MORFOMET
ULAT SUTER
OMETRI DAN KERAGAMAN
UTERA LIAR
Cricula trifenestrat
(Lepidoptera: Saturniidae)
SURIANA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011
MAN GENETIK
strata
Helfer
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Morfometri dan Keragaman Genetik Ulat Sutera Liar Cricula trifenestrata Helfer (Lepidoptera: Saturniidae) adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2011
ABSTRACT
SURIANA. Morphometry and Genetics Diversity of Wild Silk Moth Cricula
trifenestrata Helfer (Lepidoptera: Saturniidae). Under direction of: DEDY DURYADI SOLIHIN, RONNY RACHMAN NOOR dan ACHMAD MACHMUD THOHARI.
The study was conducted to describe morfometry, life cycle, fibroin and cytohcrome C oxidase subunit I (COI) gene characteristics of Cricula trifenestrata.Morphometry data were obtained from 3 locations at Campus IPB Darmaga area, namely in the parking area Graha Widya Graduation (GWW), Bara Tomb Complex, and Animal House C Fapet. Observations were done on the duration and timing of life cycle, viability, morphology, anatomy, and nine morphological traits, namely cocoon weight (CW), shell cocoon weight (SCW), cocoon lenght (CL) cocoon width (CWD), pupae weight (PW), pupae length (PL), pupae width (PWD), thorax length (TL), and wingspan (WS). Data were analyzed descriptively, and discriminant analysis were also performed. Molecular data obtained from genomic DNA extracted from silk glands of larvae. Genomic DNA was amplified
and sequenced, and then characterized in order to describe the number and type of
nucleotide, the number and types of amino acids on proteins fibroin and COI. Characterization results was compared with fibroin and COI genes sequences of other silk moth available in GenBank and using it to constructed phylogenetic tree. The results showed that C. trifenestrata spend 51.0 ± 7.3 days for one life cycle. All morphological traits significantly affected the grouping by location in each stage of development. The prediction accuracy for the parameter grouping cocoon, pupae and imago were 93.6%, 83.3%, and 92.7% respectively, with the biggest differentiator: CV, PW, and WS
respectively. The partial fibroin gene consists of first exon coding region with 42
nucleotides (14 amino acids), an introns (113 nucleotides) and partial coding region of second exon with 831 nucleotides (277 amino acids) long. The comparison with other silk moth showed that fibroin gene were conserved on C. trifenestrata, but very diverse with other species. Nucleotides 25th(C), 35th(T) of the first exon, and 12thamino acids (valine) were diagnostic for the species C. trifenestrata. Nucleotide of second exon encoding 277 amino acids that were dominated by the amino acid alanine (27.8%) and glycine (21.66%). Nucleotide introns were very diverse, more than 70% nucleotide of A and T. The partial COI gene ofC. trifenestrata size of 595 nucleotides, encoded 198 amino acids. There was no difference nucleotide sequence of intra species C. trifenestrata, but showed 23.7% diversity with other silk moth. Nucleotide dominated by thymine and adenine bases (± 70%). There are 25 nucleotide diagnostic for C. trifenestrata. Sequenced of 198 amino acids, showed only 10% variation between species of silk moth. Amino acid 107th(valine) and 138th(treinonin) are amino acid diagnostic forC. trifenestrata.
RINGKASAN
SURIANA. Morfometri dan Keragaman Genetik Ulat Sutera Liar Cricula
trifenestrata Helfer (Lepidoptera: Saturniidae). Dibimbing oleh: DEDY DURYADI SOLIHIN, RONNY RACHMAN NOOR dan ACHMAD MACHMUD THOHARI.
Cricula trifenestrata merupakan serangga penghasil sutera alam yang belum didomestikasi, sehingga dikenal sebagai ulat sutera liar. Serangga ini memiliki penyebaran yang luas, meliputi Asia terutama Asia Tenggara, bahkan di Indonesia ditemukan beberapa
spesies yang endemik di beberapa pulau (Nassig et al. 1996). Akhir-akhir ini C.
trifenestrata telah dimanfaatkan oleh sebagian kalangan masyarakat Indonesia untuk menghasilkan serat sutera yang dikenal sebagai sutera emas, sebab kokon sebagai sumber serat sutera dari C. trifenestrata memiliki warna asli kuning keemasan. Oleh karena itu, maka nilai jual serat sutera C. trifenestrata lebih tinggi dibandingkan sutera alam lainnya.
Di kalangan masyarakat Bali dan Jogyakarta kokon C. trifenestrata dimanfaakan pula
untuk membuat berbagai aksesoris pelengkap pakaian wanita seperti bros, anting, dan
hiasan tumah tangga seperti kap lampu, wall paper, dll. Kokon tersebut semuanya
diperoleh dari alam.
Ketersedian bahan baku dari alam menjadi kendala utama untuk pemenuhan kebutuhan akan serat sutera maupun kokon. Informasi mengenai C. trifenestrata, masih sangat terbatas, padahal potensi pengembangan serangga ini sangat besar. Seekor betina dapat menghasilkan telur ± 300. Serangga ini bersifat bivoltin, yaitu; setiap tahunnya dapat berlangsung dua kali siklus (dua generasi) dengan durasi perkembangannya yang relatif pendek, yaitu hanya sekitar dua bulan per siklus.
C. trifenestrata bersifat polifagus. Tanaman inangnya adalah alpukat, jambu mete, kayu manis, dan kedondong. Oleh karena itu penanganan yang baik mengarah pada domestikasi, diharapkan dapat memaksimalkan pemanfaatan serangga ini. Penanganan dan pemanfatan suatu sumber daya alam akan efisien dan efektif jika ditunjang oleh pengetahuan yang lengkap mengenai sumber daya alam tersebut. Dengan demikian, kajian mengenai morfometri, siklus hidup dan kajian molekuler sangat dibutuhkan untuk pengembangan pemanfaatkanC. trifenestratasecara berkelanjutan.
Kajian morfometri dan siklus hidup ulat sutera liar C.trifenestrata dilakukan pada bulan Desember 2008 sampai dengan April 2010. Pengamatan dilakukan pada tiga lokasi disekitar kampus IPB Darmaga, yaitu di Lapangan Parkir Gedung Graha Widya Wisuda (GWW), Kompleks Makam Bara, dan Kandang C Fapet. Pengamatan dilakukan terhadap durasi dan timing siklus hidup, viabilitas setiap tahap perkembangan, morfologi, anatomi, dan pengukuran sembilan parameter morfologi, yaitu berat kokon (BK), berat kokon tanpa pupa (BKTP), panjang kokon (PK) lebar kokon (LP), berat pupa (BP), panjang pupa (PP), lebar pupa (LP), panjang dada (PD), bentang sayap (BTGS), serta faktor fisik dan biologi lingkungan selama penelitian. Data dianalisis secara deskriptif, kecuali data morfometri menggunakan analisis diskriminan. Hasil penelitian menujukkan bahwa C. trifenestrata membutuhkan waktu 51.0 ± 7.3 hari untuk setiap satu siklus hidup, pada siklus hidup I ukuran populasi sangat kecil dibandingkan siklus hidup II. Viabilitas terbesar terdapat pada fase pupa dan viabilitas terkecil terdapat pada fase larva instar I. Kematian terbesar disebabkan oleh kekurangan pakan atau predator.
Semua karakter morfometri berpengaruh nyata terhadap pengelompokan berdasarkan lokasi sub populasi pada setiap fase perkembangan. Besarnya ketepatan prediksi pengelompokan untuk parameter pupa, kokon dan imago berturut-turut: 93,6% , 83,3%,
karakter morfometri sama antar sub populasi, yang ditunjukkan dengan daerah tumpang tindih pada plot pengelompokan. Pada fase kokon dan pupa terdapat daerah tumpang tidih antara sub populasi Kandang C dengan sub populasi Kompleks Makam Bara, tetapi pada fase imago, ketiga sub populasi memiliki daerah tumpang tindih. Hal ini berarti bahwa beberapa ciri berkembang bersama antar sub populasi, yang mengindikasikan masih adanya aliran gen antar sub populasi. Sub populasi di Kandang C memiliki daerah tumpang tindih yang lebih luas dibandingkan sub populasi lainnya.
Karakterisasi gen fibroin ulat sutera liarC. trifenestrata dilakukan untuk mendeteksi
nukleotida dan asam amino diagnostik pada serangga tersebut. Penelitian dilakukan
melalui tahapan sebagai berikut: DNA total diekstrak dari sampel yang berasal dari sub populasi Kabupaten Bogor, Purwakarta dan Bantul, masing-masing dua individu setiap sub populasi. Sumber DNA total adalah kelenjar sutera. Prosedur ekstraksi dilakukan dengan metode standar dengan beberapa modifikasi (Duryadi, 1993). DNA total yang diperoleh diamplifikasi dengan metode PCR menggunakan primer khusus untuk Saturniidae yang dirancang menggunakan software Primer3. Primer tersebut terdiri atas
primerforwardFFCt 5'-CATAACCATGAGAGTAATAGCC-3’, dan primerreverseRFCt
5'-CTGCTGAGTCTGATCCGTAA-3' dengan produk PCR sebesar 986 bp. Hasil amplifikasi dirunutkan, kemudian dikarakterisasi mengenai jumlah dan tipe nukleotida dan asam amino penyusun protein fibroinnya. Hasil karakterisasi kemudian dibandingkan dengan sekuen gen fibroin dan asam amino ulat sutera lain yang tersedia pada GenBank.
Dari hasil perbandingan tersebut terdeteksi nukleotida dan asam amino diagnostik
(penciri) C. trifenestrata. Runutan nukleotida maupun asam amino selanjutnya
dipergunakan untuk menyusun filogeni kekerabatan antar serangga penghasil serat sutera.
