, serta kandungan G+C masing-masing yaitu 52.17% dan 78.39%. Kriteria CpG island yang digunakan pada MethPrimer tersebut adalah panjang >100 bp, % GC >50.0, dan Obs/Exp >0.6. Situs CpG pada CpG island I sebanyak 8 situs CpG dan CpG island II sebanyak 29 situs CpG (Gambar 4). Kriteria dalam penentuan CpG island cukup beragam, namun kriteria yang digunakan dalam penentuan CpG island umumnya berdasarkan Gardiner-Garden dan Frommer (1987) seperti yang digunakan pada beberapa browser genom lainnya yang disajikan pada Tabel 1.
Tabel 1 Kriteria penentuan CpG island
Database/Prediksi Panjang (bp) G+C (%) CpG [o/e]
ENSEMBL ≥400 ≥50 ≥0.6
NCBI relaxed ≥200 ≥50 ≥0.6
NCBI strict ≥500 ≥50 ≥0.6
USCS >200 ≥50 >0.6
CpGProd >500 >50 >0.6
Sumber: Illingworth dan Bird (2009)
Keberadaan CpG island ini akan berpengaruh terhadap adanya pola metilasi DNA. Metilasi terlibat dalam silencing region DNA atau mencegah ekspresi (Clark dan Pazdernik 2009). CpG island umumnya termetilasi di dalam sel mamalia (Turner et al. 2007). Sitosina (C) yang termetilasi dapat mempengaruhi transkripsi melalui penghambatan faktor transkripsi untuk mengikat DNA atau perekrutan protein regulasi seperti histon memodifikasi dan kromatin-remodeling oleh enzim (Fazzari dan Greally 2004), sedangkan struktur kromatin dan nucleosomal merupakan kunci regulator dalam ekspresi gen (Segal dan Widom 2009). Jika terjadi metilasi maka ekspresi gen ditekan/tidak terekspresi, sedangkan jika tidak terjadi metilasi maka gen akan terekspresi. Metilasi DNA dikenal sebagai regulator epigenetik ekspresi gen yang relevan untuk silencing transkripsi selama pengembangan dan pencetakan genetik (Li 2002; Jaenisch dan Bird 2003). Penelitian metilasi DNA pada gen STAT5A telah dilakukan pada fetus sapi yang diproduksi melalui somatic cell nuclear transfer (SCNT) (Couldrey dan Lee 2010). Hasil penelitian tersebut menunjukkan bahwa 43 dari 58 situs CpG dapat dianalisis dan tingkat metilasi DNA di CpG island antara STAT5A dan STAT5B sebagian besar di bawah 30% pada sampel ginjal, adrenal dan liver (Couldrey dan Lee 2010).
SNPs (Single Nucleotide Polymorpihsms) Gen STAT5A
Gambar 5 SNPs pada hasil sekuen gen STAT5A promoter pada sapi bali (a. SNPs di CpG island I, b. SNPs di CpG island II)
Total SNP gen STAT5A promoter yang ditemukan adalah 11 SNP dengan jumlah SNP pada CpG island I dan II masing-masing sebanyak 8 dan 3 SNP (Tabel 2). SNP yang teridentifikasi pada daerah CpG island I yaitu g.-1448C>A,
g.-1409C>A, g.-1405C>A, g.-1401C>A, g.-1450G>A, g.-1361G>A, g.- a.
