KERAGAMAN GEN KALPASTATIN (CAST) DAN HUBUNGANNYA
DENGAN SIFAT PERTUMBUHAN DAN KUALITAS
KARKAS PADA DOMBA LOKAL
MUHAMMAD IHSAN ANDI DAGONG
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi berjudul “Keragaman Gen Kalpastatin (CAST) dan Hubungannya dengan Sifat Pertumbuhan dan Kualitas
Karkas pada Domba Lokal”, adalah karya saya dengan arahan dari komisi
pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir disertasi ini.
Bogor, Januari 2012
Muhammad Ihsan Andi Dagong
ABSTRACT
MUHAMMAD IHSAN ANDI DAGONG. Polymorphisms of Calpastatin (CAST) Gene and It’s Association with Growth Trait and Carcass Quality in Local Sheep. Supervised by CECE SUMANTRI, RONNY RACHMAN NOOR, RACHMAT HERMAN and MOHAMAD YAMIN.
Calpastatin (CAST) is an indigenous inhibitor of calpain that is involved in regulation of protein turn over and growth, therefore CAST gene is an excellent candidate gene for growth and carcass trait in livestock. The objectives of this research were to identify genetic polymorphisms in the part of intron 5 - exon 6 of CAST gene, to identify the association of CAST polymorphism with growth trait, and carcass characteristics as well as meat quality in the local sheep. A PCR-SSCP method was carried out to identify the genetic variation of CAST gene from 401 samples origin from three groups of local sheep ie. Fat Tail Sheep (FTS), Thin Tail Sheep (TTS) and Priangan Sheep. In total 157 heads of TTS were grouped based on CAST genotype and raised untill three months for growth trait measurement. A number of 33 heads of sheep representing of CAST genotype (CAST-11, CAST-12 and CAST-22) were used to identify carcass and meat characterization. The results shows that most population studied were polymorphic, with genotype frequencies of CAST-11, CAST-12, CAST-22, CAST-13, CAST-23, and CAST-33 were 29.7%, 38.2%, 24.2%, 2.5%, 4.5% and 0.7% respectively. CAST-1 and 2 alleles were most commonly found in all populations with total frequency 95.8%, while rare allele was CAST-3 (4.2%) and only found in thin tail population. There were no associations between CAST polymorphisms with body weight and body linear measurement, but a significant association was found between CAST variation and daily gain. The CAST-23 and CAST-33 genotype had higher daily gain when compared to other genotypes. There were no associations between CAST polymorphisms with meat tenderness, pH, water holding capacity and cooking loss, neither with carcass weight and dressing percentage. The shoulder proportion of CAST-11 genotype was larger when compared to those of CAST-12 or CAST-22 genotype, but the lean meat proportion of CAST-22 in shoulder, rack and loin was higher (P<0.05) when compared to the CAST-11 but it did not differ with CAST-12. The fat percentage of CAST-11 genotype was the highest among the genotypes. The CAST-22 genotype had higher lean meat percentage compared to the CAST-11 genotype. Its concluded that CAST genes were polymorphic and had a significant association with growth trait and carcass quality in local sheep.
RINGKASAN
MUHAMMAD IHSAN ANDI DAGONG. Keragaman Gen Kalpastatin (CAST) dan Hubungannya dengan Sifat Pertumbuhan dan Kualitas Karkas pada Domba Lokal. Dibimbing oleh CECE SUMANTRI, RONNY RACHMAN NOOR, RACHMAT HERMAN dan MOHAMAD YAMIN.
Domba lokal adalah salah satu sumber daya genetik yang sangat potensial untuk dikembangkan. Hal ini disebabkan domba lokal memiliki beberapa kelebihan yakni bersifat prolifik, daya adaptasi terhadap lingkungan dan penyakit yang lebih tinggi, siklus produksi relatif lebih singkat serta relatif sedikit membutuhkan modal. Selain itu, di beberapa daerah yang padat penduduknya seperti pulau Jawa, daging domba mampu mensubstitusi sebagian daging sapi yang masih harus diimpor setiap tahun. Nilai karkas domba sangat dipengaruhi oleh total produksi daging, distribusi dan proporsi daging pada karkas. Gen kalpastatin (CAST) diketahui memiliki hubungan dengan sifat kualitas karkas dan daging pada ternak. Kalpastatin terlibat dalam beragam proses fisiologi dalam tubuh ternak seperti pengaturan turn over protein, pertumbuhan, fusi dan migrasi mioblast. Diduga keragaman gen CAST mempunyai hubungan dengan pertumbuhan dan potensial digunakan sebagai marka gen untuk sifat pertumbuhan pada ternak. Kalpastatin berperan dalam menghambat fungsi enzim kalpain yang terlibat dalam mengatur turn over protein dan pertumbuhan.
Penelitian ini terbagi dalam tiga tahapan yaitu 1). Identifikasi keragaman gen CAST pada populasi domba lokal, 2). Hubungan keragaman gen CAST dengan sifat pertumbuhan pada domba lokal, 3). Hubungan keragaman gen CAST dengan kualitas daging dan karkas domba lokal. Metode PCR-SSCP digunakan pada penelitian tahap pertama untuk mengidentifikasi variasi pada gen CAST. Sebanyak 401 ekor domba lokal dari 8 sub populasi digunakan dalam penelitian ini, tiga group merupakan Domba Ekor Tipis (DET) dari Sukabumi, UP3J Jonggol dan Kissar, sedangkan sisanya antara lain domba Priangan dari Margawati dan Wanaraja serta Domba Ekor Gemuk (DEG) dari Donggala, Sumbawa dan pulau Rote. Pada tahap kedua, sebanyak 157 ekor DET yang dikelompokkan berdasarkan genotipe gen CAST dipelihara selama tiga bulan dan diukur parameter pertumbuhannya seperti bobot badan, PBB harian serta ukuran morfometriknya. Pada tahap ketiga sebanyak 33 ekor DET yang mewakili tiga genotipe CAST-11, CAST-12 dan CAST-22 diidentifikasi kualitas karkas dan dagingnya.
populasi DET. Disimpulkan bahwa gen CAST bersifat polimorfik dan informasi keragaman tersebut dapat digunakan untuk mencari hubungan dengan sifat pertumbuhan, kualitas karkas dan daging domba lokal.
Perbedaan bobot badan maupun morfometrik tidak menunjukkan perbedaan yang nyata antara variasi gen CAST pada domba yang diamati, namun variasi gen tersebut berhubungan dengan pertambahan bobot badan harian (PBB harian). Genotipe CAST-23 dan CAST-33 yang membawa alel CAST-2 dan CAST-3, diduga memiliki hubungan dengan performa pertumbuhan dan potensial untuk digunakan dalam seleksi sifat pertumbuhan. Tidak ada perbedaan sifat keempukan daging, pH, daya ikat air serta susut masak di antara ketiga genotipe (P>0.05). Tidak ada perbedaan bobot karkas maupun persentase karkas. Proporsi potongan shoulder CAST-11 lebih tinggi dibanding yang lainnya, namun proporsi daging CAST-22 pada potongan shoulder, rack dan loin nyata lebih tinggi dibanding CAST-11 namun tidak berbeda dengan CAST-12. Persentase lemak karkas CAST-11 lebih tinggi dibanding ketiga genotipe lainnya.
Genotipe CAST-11 cenderung memiliki perlemakan yang lebih tinggi dibanding CAST-22, sedangkan CAST-22 cenderung memiliki persentase daging yang lebih tinggi, sehingga untuk pengembangan ke depan guna mendapatkan domba yang perdagingannya lebih banyak sebaiknya dikembangkan domba yang memiliki genotipe CAST-22.
Keragaman gen CAST pada domba lokal cukup tinggi dan memiliki hubungan positif dengan pertambahan bobot badan harian dan kualitas karkas khususnya proporsi daging (lean) karkas. Keragaman gen CAST khususnya pada lokus intron 5 - ekson 6 kemungkinan potensial dapat digunakan sebagai marka seleksi untuk mendapatkan domba cepat tumbuh.
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
KERAGAMAN GEN KALPASTATIN (CAST) DAN HUBUNGANNYA
DENGAN SIFAT PERTUMBUHAN DAN KUALITAS
KARKAS PADA DOMBA LOKAL
MUHAMMAD IHSAN ANDI DAGONG
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : 1. Prof. (R). Dr. Ismeth Inounu
(Peneliti Utama Puslitbangnak Kementan RI)
2. Dr. Ir. Rudy Priyanto
(Staf Dosen Fakultas Peternakan IPB)
Penguji pada Ujian Terbuka : 1. Dr. Ir. Riwantoro, MM
(Sekretaris Dirjen Peternakan Kementan RI)
2. Prof. (R). Dr. Ir. Kusuma Diwyanto
Nama : Muhammad Ihsan Andi Dagong
NRP : D161080041
Program Studi / Mayor : Ilmu Produksi dan Teknologi Peternakan / ITP
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc
Ketua Anggota
Prof.Dr.Ir. Ronny R. Noor, M.Rur.Sc
Prof.(Em).Dr. drh. Rachmat Herman, M.V.Sc
Anggota Anggota
Dr. Ir. Mohamad Yamin, M.Agr.Sc
Mengetahui
Ketua Program Studi/Mayor Dekan Sekolah Pascasarjana Ilmu Produksi dan Teknologi Peternakan
Dr. Ir. Rarah R.A. Maheswari, DEA
UCAPAN TERIMA KASIH
Segala puja dan puji serta syukur tak lupa penulis persembahkan kepada Allah SWT, Tuhan semesta alam atas karunia Rahmat dan Kasih Sayang-Nya sehingga Disertasi Doktor yang berjudul Keragaman Gen Kalpastatin (CAST) dan Hubungannya dengan Sifat Pertumbuhan dan Kualitas Karkas pada Domba Lokal dapat diselesaikan.
Pada kesempatan ini penulis menyampaikan banyak terima kasih kepada : 1. Komisi pembimbing Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc, Prof. Dr. Ir.