Hasil perunutan diperoleh sekuen gen fibroin parsial yang terdiri atas daerah
pengkode ekson I sepanjang 42 nukleotida (14 asam amino), intron (113 nukleotida) dan sebagian daerah pengkode ekson II sepanjang 831 nukleotida (277 asam amino). Hasil perbandingan dengan spesies lain dari famili yang sama (Saturnnidae) dan famili lain (Bombycidae) menunjukkan bahwa ekson I terkonservasi sesama C. trifenestrata, tetapi sangat beragam dibandingkan dengan spesies lain. Namun demikian sesama Saturniidae masih dalam satu cluster dan berbeda dengan Bombycidae. Nukleotida ke 25 (C), dan ke 35 (T) dari ekson I serta asam amino ke 12-nya (valin) bersifat diagnostik bagi spesiesC. trifenestrata. Nukleotida ekson II mengkode 277 asam amino yang didominasi oleh asam amino alanin (27.08%) dan glisin (21.66%). Alanin membentuk polyalanin dengan motif
berupa: (A)2S(A)2, (A)3SS(A)3, (A)2GSAS(A)3, (A)7
SAAGSSGR-GLGGYDGSVDGGYGSGSGS(A)10, (A)10GSSGRGIGGYDGFVDGGYGSGSGS(A)12,
(A)12GSSGRGLGGYDGWV-DDGYGSGSGS(A)12. Dibandingkan dengan spesies lain,
intron C. trifenestrata, lebih pendek ukurannya yaitu 113 nukleotida. Nukleotidanya sangat beragam antar spesies ulat sutera yang dibandingkan, baik dengan famili sama (Saturniidae) maupun dengan famili berbeda (Bombycidae) dan lebih dari 70% berupa A dan T (ATrich).
Kajian keragaman genetik populasi ulat sutera liar Cricula trifenestrataberdasarkan gen Cytochrome C oxidase subunit I (COI) dilakukan dengan menggunakan sampel yang sama dengan kajian keragaman genetik gen fibroin. Hal yang berbeda hanya pada jumlah sampel dan primer yang dipergunakan untuk amplifikasi. Lima belas sampel yang diperoleh dari tiga Kabupaten yaitu Kabupaten Bogor, Purwakarta dan Bantul diekstraksi DNAnya, kemudian diamplifikasi menggunakan primer yang didesain khusus untuk gen
COI famili Saturniidae, yaitu: primer forward 5'-TGATCAAATTTATAATAC-3' dan
primer reverse 5'-GTAAAATTAAAATATAAAC-3' (Mahendran et al. 2006). Seperti
halnya dengan gen fibroin, amplikon COI dirunutkan dan dikarakterisasi jumlah dan tipe
karakterisasi dibandingkan dengan sekuen gen COI ulat sutera lain yang tersedia pada GenBank. Hasil perbandingan tersebut terdeteksi nukleotida dan asam amino diagnostik
untuk spesies C. trifenestrata. Runutan nukleotida maupun asam amino selanjutnya
dipergunakan untuk menyusun filogeni kekerabatan antar serangga penghasil serat sutera. Hasil perunutan parsial gen COI C. trifenestrata berukuran 595 nukleotida (nt),
yang menyandi 198 asam amino. Tidak ada perbedaan nukleotida intra spesies C.
trifenestrata, akan tetapi menunjukkan 23,7% keragaman dengan ulat sutera lainnya. Nukleotida dari gen ini didominasi oleh timin dan adenine (±70%). Berdasarkan 595 nukleotida tersebut, telah dideteksi 25 situs diagnostik yang dapat dipergunakan sebagai pengenal bagi spesies C. trifenestrata. Dari 198 asam amino yang dirunutkan, hanya 10% asam amino beragam antar spesies ulat sutera famili Saturniidae dan Bombycidae. Asam
amino ke 107 (valin) dan ke 138 (treonin) merupakan asam amino diagnostik bagi C.
trifenestrata.
©Hak Cipta milik IPB, 2011
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan masalah; dan pengutipan tersebut tidak merugikan kepentingan wajar IPB
MORFOMETRI DAN KERAGAMAN GENETIK
ULAT SUTERA LIAR
Cricula trifenestrata
Helfer
(Lepidoptera: Saturniidae)
SURIANA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Biologi Mayor Biosains Hewan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Prof. Dr. Ir. Aunu Rauf, M.Sc.
(Departemen Hama dan Penyakit Tanaman IPB)
Dr. Hari Sutrisno, M.Sc (Entomogi – LIPI)
Penguji pada Ujian Terbuka : Dr. Ir. Sutrisno.
(Entomologi Molekuler-Balai Besar Bioteknologi dan Sumberdaya Genetik Bogor)
Dr. Rika Raffiudin.
HALAMAN PENGESAHAN
Judul Disertasi : Morfometri dan Keragaman Genetik Ulat Sutera LiarCricula
trifenestrataHelfer (Lepidoptera: Saturniidae)
Nama Mahasiswa : Suriana
Nomor Pokok : G362070031
Mayor : Boisains Hewan
Disetujui, Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA Ketua
Prof. Dr. Ir. Ronny Rachman Noor, M.Rur, Sc Dr. Ir. Achmad Machmud Thohari, DEA
Anggota Anggota
Mengetahui
Ketua Mayor Biosains Hewan Dekan Sekolah Pascasarjana IPB
Dr. Bambang Suryobroto Dr. Ir. Dahrul Syah, M.Sc.Agr.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini adalah ulat sutera liar Cricula trifenestrata Helfer. Disertasi ini memuat 3 Bab yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal ilmiah. Deskripsi
Morfologi, Anatomi dan Perilaku Ulat Sutera Liar Cricula trifenestrata Helfer
(Lepidoptera: Saturniidae) telah diterbitkan (Jurnal Biologi Lingkungan 4:124-132; ISSN 1978-3736), Characterization of Coding Region Fibroin Gene on Wild Silk MothCricula trifenestrata Helfer (Lepidoptera:Saturniidae), telah diterbitkan pada Jurnal Media Peternakan vol. 34, No. 1: 23-29 (Terakreditasi B SK DIKTI No. 43/DIKTI/Kep/2008). Karakterisasi Relung Populasi Ulat Sutera Liar Cricula trifenestrata Helfer (Lepidoptera: Saturniidae) di Wilayah Darmaga sedang menunggu penerbitan di Jurnal Entomologi.
Selama penelitian hingga penyelesaian disertasi, penulis mendapatkan bantuan dari berbagai pihak. Pada kesempatan ini penulis menyampaikan terima kasih sedalam-dalamnya kepada Bapak Dr. Ir. Dedy Duryadi Solihin, DEA, Bapak Prof. Dr. Ir. Ronny Rachman Noor, M. Rur, Sc., dan Bapak Dr. Ir. Achmad Machmud Thohari, DEA. selaku komisi pembimbing yang telah mendukung, membimbing dan meluangkan waktu serta pikiran sejak perencanaan penelitian, pelaksaan penelitian hingga penulisan disertasi ini selesai.
Terima kasih juga penulis sampaikan kepada Dekan Fakultas MIPA dan Rektor UNHALU Kendari yang telah memberikan ijin tugas belajar, serta Tim Manajemen Beasiswa Program Pascasarjana Dirjen Dikti Depdiknas yang telah memberi dana pendidikan S3. Terima kasih kepada Rektor IPB, Dekan Pascasarjana IPB, Ketua Mayor Biosains Hewan serta seluruh staf pengajar dan administrasi BSH atas bantuannya sehingga penulis dapat menyelesaikan pendidikan S3 di Sekolah Pascasarjana IPB.
Terima kasih penulis tujukan kepada kepala dan staf Laboratorium Biologi Molekuler PAU; Pak Heri dan teman-teman di Lab PAU: Bu Fahma, Bu Nungki, Bu Butet, Bu Fifi, Bu Wahyu, Bu Rini, Bu Irma, Bu Nanna (alm), Bu Septi, Bu Andri, Pak Jusak, Pak Tri, Pak Rahman.
Terima kasih kepada kepala dan staf Laboratorium Genetika Molekuler Ternak Fapet IPB atas segala fasilitas dan bahan yang dapat penulis gunakan selama amplifikasi DNA dan pengamatan serat sutera, kepada Bapak Prof. Dr. Ir. Ronny Rachman Noor, M. Rur, Sc., Prof. Dr. Ir. Muladno, MSA, Prof Dr. Ir. Cece Sumantri, M. Agr.Sc., Eryk Andreas, S.Pt, M.Si, Restu Miranti, S.Pt, dan teman-teman di Lab. GMT, Andi Baso Malompengen, Pak Ihsan, Surya. Terima kasih kepada Drs. Sakka, M.Si, Dr. Muzuni, M.Si, teman-teman di BSH: Bu Tetri, Bu Irma, Mbak Kanti, Bu Ayu dan teman-teman yang tidak saya sebut satu persatu, atas segala bantuan, dan ukhuah yang terbina selama ini.
Akhirnya, kepada keluarga tercinta, anak, suami, dan keluarga besar, penulis menyampaikan terima kasih atas pengorbanan, pengertian, dorongan dan doanya yang tak pernah putus. Semoga karya ilmiah ini bermanfaat bagi pengembangan ilmu.
RIWAYAT HIDUP
xi
xii
KERAGAMAN GEN CYTOCHROME C OXIDASE (COI)
ULAT SUTERA LIARCricula trifenestrataHelfer (Lepidoptera: Saturniidae)
Abstrak . . . 81
Pendahuluan . . . 81
Materi dan Metode . . . 83
Hasil . . . . Pembahasan . . . 90
Simpulan Saran . . . . . . . . 93 93 PEMBAHASAN UMUM . . . 94
SIMPULAN DAN SARAN . . . 98
DAFTAR PUSTAKA . . . 99
xiii
6 Rata-rata durasi siklus hidup dan viabilitasC. trifenestrata . . . . 38
7 Hasil uji kesamaan rata-rata kelompok . . . .. . . . 41
8 Fungsi diskriminan, Eigenvalue, prosentase keragaman, dan korelasi canonical sub populasiC. trifenestrata . . . . 42
9 Matriks korelasi absolute nilai parameter yang diamati pada populasi C. trifenestrata . . . 43
10 Hasil pengelompokan populasi C. trifenestrata berdasarkan asal populasi dengan parameter kokon, pupa, dan imago . . . 44
11 Posisi runutan nukleotida ekson I pada C. trifenestrata,Antheraea danB. mori . . . 64
12 Jumlah perbedaan nukleotida ekson I gen fibroin C. trifenestrata, Antheraea, danB. mori. . . 64
13 Situs asam amino berbeda yang dikode oleh ekson I gen fibroinC. trifenestrata, Antheraea, danB. mori . . . . 65
14 Jumlah perbedaan nukleotida ekson II gen fibroin C. trifenestrata, Antheraea, danB. mori . . . . 67
15 Matriks perbedaan asam amino ekson II gen fibroin C. trifenestrata, AntheraeadanB. mori . . . . 68
16 Prosentase asam amino utama yang dikode oleh ekson II gen fibroin C. trifenestrata, AntheraeadanB. mori . . . . 68
xiv
19 Perbandingan nukleotida dan asam amino ekson 1 pada C.
trifenestrata, AntheraeadanB. mori . . . .
