Identifikasi Dan Ekspresi Gen Protein Selubung Pepper Vein Yellows Virus Pada Escherichia Coli
Teks penuh
Gambar


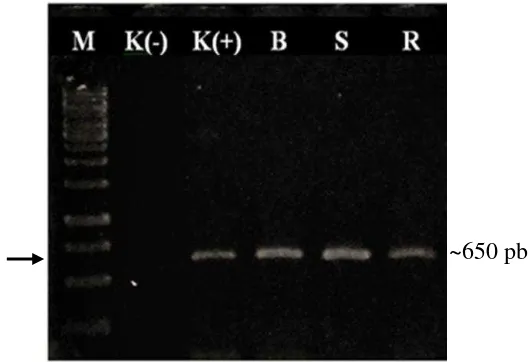




Dokumen terkait
Pada analisis SDS-PAGE, ekspresi protein rekombinan XynA BCRS-01 memiliki bobot molekul protein yang sama dengan yang dilakukan oleh Qu dan Shao (2011) dan Yasinok et
gen TcPIN yang diisolasi dari biji kakao, ekspresi proteinnya tidak menunjukkan aktivitas sebagai inhibitor tripsin, baik pada protein hasil ekspresi secara
Skripsi berjudul “ Identifikasi Profil Protein Daun Mimba ( Azadirachta Indica Juss ) dengan Analisis SDS – PAGE ” telah diuji dan disahkan oleh Fakultas Kedokteran
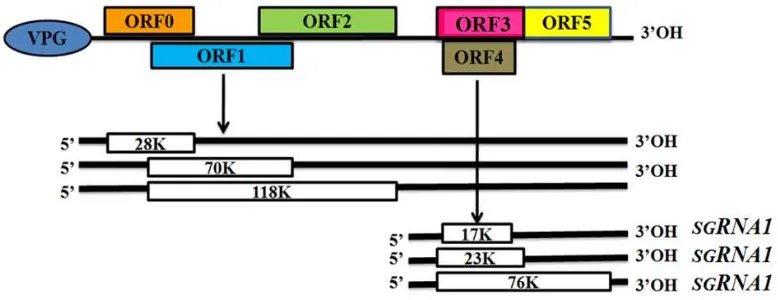
Identifikasi Tomato infectious chlorosis virus penyebab penyakit klorosis pada tanaman tomat di Cipanas Jawa Barat melalui perunutan nukleotida gen protein selubung utama..
• Mutan tsA (antigen T thermolabile) Æ tidak dapat memulai replikasi kromosom virus & tidak dapat memulai transkripsi gen-gen late untuk protein kapsid virus. Antigen T Æ
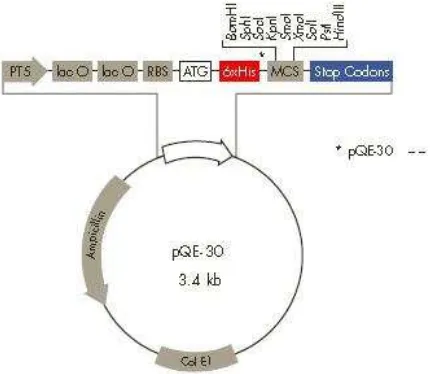
Ketujuh klon transforman dilakukan uji ekspresi protein pada skala kecil dengan menggunakan medium produksi yaitu media LB cair 10 ml yang ditambah ampisilin dan diinkubasi pada
Antibodi poliklonal adalah antibodi yang dapat diperoleh dari hasil hiperimunisasi. Hiperimunisasi merupakan imunisasi yang dilakukan secara sengaja terhadap hewan
Identifikasi Tomato infectious chlorosis virus penyebab penyakit klorosis pada tanaman tomat di Cipanas Jawa Barat melalui perunutan nukleotida gen protein selubung utama..