1384A>G dan g.-1350G>T. SNP g.-1448C>A, g.-1450G>A, g.-1361G>A, g. -1384A>G dan g.-1350G>T ditemukan pada ketiga populasi sapi bali. Selain itu, SNP g.-1409C>A, g.-1405C>A, dan g.-1401C>A hanya ditemukan pada populasi sapi bali yang berasal dari NTB. SNP yang ditemukan pada daerah CpG island II yaitu g.-1167C>T, g.-1128G>A, dan g.-1093G>A. SNP basa C>T pada posisi g.-1167 hanya ditemukan pada populasi sapi bali di pulau Bali. Selain itu, SNP pada posisi g.-1128G>A dan g.-1093G>A ditemukan pada populasi sapi bali yang berasal dari pulau Bali dan NTB, sedangkan pada sapi bali dari Barru tidak ditemukan. Single nucleotide polymorphism (SNP) merupakan perbedaan akibat adanya subtitusi basa tunggal sehingga terjadi polimorfisme atau keragaman (Kwok dan Chen 2003). Perbedaan SNP tersebut dapat dijadikan penciri sapi bali pada lokasi tersebut. Efek adanya pasangan basa substitusi sederhana pada SNP dapat berhubungan juga dengan sifat atau penyakit tertentu (Schork et al. 2000). SNP di wilayah coding dapat langsung berdampak pada protein terkait, SNP di intron dapat mempengaruhi splicing dan SNP di promoter dapat berpengaruh terhadap ekspresi gen (Schork et al. 2000). SNP gen STAT5A pada sapi bali telah diteliti pada posisi 6853 C>T pada ekson 7 pada sapi bali yang berasal dari 3 daerah yaitu Bali, NTB dan Sulawesi Selatan (Paramitasari et al. 2015). Hasil penelitian tersebut menunjukkan bahwa lokus STAT5A|AvaI memiliki alel monomorfik C (Paramitasari et al. 2015). Selain itu, SNP gen STAT5A bagian promoter telah diteliti sebelumnya pada sapi dan ditemukan subtitusi basa A/G di posisi -488 (Flisikowski et al. 2004).
Tabel 2 Nilai frekuensi genotipe dan alel SNPs di CpG islands gen STAT5A pada sapi bali
No
SNPs Posisi SNPs Genotipe Alel
Lokasi
Bali Barru NTB
CC CA AA C A CC CA AA CC CA AA CC CA AA
CpG
island
I*
1 g.-1448C>A 0.78 (31) 0.20 (8) 0.02 (1) 0.88 0.12 0.22 (9) 0.15 (6) 0 (0) 0.37 (15) 0.02 (1) 0 (0) 0.20 (8) 0.02 (1) 0.02 (1) 2 g.-1409C>A 0.86 (36) 0.14 (6) 0 (0) 0.93 0.07 0.38 (16) 0 (0) 0 (0) 0.38 (16) 0 (0) 0 (0) 0.10 (4) 0.14 (6) 0 (0) 3 g.-1405C>A 0.93 (39) 0.05 (2) 0.02 (1) 0.95 0.05 0.38 (16) 0 (0) 0 (0) 0.38 (16) 0 (0) 0 (0) 0.17 (7) 0.05 (2) 0.02 (1) 4 g.-1401C>A 0.90 (38) 0.10 (4) 0 (0) 0.95 0.05 0.38 (16) 0 (0) 0 (0) 0.38 (16) 0 (0) 0 (0) 0.14 (6) 0.10 (4) 0 (0)
GG GA AA G A GG GA AA GG GA AA GG GA AA
5 g.-1450G>A 0.57 (24) 0.43 (18) 0 (0) 0.79 0.21 0.07 (3) 0.31 (13) 0 (0) 0.36 (15) 0.02 (1) 0 (0) 0.14 (6) 0.10 (4) 0 (0) 6 g.-1361G>A 0.73 (29) 0.27 (11) 0 (0) 0.86 0.14 0.30 (12) 0.05 (2) 0 (0) 0.35 (14) 0.05 (2) 0 (0) 0.07 (3) 0.18 (7) 0 (0)
AA AG GG A G AA AG GG AA AG GG AA AG GG
7 g.-1384A>G 0.93 (39) 0.07 (3) 0 (0) 0.96 0.04 0.36 (15) 0.02 (1) 0 (0) 0.36 (15) 0.02 (1) 0 (0) 0.21 (9) 0.02 (1) 0 (0)
GG GT TT G T GG GT TT GG GT TT GG GT TT
8 g.-1350G>T 0.83 (35) 0.17 (7) 0 (0) 0.92 0.08 0.31 (13) 0.07 (3) 0 (0) 0.31 (13) 0.07 (3) 0 (0) 0.21 (9) 0.02 (1) 0 (0)
CpG
island
II
CC CT TT C T CC CT TT CC CT TT CC CT TT