20 Variasi panjang intron gen fibroin pada Saturniidae dan Bombycidae 78
21 Matriks perbedaan nukleotida COI (595 nt) antara C. trifenestrata dengan COI dari dataGenBank . . . .. . . . .
22 Situs diagnostik antar spesies berdasarkan 595 nukleotida gen COI C. trifenstrata . . . .. . 86
23 Jarak genetik antara C. trifenestrata dengan spesies dari data
Genbank,berdasarkan runutan 595 nukleotida COI . . . .
24 Situs asam amino berbeda pada gen COI C. trifenestrata dan asam
amino dataGenBank . . . .. . . 88
25 Jumlah dan prosentase asam amino COI yang berbeda antar spesies
Matriks jarak genetik berdasarkan runutan asam amino COI . . . .
89
89 26
86
xv
DAFTAR GAMBAR
Halaman
1 Alur penelitian . . . 9
2 Imago jantan dan betinaC.trifenestrata . . . 11
3 Kerajinan dengan bahan dasar dari kokonC.trifenestrata . . 11
4 Larva dan pupaC.trifenestrata . . . 11
5 SebaranC.trifenestrata di Asia . . . 12
6 Kokon ulat sutera . . . 13
7 Kelenjar suteraB. mori (Mondalet al2007) . . . 14
8 Skema organisasi asam amino pada protein fibroin B. mori 14 9 Struktur gen fibroinB. mori . . . .. . 15
10 Skema posisi protein fibroin dan protein sericin . . . 18
11 Skema kedudukan bagian amorf danB-sheet . . . 19
12 Serat suteraB. mori . . . .. 19
13 Mekanisme metamorphosis serangga holometabola . . . 20
14 Pengaruh faktor lingkungan terhadap plastisitas fenotipik 20 15 Pengaruh kualitas makanan terhadap keturunan . . . 22
16 Beberapa sifat dasar norma reaksi . . . 24
17 Lokasi pengambilan sampel . . . 32
18 Tekhnik pengukuran . . . 35
19 Perilaku peletakan telur C. trifenestrata . . . 38
20 Ringkasan pembentukan kokon padaC. trifenestrata . . . 39
21 Eklosi padaC. trifenestrata . . . 40
xvi
23 Lokasi Pengambilan sampel . . . 57
24 SEM . . . 57
25 Skema gen fibroin utuh A. pernyi(9282 bp) . . . 59
26 Pola serat penyusun kokon . . . 61
27 Perbandingan serat kokon C. trifenestrata . . . 61
28 Serat sutera hasil degumming . . . 62
29 Kelenjar suteraC. trifenestrata . . . 62
30 Produk PCR gen fibroin . . . 63
31 Skema posisi primer FfCt dan RfCt pada fragmen gen . . . 63
32 Filogeni berdasarkan 42 runutan nukleotida ekson I . . . 65
33 Filogeni berdasarkan runutan 14 asam amino . . . .. . . 66
34 Filogeni berdasarkan runutan nukleotida ekson II . . . 67
35 Filogeni berdasarkan runutan asam amino . . . 69
36 Filogeni berdasarkan runutan nukleotida total ekson I . . . 70
37 Filogeni berdasarkan runutan intron gen fibroin . . . 72
38 Perbandingan kelenjar sutera . . . 73
39 Peta linier mtDNAA. pernyiutuh sepanjang 15.566 bp . . . 83
40 Profil DNA COIC. trifenetratahasil amplifikasi dengan primer RCOI dan FCOI (Mahendran 2005) . . . 85
41 Skema penempelan primer FCOI dan RCOI pada sekuen gen COI C. trifenestrata . . . 85
42 Filogeni berdasarkan runutan 595 nukleotida COI . . . 87
xvii
DAFTAR LAMPIRAN
Halaman
1 Data iklim di kawasan Darmaga . . . . .. . . 108
2 Posisi penempelan primer FFCt dan RFCt . . . 110
3 Pensejajaran berganda 904 nukleotida ekson II gen fibroin . . . 111
4 Komposisi nukleotida ekson II gen fibroin. . . 116
5 Pensejajaran berganda 317 asam amino ekson II gen fibroin. . .. . 117
6 Pensejajaran berganda nukleotida intron gen fibroin . . . 121
7 Posisi penempelan primer FCO I dan RCOI pada . . . 122
8 Pensejajaran berganda 595 nukleotida COI . . . 123
9 Komposisi asam amino COI . . . 129
PENDAHULUAN
Latar Belakang
Serangga dari famili Bombycidae dan Saturniidae merupakan serangga (ngengat) yang menguntungkan (beneficial insect), sebab serangga tersebut menghasilkan kokon sebagai sumber serat sutera alam. Terdapat sekitar 400-5000 spesies ngengat penghasil sutera, tetapi hanya delapan spesies yang telah dimanfaatkan secara komersial, dan yang telah
didomestikasi hanya Bombyx mori (famili Bombycoidae). Ngengat B. mori
ini telah menghasilkan 99% dari sutera di dunia (Dingle et al.2005). Hanya 1% serat sutera lainnya yang dihasilkan oleh beberapa spesies ulat sutera liar famili Saturniidae, yaitu; Antheraea mylitta (tasar silkworm) dan A. assama (Muga silkworm), A. roylei dan Samia cinthya ricini endemik di
India (Mahendran et al. 2006; Arunkumar et al. 2006), A. yamamai
(Japanese oak silkworm) endemik di Jepang (Tamuraet al.1987), A. pernyi (Chinese oak silkworm) endemik di Cina (Liu et al. 2008), sedangkan A. proylei merupakan hybrid dari A. pernyi dan A. roylei, terdapat di India (Arunkumar et al. 2006; Kakati & Chutia 2009), Attacus atlas (fagaria silkmoth) terdapat di India dan Indonesia (Kakati & Chutia 2009; Mahendran et al. 2006; Kalshoven 1981), dan Cricula trifenestrata yang tersebar di kawasan Asia (Nassigat al.1996; Kakati & Chutia 2009).
Usaha domestikasi terhadap ulat sutera terkait dengan kesadaran, kepentingan dan kebijakan pelakunya. Di India misalnya, ulat sutera liar dipelihara di habitat aslinya dengan pertimbangan lebih murah dan jika tertangani dengan baik, maka dapat mendukung usaha konservasi (Reddy et
al. 2010). Kendala yang dihadapi adalah hama dan penyakit yang belum
tertangani (Kakati & Chutia 2009). Sementara itu, di Australia kebijakan
untuk mengesplorasi penggunaan B. mori sebagai penghasil sutera
didasarkan pertimbangan banyaknya pilihan strain yang dapat
dikembangkan dan disesuaikan dengan kondisi alam Australia (Dingle et al. 2005)
Umumnya Saturniidae terdapat pada hutan tropis dan subtropis, dengan keragaman terbesar di daerah tropis Dunia Baru (Amerika Utara)
dan Mexico (Tuskes et al. 1996). Nassig et al. (1996) telah mempublikasikan Saturniidae Indonesia, khususnya yang ditemukan di Sumatera. Famili Saturniidae yang ditemukannya meliputi delapan genus yang terdiri dari 25 spesies, yaitu genus: Attacus, Archaeoattacus, Samia, Actias, Cricula, Anthereae, Leopa, dan Lemaireia. Sejauh ini jenis dan sebaran genus Cricula di Asia adalah tiga spesies ditemukan di Vietnam; yaitu C. trifenestrata Helfer 1837, C. jordan Bryle 1944, dan C. vietname nsp (Nassig & Naumann 1999). Di Malaysia Barat ditemukan tiga spesies, yaituC. elaeziaJordan 1909,C. cameronensis U.&L.H.Paukstadt, 1998 dan C. trifenestrata Helfer 1873. Di pulau Andamanica India, C. andamanica Jordan 1909, merupakan salah satu spesies Cricula endemik. Spesies lain yang ditemukan di daerah Mindanao (Philipina), sebelumnya diidentifikasi
sebagai kelompok spesies C. elaezia Jordan ternyata adalah C.
mindanaensis. Spesies ini berhubungan dekat dengan C. quinquefenestrata Roepke, yang ditemukan di Pulau Sulawesi tetapi berbeda alat kelamin jantannya. Selain itu, sub spesies yang juga ditemukan di daerah Mindano adalahC. luzonica kardi ssp.nov(Nassiget al.1996).
Spesies Cricula yang ditemukan oleh Nassig et al. (1996) di
Indonesia ada tiga, yaitu:C. sumatrensis Jordan,C. elaezia Jordan, dan C. trivenestrata javana Watson. Sejauh ini belum ada publikasi khusus
mengenai C. trifenestrata di Pulau Jawa, namun Kalshoven (1981) telah
C. trifenestrata, dikenal sebagai ulat kipat atau ulat alpukat. Habitat asli serangga tersebut adalah pohon inang seperti alpukat, jambu mete,
mangga, dan kayu manis (Ali & Karim 1991; Wikardi et al.1996;
Syafaruddin & Rahmatia 1999). C. trifenestrata memiliki keistimewaan
pada kokonnya. Berbeda dengan spesies Saturniidae lainnya, kokon C.
trifenestrata tidak tertutup rapat, tetapi membentuk jala, sehingga dapat langsung diproses menjadi barbagai macam aksesoris misalnya bros, kap lampu, aksesoris sepatu atau tas dan lain-lain. Apabila kokonnya dipintal menjadi benang, maka benang yang dihasilkannya lebih halus dan berwarna khas, yaitu warna kuning keemasan. Oleh karena itu, produk sutera dari serangga ini eksklusif dan harganya cukup tinggi.
Meskipun distribusinya luas, dan telah dimanfaatkan untuk produksi serat sutera serta aksesoris, kajian-kajian yang berhubungan dengan ulat alpukat, masih kurang. Hasil analisa karyotipe, diketahui serangga ini memiliki kromosom haploid sebanyak 31 buah (Narang & Gupta 1979). Kajian lainnya hanya terbatas pada usaha menekan laju pertumbuhan populasinya pada tanaman pertanian khususnya jambu mete ( Ali & Karim 1991; Wikardi et al. 1996; Syafaruddin & Rahmatia 1999), padahal dilain
pihak outbreak populasi ulat alpukat dapat dikendalikan dengan cara
memanfaatkannya sebagai sumber kokon untuk menghasilkan serat sutera.