Ronny Rachman Noor, M.Rur.Sc, Prof. (Em). Dr. drh. Rachmat Herman, M.VSc, dan Dr. Ir. Mohamad Yamin, M.Agr.Sc yang selalu dengan sabar dan bijaksana memberikan bimbingan dan arahan, masukan dan motivasi kepada penulis dalam menyelesaikan karya ilmiah ini.
2. Rektor Institut Pertanian Bogor, Dekan Sekolah Pascasarjana dan Dekan Fakultas Peternakan beserta seluruh stafnya yang telah memberikan kesempatan untuk melanjutkan studi S3 di Program Studi Ilmu Produksi dan Teknologi Peternakan.
3. Direktorat Jenderal Pendidikan Tinggi (DIKTI) atas beasiswa untuk studi S3 melalui program beasiswa BPPS serta atas kesempatan yang diberikan untuk mengikuti program sandwich-like di Gene Marker Laboratory, Lincoln University New Zealand dan bantuan dana Hibah Disertasi Doktor melalui dana DIPA IPB.
4. Prof. (R). Dr. Ir. Ismeth Inounu (Peneliti Utama Puslitbangnak Kementan RI) dan Dr. Ir. Rudy Priyanto (Staf Dosen Fapet IPB) atas kesediaanya sebagai penguji luar komisi dalam ujian tertutup, serta Dr. Ir. Riwantoro, MM (Sekretaris Dirjen Peternakan Kementan RI) dan Prof. (R). Dr. Ir. Kusuma Diwyanto (Peneliti Utama Puslitbangnak Kementan RI) atas kesediaannya sebagai penguji luar komisi pada ujian terbuka. Saran dan kritikan yang diberikan sangat membantu demi kesempurnaan tulisan ini.
5. Dr. Jonathan G.H. Hickford (Associate Professor), kepala Lab. Gene Marker Lincoln University, New Zealand atas kesediaannya untuk menerima penulis untuk melaksanakan program sandwich selama kurang lebih tiga bulan. 6. Prof. Dr. Ir. Muladno, MSA (Ketua Lab. Genetika Molekuler Ternak Fapet
IPB) beserta Dr. Ir. Jakaria atas kesempatan yang diberikan untuk melaksanakan penelitian di Lab. LGMT serta teman-teman seperjuangan di LGMT dan ABGSci (Pak Andi Baso Lompengeng, Eryk Andreas, Surya Nur Rahmatullah, Restu Misrianti, Ferdy Syaputra, Irene), teman seperjuangan S3 ITP 2008 serta adik-adik S1 (Widha, Dari, Monai dan kawan-kawan).
7. Keluarga besar di Watansoppeng (Pung Aji dan Mama Aji serta Etta dan Ibu Nene, Ibu, Faiz dan Meuthia) atas doa, dukungan dan motivasi yang diberikan.
8. Semua pihak yang tidak dapat penulis sebutkan satu per satu atas segala doa, bantuan dan dukungannya.
Semoga hasil penelitian ini dapat bermanfaat
RIWAYAT HIDUP
Penulis dilahirkan di Watansoppeng, Kabupaten Soppeng Sulawesi Selatan pada tanggal 26 Mei 1977 sebagai anak tunggal dari pasangan H. Andi Dagong dan Hj. Suhartati. Pendidikan sarjana ditempuh di Program Studi Produksi Ternak, Fakultas Peternakan Universitas Hasanuddin (UNHAS), lulus pada tahun 1999. Pada tahun 2000, penulis diterima di Program Studi Agribisnis pada Program Pascasarjana UNHAS dan menamatkannya pada tahun 2003. Kesempatan untuk melanjutkan ke program Doktor pada program studi Ilmu dan Teknologi Peternakan IPB diperoleh pada tahun 2008. Beasiswa pendidikan pascasarjana (BPPS) diperoleh dari Direktorat Pendidikan Tinggi (Dikti) Kementerian Pendidikan Nasional Republik Indonesia.
Penulis bekerja sebagai staf pengajar (Dosen) pada jurusan Produksi Ternak, Fakultas Peternakan UNHAS sejak tahun 2002. Mata kuliah yang diampu antara lain Ilmu Produksi Ternak Domba dan Kambing, Ilmu Produksi Ternak Potong dan Kerja, Manajemen Ternak Potong, Anatomi Ternak dan Pengantar Anatomi Ternak.
Selama mengikuti program Doktor (S3), penulis telah mendapat kesempatan untuk mengikuti program Sandwich-like di Laboratorium Gene Marker, Life Science and Agriculture Department, Lincoln University, New Zealand yang dibiayai oleh Direktorat Pendidikan Tinggi (Dikti) pada tahun 2009 selama tiga bulan. Karya ilmiah berjudul Genetic Polymorphisms of the Coding Region (Exon 6) of Calpastatin (CAST) in Indonesian Sheep diterbitkan pada jurnal Media Peternakan (Tahun 2011, Vol.34 No.3 pp.190 - 195). Artikel lain berjudul Karakteristik karkas Domba Ekor Tipis (DET) berdasarkan variasi genotipe gen kalpastatin (CAST) (lokus intron 5 – ekson 6) dalam proses review pada Jurnal Ilmu Ternak dan Veteriner (JITV). Karya-karya ilmiah tersebut merupakan bagian dari program S3 penulis.
DAFTAR PUBLIKASI
1. Genetic polymorphisms of the coding region (Exon 6) of calpastatin (CAST) in Indonesian sheep. Media Peternakan, Vol 34. No.3 pp. 190 – 195 (2011) (Akreditasi B DIKTI).
DAFTAR SINGKATAN
SNP = Single Nucleotide Polymorphisms
QTL = Quantitative Trait Loci
kDa = Kilo Dalton (ukuran berat molekul protein)
DNA = Deoxyribosa nucleat acid
3’ UTR = 3’ Untranslated region
WBSF = Warner Bratzler Shear Force
MAS = Marker Assisted Selection
PCR = Polymerase Chain Reaction
SSCP = Single Strand Conformational Polymorphisms
RFLP = Restriction Fragment Length Polymorphisms
BLAST = Basic Local Alignment Search Tool
DMA = Daya Mengikat Air
M/B ratio = Meat to Bone ratio
M/F ratio = Meat to Fat ratio
CAST = Kalpastatin (Calpastatin) SD = Standar Deviasi
DAFTAR ISI
Halaman
DAFTAR TABEL ……….. i
DAFTAR GAMBAR ………. ii
DAFTAR LAMPIRAN ……….. iv
PENDAHULUAN
Latar Belakang ……….. 1
Tujuan Penelitian ………. 2
Manfaat Penelitian ……… 3
Ruang Lingkup Penelitian ……… 3
TINJAUAN PUSTAKA
Keragaman Domba Lokal Indonesia ……… 5
Sistem Kalpain Kalpastatin dan Struktur Molekuler Gen Kalpastatin.. 7