1 g.-1167C>T 0.88 (37) 0.12 (5) 0 (0) 0.94 0.06 0.26 (11) 0.12 (5) 0 (0) 0.38 (16) 0 (0) 0 (0) 0.24 (10) 0 (0) 0 (0)
GG GA AA G A GG GA AA GG GA AA GG GA AA
2 g.-1128G>A 0.57 (24) 0.43 (18) 0 (0) 0.79 0.21 0.12 (5) 0.26 (11) 0 (0) 0.38 (16) 0 (0) 0 (0) 0.07 (3) 0.17 (7) 0 (0) 3 g.-1093G>A 0.88 (37) 0.10 (4) 0.02 (1) 0.93 0.07 0.36 (15) 0 (0) 0.02 (1) 0.38 (16) 0 (0) 0 (0) 0.14 (6) 0.10 (4) 0 (0)
SNP yang teridentifikasi pada CpG island I sebanyak 8 SNP yaitu g. -1448C>A, g.-1409C>A, g.-1405C>A, g.-1401C>A, g.-1450G>A, g.-1361G>A,
g.-1384A>G, dan g.-1350G>T. Pada SNP kesatu dan ketiga ditemukan tiga macam genotipe yaitu CC, CA, dan AA, sedangkan pada SNP kedua dan keempat hanya ditemukan dua tipe genotipe yaitu CC dan CA. SNP kelima, keenam dan ketujuh hanya ditemukan dua tipe genotipe yaitu GG dan GA serta AA dan AG. Pada SNP kedelapan ditemukan dua tipe genotipe yaitu GG dan GT. Frekuensi alel pada seluruh SNP CpG island I memiliki nilai ≤ 0.99, sehingga dapat diketahui bahwa gen STAT5A promoter daerah CpG island I pada sapi bali bersifat polimorfik. Suatu alel tergolong polimorfik jika memiliki frekuensi alel ≤0.99 (Nei 1987).
Tabel 3 SNPs gen STAT5A serta asosiasinya pada beberapa bangsa sapi
SNP Region Bangsa sapi Asosiasi Sumber 9501A/G Intron 9 Jersey Produksi susu,
lemak dan kadar protein.
Brym et al. (2004)
6853C>T Ekson 7 Podolica Performa pertumbuhan
Black and White Pertumbuhan, kualitas karkas, dan konversi pakan
Oprządek et al. (2003) Iranian holstein Produksi susu Sadeghi et al. (2009) Italian Brown Produksi susu Selvaggi et al. (2009) Jersey Komposisi susu Dario dan Selvaggi
(2011) 12195G/C Ekson 8 Komposisi susu dan
daya tahan hidup
12743T>C Ekson 16 Jersey Komposisi susu Selvaggi et al. (2013) -488A>G promoter pengikatan
promoter Sapi bali - Hasil penelitian
PCR-heteroduplex dan sekuensing (Flisikowski et al. 2004). Hasil ekspresi gen dari SNP tersebut menunjukkan bahwa tingkat ekspresi pada liver sapi lebih tinggi pada genotipe AA dibandingkan GG (Flisikowski et al. 2004). Selain itu, hasil dari electrophoretic mobility shift assay (EMSA) menunjukkan bahwa transisi A→G pada posisi −488 pada gen STAT5A promoter dapat meningkatkan pengikatan kapasitas untuk nuclear proteins hati [diduga hepatic nuclear factor (HNF)-3] (Flisikowski et al. 2004). Keberadaan SNP pada CpG island dapat dijadikan dasar penelitian pola metilasi pada gen STAT5A sapi bali yang berpengaruh terhadap ekspresi gen.
SIMPULAN
Jumlah SNPs yang ditemukan yaitu 11 SNP dengan lokasi masing-masing pada CpG island I dan II yaitu 8 SNP dan 3 SNP. SNP yang teridentifikasi pada daerah CpG island I yaitu g.-1448C>A, g.-1409C>A, g.-1405C>A, g.-1401C>A,
g.-1450G>A, g.-1361G>A, g.-1384A>G, dan g.-1350G>T. SNP yang ditemukan pada daerah CpG island II yaitu g.-1167C>T, g.-1128G>A, dan g.-1093G>A. Hasil identifikasi keragaman pada kedua CpG island yang berbeda menunjukkan adanya sifat polimorfik yang dapat dijadikan kriteria awal dalam melakukan seleksi.