Di India, C. trifenestrata lebih dikenal sebagai hama tanaman
mangga (devoliator mango) (Sheikh 2004; Rono et al. 2008). Meskipun
demikian, C. trifenestratatelah dimanfaatkan dalam industri rumah tangga untuk penghasil serat sutera dan telah dikaji beberapa aspek biologinya (Kakati & Chutia 2009). Hal yang sama juga telah dilakukan oleh beberapa kalangan di Indonesia, terutama padaC. trifenestratadengan tanaman inang jambu mete. Penelitian serupa sangat dibutuhkan, sehingga paradigma sebagai hama dapat berubah menjadi serangga menguntungkan (beneficial insect).
ini terbukti dengan ditemukannya beberapa spesies yang bersifat endemik di beberapa pulau di Indonesia, maka dengan penanganan yang baik
diharapkan dapat meningkatkan produktivitasnya; (b) kerusakan pada
tanaman (bersifat defoliator) dapat memicu pertumbuhan generatif, sebab C. trifenestrata memakan daun tanaman muda maupun tua. Setelah daun habis, tanaman terpicu untuk menghasilkan pucuk baru hanya dalam waktu sekitar dua minggu saja, selanjutnya setelah tanaman menjadi gundul, maka munculnya bunga dan buah lebih cepat; (c) kontinuitas dan produktivitas sebagai hewan budidaya, seekor betina dapat menghasilkan telur ±
300/siklus reproduksi dan untuk mendapatkan kokon hanya diperlukan
waktu 40 hari saja. Jika potensi ini dapat dimaksimalkan dalam
menghasilkan benang sutera, maka paradigma sebagai hama, berubah
menjadi serangga menguntungkan; (d) kualitas serat sutera yang khas, serat
yang dihasilkan oleh C. trifenestrata memiliki warna alami (kuning
keemasan) sehingga tidak memerlukan pewarna tambahan.
Serat sutera yang dihasilkan oleh C. trifenestrata sebagai hasil ekspresi gen fibroin. Kajian genetika molekuler dari gen fibroin ini belum banyak dilakukan (GenBank2010). Oleh karena itu karakterisasi molekuler dari gen fibroin akan dapat membantu dalam jangka panjang untuk usaha rekayasa dan perbaikan genetik dari sutera yang dihasilkannya.
Populasi alamiah mengalami lingkungan yang bervariasi menurut
ruang dan waktu. Fenotip yang diekspresikan oleh suatu organisme
merupakan produk genotip, lingkungan yang dihadapi dan interaksi antara genotip dan lingkungannya (P=G+E+GE) (Noor 2008). Oleh karenanya, keragaman dalam populasi dapat dikatakan sebagai manifestasi keragaman variasi lingkungan (Perotti et al. 2006). Lingkungan yang bervariasi dan ekstrim merupakan stimulator bagi organisme untuk mengekspresikan kemampuan genotipnya dalam menghadapi lingkungan tersebut.
apabila tidak mampu mengatasi stres akan berakibat fatal yaitu kematian pada level individual dan kepunahan pada level populasi (Badyaev 2005). Menghadapi stres lingkungan, organisme melakukan strategi menghidar atau tetap pada lingkungan tersebut dengan segala konsekuensinya. Beberapa organisme mampu menghadapi stres ini, yaitu dengan jalan merubah strategi fisiologi, morfologi maupun perilaku (Noor 2008). Bergantung pada jenis stres lingkungan, organisme akan menghadapinya dengan strategi yang bermacam-macam. Jika stres lingkungan disebabkan oleh faktor biotik, maka beberapa perubahan terutama perubahan perilaku akan dilakukan sesuai dengan interaksinya dengan faktor biotik tersebut, ada yang bersifat mutual, parasit, atau pemangsa dan mangsa (Badyaev 2005). Sebaliknya jika stres lingkungan berasal dari faktor abiotik, umumnya respon organisme akan berupa perubahan fisiologi yang dimanifestasikan dengan perubahan fenotip, misalnya memperpendek waktu tahap tertentu dalam siklus hidup sebagai strategi bertahan dan melanjutkan tahap kehidupan selanjutnya (Whitman & Agrawal 2009).
Sejak dilaksanakannya Jogja International Silk Ekshibition and Conference di Jogjakarta pada tgl 23-26 April 2002, yang dilanjutkan dengan Jogja Expo Center, maka penggunaan serat sutera yang berasal dari
sutera liar alam (A. atlas dan C. trifenestrata) makin berkembang,
setidaknya bagi kalangan tertentu yang berkecimpung di dunia persuteraan. Hal ini berlanjut dengan ditetapkannya Desa Karang Asem Bantul sebagai Eco Tourims. Sutera Liar. Atas kerjasama masyarakat pencinta sutera dengan Garuda Air Lines, ditetapkan sumbangan untuk penanaman satu pohon dari setiap satu penumpang. Sampai sekarang daerah tersebut telah memanfaatkan lahan yang dulunya tandus, menjadi tempat ‘pemeliharaan’ sutera liar dengan tanaman jambu mete, sirsak, dan mahoni sebagai sumber pakan. Dengan demikian, telah terjadi pergeseran paradigma insekta dalam hal ini ulat sutera liar sebagai hama menjadi insekta bermanfaat sebagai penghasil serat sutera. Salah satu kelebihan serat sutera yang dihasilkan dari
spesies ini adalah warnanya yang kuning keemasan. Dengan berbagai
yang lebih tinggi dibanding serat sutera lainnya. Melalui PT Yarsilk Gora Mahottana, benang maupun kokon dari A. atlas dan C. trifenestata telah diekspor ke Jepang. Meskipun demikian ekspor ini masih sangat kecil, yaitu 10 kg sekali ekspor. Satu kg benangA. atlas dihargai sebesar Rp.1,5 juta,
dan 1kg benang C. trifenestrata dihargai Rp.1,7 juta. Kokon A. atlas
seberat 1 kg dihargai Rp 65.000 dan 1kg kokonC. trifenestratadihargai Rp 70.000 (BAPEDA PEMDA DIY 2007).
Database taksa sebagai referensi untuk identifikasi maupun konservasi hewan akan sangat bermanfaat jika metode identifikasinya akurat. Marka genetik dapat dipergunakan untuk memenuhi tujuan tersebut. Database tersebut belum tersedia sampai sekarang, baik untuk spesies
maupun genus Cricula (Nassig et al. 1996; GenBank 2010). Gen penyandi
cytochrome C sub unit 1 (COI) telah diusulkan sebagai DNA barkode hewan (Hebert et al. 2003a, b). Hal ini didasarkan oleh kenyataan bahwa gen tersebut memiliki laju evolusi yang cepat dibandingkan gen inti (Mahendran el al. 2006), sehingga setiap organisme diyakini memiliki gen COI yang unik dan bervariasi untuk masing-masing organisme. Marka genetik ini telah berhasil dipergunakan sebagai alat barcoding untuk ordo Lepidoptera (Hajibabaei et al.2005), ngengatDiatraea saccharalis (Bravo et.al.2008), beberapa spesies ikan, burung dan mamalia (Hebertet.al2004).
Tujuan Penelitian
1. Mengkaji siklus hidup C. trifenestrata di alam dan berapa kali siklus tersebut berulang dalam setahun untuk memperoleh data mengenai durasi dan timingsiklus hidupC. trifenestrata.
2. Melakukan karakterisasi serat kokon dan serat fibroinC. trifenestrata. 3. Melakukan karakterisasi morfologi C. trifenestrata dari populasi asal
yang berbeda, untuk memperoleh data variasi morfologi antara sub populasi.
4. Mendeskripsikan keragaman gen fibroin pada C. trifenestrata dan
5. Mendeskripsikan keragaman gen COI pada C. trifenestrata, dan membandingkan dengan gen COI pada serangga penghasil sutera
lainnya, sehingga diperoleh marka pengenal spesies bagi C.
trifenestrata.
Manfaat Penelitian
1. Studi komparatif dan seleksi populasi terbaik dalam proses domestikasi, dimungkinkan berdasarkan informasi siklus hidup, morfometri, dan faktor-faktor yang mempengaruhi siklus hidupC. trifenestrata.
2. Informasi mengenai sekuen gen dan asam amino fibroin dapat dijadikan sebagai dasar untuk seleksi populasi dalam hubungan dengan produksi serat sutera dan pengenal spesiesC. trifenestrata.
3. Informasi mengenai keragaman genetik dengan penanda COI dapat
dipergunakan sebagai pembeda antar spesiesC. trifenestrata.
Kerangka Pemikiran
Keragaman genetik merupakan variasi genetik dalam spesies, yang mencakup variasi genetik di dalam populasi yang sama atau antar populasi yang berbeda. Variasi tersebut merupakan variasi internal, yang terjadi akibat mutasi gen dan rekombinasi, dan variasi akibat adaptasi terhadap
sebaran georgafis, perbedaan preferensi terhadap tanaman inang, dan
kondisi lingkungan eksternal lainnya. Variasi genetik dalam populasi diperlukan untuk merespon perubahan lingkungan. Makin besar variasi dalam populasi makin besar kemampuan adaptasi populasi tersebut. Oleh karena itu penting untuk mengkaji bioekologi suatu hewan untuk pemanfaatannya secara maksimal dan berkelanjutan.
tersebut dapat peroleh dari ekplorasi genom inti maupun genom mitokondria.
PengembanganC. trifenestratasebagai penghasil serat sutera sangat potensil. Produksi sutera yang berasal dari sutera liar dapat dilakukan melalui pendekatan seleksi berdasarkan kemampuan atau keunggulan
produksi dari masing-masing spesies yang ada. Namun demikian,
nampaknya terdapat dua faktor penting yang sangat menentukan, yaitu faktor genetik dan lingkungan. Faktor genetik berperan menentukan kemampuan kelenjar sutera untuk menghasilkan sutera dengan kualitas tinggi, sedangkan faktor lingkungan terutama makanan dan menejemen pemeliharaan dapat mengoptimalkan kemampuan genetiknya.
Serat sutera diproduksi oleh kelenjar sutera dan pada kelenjar tersebut terdapat gen struktural fibroin yang berperan aktif dan menetukan produksi serat sutera (Agrawal & Gopinathan 1988; Hwang et al. 2001). Struktur maupun keragaman dari gen ini akan dipelajari dan dibandingkan
dengan struktur gen penyandi sutera yang telah ada di GenBank.