Keragaman Gen Kalpastatin dan Hubungannya dengan Sifat Produksi 10
Pertumbuhan Otot ……… 11
Hubungan Kualitas Daging dengan Pertumbuhan Otot ……… 12
Karkas dan Komponennya ……… 13
Potongan Komersial Karkas Domba ……… 15
Marker Assisted Selection (MAS) ……… 16
KERAGAMAN GEN KALPASTATIN (CAST) PADA DOMBA LOKAL INDONESIA
Abstrak ………. 19
Abstract ……… 19 Pendahuluan ………. 20
Bahan dan Metode ……… 21
Hasil dan Pembahasan ……….. 25
KERAGAMAN GEN KALPASTATIN (CAST) DAN HUBUNGANNYA DENGAN PERFORMA PERTUMBUHAN PADA DOMBA EKOR TIPIS
Abstrak ………. 31
Abstract ……… 31 Pendahuluan ………. 32
Bahan dan Metode ……… 33
Hasil dan Pembahasan……… 34
Simpulan ……….. 37
KARAKTERISTIK KARKAS DAN KUALITAS FISIK DAGING PADA BERAGAM GENOTIPE GEN KALPASTATIN (CAST) DOMBA EKOR TIPIS
Abstrak ………. 39
Abstract ……… 39 Pendahuluan ………. 40
Bahan dan Metode ……….. 41
Hasil dan Pembahasan……… 47
Simpulan ……….. 63
PEMBAHASAN UMUM ………. 65
SIMPULAN DAN SARAN……….. 69
Simpulan………. 69
Saran ………. 69
DAFTAR PUSTAKA ……….. 71
DAFTAR TABEL
Halaman
1 Frekuensi genotipe dan alel gen CAST pada domba lokal ……… 27
2 Frekuensi genotipe observasi dan harapan gen CAST pada domba lokal ………. 27
3 Nilai heterosigositas pengamatan dan harapan gen CAST pada domba lokal ………..……… 28
4 Sebaran genotipe gen CAST domba Ekor Tipis (DET) ..……….. 34
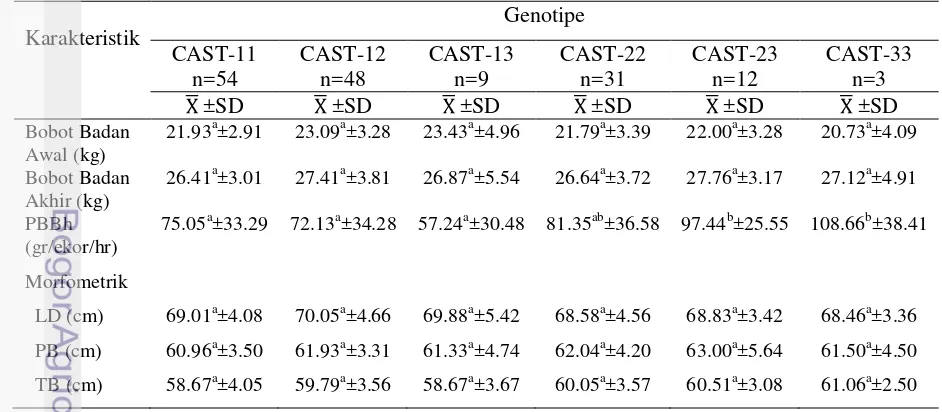
5 Rataan bobot badan, PBBh dan karakteristik morfometrik domba Ekor Tipis (DET) berdasarkan genotipe gen CAST ……….. 35
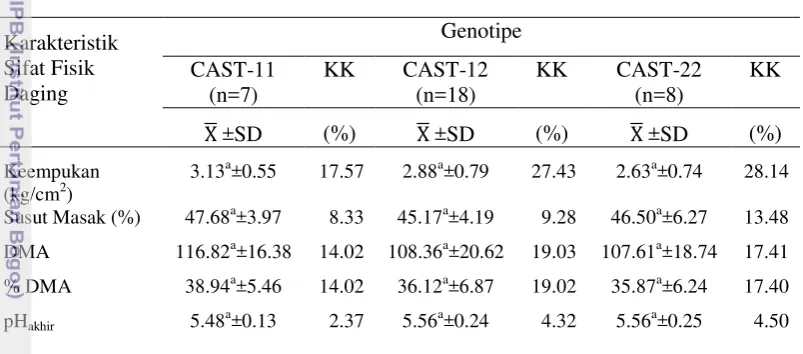
6 Kualitas sifat fisik daging berdasarkan variasi genotipe gen CAST…… 47
7 Rataan bobot potong, bobot kosong, bobot karkas hangat dan dingin, serta persentase karkas berdasarkan variasi genotipe gen CAST……… 50
8 Rataan bobot komponen non karkas berdasarkan variasi genotipe gen CAST ……… 51
9 Rataan bobot komponen potongan utama karkas (shoulder, rack, loin dan leg) berdasarkan variasi genotipe gen CAST ……… 52 10 Rataan bobot potongan karkas (neck, breast-foreshank dan flank)
berdasarkan variasi genotipe gen CAST ……..………. 53
11 Rataan persentase komponen potongan utama karkas (shoulder, rack,
loin dan leg) berdasarkan variasi genotipe gen CAST ……… 56 12 Rataan persentase komponen potongan karkas (neck, breast-foreshank
dan flank) berdasarkan variasi genotipe gen CAST……….. 57 13 Rataan rasio daging dengan tulang (M/B), dan rasio daging
dengan lemak (M/F) potongan karkas berdasarkan variasi genotipe gen CAST ……… 59
15 Rataan bobot otot utama karkas berdasarkan variasi genotipe gen CAST ……… 62
DAFTAR GAMBAR
Halaman
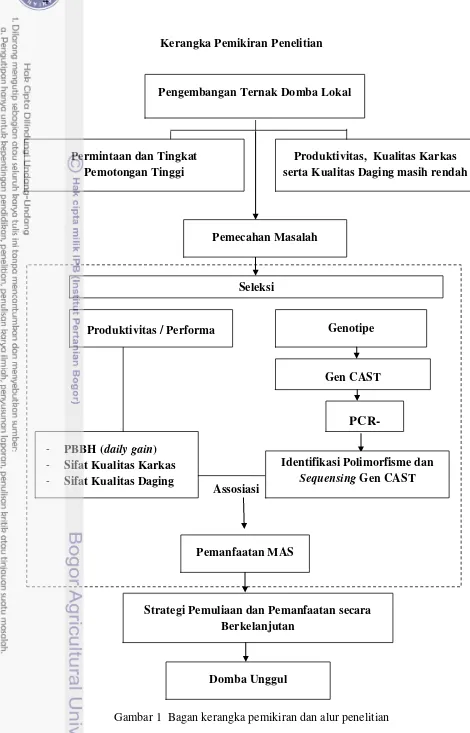
1 Bagan kerangka pemikiran dan alur penelitian……… 4
2 Struktur domain protein kalpastatin ……… 8
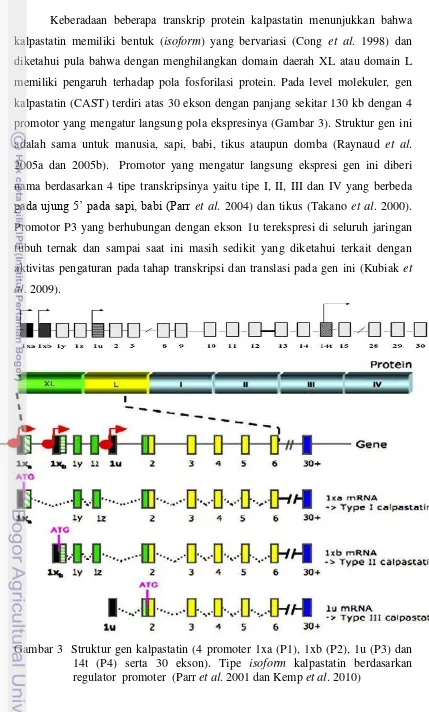
3 Struktur gen kalpastatin (CAST) ………. 9
4 Produk PCR gen kalpastatin(CAST)……… 25
5 Pola pita SSCP (single strandconformational polymorphism) gen
kalpastatin (CAST)……….………... 26
6 Runutan nukleotida alel CAST-1, CAST-2 dan CAST-3 ……….. 30
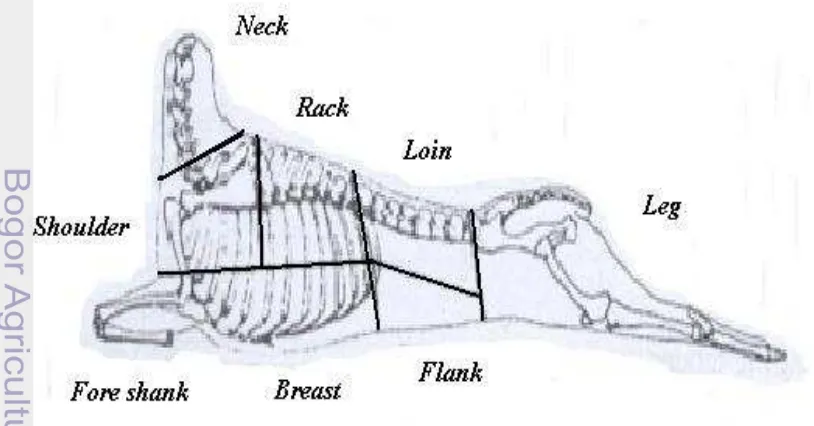
7 Potongan komersial karkas domba ………. 43
DAFTAR LAMPIRAN
Halaman
1 Runutan nukleotida alel CAST-1 hasil DNA sekuensing ………. 83
2 Runutan nukleotida alel CAST-2 hasil DNA sekuensing………….…. 84
3 Runutan nukleotida alel CAST-3 hasil DNA sekuensing ……….. 85
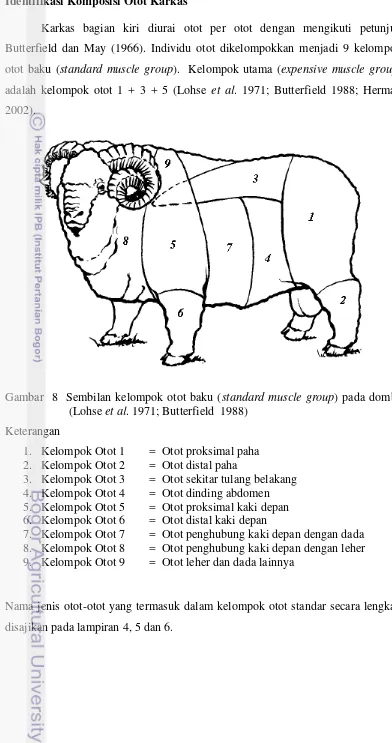
4 Jenis otot pada kelompok otot 1 (otot proksimal paha)……… 86
5 Jenis otot pada kelompok otot 2 (otot distal paha) dan kelompok otot 3 (otot sekitar tulang belakang) ……… 87
6 Jenis otot pada kelompok otot 4 (otot dinding abdomen) dan kelompok otot 5 (otot proksimal kaki depan).………. 88
7 Jenis otot pada kelompok otot 6 (otot distal kaki depan), kelompok otot 7 (otot penghubung kaki depan dengan leher) dan kelompok otot 8 (otot penghubung kaki depan dengan dada )……… 89
8 Jenis otot pada kelompok otot 9 (otot leher dan dada lainnya) ……….. 90
9 Rataan bobot otot-otot dalam kelompok 1 (otot proksimal paha) berdasarkan variasi genotipe gen CAST ………. 91
10 Rataan bobot otot-otot dalam kelompok otot 3 (otot sekitar tulang belakang) ……….. 92
11 Rataan bobot otot-otot dalam kelompok otot 5 (otot proksimal kaki depan) ……….. 93
12 Otot M. bicep femoris dan M. semimembranosus……… 94 13 Otot M. semitendinosus dan M. gastrocnemius………. 95 14 Otot M. rectus femoris dan M. vastus lateralis……….. 96 15 Otot M. gracilis dan M. adductor femoris……… 97 16 Contoh hasil uji non parametrik (Kruskal-Wallis) untuk sifat fisik
daging………..… 98
PENDAHULUAN
Latar Belakang
Domba lokal adalah salah satu sumber daya genetik yang sangat potensial
untuk dikembangkan. Hal ini disebabkan domba lokal memiliki beberapa
kelebihan yakni bersifat prolifik (beranak kembar), daya adaptasi terhadap
lingkungan dan penyakit yang lebih tinggi, siklus produksi relatif lebih singkat
serta relatif sedikit membutuhkan modal. Selain itu, di beberapa daerah yang
padat penduduknya seperti pulau Jawa, daging domba mampu mensubstitusi
sebagian daging sapi yang masih harus diimpor setiap tahun. Berdasarkan data
tahun 2008, kebutuhan domba khusus di Jawa Barat mencapai kisaran 3.343.365
ekor (Ditjennak 2009). Besarnya permintaan tersebut menunjukkan bahwa
prospek usaha budidaya domba masih terbuka lebar.