SARAN
SNPs yang ditemukan pada penelitian ini perlu divalidasi dalam populasi yang lebih besar, terutama pada SNPs yang konsisten ditemukan pada ketiga populasi sapi bali. SNPs gen STAT5A promoter yang teridentifikasi pada sapi bali lebih lanjut diperlukan adanya penelitian terhadap tingkat metilasi dan ekspresi gen tersebut sehingga dapat dijadikan dasar dalam melakukan seleksi.
DAFTAR PUSTAKA
Argetsinger LS, Carter-Su C. 1996. Growth hormone signalling mechanisms: involvement of the tyrosine kinase JAK2. Horm. Res. 45:22–24.
Brandenberg O, Dhlamini Z, Sensi A, Ghosh K, Sonnino A. 2011. Module Biosafety Resource Book: Introduction to Molecular Biology and Genetic Engineering. Rome (IT): FAO of the United Nations.
Brym P, Kamiñski S, Rusc A. 2004. New SSCP polymorphism within bovine STAT5A gene and its associations with milk performance traits in Black-and-White and Jersey cattle. J Appl Genet. 45(4):445-452.
Couldrey C, Lee RSF. 2010. DNA methylation patterns in tissues from mid-gestation bovine foetuses produced by somatic cell nuclear transfer show subtle abnormalities in nuclear reprogramming. BMC Dev Biol. 10:27. Dario C, Selvaggi M, Carnicella D, Bufano G. 2009. STAT5A/AvaI
polymorphism in Podolica bulls and its effect on growth performance traits. Livest Sci. 123:83–87.
Dario C, Selvaggi M. 2011. Study on the STAT5A/AvaI polymorphism in Jersey cows and association with milk production traits. Mol Biol Rep. 38(8):5387-92. doi: 10.1007/s11033-011-0691-8.
Darnell JrJE, Kerr IM, Stark GR. 1994. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science. 264(5164):1415–1421.
Deaton AM, Bird A. 2011. CpG islands and the regulation of transcription. Genes Dev. 25:1010–1022.
Fazzari MJ, Greally JM. 2004. Epigenomics: beyond CpG islands. Nat Rev Genet. 5:446–455.
Flisikowski K, Oprządek J, Dymnicki E, Zwierzchowski L. 2003. New polymorphism in the bovine STAT5A gene and its association with meat production traits in beef cattle. Anim Sci Pap Rep. 21(3):147-157.
Flisikowski K, Starzyński RR, Zwierzchowski L. 2004. Promoter variant-dependent expression of the STAT5A gene in bovine liver. BBA-Gene Struct Expr.1679(2):195-199 doi:10.1016/j.bbaexp.2004.05.005.
Gardiner-Garden M, Frommer M. 1987. CpG islands in vertebrate genomes. J Mol Biol. 196:261-282.
Goldammer T, Meyer L, Seyfert HM, Brunner RM, Schwerin M. 1997. STAT5A encoding gene maps to chromosome 19 in cattle and goat and chromosome 11 in sheep. Mamm Genome. 8:705–706.
Gunawan A, Sari R, Parwoto Y. 2011. Genetic analysis of reproductive traits in bali cattle maintained on range under artificially and naturally bred. J Indonesian Trop Anim Agric. 36(3).
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp. Ser. 41: 95-98. Handiwirawan E, Subandriyo. 2004. Potensi dan keragaman sumberdaya genetik
sapi bali. Wartazoa. 14:3.
Herrington J, Smit L, Schwartz J, Carter-Su C. 2000. The role of STAT proteins in GH signaling. Oncogene. 19(21):2587–2597.
Illingworth RS, Bird AP. 2009. CpG islands-‘A rough guide’. FEBS Letters. 583:1713-1720.