Morfologi dan anatomi
Siklus Hidup (Telur - imago)
Gambar 1 Bagan alur penelitian
Pengamatan timing, durasi, dan viabilitasC. trifenestrata,variasi-variasi ukuran morfologi antar sub populasi, apalagi data molekuler baik dari gen inti maupun dari gen mitokondria)
Dasar Seleksi Dalam Usaha Domestikasi Ulat Alpukat (C. trifenestrata)
TINJAUAN PUSTAKA
Morfologi dan Klasifikasi
Cricula trifenestra
Helfer
Cricula trifenestrata merupakan salah satu spesies dari famili Saturniidae. Famili ini ditandai dengan adanya spot; pola cincin konsentrik seperti jendela (fenestra) pada sayapnya (Gambar 2). Meskipun umumnya
Saturniidae menghasilkan sutera, namun berkerabat jauh dengan Bombyx
mori, ngengat penghasil sutera yang telah dikenal selama ini. Saturnaiidae, dalam beberapa tulisan disingkat saturniid, atau ulat sutera liar atau ada yang menyebutnya ulat sutera non-mulberi. Saturniidae merupakan super
famili Bombycidae dengan anggota terbesar, meliputi sekitar 1.861
spesies, 162 genus dan sembilan sub famili (Tuskes el al. 1996). Saturniid
meliputi beberapa ngengat yang banyak dimanfaatkan seperti Antheraea,
Actias, Attacus, Saturnia, dan Cricula (Kakati & Chutia 2009). C. trifenestra akhir-akhir ini mendapat perhatian besar, karena kokonnya yang berwarna khas kuning kemasan sehingga memiliki nilai jual yang lebih tinggi dibandingkan dengan kokon ulat sutera lainnya (BPS 2009). Selain sebagai bahan dasar pembuatan serat sutera, kokon juga merupakan sumber bahan industri lainnya misalnya untuk bahan membuat aksesoris (Gambar 3), bahan dasar kosmetik, bahkan larva dan pupa satuniid pada beberapa tempat merupakan sumber makanan bagi manusia (Mondalet al.2007).
C. trifenestrata mengalami metamorfosis, sehingga ditemukan tiga bentuk tubuh berbeda selama siklus hidupnya, yaitu larva, pupa dan imago (Gambar 2 dan 4). Sebagaimana halnya serangga pada umumnya, tubuh imagoC. trifenestrata dapat dibedakan atas tiga bagian, yaitu kepala-dada-perut. Hampir semua bagian tubuh ditutupi oleh bulu halus. Pada kepala ditemukan mata dan antenna. Dada terdiri atas tiga ruas, masing-masing dilekati oleh sepasang kaki. Pada ruas pertama dan kedua terdapat dua pasang sayap, yaitu sayap depan dan sayap belakang. Perut terdiri atas
sembilan ruas, pada ruas ke-1 sampai ruas ke-6 terdapat enam pasang
lubang trakea (spirakel) sebagai alat pertukaran gas, dan pada ruas terakhir
terdapat saluran pelepasan (telur bagi hewan betina, sperma bagi hewan jantan).
Larva C. trifenestrata juga dibedakan atas: kepala, perut dan dada. Berbeda halnya dengan imago, pada kepala larva ditemukan alat mulut, sebab pada tahap ini larva menghabiskan waktu untuk tumbuh dan berkembang dengan kegiatan makan. Tubuh larva bagian belakang (dada dan perut) ditutupi oleh bulu.
Fenestra (jendela)
B
Gambar 2 ImagoC. trifenestrata jantan (A) dan betina (B) (www.arbec.saturniidae.com;http://www.saturnia.com.)
Gambar 3 Kerajinan dengan bahan dasar dari kokonC. trifenestrataanting (A), cepuk (B) dan kap lampu (C)
(www.houseofcocoon.blogspot.com; www.cis.kit.ac.jp/~kaikoga/randde.htm)
A B C
A B
Gambar 4 Larva(A) dan pupa C. trifenestrata (B)
Berdasarkan cheklist Saturniidae asal Sumatera dan Malaysia (Nassiget al.1996),C. trifenestrata merupakan salah satu dari tiga spesies Cricula yang ada disana. Kedudukan taksonomi C. trifenestrata menurut Triplehorn & Johnson (2005), klasifikasi adalah sebagai berikut:
Kingdom : Animalia
Filum : Arthropoda
Kelas : Insekta
Ordo : Lepidoptera
Super famili : Bombycoidea
Famili : Saturniidae
Sub famili : Saturniinae
Genus :Cricula
Spesies :Cricula trifenestrataHelfer, 1837
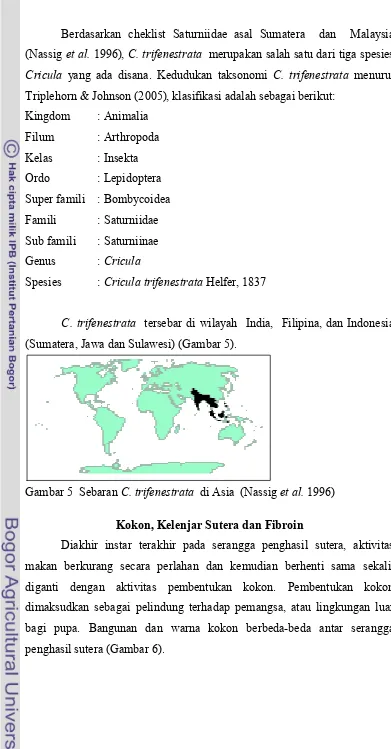
C. trifenestrata tersebar di wilayah India, Filipina, dan Indonesia (Sumatera, Jawa dan Sulawesi) (Gambar 5).
Gambar 5 SebaranC. trifenestrata di Asia (Nassiget al.1996)
Kokon, Kelenjar Sutera dan Fibroin
Diakhir instar terakhir pada serangga penghasil sutera, aktivitas makan berkurang secara perlahan dan kemudian berhenti sama sekali,
diganti dengan aktivitas pembentukan kokon. Pembentukan kokon
Kelenjar labial insekta selalu menghasilkan saliva untuk pencernaan, tetapi pada larva beberapa ordo Tricoptera dan ordo Lepidoptera, kelenjar salivanya mensekresikan fiber atau serat padat yaitu serat sutera yang berfungsi membentuk kokon pelindung pupa, dan ‘tangkai telur’ pada ordo Neuroptera (Sezutsu & Yukuhiro 2000). Oleh karena mensekresikan serat sutera, maka kelenjar tersebut diberi nama kelenjar sutera. Kelenjar sutera telah dikaji secara intensif pada B. mori (Hyodo et al. 1982; Zhou et al. 2000; Mondalet al.2007). Kelenjar ini terdiri atas tiga bagian, yaitu bagian depan, bagian tengah dan bagian belakang (Gambar 7). Bagian depan merupakan tempat dimana serat sutera disalurkan saat pembentukan kokon, bagian tengah kelenjar menghasilkan protein sericin yang lengket dan menyebabkan serat sutera terbungkus oleh semacam lem sedangkan bagian belakang kelenjar menghasilkan rantai berat, rantai ringan dan protein lain yang dirakit menjadi protein fibroin. Protein fibroin tersebut merupakan bahan dasar pembentukan serat sutera (Zurovec et al.1998; Sezutsu & Yukuhiro 2000). Dari bentuk dan peranannya yang khas ini, serta mudah dipelajari dan dipakai oleh masyarakat ilmiah maupun mahasiswa, beberapa
ahli menyarankan kelenjar sutera dapat dijadikan suatu model untuk
mempelajari biologi sel maupun genetika molekuler. Secara khusus kelenjar sutera dipakai sebagai model dalam hal pengaturan ekpresi gen pada tahap perkembangan tertentu. Kelenjar sutera ini disusun oleh sel dengan inti poliploid. Inti poliploid dihasilkan dari sintesa DNA berulang kali tanpa diikuti pembelahan sel (Agrawal & Gopinathan 1988).
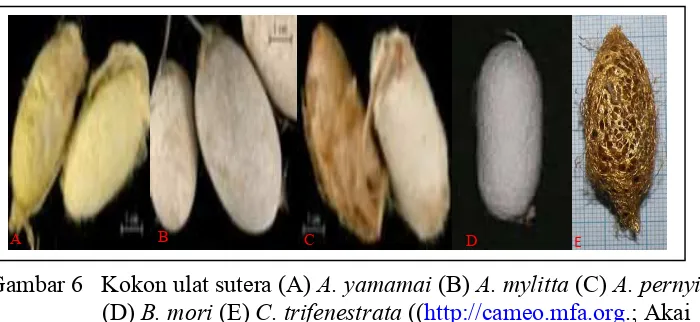
Gambar 6 Kokon ulat sutera (A)A. yamamai(B)A. mylitta(C)A. pernyi (D)B. mori(E)C. trifenestrata((http://cameo.mfa.org.; Akai 2002).
Selain B. mori, serat sutera juga dihasilkan oleh beberapa spesies ulat sutera liar, famili Saturniidae, yaitu dari genusAntheraea, Attacus dan Cricula. Anggota Genus Saturniidae penghasil sutera hingga saat ini belum berhasil dibudidayakan atau dikenal sebagai ulat sutera liar. Dari data yang ada ternyata beberapa spesies memiliki variasi fenotipik dari sutera yang dihasilkannya dalam hal warna maupun teksturnya. Variasi yang terjadi kemungkinan disebabkan oleh karena komposisi penyusun material utama dari serat sutera tersebut, yaitu protein fibroinnya (Gambar 8).
Gambar 8 Skema organisasi asam amino pada protein fibroin B. mori.
Jarak antara lapisan bervariasi bergantung pada banyaknya residu asam amino glisin atau alanin (A). Komposisi kimia asam amino penyusun protein fibroin (B) (Nelson & Cox 2003).
Pada famili Bombicoidae, fibroin disusun oleh rantai berat (H-fib), rantai ringan (L-fib), dan P25, sedangkan famili Saturniidae hanya disusun oleh dimerH-fib(Mondal et al.2007;Sehnal & Sutherland 2008; Suzetzuet al. 2008). Perbedaan antara gen fibroin pada B. mori dan gen fibroin pada
kelompok Saturniidae hanya pada komposisi dan jumlah pasang basa
penyusunnya ( Tabel 1). Oleh karena itu, serat sutera yang dihasilkan oleh B. mori berbeda dengan serat sutera yang dihasilkan oleh ulat sutera liar (Famili Saturniidae).