Terkait dengan upaya pengembangan domba, beberapa kelemahan yang
dimiliki oleh domba lokal yang perlu diperhatikan secara serius antara lain
bobot potong dan pertambahan bobot badan harian yang masih rendah dengan
variasi yang cukup tinggi (54 – 174 g/ekor/hari) (Yamin et al. 2009). Kualitas karkas dan daging domba lokal juga sangat bervariasi dan belum memenuhi
standar pasar internasional, sehingga dibutuhkan upaya perbaikan mutu genetik
yang dapat meningkatkan produktivitas, kualitas karkas dan daging domba lokal
yang berdampak pada peningkatan kontribusi produksi daging domba terhadap
total produksi daging dalam negeri yang saat ini hanya berkisar 5 % (Ditjennak
2009).
Kemajuan bioteknologi molekuler memungkinkan seleksi dapat
dilakukan pada tingkat DNA melalui pemanfaatan marker gen yang mempunyai
hubungan dengan sifat ekonomis. Salah satu marker gen yang ada hubungannya
dengan bobot badan pada domba lokal yakni kalpastatin (Sumantri et al. 2008). Sejumlah penelitian lain juga menunjukkan hubungan gen kalpastatin dengan
Kalpastatin adalah inhibitor spesifik bagi calpain yang merupakan enzim
protease yang tergantung pada ion Ca2+ (µ-calpain dan m-calpain) (Goll et al. 2003). Sistem kalpain kalpastatin ditemukan hampir di semua jaringan tubuh
hewan dan mempengaruhi banyak proses penting seperti pengaturan fusi dan
migrasi mioblas (Barnoy et al. 1996; Dedieu et al. 2004), turn over protein dan pertumbuhan otot (Huang & Forsberg 1998), serta longevity dan fertilitas (Garcia et al. 2006). Sistem kalpain kalpastatin banyak mendapat perhatian khusus karena peranan utamanya dalam produksi daging dan diyakini terlibat
dalam proses degradasi protein miofibril dalam proses keempukan pasca
pemotongan melalui pengaturan aktivitas kalpain (Koohmaraie1992).
Informasi mengenai hubungan gen kalpastatin dengan sifat pertumbuhan,
kualitas karkas serta komposisi dan perkembangan otot pada domba lokal saat ini
belum tersedia, sehingga dibutuhkan informasi mengenai pengaruh gen tersebut.
Keragaman genetik gen kalpastatin diharapkan dapat digunakan sebagai salah
satu penciri pada program seleksi untuk mendapatkan domba dengan sifat
pertumbuhan dan kualitas karkas yang lebih unggul.
Tujuan Pen eli tian
Secara khusus penelitian ini mempunyai tujuan :
1. Mendapatkan informasi keragaman genotipe dan frekuensi alel gen
kalpastatin (CAST) khususnya pada daerah intron 5 - ekson 6 pada
beberapa sub populasi domba lokal di Indonesia.
2. Mempelajari dan mendapatkan informasi hubungan antara keragaman gen
CAST dengan sifat pertumbuhan dan kualitas karkas pada domba lokal.
3. Mempelajari gambaran komposisi otot-otot domba berdasarkan keragaman
Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan gambaran mengenai
keragaman genetik gen kalpastatin (CAST) pada intron 5 - ekson 6. Berdasarkan
keragaman genetik tersebut diharapkan dapat menjadi penanda (marker) DNA
yang berhubungan dengan sifat pertumbuhan dan kualitas karkas domba lokal
serta dapat dimanfaatkan sebagai salah satu penciri seleksi untuk mempercepat
kemajuan genetik dalam program seleksi mendapatkan domba lokal unggul.
Hasil penelitian ini juga diharapkan menjadi salah satu informasi dasar dalam
melengkapi data riset genetika molekuler ternak dalam upaya perbaikan dan
penentuan kebijakan peningkatan mutu genetik dan pemanfaatan domba lokal
secara berkelanjutan.
Ruang Lingkup Penelitian
Ruang lingkup dan batasan dalam penelitian ini adalah :
1. Identifikasi keragaman genetik gen kalpastatin (CAST) pada domba lokal
Indonesia.
2. Hubungan keragaman genetik gen CAST dengan sifat pertumbuhan pada
domba lokal.
3. Hubungan keragaman genetik gen CAST dengan sifat kualitas karkas dan
daging pada domba lokal, serta profil dan gambaran distribusi otot karkas
Kerangka Pemikiran Penelitian
Gambar 1 Bagan kerangka pemikiran dan alur penelitian Domba Unggul
Strategi Pemuliaan dan Pemanfaatan secara Berkelanjutan
Produktivitas, Kualitas Karkas serta Kualitas Daging masih rendah
Gen CAST Genotipe Seleksi
Pemanfaatan MAS Assosiasi
PCR-- PBBH (daily gain)
- Sifat Kualitas Karkas
- Sifat Kualitas Daging
Produktivitas / Performa
Pengembangan Ternak Domba Lokal
Pemecahan Masalah Permintaan dan Tingkat
Pemotongan Tinggi
Identifikasi Polimorfisme dan
TINJAUAN PUSTAKA
Keragaman Domba Lokal Indonesia
Domba merupakan ternak yang pertama kali didomestikasi, dimulai dari
daerah Kaspia, Iran, India, Asia Barat, Asia Tenggara, dan Eropa sampai ke
Afrika (Gatenby 1991). Indonesia mempunyai tiga jenis domba yang telah
beradaptasi dengan baik pada kondisi lingkungan dan pengelolaan secara intensif
maupun ekstensif. Dua jenis di antaranya adalah Domba Ekor Tipis (DET)
(Javanese Thin Tailed) dan Domba Ekor Gemuk (DEG) (Javanese Fat Tailed) (Subandriyo 1993), serta domba Priangan yang dikenal sebagai domba Garut
(Mulyaningsih 1990). Domba-domba lokal lainnya yang tersebar di Indonesia
diduga merupakan rumpun dari ketiga jenis domba tersebut. DET diduga berasal
dari India/Bangladesh sedangkan DEG kemungkinan berasal dari Asia Barat.
Selama kurun 150 tahun terakhir juga telah dintroduksikan bangsa domba Eropa
dari Belanda, Australia dan New Zealand namun kontribusinya terhadap pool gen
domba lokal hanya sedikit diakibatkan tingginya mortalitas domba import tersebut
(Bradford & Inounu 1996).
Domba Ekor Tipis (DET)
Domba ekor tipis merupakan domba yang banyak terdapat di Jawa Barat
dan Jawa Tengah. Domba ini termasuk golongan domba kecil, dengan bobot
potong pada jantan berkisar 30 – 40 kg, sedangkan pada betina berkisar
15 - 20 kg. Warna bulu putih dan biasanya memiliki bercak hitam di sekeliling
matanya. Domba jantan memiliki tanduk melingkar, sedangkan yang betina
biasanya tidak bertanduk. Bulunya berupa wol yang kasar (Subandriyo 1993).
Ekornya tidak menunjukkan adanya deposisi lemak, dengan ukuran panjang ekor
rata-rata 19.3 cm, lebar pangkal 5.6 cm dan tebal 2.7 cm (Tiesnamurti 1992).
Salah satu keunggulan Domba Ekor Tipis (DET) adalah sifatnya yang
2.2 kg, rataan bobot sapih 10.0 kg, rataan pertambahan bobot badan harian 87.96 g
(Priyanto et al. 1992).
Sumantri et al. (2007) melaporkan domba Jonggol jantan dewasa mempunyai bobot badan sebesar 34.9 kg, sedangkan bobot badan betina sebesar
26.11 kg. Bobot badan domba Jonggol lebih tinggi bila dibandingkan sejumlah
domba lokal lainnya, misalnya bila dibandingkan dengan bobot badan jantan
dewasa dan betina dari domba Donggala (24.0 dan 25.3 kg), Kisar (25.8 dan 18.9
kg), dan Rote (27.9 dan 20.3 kg), akan tetapi hampir sama dengan bobot badan
dewasa domba jantan dan betina dari Sumbawa (33.8 dan 26.9 kg).
Domba Ekor Gemuk (DEG)
Domba Ekor Gemuk (DEG) banyak terdapat di Jawa Timur dan Madura,
serta pulau-pulau di Nusa Tenggara (pulau Rote dan Sumbawa). Telinga DEG
umumnya berukuran medium dengan posisi agak tegak menggantung
(semipendulous). Di Sulawesi Tengah dikenal sebagai domba Palu atau Donggala. Tanda-tanda yang merupakan karakteristik khas domba ekor gemuk
adalah ekor yang besar, lebar dan panjang. Bagian pangkal ekor membesar
merupakan timbunan lemak, sedangkan bagian ujung ekor kecil tidak
berlemak. Warna bulu putih, tidak bertanduk. Bulu wolnya kasar. Domba ini
dikenal sebagai domba yang tahan terhadap panas dan kering. Domba ini diduga
berasal dari Asia Barat Daya yang dibawa oleh pedagang bangsa Arab pada abad
ke-18 (Salamena 2006).
Bentuk tubuh domba ekor gemuk lebih besar dari pada domba ekor
tipis. Domba ini merupakan domba tipe pedaging, bobot jantan dewasa antara 40
– 60 kg, sedangkan bobot badan betina dewasa 25 – 35 kg. Tinggi badan pada jantan dewasa antara 60 – 65 cm, sedangkan pada betina dewasa 52 – 60 cm.
Domba Priangan / Domba Garut
Jenis domba lokal lainnya adalah domba Priangan yang tersebar di daerah
Bandung, Garut, Sumedang, Ciamis, dan Tasikmalaya. Domba Garut
dikelompokkan dalam dua kelompok yakni domba Garut Wanaraja (Garut
pedaging) dan domba Garut Cibuluh (Garut tangkas). Kedua jenis domba ini
memiliki penampilan yang berbeda. Domba Garut Wanaraja umumnya berbulu
putih dan bulunya lebih halus. Domba Garut Cibuluh mudah dikenali karena daun
telinganya yang kecil (rudimenter), memiliki tubuh yang kekar dan besar, serta umumnya berwarna hitam. Domba jantan bertanduk besar sehingga sering
dipertandingkan, khususnya untuk diadu tangkas, sedangkan domba betina
umumnya tidak bertanduk (Wiradarya 2005).