Jaenisch R, Bird A. 2003. Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genet. 33 (Suppl 1):245–254.
Khatib H, Huang W, Wang X, Tran AH, Bindrim AB, Schutzkus V, Monson RL, Yandell BS. 2009. Single gene and gene interaction effects on fertilization and embryonic survival rates in cattle. J Dairy Sci. 92(5):2238– 2247.
Kwok PY, Chen X. 2003. Detection of single nucleotide polymorphisms. Curr Issues Mol Biol. 5:43-60.
Lechner J, Welte T, Dopler W. 1997. Mechanism of interaction between the glucocorticoid receptor and STAT5: role of DNA-binding. Immunobiology. 198(1-3):112-123.
Li E. 2002. Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet. 3:662–673.
Li MJ, Liu M, Liu D, Lan XY, Lei CZ, Yang DY, Chen H. 2013. Polymorphisms in the promoter region of the Chinese bovine PPARGC1A gene. Asian-Aust J Anim Sci. 26:483-487.
Nei M. 1987. Molecular Evolutionary Genetics. New York (US): Columbia Univ Pr.
Nei M, Kumar S. 2000. Molecular Evolution and Phylogenetics. New York (US): Oxford Univ Pr.
Oikonomou G, Michailidis G, Kougioumtzis A, Avdi M, Banos G. 2011. Effect of polymorphisms at the STAT5A and FGF2 gene loci on reproduction, milk yield and lameness of Holstein cows. Res Vet Sci. 91:235–239.
Oprządek J, Flisikowski K, Zwierzchowski L, Dymnicki E. 2003. Polymorphisms at loci of leptin (LEP), Pit1 and STAT5A and their association with growth, feed conversion and carcass quality in Black-and-White bulls. Anim Sci Pap Rep. 21(3):135-145.
Paramitasari KA, Sumantri C, Jakaria. 2015. The genetic variability of prolactin and signal transducers and activators of transcription 5A (STAT5A) genes in bali cattle. Media Petern. 38(1):1-11.
Payne WJA, Rollinson DHL. 1973. Bali Cattle. World Anim Rev. 7:13-21.
Pelt-Verkuil van E, Belkum van A, Hays JP. 2008. Principles and Technical Aspects of PCR Amplification. Netherlands (NL): Springer.
Sadeghi M, Shahrbabak MM, Mianji GR, Javaremi AN. 2009. Polymorphism at locus of STAT5A and its association with breeding values of milk production traits in Iranian Holstein bulls. Livest Sci. 123:97–100
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning: A Laboratory Manual. 2nd ed. USA (US): Cold Spring Harbour Laboratory Pr.
Saxonov S, Berg P, Brutlag DL. 2006. A genome-wide analysis of CpG dinucleotides in the human genome distinguishes two distinct classes of promoters. Proc Natl Acad Sci. 103:1412–1417.
Schennink A, Bovenhuis H, Léon-Kloosterziel KM, Van Arendonk JAM, Visker MHPW. 2009. Effect of polymorphisms in the FASN, OLR1, PPARGC1A, PRL and STAT5A genes on bovine milk-fat composition. Anim Genet. 40(6): 909–916.
Schindler C, Darnell JrJE. 1995. Transcriptional responses to polypeptide ligands. The JAK- STAT pathway. Annu Rev Biochem. 64:621–651.
Schork NJ, Fallin D, Lanchbury S. 2000. Single nucleotide polymorphisms and the future of genetic epidemiology. Clin Genet. 58:250–264.
Segal E, Widom J. 2009. From DNA sequence to transcriptional behaviour: a quantitative approach. Nat Rev Genet. 10:443–456.
milk composition in Italian Brown cattle. J Dairy Res. 76:1–5. doi:10.1017/S0022029909990070.
Selvaggi M, Tufarelli V, Pinto F, Centoducati G, Dambrosio A, Santacroce MP, Dario C. 2013. Bovine STAT5A gene polymorphism analysis and its association with milk composition traits in Jersey cows. International J Bios Bioch and Bioinf. 3:4. doi: 10.7763/IJBBB.2013.V3.227.