Gambar 7 Kelenjar suteraB. mori A-B : bagian depan, B-C : bagian tengah, C-D : bagian belakang (Mondalet al.2007)
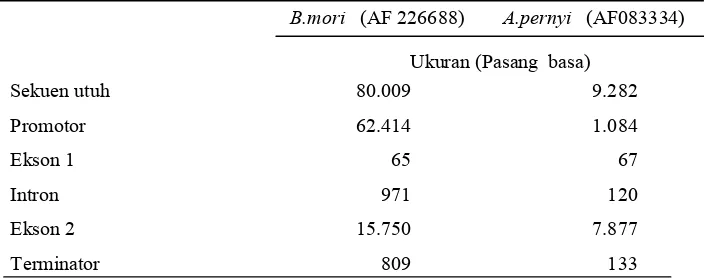
Tabel 1 Struktur gen fibroin padaB. mori (Zouet al. 2000) dan A. pernyi (Sezutsu & Yukuhiro 2000)
Protein fibroin merupakan hasil ekpresi gen H-fib dengan struktur yang relatif sama antara Bombicoidae dengan Saturniidae. Gen fibroin B. moriterdapat pada kromosom 23 (Fangzhouet al.2002). Gen ini terdiri dari
2 ekson dan 1 intron. Struktur gen fibroin B. mori tersusun secara
skematis pada Gambar 9.
+1 s/d +65 +1037
62.414 bp 971bp 15.750 bp
5’
Pensejajaran sekuen gen fibroin B. mori (Bombicidae) dan A.
yamamai (Saturniidae) menunjukkan bahwa daerah pengapit pada ujung 5’ (5’ flanking region) dan ekson I antara kedua spesies ini memiliki homologi yang tinggi (>90%), dan daerah pengapit ini dikenal sangat terkonservasi. Sama halnya dengan A. pernyi, ekson I pada kedua spesies
hanya mengekspresikan 14 asam amino. Namun komposisi nukleotida
pada daerah intron dan ekson II antara ketiga spesies tersebut (B. mori, A. pernyi, dan A. yamamai) sangat berbeda (Sezutsu & Yukuhiro 2000; Zhou et al2000; Hwanget al.2001).
Asam amino penyusun protein fibroin A. pernyi berjumlah 2.639.
Asam-asam amino tersebut didominasi oleh asam amino glisin (27.283%) B.mori (AF 226688) A.pernyi (AF083334)
Ukuran (Pasang basa)
Gambar 9 Struktur gen fibroin B. mori (No akses pada GenBank AF
226688); P: promotor, E1 : Ekson 1, E2 : Ekson 2 dan T:
dan alanin (43.085%). Asam amino alanin (A) membentuk blok polyalanin, mulai dari tiga sampai 13 residu asam amino alanin. Sedangkan asam amino glisin (G) berikatan dengan asam amino lainnya yang juga tersusun secara berurutan dan mengalami pengulangan (Tabel 2).
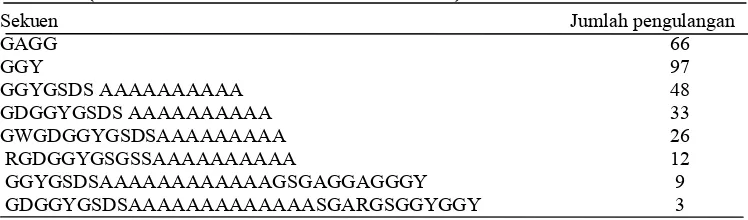
Tabel 2 Beberapa sekuen berulang pada asam amino fibroin A. pernyi
(diolah dari Sezutsu & Yukuhiro 2000)
A:alanin, G:glisin, Y:tirosin, D:Asam aspartat, S: serin, R:Arginin.
Protein fibroin B. mori terdiri atas 5.249 asam amino. Berbeda
halnya dengan protein pada fibroin A. pernyi, protein fibroin B. mori
didominasi oleh asam amino glisin (45.886%) dan alanin (30.268%). Selain itu, terdapat perbedaan dalam hal urutan dan susunan pengulangan
asam-asam amino tersebut. Pada protein B. mori susunan dan urutan asam
amino didominasi oleh urutan glisin-alanin (GA), dengan jumlah
pengulangan mulai dari dua sampai dengan delapan GA. Setelah GA maka urutan berulang yang dijumpai adalah GS (glisin-serin), GY (glisin-tirosin) dan GT (glisin-treonin) (Tabel 3).
Tabel 3 Beberapa sekuen berulang pada asam amino fibroinB. mori(diolah dari Zhouet al.2000)
Fibroin adalah glikoprotein yang terdiri atas dua sub unit protein berukuran 370 dan 25 kDa. Fibroin tersebut membentuk suatu filament yang terdiri atas daerah amorfus dan daerah kristalin. Daerah amorfus berbeda dengan daerah kristalin berdasarkan jenis asam aminonya. Pada daerah
amorfus, rantai samping asam amino membentuk bulkier, sedangkan di
daerah kristalin, rantai samping asam aminonya pendek dan memungkinkan
untuk dikemas dalam bentuk B-sheet (Gambar 8) yang mengandung 12%
asam amino alanin, 30% asam amino glisin dan 44% asam amino serin.
Struktur B-sheet dengan B-sheet lainnya tersusun berlawanan arah
(antiparalel) selanjutnya membentuk mikrofibril. Mikrofibril-mikrofibril akan membentuk suatu bundel, dan beberapa bundel akhirnya membentuk satu serat tunggal fibroin (Padamwar & Pawar 2004).
Selain protein fibroin, serat sutera dibungkus oleh protein yang bersifat perekat bagi protein fibroin, sehingga serat menjadi kaku. Protein
ini disebut sericin. Protein sericin mencakup 20-30% dari total berat
kokon dan komposisinya didominasi oleh asam amino serin (32%). Dalam beberapa tulisan, protein sericin disebut protein tipe II, sebab merupakan protein dengan proporsi terbesar kedua sebagai penyusun serat sutera, setelah protein fibroin. Jadi menurut proporsinya, protein fibroin sebagai protein utama (tipe I) dan sericin sebagai protein ke-2 (Padamwar & Pawar
2004; Mondal et al. 2007). Sericin merupakan polipeptida dengan
komposisi dan ukuran asam amino bervariasi. Polipeptida ini diekresikan pada bagian tengah kelenjar sutera B. mori,yang dikode oleh dua gen yang berbeda, yaitu gen ser1 dan genSer2 tetapi terdapat pada kromosom yang
sama, yaitu kromosom 11 (Takasu et al. 2005). Gen ser1 mengkode
polipeptida dengan ukuran 76-330 kDa, sedangkan gen ser 2 mengkode
polipeptida dengan ukuran sekitar 226.5 kDA.
Berdasarkan sifat kelarutannya, maka protein sericin dapat
sama dengan protein sericin A, tetapi dengan penambahan asam amino triptofan. Protein sericin B mengandung 16.8% nitrogen. Protein sericin C terdapat pada bagian paling dalam. Komposisinya sama dengan protein sericin B, tetapi dengan penambahan asam amino prolin, sulfur dan nitrogen. Berbagai modifikasi penamaan protein sericin berdasarkan kelarutannya, tidak berdasarkan pengelompokan A, B dan C, tetapi menjadi kelompok sericin I, II dan III (Padamwar & Pawar 2004).
Satu serat tunggal dari sutera, pada dasarnya disusun oleh dua protein fibroin yang terbentuk melalui ikatan disulfida antar asam amino penyusunnya. Beberapa filament (fibril) akan membentuk suatu berkas filament (bunch of fibrils) yang menempati bagian core dari serat sutera. Berkas filament tersebut selanjutnya dibungkus oleh beberapa lapis protein sericin, yang selanjutnya membentuk serat tunggal sutera (Gulrajani et al. 2011) (Gambar 10).
Berdasarkan kedudukannya dalam pembentukan serat sutera, sericin adalah protein tipe II, mengandung 18 asam amino termasuk asam amino esensil. Asam amino penyusun protein sericin terdiri atas 32% serin, 45,8% asam amino hidroksil dan 12.2 % asam amino polar (Gulrajaniet al.2011). Sama halnya dengan protein fibroin, sericin juga membentuk struktur Β -sheet, yang berada diantara bagian amorf tetapi dapat berubah menjadi bentuk coil dengan jalan menyerap air atau adanya tekanan mekanik, yang menyebabkan susunan serat menjadi rapat atau renggang (Zhuet al.1998). Gambar 11 menyajikan posisi/keadaanΒ-sheetpada satu serat sutera.
Pada pembuatan benang sutera, kokon diperlakuan dengan air panas (dapat dicampur dengan sabun atau bahan alkalis lainnya) untuk
menghilangkan sericin. Proses yang disebut degumming ini menghasilkan
serat sutera yang lebih lembut dan halus. Perbandingan serat sutera B. mori sebelum dan setelahdegummingdisajikan pada Gambar 12.
Metamorfosis
Serangga holometabola (termasuk C. trifenestrata) mengalami
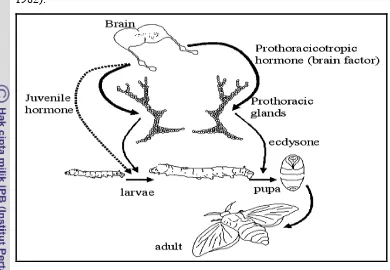
metamorphosis sempurna, yaitu larva berlangsung pada periode tertentu yang diantarai oleh pergantian kulit (molting). Tahap selanjutnya adalah pupa, dan diakhiri dengan tahap imago (serangga dewasa) (Chapman 1982).
Metamorfosis dikendalikan oleh system neuroendokrin (Gambar 13). Ketika larva mengalami ukuran maksimal, maka kelenjar pada otak (corpora alata) terpicu untuk melepaskan hormon prototorasik (PTTH). Selanjutnya PTTH memicu kelenjar prototoraks untuk menghasilkan hormon ekdison. Hormon ekdison mengaktifasi sel epidermis untuk memulai apolisis (terlepas dari sel efidermis) yang diikuti oleh proses ekdisis (tanggalnya kutikula lama). Selain PTTH, kelenjar otak juga melepaskan hormon juvenile (JH) yang berfungsi mempertahankan sifat larva. Pada akhir instar, titer PTTH mengalami puncaknya dan menghambat pelepasan JH. Dengan
Bagianamorf
Fibril sutera
Β-sheet
Gambar 11 Skema kedudukan bagian amorf dan B-sheet pada serat sutera
(www. optic.rochester.edu.)
Gambar 12 Serat suteraB. mori(A) serat mentah (B) Hasildegumming dengan penambahan sabun dan Na2CO3(Sasithorn & Luepong 2009)
B
demikian tidak adanya JH menyebabkan pupasi pada tahap ini (Chapman 1982).