Domba Priangan merupakan domba utama di Jawa Barat yang telah
beradaptasi baik dengan kondisi lingkungan panas dan kelembaban tinggi. Domba
ini memiliki ciri-ciri profil kepala cembung dan mempunyai bentuk muka bagian
atas lebar, pendek, sedikit cembung, kelopak mata agak menonjol. Pada jantan
mempunyai tanduk berat, melingkar mendekati leher karena besar, panjang
mencapai 55 cm, dasar tanduk 21 cm, jarak dasar tanduk hampir bersentuhan satu
sama lain. Pada betina tidak bertanduk hanya kadang-kadang dijumpai benjolan,
kepala kecil terutama bagian atas tidak demikian sempurna, hanya betina
bentuknya lebih halus. Bobot badan dewasa dapat mencapai kisaran 50 – 62 kg
pada jantan dan 35 – 40 kg pada betina (Davendra & McLeroy 1992).
Sistem Kalpain Kalpastatin dan Struktur Molekuler Gen Kalpastatin
Pada level fisiologik, kalpastatin adalah inhibitor spesifik bagi kalpain
yang merupakan enzim protease yang tergantung pada ion Ca2+ (µ-calpain dan
m-calpain) (Goll et al. 2003). Sistem kalpain kalpastatin ditemukan hampir di semua jaringan tubuh hewan dan mempengaruhi banyak proses penting seperti
dalam proses degradasi protein miofibril dalam proses keempukan pasca
pemotongan melalui pengaturan aktivitas kalpain (Koohmaraie 1992).
Banyak penelitian dalam bidang peternakan, khususnya pada ternak
pedaging berusaha untuk mengungkap peranan kalpastatin secara genetik dan
fisiologis di dalam tubuh ternak. Level kalpastatin bervariasi di antara spesies
ternak (Koohmaraie 1992), bangsa (Shackelford et al. 1995), otot (Geesink & Koohmaraie 1999), dan berpengaruh secara genetik terhadap kualitas karkas
(Bickerstaffe et al. 2006; Schenkel et al. 2006) dan daging (Perry et al. 2009; Van Eeneenaam et al. 2007).
Pada level protein, kalpastatin merupakan protein dengan lima domain
penghambat (Gambar 2) yang diperkirakan memiliki bobot molekul sekitar 65 -
76 kDa yang ditemukan pada eritrosit (Killefer & Koohmaraie 1994) dan sekitar
107 – 172 kDa pada jaringan yang lebih besar (Raynoud et al. 2005a). Kalpastatin ada diseluruh jaringan otot yang mengekspresikan kalpain. Salah satu domain
yakni N-terminal leader (L) tidak menghambat aktivitas kalpain (Emori et al. 1987), namun kemungkinan terlibat dalam lokalisasi intraselular (Averna et al. 2001) sedangkan 4 domain lainnya (domain I – IV) memiliki homologi yang
tinggi dan masing-masing mempunyai kemampuan menghambat aktivitas kalpain
(Emori et al. 1987; Cong et al. 1998). Domain penghambat pada kalpastatin memiliki tiga region yang sangat terkonservasi yakni region A, B dan C. Region
A dan C terikat pada kalpain yang tergantung pada ion Ca2+ namun tidak memiliki
aktivitas penghambat sedangkan region B menghambat aktivitas kalpain (Tompa
et al. 2002).
Gambar 2 Struktur domain protein kalpastatin. 4 domain penghambat dengan 786 asam amino. A, B dan C adalah domain penghambat yang sangat
conserve (Odeh 2003)
Keberadaan beberapa transkrip protein kalpastatin menunjukkan bahwa
kalpastatin memiliki bentuk (isoform) yang bervariasi (Cong et al. 1998) dan diketahui pula bahwa dengan menghilangkan domain daerah XL atau domain L
memiliki pengaruh terhadap pola fosforilasi protein. Pada level molekuler, gen
kalpastatin (CAST) terdiri atas 30 ekson dengan panjang sekitar 130 kb dengan 4
promotor yang mengatur langsung pola ekspresinya (Gambar 3). Struktur gen ini
adalah sama untuk manusia, sapi, babi, tikus ataupun domba (Raynaud et al. 2005a dan 2005b). Promotor yang mengatur langsung ekspresi gen ini diberi
nama berdasarkan 4 tipe transkripsinya yaitu tipe I, II, III dan IV yang berbeda
pada ujung 5’ pada sapi, babi (Parr et al. 2004) dan tikus (Takano et al. 2000). Promotor P3 yang berhubungan dengan ekson 1u terekspresi di seluruh jaringan
tubuh ternak dan sampai saat ini masih sedikit yang diketahui terkait dengan
aktivitas pengaturan pada tahap transkripsi dan translasi pada gen ini (Kubiak et al. 2009).
Keragaman Gen CAST dan Hubungannya dengan Sifat Produksi
Gen CAST dikenal bersifat polimorfik pada ternak sapi, dan variasi gen
CAST telah digunakan sebagai marker gen pada dua marker komersial yakni
GeneStar Tenderness dan Igenity TenderGENE. GeneStar Tenderness
menggunakan SNP G/A pada daerah 3’ UTR (posisi basa 2959 dari gen Bank :
AF159246) (Barendse et al. 2002), sementara Igenity TenderGENE menggunakan SNP G/C pada daerah intron 5 (posisi basa 282 dari gen Bank : AY008267) (Van
Eeneenam et al. 2007). Gen CAST juga diketahui bersifat polimorfik pada babi (Krzęcio 2008), dan kambing (Zhou & Hickford 2008c). Pada domba gen CAST
juga diketahui sangat polimorfik dengan 5 allel pada daerah intron 5– ekson 6
(Zhou et al. 2007) dan 4 allel pada daerah intron 12 (Roberts et al. 1996, Byun et al. 2009a). Namun sampai saat ini keragaman gen CAST pada daerah promotor baru dilaporkan pada sapi (Kubiak et al. 2009) dan belum ada informasi terkait dengan polimorfisme tersebut pada ternak domba.
Hasil analisis Quantitative Traits Loci (QTL) menunjukkan bahwa gen CAST berasosiasi kuat dengan sifat pertumbuhan pada domba silang balik antara
Domba Ekor Tipis (DET) dengan domba Merino (Margawati 2005). Sumantri et al. (2008) melaporkan gen CAST pada fragmen intron 1 memiliki hubungan yang kuat dengan bobot badan pada domba lokal, individu bergenotipe MN mempunyai
bobot badan lebih besar daripada individu bergenotipe NN. Frekuensi alel M
sangat bervariasi tertinggi pada domba Garut tangkas Ciomas/Bogor (0.29), Garut
Margawati (0.24), DET Jonggol/Bogor (0.16) dan terendah pada domba DEG
Madura dan Sumbawa (0.04). Lebih lanjut Sumantri et al. (2008) melaporkan frekuensi alel M di tiga populasi berkisar 0.16 – 0.29, tetapi individu bergenotipe
MM tidak ditemukan pada domba lokal yang diobservasi. Fenomena ini
kemungkinan disebabkan oleh seleksi negatif, domba berbobot badan besar
kemungkinan besar bergenotipe MM banyak dipotong.
Nassiry et al. (2006) juga melaporkan gen CAST intron 1 memiliki hubungan yang kuat dengan pertambahan bobot badan harian (daily gain) pada domba Kurdi. Namun variasi tersebut dilaporkan tidak ada hubungannya dengan
bobot lahir. Namun hubungan keragaman tersebut tidak ada hubungannya dengan
lama hidup (longevity) dan fertilitas pada domba (Byun et al. 2009c) hal berbeda yang diketahui pada sapi perah (Garcia et al. 2006). Bickerstaffe et al. (2006) melaporkan variasi di ekson 6 berhubungan dengan kualitas karkas, dimana alel A
berpengaruh pada peningkatan 15% otot longissimus dorsi (shortloins).
Penelitian lain pada ternak sapi menunjukkan beberapa polimorfisme dari
gen CAST memiliki hubungan dengan kualitas karkas, ternak dengan genotipe CC
cenderung memiliki persentase daging tanpa lemak (lean) dan tulang yang lebih rendah dibanding genotipe GG (Schenkel et al. 2006). Polimorfisme gen CAST juga memiliki hubungan dengan nilai keempukan Warner-Bratzler Shear Force
(WBSF) (Barendse 2002; Koohmaraie et al. 1995). Secara khusus korelasi genetik antara aktivitas kalpastatin dengan keempukan daging telah diukur oleh Casas et al. (2006) dengan menggunakan WBSF. Hasil penelitian tersebut menunjukkan hubungan antara marker gen CAST dengan WBSF dan nilai keempukan sangat
signifikan (P<0.01). Ternak dengan genotipe CC dan CT menghasilkan daging
yang lebih alot dibandingkan dengan genotipe TT. Estimasi heritabilitas dari
keempukan uji organoleptik berkisar 0.22 sampai 0.50, sedangkan heritabilitas
WBSF sekitar 0.12 sampai 0.58 (Gregory et al. 1995; Wheeler et al. 1996;
O’Connor et al. 1997). Heritabilitas yang cukup tinggi dari sifat keempukan daging menunjukkan bahwa gen tersebut sangat potensial digunakan untuk
memprediksi dan menekan variabilitas dari sifat tersebut dengan seleksi
menggunakan teknologi biologi molekuler (Costello et al. 2007).
Pertumbuhan Otot
Hyperplasia (peningkatan jumlah sel) dan hypertrophy (peningkatan ukuran sel) adalah penentu utama massa otot. Jika hyperplasia didefinisikan sebagai jumlah sel secara aktual, maka sifat ini ditentukan oleh proliferasi sel-sel
embrional. Namun, jika hyperplasia didefinisikan sebagai total kandungan DNA sel-sel otot, maka hal tersebut ditentukan oleh proliferasi sel-sel sebelum lahir,
dan pertumbuhan setelah lahir dan perkembangan sel-sel satelit. Ternak yang
potensi perototan yang lebih besar, demikian pula dengan ternak yang memiliki
sel-sel satelit yang lebih aktif juga akan memiliki potensi untuk memiliki
perototan yang besar (Koohmaraie et al. 2002).