Seyfert H, Pitra C, Meyer L, Brunner RM, Wheeler TT, Molenaar A, McCracken JY, Herrmann J, Thiesen H, Schwerin M. 2000. Molecular characterization of STAT5A and STAT5B encoding genes reveals extended intragenic sequence homogenity in cattle and mouse and different degrees of divergent evolution of various domains. J Mol Evol. 50:550-561.
Talib C. 2002. Sapi bali di daerah sumber bibit dan peluang pengembangannya. Wartazoa. 12:3.
Talib C, Entwistle K, Siregar A, Budiarti-Turner S, Lindsay D. 2003. Survey of population and production dynamics of bali cattle and existing breeding programs in Indonesia. ACIAR Proceedings. 110.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution.
Turner P, McLennan A, Andy B, Mike W. 2007. Molecular Biology 3rd Ed. New York (US): Taylor and Francis Group.
LAMPIRAN
Lampiran 1 Sekuens gen STAT5A yang diakses di GenBank kode akses AY280369.1
LOCUS AY280369 978 bp DNA linear MAM 30-JAN-2006 DEFINITION Bos taurus STAT5A gene, promoter region and exon 1. ACCESSION AY280369
VERSION AY280369.1 GI:30351072 KEYWORDS .
SOURCE Bos taurus (cattle) ORGANISM Bos taurus
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Laurasiatheria;
Cetartiodactyla; Ruminantia; Pecora; Bovidae; Bovinae; Bos. REFERENCE 1 (bases 1 to 978)
AUTHORS Flisikowski,K., Starzynski,R.R. and Zwierzchowski,L.
TITLE Promoter variant-dependent expression of the STAT5A gene in bovine
liver
JOURNAL Biochim. Biophys. Acta 1679 (2), 195-199 (2004) PUBMED 15297151
REFERENCE 2 (bases 1 to 978)
AUTHORS Flisikowski,K., Hiendleder,S. and Zwierzchowski,L. TITLE Nucleotide sequence variation in the transcription factor STAT5A
5'-noncoding region in Bos taurus and Bos indicus cattle JOURNAL Biochem. Genet. 43 (7-8), 459-464 (2005)
PUBMED 16187168 REFERENCE 3 (bases 1 to 978)
AUTHORS Flisikowski,K. and Zwierzchowski,L.
TITLE Sequence variations in the transcription factor STAT5A gene 5'-flanking region in the cattle and European bison (Bison
bonasus) JOURNAL Unpublished REFERENCE 4 (bases 1 to 978)
AUTHORS Flisikowski,K. and Zwierzchowski,L. TITLE Direct Submission
/gene="STAT5A" DEFINITION Bos taurus partial stat5A gene, exons 1-4 and joined CDS. ACCESSION AJ242522
VERSION AJ242522.1 GI:5701819
KEYWORDS STAT5A gene; STAT5A, mammary gland factor. SOURCE Bos taurus (cattle)
ORGANISM Bos taurus
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Laurasiatheria;
AUTHORS Seyfert,H.M., Pitra,C., Meyer,L., Brunner,R.M., Wheeler,T.T., Molenaar,A., McCracken,J.Y., Herrmann,J., Thiesen,H.J. and Schwerin,M.
TITLE Molecular characterization of STAT5A- and STAT5B- encoding genes reveals extended intragenic sequence homogeneity in cattle and mouse and different degrees of divergent evolution of various domains
JOURNAL Submitted (08-JUN-1999) Seyfert H.M., Molecular Biology, Research Institute for the Biology of Farm Animals, Wilhelm- Stahl-Allee-2, 18196 Dummerstorf, GERMANY
2941 gggggtgtgggaagggaagtaaaataggaacccagggccaggactataatggaaagtacc DEFINITION Bos taurus partial stat5A gene, exons 5-19.