Pengaruh Faktor Lingkungan Terhadap Keragman Fenotipik
a. Munculnya Keragaman Sebagai Respon Adaptif
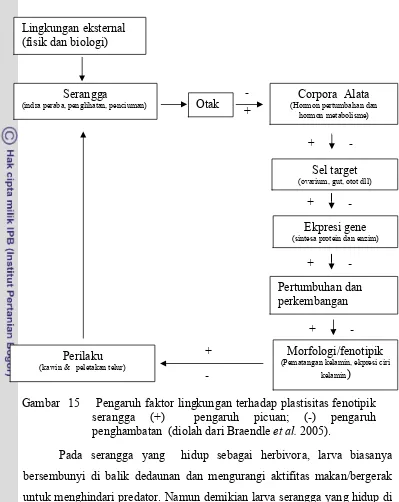
Sebagai suatu sistem, organisme sangat dipengaruhi oleh
lingkungannya. Oleh karena itu, fenotip organisme dapat berbeda sesuai dengan keadaan lingkungannya (Gambar 14). Kemampuan suatu organisme mengekpresikan fenotip yang berbeda pada lingkungan yang berbeda pula dikenal dengan istilah plastisitas fenotipik (Whitman & Agrawal 2009). Ekspresi fenotipik merupakan hasil dari ekspresi genotip, faktor lingkungan dan interaksi antara genotip dan lingkungan (Noor 2008).
Fenotip 1 Fenotip 2 Fenotip n
Gambar 14 Ragam Fenotip dipengaruhi oleh ragam genotip dan lingkungan (Pigliucci & Massimo 2001 ).
Genotip polimorfik (ragam sudah diketahui)
E1 E2 En
Gen-gen yang bersifat plastis dapat berupa gen autosomal, maupun gen terpaut kelamin (Noor 2008; Mikolajewsky et al. 2008). Plastisitas
fenotipik merupakan fenomena yang umum pada serangga, yang dapat
diamati pada ukuran tubuh, warna tubuh, ada tidaknya sayap, warna dan pola sayap, pergantian kasta, ada tidaknya capid, dll (Braendle et al. 2005; Whitman & Agrawal 2009). Plastisitas fenotipik juga teramati pada serangga tertentu meliputi perilaku makan, perilaku kawin, peletakan telur, perilaku anti predator dll.
Pada dasarnya plastisitas fenotipik merupakan proses fisiologi,
tetapi dimanifestasikan sebagai perubahan biokimia, morfologi, perilaku dan life history(Tublitz & Loi 1993; Holbrook & Schal 2004 ; Whitman & Agrawal 2009).
Sinyal yang menentukan adanya plastisitas berasal dari faktor lingkungan. Faktor lingkungan tesebut dapat berupa faktor lingkungan internal atau eksternal, faktor lingkungan fisik/abiotik maupun biotik. Faktor lingkungan internal antara lain: titer hormon atau enzim yang berperan dalam proses fisiologi dan biokimia (Nijthout 2003), beberapa peptida mirip hormon atau enzim, zat bersifat ligand, koenzim dll. Faktor-faktor ekternal antara lain: suhu, kelembaban, gas dan unsur tertentu, makanan dan ketersediaannya, predator, parasitoid dll (Gibert et al. 2007; Braendle et al. 2005). Faktor-faktor tersebut secara sendiri-sendiri ataupun melalui interaksi yang kompleks akan mempengaruhi organisme pada level molekuler sampai level yang lebih tinggi dengan implikasi pada perubahan fenotip dan atau perilaku (Gambar 15).
Pada serangga yang hidup sebagai herbivora, larva biasanya bersembunyi di balik dedaunan dan mengurangi aktifitas makan/bergerak untuk menghindari predator. Namun demikian larva serangga yang hidup di air, untuk menghidari predator mereka mengembangkan kemampuan untuk berenang lebih cepat atau bersembunyi dibalik substrat di bawah permukaan air (Stokset al.2003).
b. Plastisitas Merupakan Strategi Adaptif
Hewan yang terpapar perubahan lingkungan yang ekstrim hanya memiliki 2 pilihan, yaitu mengembangkan plastisitasnya atau tereliminasi dari lingkungan tersebut (Badyaev 2005). Demikian pula hewan yang Lingkungan eksternal
Gambar 15 Pengaruh faktor lingkungan terhadap plastisitas fenotipik
serangga (+) pengaruh picuan; (-) pengaruh
mengembangkan plastisitasnya hanya akan mengekspresikan bentuk alternatif saja (Nijhout 2003).
Plastisitas fenotipik berperan penting dalam kelangsungan hidup organisme. Untuk dapat bertahan hidup organisme mengembangkan plastisitas, meskipun harus dibayar mahal (adatrade off dancost plasticity). Ekspresi sifat atau perilaku yang bersifat plastis membutuhkan biaya yang besar dan harus dibayar ketika hal tersebut berlangsung. Dilain pihak, ekpresi sifat tersebut membutuhkan trade off yang konsekuensinya terjadi pada kehidupan selanjutnya. Oleh karena itu, hewan (serangga) yang telah
didomestikasipun, misalnya ulat sutera B. mori masih memiliki
kemungkinan untuk plastis terhadap faktor-faktor lingkungan dalam batas tertentu, walaupun rentangnya relatif sempit. Plastisitas tersebut meliputi plastisitas terhadap perubahan suhu, kualitas makanan, dan fotoperiode (Iqbalet al.2008). Dengan mengembangkan plastisitas, maka performa ulat sutera (B. mori) yang dikembangkan di Indonesia cenderung kurang baik dibandingkan dengan performa ulat sutera yang dikembangkan di tempat asalnya. Misalnya kokon relatif lebih kecil/ringan sehingga benang sutera yang dihasilkan juga relatif lebih sedikit. Hal tersebut terjadi sebab ulat sutera harus menghabiskan sebagian energi untuk menghadapi perubahan
lingkungan, yang seharusnya energi tersebut dipergunakan untuk
bereproduksi secara maksimal.
Pada ulat sutera liar, plastisitas ‘lebih lebar’ lagi, terutama bagi ulat sutera yang bersifat polifagus. Pada Attacus atlas misalnya, warna kokon
bervariasi sesuai dengan tanaman inangnya. Pada Antheraea mylitta
plastisitas tidak terbatas hanya pada sumber makanan, tetapi juga pada
ketinggian, musim, dan faktor iklim lainnya (Mahendran et al. 2006).
Berbeda halnya dengan ulat sutera yang telah didomestikasi, ekspresi sifat plastis pada ulat sutera liar cenderung bernilai positif, sebab keragaman yang ditimbulkan bernilai ekonomi yang berarti memberi nilai tambah bagi ulat sutera liar tersebut.
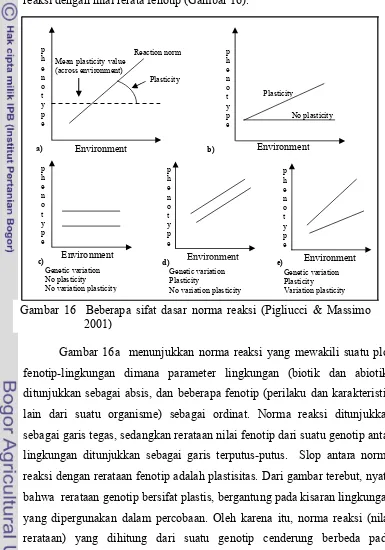
menghubungkan genotip yang terpapar lingkungan tertentu dengan fenotip yang mungkin diekspresikan oleh genotip tersebut (Pigliucci & Massimo 2001). Norma reaksi memiliki beberapa sifat dasar yang disederhanakan melalui garis tegas yang menunjukkan respon linier suatu genotip terhadap variasi lingkungan, sedangkan plastisitas merupakan slop antara norma reaksi dengan nilai rerata fenotip (Gambar 16).
Gambar 16a menunjukkan norma reaksi yang mewakili suatu plot fenotip-lingkungan dimana parameter lingkungan (biotik dan abiotik) ditunjukkan sebagai absis, dan beberapa fenotip (perilaku dan karakteristik lain dari suatu organisme) sebagai ordinat. Norma reaksi ditunjukkan sebagai garis tegas, sedangkan rerataan nilai fenotip dari suatu genotip antar lingkungan ditunjukkan sebagai garis terputus-putus. Slop antara norma reaksi dengan rerataan fenotip adalah plastisitas. Dari gambar terebut, nyata bahwa rerataan genotip bersifat plastis, bergantung pada kisaran lingkungan yang dipergunakan dalam percobaan. Oleh karena itu, norma reaksi (nilai rerataan) yang dihitung dari suatu genotip cenderung berbeda pada Gambar 16 Beberapa sifat dasar norma reaksi (Pigliucci & Massimo
lingkungan yang berbeda atau dengan kata lain tidak ada produk genotip khusus tanpa lingkungan khusus. Secara biologi, rerataan sifat dan plastisitas sifat bisa saja berhubungan atau tidak berhubungan, bergantung mekanisme genetik yang mengontrol ekspresi sifat tersebut. Oleh karena
norma reaksi merupakan suatu fungsi yang mendeskripsikan hubungan
genotip tertentu antara fenotip dengan lingkungan, dan plastisitas adalah atribut dari norma reaksi, maka suatu genotip bisa bersifat plastis dan tidak plastis, tetapi memiliki norma reaksi yang tegas (Gambar 16b). Dengan demikian norma reaksi adalah sifat atau milik populasi, bukan sifat genotip individual.
Gambar 16c, d, dan e menunjukkan perbedaan antara variasi genetik untuk rerata sifat, plastisitas, dan variasi genetik untuk plastisitas. Gambar 16c menunjukkan dua genotip yang memiliki norma reaksi berbeda, keduanya datar, cenderung ke sumbu mendatar (lingkungan) dan karenanya secara harfiah genotip tersebut tidak plastis. Oleh karena tidak plastis, berarti tidak ada variasi genetik untuk plastisitas. Meskipun demikian kedua genotip tersebut berbeda reratanya terhadap lingkungan, satu lebih tinggi terhadap yang lainnya. Jadi terdapat variasi genetik dalam populasi tersebut. Ilustrasi yang berbeda ditunjukkan pada Gambar 16d. Norma reaksi kedua genotip tersebut berbeda, tetapi bersifat plastis. Sifat plastis ditunjukkan dengan model sama (paralel). Jadi terdapat variasi genetik dan plastisitas, tetapi tidak ada variasi genetik untuk plastisitas. Dari sudut pandang
evolusioner, rerata sifat populasi pada gambar 16c dan d, dapat
Penelusuran Keragaman Genetik Hewan
Implikasi biodiversitas khususnya keragaman genetik hewan sangat beragam, mulai usaha konservasi hewan langka, pemuliaan hewan ternak dan hewan liar guna meningkatkan nilai plasma nutfah, aspek lain yang berhubungan dengan kesehatan maupun farmakologi. Nilai plasma nutfah akan meningkat jika dilengkapi dengan data keragaman genetik disamping data keragaman morfologi dan anatominya.