Besar ukuran otot ditentukan oleh keseimbangan antara jumlah protein
otot yang disintesa dengan yang didegradasi. Jika jumlah protein otot yang
disintesa lebih besar dibanding jumlah protein otot yang didegradasi, maka akan
menyebabkan hypertrophy. Ada beberapa skenario yang dapat menyebabkan
hypertrophy yaitu : 1). Peningkatan sintesa protein yang diikuti dengan penurunan degradasi protein, 2). Peningkatan sintesa protein dan degradasi
protein, namun peningkatan sintesa protein lebih tinggi dibanding peningkatan
degradasi protein, dan 3). Penurunan sintesa protein dan degradasi protein,
namun tingkat penurunan degradasi protein lebih rendah dibanding dengan
tingkat penurunan sintesa (Koohmaraie et al. 2002).
Protein otot yang memegang peranan penting dalam pertumbuhan otot
rangka adalah protein miofibril yang merupakan fraksi protein utama dalam otot
rangka. Degradasi dari protein miofibril yang menyebabkan variasi dalam
pertumbuhan otot serta kualitas daging (keempukan) khususnya pada otot
longissimus (Koohmaraie et al. 2002). Goll et al. (1989) menunjukkan pentingnya degradasi protein terhadap efisiensi deposisi massa daging tanpa
lemak (lean mass deposition). Pengurangan 10% dari tingkat degradasi protein (dari 3%/hari menjadi 2.7%/hari) akan menyebabkan penggandaan pertambahan
bobot badan secara hipotesis pada sapi yang memiliki bobot 454 kg.
Hubungan Kualitas Daging (Keempukan) dengan Pertumbuhan Otot
Jika pertumbuhan otot adalah hasil dari hyperplasia (peningkatan jumlah sel pada fase embrional atau peningkatan kandungan DNA melalui aktivitas
sel-sel satelite), maka tidak akan ada pengaruh negatif terhadap sifat keempukan
daging (Koohmaraie et al. 2002). Sebagai contoh adalah fenotip double muscling pada sapi, daging yang berasal dari sapi ini lebih empuk khususnya pada daging dimana jaringan ikat sebagai penentu keempukan (Wheeler et al. 2001). Namun hal yang berbeda jika pertumbuhan otot adalah hasil dari
Hypertrophy terjadi akibat penurunan laju degradasi protein otot dan dapat mempengaruhi kualitas daging khususnya keempukan. Hal ini ditunjukkan
pada domba dengan fenotip Callipyge. Otot domba Callipyge mengalami hypertrophy akibat penurunan laju degradasi protein otot. Hal ini ditandai dengan
aktivitas kalpastatin otot yang tinggi dan menyebabkan domba memiliki fenotipe
perdagingan yang ekstrim pada bagian hindquarter, namun dagingnya akan sangat alot (Freking et al. 1998 & 2004; Koohmaraie et al. 1995).
Karkas dan Komponennya
Karkas adalah bagian tubuh ternak setelah dikurangi bagian saluran
pencernaan, kepala, kulit dan keempat kaki. Bobot karkas adalah bobot hidup
setelah dikurangi bobot saluran pencernaan, darah, kepala, kulit dan keempat kaki
dari persendian carpus atau tarsus ke bawah (Berg & Butterfield 1976). Terkadang dijumpai beberapa modifikasi dengan atau tanpa ginjal, lemak ginjal,
lemak pelvis, lemak sekitar ambing, diaphragma dan ekor. Perbedaan sangat besar
adalah lemak atau ginjal termasuk dalam karkas atau tidak (Colomer-Rocher et al. 1987).
Persentase karkas dapat diukur dalam beberapa cara yakni dengan
mengukur perbandingan antara bobot karkas dengan bobot hidup saat dipotong
atau perbandingan bobot hidup dengan bobot kosong (empty body weight) yakni bobot setelah dikurangi isi saluran pencernaan dan urine (Ekiz et al. 2009). Persentase karkas dipengaruhi oleh bobot karkas, bobot ternak, kondisi, bangsa
ternak, proporsi bagian-bagian non karkas, ransum yang diberikan serta cara
pemotongan (Berg & Butterfield 1976).
Herman (2005) menyatakan bahwa persentase karkas domba Priangan
adalah sebesar 48.70% dan domba Ekor Gemuk adalah sebesar 52.39% pada
bobot potong 25 kg. Pada bobot potong 40 kg, domba Priangan memiliki
persentase karkas sebesar 54.90% dan domba Ekor Gemuk sebesar 55.49%
(Herman 2002). Persentase karkas bervariasi karena bobot hidup dan perlemakan
dari domba tersebut, sedangkan persentase tulang, otot dan lemak dalam karkas
Persentase karkas domba lokal tidak terlalu jauh berbeda dengan
persentase karkas pada beberapa bangsa domba lainnya seperti yang dilaporkan
pada domba Omani (48%) oleh Mahgoub et al. (2000), domba Mexican Pelibuey (44%), Gutierrez et al. (2005); domba berambut yang dipelihara secara intensif (52%), Burke et al. (2003) dan Kawas et al. (2007), domba Ovin Martinik (57%) (Archimede et al. 2008), dan domba Chios (54.59%) Ekiz et al. (2009).
Komponen utama karkas terdiri atas jaringan otot, tulang dan lemak.
Kualitas karkas sangat ditentukan oleh ketiga komponen tersebut. Perubahan
proporsi dari satu komponen akan mempengaruhi proporsi komponen lainnya.
Proporsi relatif dari ketiga komponen tersebut yang menentukan kualitas karkas
(Aberle et al. 2001). Tulang sebagai kerangka tubuh merupakan komponen karkas yang tumbuh dan berkembang paling dini (early maturing tissue), kemudian disusul oleh jaringan otot, dan yang paling akhir adalah jaringan lemak
(late maturing tissue) (Aberle et al. 2001; Abdullah et al. 2008). Proporsi komponen karkas dan potongan karkas yang dikehendaki oleh konsumen adalah
karkas atau potongan karkas yang terdiri atas proporsi daging tanpa lemak (lean) yang tinggi, tulang yang rendah dan lemak yang optimal (Macit 2002).
Komponen karkas yang dapat memberikan nilai ekonomis adalah lemak,
karena lemak berfungsi sebagai pembungkus daging dan memberikan keempukan
pada daging (Berg & Butterfield 1976). Namun, seiring dengan meningkatnya
kesadaran konsumen akan makanan yang sehat, perlemakan yang terlalu banyak
pada karkas cenderung dihindari. Kandungan lemak (perlemakan) pada domba
lokal memperlihatkan perbedaan yang nyata karena perbedaan bangsa. Herman
(2002) menyatakan bahwa pada bobot potong 25 kg, persentase karkas, otot,
tulang dan lemak pada domba Priangan berturut-turut adalah sebesar 48.80%,
62.28%, 17.05%, dan 18,67%, sedangkan pada domba Ekor Gemuk berturut-turut
adalah 52.73%, 53.55%, 15.50%, dan 29.30%.
Pertumbuhan jaringan lemak lebih lambat dibandingkan dengan
pertumbuhan otot, tetapi dengan berlangsungnya pertumbuhan ternak, peletakan
lemak secara bertahap akan melebihi pertumbuhan otot, selanjutnya begitu tingkat
kedewasaan ternak tercapai pertumbuhan otot cenderung stabil tetapi perletakan
artinya secara konstan lemak disimpan dan dimobilisasi dalam tubuh, lemak
banyak terkumpul dalam dinding rongga perut dan ginjal (Aberle et al. 2001). Pada ternak muda, lemak dideposisi pertama kali pada daerah organ dalam
tubuh (visceral), kemudiaan saat mendapatkan nutrisi yang cukup, maka lemak
akan dideposisi di bawah kulit (lemak subkutan), kemudian lemak antar otot
(intermuskular) dan lemak yang paling terakhir dideposisi adalah lemak di antara
serat otot (intramuskular) (Aberle et al. 2001).
Rasio perbandingan antara ketiga komponen karkas baik berupa rasio
daging dengan tulang, ataupun rasio daging dengan lemak umumnya dikaitkan
dengan tingkat perdagingan ternak (Purchas et al. 1991). Rasio daging dengan tulang atau rasio daging dengan lemak yang tinggi menunjukkan perdagingan
yang bagus, rasio daging dan lemak pada daerah leg dan shoulder umumnya lebih tinggi dibanding pada daerah rack atau loin, yang disebabkan pola pertumbuhan distoproksimal pada ternak. Artinya bagian tubuh ternak khususnya pada daerah
shoulder dan leg berkembang lebih awal dibandingkan pada bagian rack atau loin
(Abdullah et al. 2008).
Potongan Komersial Karkas Domba
Cara pemotongan potongan komersial karkas ditentukan oleh spesies
ternak dan selera konsumen, oleh karena itu ditemukan cara pemotongan yang
berbeda-beda pada satu negara dengan negara yang lainnya. Bicer et al. (1995) hanya membagi potongan karkas domba dalam empat potongan utama yakni
shoulder-neck-brisket, back-chest, loin-belly dan rump-leg-hindshanks. Colomer-Rocher et al. (1987) membagi karkas dalam enam potongan yakni leg, shoulder, neck, ribs, loin, dan breast. Abdullah et al. (2008) membagi potongan utama karkas (primal cut) hanya dalam empat potongan utama yakni shoulder, rack, loin
dan leg. Rodrigues et al. (2006) membagi potongan karkas dalam tiga kategori, yakni potongan leg, chump dan loin sebagai kategori/kualitas pertama, ribs,
anterior ribs dan shoulder sebagai kualitas kedua, sedangan breast, shank dan
neck adalah kualitas ketiga.
Herman (1993) menyatakan bahwa pada irisan karkas utama,
lebih besar dengan persentase otot lebih tinggi dan lemak lebih rendah
dibandingkan dengan irisan shoulder domba Ekor Gemuk. Ekor Gemuk mempunyai potongan leg yang lebih besar daripada Priangan, tetapi persentase lemaknya lebih tinggi dan ototnya lebih rendah. Persentase potongan leg pada Priangan (30.8%) pada Ekor Gemuk (32.3%), loin pada Priangan (9.1%) pada Ekor Gemuk (10.1%), rack pada Priangan (9.4%) pada Ekor Gemuk (8.6%) dan
shoulder pada Priangan (28.2%) pada Ekor Gemuk (27.3%).