ACCESSION AJ237937
VERSION AJ237937.1 GI:5701818
KEYWORDS Mammary Gland Factor STAT5A; STAT5A gene. SOURCE Bos taurus (cattle)
ORGANISM Bos taurus
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Laurasiatheria;
Cetartiodactyla; Ruminantia; Pecora; Bovidae; Bovinae; Bos. REFERENCE 1
AUTHORS Seyfert,H.M., Pitra,C., Meyer,L., Brunner,R.M., Wheeler,T.T., Molenaar,A., McCracken,J.Y., Herrmann,J., Thiesen,H.J. and Schwerin,M.
TITLE Molecular characterization of STAT5A- and STAT5B- encoding genes reveals extended intragenic sequence homogeneity in cattle and mouse and different degrees of divergent evolution of various domains
14461 tgggatgaccacaggcttgggagtcacaaaactggaattccagtcctgactccacctgcc 14521 taccaggtggctctgaacaagcttcctaacatggttgagcctcagtttcttcctcagtgg 14581 ctgtggggattggcataaagtggttgaaaggcccaggccttcataggtgctcagtgtctc 14641 gtgcctgagatctcatgggcagagctccttcacccaaccacctgtttttacagaggaggg 14701 cagggacaggggaaatgaattccctgagatccctcagtgagcttgtgagtgatgagacta 14761 gaaccggggtgtcttccgacaaactagtgactgactgattcagcttggtggtgcgggcat 14821 ctagccagagcaggtcctgttcccatgagactgaacggggatcaggctgggcgctgtccc 14881 ccaggagcctggggctctgcggcaggccagctccctctgatacccctctccccctcccag 14941 ccctgacccggtgctcgaccaggatggagaattcgacctggacgagaccatggatgttgc 15001 ccggcacgtggaagaactcctccgccgcccaatggacagtctggagccctctctcccccc 15061 gcccactggtctctttacccccggcagaggctcgctgtcctgaatgtttgttcgaacact 15121 gcactcctctgtggaaacaatccccagtgtgcagggtcctattcattgtgattttgtatt 15181 tgtatctctgtgcatactgatgcctttgcaggcagcccatgtacacatgtagatgtgcgc 15241 gcatttgtgtacgaggtgtgcccacctcgcctccgcagtcttaggtgtgtggctgccttg 15301 ttcttcttcagatgaaggcattccaagagctcactgggtctgtctgtggagagagaccaa 15361 gccgcttctgtggtcacaggtacgccaggcaggcagacctattcctgccagagccttagt 15421 ctgctcagcagctgtttgaatggaattatttgckaagggaaaggagacaggtaatttytc 15481 taagctcaagttttattcctgagttagctagcagcttcgcctctcagttgtgtrcgtgga 15541 tctgatctggtacagtgttggtcttttgcttcttcccctcaggagggaggtgatgagtga 15601 tattttatactttatctcctgaatagggactccagagcttttgtgaaccgagagcttgtc 15661 taggctgtggttatgtccctaaacacagcagaaaggggtcttttgcctcgggcctggagt 15721 tccctggcctcttggcccccagctcggggggtccggcttcatgtgtataatgctgctgcc 15781 cacattcctgctcttcattcccccgaccccaatcccattccctccatctggccccagcct 15841 ttccttgcctctgtctccaacyttgctcactatggaagaagttgggggagggggagagag 15901 gtycagggcttgcagggttgatcccctctcctggaaaaaacctgcag
//
Lampiran 5 Komposisi basa sekuen gen STAT5A promoter dan CpG islands pada sapi bali
Sekuen Panjang (bp)
Komposisi basa (%)
A T G C A+T G+C
RIWAYAT HIDUP
Penulis dilahirkan di Tangerang pada tanggal 04 Juni 1992. Penulis adalah anak ketiga dari pasangan Zaenal Arifin dan Erius Erina. Penulis menamatkan pendidikan SMA di Sekolah Menengah Atas Negeri (SMAN) 01 Curug pada tahun 2010. Pada tahun yang sama penulis diterima sebagai mahasiswi Program Diploma III Institut Pertanian Bogor, pada Program Keahlian Teknologi dan Manajemen Ternak melalui jalur Ujian Seleksi Masuk IPB (USMI). Pada tahun 2013 penulis melanjutkan pendidikan program sarjana alih jenis di Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.