Satu hal yang sangat menguntungkan adalah tersedianya berbagai alat dan metode untuk penelusuran keragaman genetik (penanda genetik), baik hewan maupun tumbuhan. Penanda genetik adalah sekuen DNA yang dapat menunjukkan suatu variasi, yang mungkin disebabkan oleh mutasi atau perubahan materi genetik, dan dapat diamati. Sekuen DNA untuk penanda genetik, dapat berupa sekuen pendek, yang terdapat disekitar pasangan basa tunggal yang berubah (polimorfisme nukleotida tunggal) atau berupa sekuen panjang yang mengkode asam amino tertentu. Penanda genetik yang digunakan pada hewan dan manusia berdasarkan genom mitokondria maupun genom inti. Gen-gen dalam masing-masing genom tersebut telah terbukti banyak digunakan untuk studi evolusi, genetika populasi, penelusuran kekerabatan dan ekspresi gen terkait dengan faktor lingkungan dan faktor-faktor lain yang mempengaruhinya.
Genom mitokondria
Genom mitokondria sering disebut mitogenom, atau disingkat mtDNA berbentuk sirkuler, utas ganda, dengan BM kecil, panjangnya
berkisar 15.000-16.000 bp (Avisa et al. 1987). Pada Antherae pernyi,
ukuran mtDNA lengkapnya adalah 15.566 bp, pada Eriogyna pyretorum
(Lepidoptera: Saturniidae) terdiri dari 15.327 bp. mtDNA yang lebih panjang antara lain ditemukan pada manusia, yaitu 16.569 bp. mtDNA tersebut meliputi 13 gen pengkode protein (PCGs), 2 gen pengkode rRNA,
22 gen pengkode tRNA dan daerah pengontrol. Susunan PCGs
menunjukkan bahwa T lebih banyak dibanding dengan A. Komposisi
kodon ATN kecuali gen cytohrome oxidase (CO) I dan II. Semua gen tRNA memiliki struktur clover-leaf, dengan pengecualiantrnSI(AGN) dan trnS2 (UCN) (Liuet al.2008)
Diantara protein yang dikode oleh gen-gen mitokondria, adalah
protein yang berfungsi sebagai enzim, misalnya cytochrome oxidase ke
1,2,3 (CO I, CO II dan CO III), NADH dehidrogenase (1-6), dan ATP sintetase. Karena struktur dan evolusinya, maka sekuen mtDNA digunakan secara luas sebagai penanda genetik pada studi evolusi. Terdapat beberapa alasan mengapa mtDNA dapat digunakan sebagai marker genetik, antara lain; jumlah copy mitokondria dalam sel atau jaringan cukup melimpah, terutama sel-sel yang aktif dalam proses metabolisme, sehingga amplifikasi mtDNA lebih mudah dibandingkan dengan amplifikasi lokus gen inti.
Selain itu, bagian pengendali bukan pengkode (d-loop) pada molekul
mtDNA memiliki variabilitas tinggi dan secara ekstensif telah
dikarakterisasi pada sejumlah taksa. Dengan demikian mtDNA representatif untuk mempelajari keragaman genetik intraspesies dan interspesies (Luntet
al. 1996; Hebert at al. 2003a). Sebagai penanda genetik, mtDNA mudah
diamplifikasi sebab bersifat haploid sehingga tidak perlu dikloning; mtDNA memiliki laju evolusi yang tinggi, dan jumlah gen yang dimiliki terbatas dibanding genom inti; oleh karena frekuensi rekombinasinya rendah, maka diasumsikan bahwa molekul keseluruhan memiliki genealogikal sejarah yang sama; mtDNA telah direkomendasikan dalam studi taksonomi dengan usulan bahwa semua spesies dideskripsikan adalah sekuen tag mtDNA atau barcode(Hebertet al.2003a).
CO I dari mtDNA sudah dipergunakan sebagai suatu penanda DNA untuk spesies hewan dan sejumlah studi-studi di dalam bermacam taksa (Hebertet al.2004; Hajibabaeiet al.2005; Harmonet al.2006; Hulcret al.
2007; Bravo et al. 2008). DNA barcoding memisahkan kebanyakan jenis
Gen COI merupakan salah satu gen yang dapat digunakan sebagai penanda genetik dalam studi molekuler untuk mempelajari karakteristik genetik antar spesies maupun antar individu. Identifikasi spesies melalui DNA barcode untuk spesies hewan adalah fragmen 648 bp dari gen COI. DNA barcode ini telah sukses mengidentifikasi/membedakan spesies berbagai Invertebrata dan Vertebrata, dari Lepidoptera sampai burung dan dalam geografi yang berbeda (Hajibabaei et al. 2007). Hasil penelitian menunjukkan bahwa sistem identifikasi hewan berdasarkan gen COI cukup efektif. Genom mitokondria dari spesies yang berkerabat dalam satu filum yang dibandingkan menunjukkan diversitas yang cukup untuk membedakan antar hewan tersebut.
Konstruksi Pohon filogeni
Konstruksi pohon filogeni antara lain ditujukan untuk memahami
sejarah evolusioner makhluk hidup, kajian biodiversitas, untuk
memprediksi fungsi kebaharuan gene, pemetaan keragaman strain pathogen untuk pembuatan vaksin, dan tujuan-tujuan lainnya. Beberapa hal yang mesti ditentukan sebelumnya adalah: (1) jenis pohon yang akan dikonstruksi (2) Sekuen yang akan akan dibandingkan (3) data yang harus dihitung.
Sekuen yang akan dibandingkan, biasanya berupa sekuen DNA, RNA atau sekuen asam amino suatu protein, bergantung tujuan pembuatan pohon filogeni. Sekuen tersebut dijajarkan untuk mengetahui similaritas sekaligus variabilitasnya. Tersedia beberapa macam metode penjajaran yang dapat di akses secarafree, tetapi yang umum dipergunakan adalah clustal W (Tamuraet al. 2007).
MORFOMETRI DAN SIKLUS HIDUP ULAT SUTERA
LIAR
Cricula trifenestrata
Helfer (Lepidoptera: Saturniidae)
ABSRACT
The study was conducted to describe morphometric character and life cycle of Cricula trifenestrata exist in 3 locations, namely in the parking area of Graha Widya Wisuda (GWW), Bara Tomb Complex, and Animal House C in the campus IPB Darmaga areas. Observations were done on the life cycle, viability, and measurement of nine parameters, namely cocoon weight (CW), cocoon shell weight (CSW), cocoon lenght (CL) cocoon widht (CWD), pupae weight (PW), pupae length (PL), pupae width (PWD), thorax length (TL), and wingspan (WS). The data were analyzed descriptively, except morphometrical data, conducted by stepwise discriminant analysis method to determine the groupings accuracy on each development stage, and specify the parameters that gave the biggest differentiator for each stage of development. The results showed that C. trifenestrata spent 51.0 ± 7.3 day for each life cycle. The greatest viability was found in the pupae stage, while the smallest viability found in first instar larvae. Most of the deaths caused by lack of food or predators. All morphological parameters significantly affected the grouping by location in each stage of development. The prediction accuracy for the grouping by parameters the cocoon, pupae and imago are: 93.6%, 83.3% and 92.7% respectively with the biggest differentiator each: CW, PW, and WS. The three sub-populations clearly clustered. In the cocoon and pupae stage, sub-populations in the GWW develop its own characteristics, but in the adult phase, several characteristics of developing shared among sub-populations, which was indicated the persistence of gene flow between sub populations.
Key word: Wild silk moth, C. trifenestrata, life cycle, viability, cocon weight, pupae weight, wingspan.
PENDAHULUAN
Kajian mengenai biologi dan karakteristik populasi umumnya dilakukan dengan pengamatan variasi morfologi dan perilaku. Hal ini disebabkan antara lain oleh karena: variasi morfologi secara tegas dapat menjadi pembeda antar individu dan menjadi pengenal spesies, karakter morfologi mengindikasikan fenomena plastisitas (Moczek & Nijhoout 2002; Nijhoout 2003; Withman & Agrawal 2009). Dalam kajian evolusi variasi
morfologi umumnya merupakan fenomena awal isolasi reproduksi
(Vishalakshi & Singh 2008). Fenomena adanya interaksi, baik antar
individu dalam populasi, maupun antar populasi dapat ditunjukkan dengan adanya variasi morfologi. Bahkan variasi tersebut dapat dipicu oleh interaksi antar mangsa dan pemangsa (Nosil & Crespi 2006). Oleh karena itu, populasi yang berkembang pada waktu dan tempat yang berbeda dapat memiliki variasi morfologi yang berbeda pula.
Dari sudut pandang genetika populasi, seleksi alam dan aliran gen dapat mempengaruhi derajat perbedaan populasi alami. Ketika populasi berkembang pada habitat yang berbeda, seleksi alam yang bersifat divergen akan mempengaruhi perbedaan adaptif populasi lokal. Sebaliknya antara populasi yang telah mengalami divergensi, aliran gen justru akan menyebabkan homogenisasi antar populasi (Kawecki & Albert 2004). Dari
uraian di atas, diduga bahwa durasi dan timing siklus hidup juga akan
bervariasi pada ruang dan waktu yang berbeda.
Kajian mengenaiC. trifenestrata masih sangat minim, terbatas pada usaha mengendalikan populasi pada tanaman pertanian ( Ali & Karim 1991; Wikardiet al. 1996; Syafaruddin & Rahmatia 1999). Dilain pihak, terdapat spesies yang bersifat endemik di beberapa pulau di Indonesia (Nassiget al. 1996; Nassig & Nauman 1999). Hal tersebut menunjukkan adanya adaptasi yang besar terhadap lingkungan di Indonesia dan merupakan potensi untuk
pengembangan C. trifenestrata sebagai sumber serat sutera alami. Oleh
karena itu, kajian mengenai morfometri dan siklus hidup serangga ini sangat diperlukan.
Penelitian ini bertujuan untuk: (1) membandingkan durasi dantiming
siklus hidup, (2) membandingkan viabilitas antar sub populasi C.