Marker Assisted Selection (MAS)
Penggunaan Marker Assisted Selection (MAS) didasarkan pada gagasan bahwa terdapat gen yang memegang peranan utama dan menjadi sasaran atau
target secara spesifik dalam seleksi (Van der Werf 2000). Beberapa sifat yang
dikendalikan oleh gen tunggal seperti warna bulu merupakan pola pewarisan sifat
yang sederhana, namun beberapa sifat utamanya sifat produksi yang kompleks
(kuantitatif) dikontrol oleh banyak gen (poly gene) (Nicholas 1996; Noor 2008). Gen-gen sifat kuantitatif yang memiliki pengaruh besar merupakan gen-gen yang
disebut sebagai gen utama (major gene) yang terletak pada lokus sifat kuantitatif (Quantitative Trait Loci/QTL). Marker gen telah banyak digunakan untuk mengidentifikasi ternak sapi yang memiliki performa lebih bagus pada beberapa
sifat komersial seperti kualitas daging (keempukan) (Barendse et al. 2008).
Dua dekade terakhir telah diidentifikasi beberapa gen yang memiliki
pengaruh utama (major gene) terhadap sifat-sifat ekonomis pada ternak. Beberapa
gen utama seperti gen Myostatin yang menyebabkan double muscling dan berpengaruh pada produksi daging sapi (Grobet et al. 1997), gen Callipyge yang berpengaruh pada produksi daging domba (White et al. 2008), serta gen Booroola
atau gen prolifik (banyak anak) pada domba (Fogarty 2009).
Walaupun penggunaan MAS adalah suatu harapan yang optimis,
penerapan MAS akan lebih tepat digunakan pada skala industri pemuliaan ternak
atau industri peternakan sehingga keberhasilan penerapannya memerlukan strategi
terpadu yang menyeluruh untuk skala usaha peternakan besar (Dekkers 2004).
Meuwissen (2001) menyatakan bahwa MAS penting pada situasi yang ketepatan
digunakan, ketepatan seleksi diharapkan dapat lebih baik sejak periode anak atau
fase embrional.
Marker Assisted Selection (MAS) dapat membantu dalam meningkatkan laju peningkatan mutu genetik ternak melalui pemanfaatan informasi molekuler
KERAGAMAN GENETIK GEN KALPASTATIN (CAST)
PADA DOMBA LOKAL INDONESIA
Abstrak
Kalpastatin (CAST) berperan dalam menghambat fungsi enzim kalpain yang terlibat dalam mengatur turn over protein dan pertumbuhan, sehingga gen kalpastatin (CAST) diyakini sebagai salah satu kandidat gen untuk sifat pertumbuhan dan kualitas karkas. Tujuan penelitian ini adalah untuk mengidentifikasi keragaman gen CAST khususnya pada daerah sebagian intron 5 dan seluruh exon 6 pada domba. Metode PCR-SSCP digunakan untuk mengidentifikasi variasi pada gen CAST. Sebanyak 401 ekor domba dari 8 sub populasi digunakan dalam penelitian ini, tiga group merupakan Domba Ekor Tipis (DET) dari Sukabumi, UP3J Jonggol dan Kissar. Sisanya adalah antara lain domba Priangan dari Margawati dan Wanaraja serta Domba Ekor Gemuk (DEG) dari Donggala, Sumbawa dan pulau Rote. Analisis SSCP menunjukkan tiga pola SSCP berbeda yang merujuk pada tiga alel CAST berbeda yaitu alel CAST-1, 2 dan 3 dengan enam genotipe berbeda. Variasi genetik di antara populasi dihitung berdasarkan frekuensi alel dan genotipenya. Sebagian besar populasi yang diteliti menunjukkan polimorfisme gen CAST dengan frekuensi masing-masing genotipe CAST-11, CAST-12, CAST-22, CAST-13, CAST-23, dan CAST-33 adalah 29.7%, 38.2%, 24.2%, 2.5%, 4.5% dan 0.7%. CAST-1 dan 2 adalah alel yang paling umum pada seluruh populasi dengan total frekuensi 95.8%, sementara alel yang langka adalah CAST-3 (4.2%) dan hanya ditemukan di populasi DET. Dapat disimpulkan bahwa gen CAST bersifat polimorfik dan informasi tersebut dapat digunakan untuk mencari hubungan dengan sifat pertumbuhan, kualitas karkas dan daging domba lokal.
Abstract
Pendahuluan
Kalpastatin (CAST) merupakan salah satu enzim yang termasuk dalam
bagian sistem kalpain kalpastatin yang melibatkan tiga molekul yakni : µ-kalpain,
m-kalpain dan kalpastatin yang berfungsi sebagai penghambat spesifik dari kedua
jenis kalpain. Sistem ini berperan dalam berbagai macam proses fisiologik dan
patologik (Kidd et al. 2000; Huang et al. 2001; Goll et al. 2003; Raynaud et al. 2004) dan terlibat dalam proses pengaturan turn over protein dan pertumbuhan (Goll et al. 1992), migrasi myoblast (Dedieu et al. 2003) dan fusi myoblast
(Temm-Grove et al. 1999). Berdasarkan hal tersebut, gen CAST diyakini sebagai salah satu kandidat gen untuk sifat pertumbuhan dan kualitas karkas pada
ternak.
Telah banyak penelitian yang menunjukkan hubungan polimorfisme gen
CAST dengan kualitas karkas dan daging, khususnya sifat keempukan pada
beberapa ternak (Schenkel et al. 2006; Casas et al. 2006; Curi et al. 2009). Pada domba, polimorfisme gen CAST dilaporkan mempunyai hubungan dengan bobot
lahir (Byun et al. 2008), dan bobot badan (Sumantri et al. 2008,) namun tidak mempengaruhi sifat keempukan daging anak domba (Zhou & Hickford 2008a).
Beberapa penelitian terdahulu melaporkan polimorfisme gen CAST pada
domba lokal, namun penelitian tersebut hanya fokus pada daerah intron
(Sumantri et al. 2008; Sutikno et al. 2011), dan belum ada laporan keragaman pada daerah penyandi asam amino (coding region) khususnya pada domba lokal. Ekson 6 adalah ekson terbesar pada gen kalpastatin domba, dan diketahui bersifat
polimorphik pada domba dengan ditemukannya lima alel yang berbeda (Zhou et al. 2007, Byun et al. 2009a). Tullio et al. (2009) melaporkan bahwa sekuen yang disandi oleh ekson 6 diketahui memiliki beberapa situs posporilasi, dan terlibat
langsung dalam menentukan lokalisasi sel. Hasil tersebut menunjukkan bahwa
variasi yang ada dalam sekuens ekson 6 diduga dapat mempengaruhi aktivitas
ion Ca2+ channels dan kemudian dapat mengatur atau memodulasi aktivitas kalpain. Tujuan penelitian ini adalah untuk mengidentifikasi keragaman
(polimorfisme) gen CAST khususnya pada daerah penyandi asam amino (ekson
Bahan dan Metode
Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan mulai bulan April 2010 sampai April 2011.
Penelitian laboratorium dilaksanakan di Laboratorium Genetika Molekuler
Ternak Bagian Pemuliaan dan Genetika Ternak Fakultas Peternakan IPB untuk
isolasi dan purifikasi DNA, serta analisis PCR-SSCP (single strand conformational polymorphism).
Identifikasi Keragaman Gen Kalpastatin (CAST) pada Domba Lokal
Materi penelitian adalah sampel DNA genom koleksi Laboratorium
Molekuler dan Genetika Ternak (LGMT) Bagian Ilmu Pemuliaan dan Genetika
Fakultas Peternakan IPB, yang berasal dari tiga jenis domba lokal yakni Domba
Ekor Tipis (DET), Domba Ekor Gemuk (DEG) dan Domba Priangan/Garut yang
diperoleh dari beberapa daerah sub populasi domba lokal di Indonesia. Total 401
sampel DNA domba yang terbagi atas kelompok DEG dari Donggala (54),
Sumbawa (29), dan pulau Rote (12), kelompok DET dari UP3J Jonggol (52)
Kab. Bogor milik Fakultas Peternakan IPB, Kissar Maluku (32 ekor), dan
Sukabumi (161) serta kelompok domba Priangan/Garut dari Margawati (26 ekor)
(Garut Pedaging) dan Wanaraja (35 ekor) (Garut Tangkas) Kabupaten Garut,
Jawa Barat.
Teknik Pengambilan Sampel Darah
Pengambilan sampel darah dilakukan dengan mengumpulkan sekitar 5 ml
sampel darah dari domba melalui vena jugularis dengan menggunakan venojet
dan tabung vacuttainer tanpa heparin. Sampel darah tersebut kemudian diawetkan dengan etanol absolute dengan perbandingan 1:1 dan kemudian
Ekstraksi DNA Genom
Ekstraksi DNA genom dari sampel darah dilakukan dengan menggunakan
metode standar phenol chloroform (Sambrook et al. 1989) yang dimodifikasi oleh Andreas et al. (2010) dengan menggunakan enzim Proteinase K dan Phenol-Chloroform untuk mendegradasi protein dan lemak kemudian dipresifitasi
menggunakan etanol absolut. DNA hasil ekstraksi kemudian ditambahkan
larutan TE 80% dan siap untuk analisis PCR.
Analisis PCR-SSCP (Single Strand Conformational Polymorphism)
Fragmen gen CAST domba diidentifikasi dengan menggunakan metode
PCR-SSCP (single strand conformational polymorphism). Fragmen intron 5 - ekson 6 diamplifikasi dengan runutan primer F:5’-GTTATGAAT
TGCTTTCTACTC-3’ dan primer R: 5’-ATACGATTGAGAGACTTCAC-3’
berdasarkan Zhou et al. (2007) dengan prediksi panjang produk PCR sekitar 254 bp.
Sampel DNA dari masing-masing individu digunakan sebagai cetakan
untuk mengamplifikasi lokus gen CAST melalui reaksi PCR dengan sekuen
nukleotida pengapit masing-masing (Primer Forward dan Reverse). Amplifikasi
dilakukan dengan PCR dikondisikan pada volume reaksi 25 μl yang terdiri atas
50 - 100 ng DNA template, 0.25 μM primer, 150 μM dNTP (Fermentas), 2.5
mM Mg2+, 0.5 U Taq DNA polymerase (Fermentas) dan 1x buffer. Kondisi mesin PCR dimulai dengan denaturasi awal pada suhu 94oC selama 5 menit,
diikuti dengan 35 siklus berikutnya masing-masing 94oC selama 30 detik (tahap
denaturasi), 56oC selama 45 detik (tahap annealing), 72oC selama 45 detik
(tahap ekstensi), yang diakhiri dengan satu siklus ekstensi akhir pada suhu 72oC
5 menit dengan menggunakan mesin PCR (Mastercycler Personal 22331,
Eppendorf, Jerman). Produk PCR kemudian dielektrophoresis pada gel agarose
Identifikasi konformasi DNA utas tunggal (single strand conformational), dilakukan dengan mencampur 5 μl produk PCR dengan 5 μl formamide dye
(98% formamide, 10 mM EDTA, 0.025% bromophenol blue, 0.025% xylene
cyanol). Kemudian sampel didenaturasi pada suhu 95oC selama 5 menit, dan
langsung didinginkan dalam ice bath dan diloading pada gel poliakrilamid (29:1) 12%. Elektroforesis dilakukan dengan menggunakan Protean II xi cells (Bio-Rad)
pada kondisi 300 V, suhu 5 oC selama 18 jam pada larutan TBE buffer 0.5x.
Setelah elektrophoresis gel kemudian dilakukan pewarnaan dengan
silver-stainning berdasarkan metode Byun et al. (2009b) yang dimodifikasi pada larutan stainning (0.1% AgNO3, 0.04% NaOH 10 N dan 0.4% NH3).
Penentuan Posisi Pita DNA dan Genotipe Gen CAST
Penentuan posisi pita DNA pada gel poliakrilamid dilakukan secara
manual. Ukuran dan jumlah dari alel yang muncul pada gel ditentukan
berdasarkan asumsi bahwa semua pita DNA dengan laju migrasi yang sama
adalah homolog, sedangkan alel dengan migrasi paling cepat ditetapkan sebagai
alel 1, berikutnya adalah alel 2 dan seterusnya.
Analisis Sekuensing DNA
Sampel yang menunjukkan pola pita SSCP yang berbeda dilanjutkan
untuk analisis sekuensing. Analisis ini untuk menyakinkan hasil polimorfisme
yang diperoleh dari hasil analisis PCR-SSCP. Sampel-sampel yang menunjukkan
pola SSCP berbeda dan dalam keadaan homozigot yang digunakan sebagai
cetakan DNA untuk sekuensing. Sebelum sekuensing, salah satu pita unik yang
mewakili tiap-tiap alel diambil dari gel dan kemudian dipurifikasi mengikuti
metode yang dijelaskan oleh Hu et al. (2010). Sampel tersebut kemudian diamplifikasi (PCR) ulang, untuk memastikan identitas produk yang dihasilkan,
maka produk PCR hasil purifikasi dikonfirmasi ulang dengan produk PCR
sampel DNA genom melalui analisis PCR-SSCP secara bersamaan.
Perbandingan, translasi serta aligment sekuens dilakukan dengan
(basic local alignment search tool) digunakan untuk mencari homologi sekuens dengan sekuens yang ada di GenBank NCBI
(http://www.ncbi.nlm.nih.gov/BLAST).
Analisis Statistik
Frekuensi alel dan genotipe dihitung berdasarkan rumus Nei dan Kumar
(2000).
dimana Xii = frekuensi genotipe ii, Xi = frekuensi alel ke-i, nii = jumlah sampel
dengan genotipe ii, nij=jumlah sampeldengan genotipe ij, dan N= total sampel. Test keseimbangan Hardy-Weinberg (HWE) dengan uji chi-square (Kaps
& Lamberson 2004).
dimana χ² = chi-square , Obs = jumlah genotipe ke-ii hasil pengamatan, dan Exp
= jumlah genotipe ke-ii yang diharapkan.
Nilai heterozigositas pengamatan (Ho) dan heterozigositas harapan (He)
berdasarkan rumus heterozigositas Nei dan dihitung dengan menggunakan
software PopGene32 versi 1.31 (Yeh et al. 1999).
dimana Ho = heterozigositas pengamatan di antara populasi, He = heterozigositas
harapan di antara populasi, = ukuran relatif populasi, Xkij(i≠j) = frekuensi AiAj
Hasil dan Pembahasan
Keragaman Gen Kalpastatin (CAST)
Sebagian daerah intron 5 dan seluruh bagian ekson 6 pada fragmen gen
CAST berhasil diamplifikasi dengan primer forward dan reverse, dengan prediksi panjang produk PCR 254 bp (Gambar 4). Hasil analisis PCR-SSCP menunjukkan
polimorfisme pada daerah ini dengan ditemukannya tiga pola SSCP yang berbeda
yang merujuk pada tiga alel yang berbeda, alel CAST-1, CAST-2 dan CAST-3.
Baik satu atau dua pita unik yang berbeda ditemukan pada tiap individu yang
menunjukkan individu tersebut homozigot atau heterozigot dengan enam genotipe
yang berbeda, yakni CAST-11, CAST-12, CAST-22, CAST-13, CAST-23 dan
CAST-33.
Level polimorfisme yang ditemukan pada penelitian ini lebih rendah
dibanding yang dilaporkan oleh Zhou et al. (2007) yang menemukan lima alel berbeda pada domba Merino, Corriedale, Poll Dorset dan NZ crossbreed. Namun,
hasil ini lebih tinggi dibanding yang dilaporkan pada domba lokal Indonesia
(dengan PCR-RFLP) (Sumantri et al. 2008; Sutikno et al. 2011), domba Karakul Iran (Shahroudi et al. 2006) dan domba Kurdi (Nassiry et al. 2006)
Alel yang paling umum ditemukan adalah alel CAST-1 dan CAST-2
dengan frekuensi sebesar 50% dan 45.8% dengan total frekuensi keduanya
95.8%, sementara alel CAST-3 adalah alel langka dengan frekuensi dipopulasi
hanya berkisar 4.2%. Genotipe yang paling umum ditemukan adalah CAST-12
(38.2%). Zhou et al. (2007) juga menemukan alel CAST-1 dan CAST-2 sebagai alel yang paling umum, dengan total frekuensi 82% pada populasi domba
Merino, Corriedale, Romney, Poll Dorset dan NZ cross-breed, sementara alel
yang langka adalah CAST-3 (13%), CAST-4 (2%) dan CAST-5 (3%). Gambar 5
menunjukkan genotipe gen CAST hasil elektroforesis setelah SSCP.
Gambar 5 Pola pita SSCP (single strand conformational polymorphism) gen kalpastatin (CAST)
Frekuensi genotipe CAST-11 pada kelompok DEG dari populasi
Sumbawa, Rote dan Donggala berkisar 8% sampai 16.7%. Nilai tersebut lebih
rendah dibanding dengan kelompok domba Priangan dari populasi Margawati
(85.0%) atau DET dari populasi Jonggol (42.3%). Frekuensi genotipe CAST-12
pada kelompok DEG berkisar 10.3% sampai 83.3% dan lebih tinggi dibanding
populasi domba Priangan dari Wanaraja (57.1%) dan populasi DET dari Kissar
(53.1%). Sementara frekuensi CAST-22 pada DEG berkisar 8.3% sampai 79.3%
lebih tinggi dibanding populasi domba priangan dan DET. Genotipe CAST-23
hanya ditemukan pada populasi DET (Sukabumi, Kissar dan Jonggol), dengan
frekuensi tertinggi pada sub populasi Kissar (15.6%). Genotipe CAST-13 (6.2%)
Tabel 1 Frekuensi genotipe dan alel gen CAST pada domba lokal
Penelitian lain dengan menggunakan teknik PCR-RFLP melaporkan
polimorfisme kalpastatin (lokus CAST-Msp1) pada domba lokal dengan dua tipe alel (M dan N), namun hanya menemukan dua genotipe (MN dan NN) dengan
frekuensi masing-masing 25% dan 75%, dan 13% dan 87% untuk frekuensi alel
M dan N (Sumantri et al. 2008).
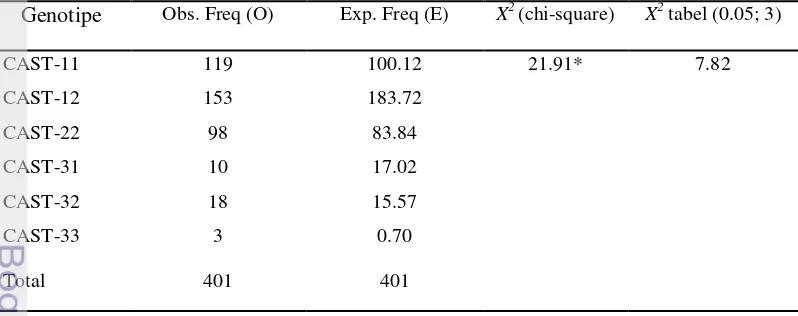
Tabel 2 Frekuensi genotipe observasi dan harapan gen CAST pada domba lokal
Genotipe Obs. Freq (O) Exp. Freq (E) X2 (chi-square) X2 tabel (0.05; 3)
* = Tidak setimbang (not equilibrium)
Hasil uji chi-square (X2) menunjukkan distribusi keenam genotipe gen CAST pada populasi tidak tersebar sesuai dengan kesetimbangan
Hardy-Weinberg (Tabel 2). Alel CAST-3 yang hanya ditemukan pada populasi DET,
kemungkinan disebabkan perkawinan tidak terjadi secara acak atau disebabkan