TANGGAP FUNGSIONAL PARASITOID TELUR
Trichogramma pretiosum
Riley terhadap TELUR INANG
Corcyra
cephalonica
Stainton pada PERTANAMAN KEDELAI
Oleh :
Mia Nuratni Yanti Rachman A44101051
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
ABSTRAK
MIA NURATNI YANTI RACHMAN. Tanggap Fungsional Parasitoid Telur
Trichogramma pretiosum Riley terhadap Telur Inang Corcyra cephalonica
Stainton pada Pertanaman Kedelai. Dibimbing oleh PUDJIANTO.
Penelitian bertujuan mengetahui hubungan antara tingkat parasitisasi
T. pretiosum pada kepadatan inang C. cephalonica berbeda dan mengetahui pengaruh pemanfaatan parasitoid terhadap serangga lain pada pertanaman kedelai. Penelitian dilaksanakan Desember 2004-Juli 2005 di Laboratorium Bioekologi Parasitoid dan Predator serta kebun percobaan Cikabayan. Penelitian menggunakan RAK dengan tiga ulangan. Data diolah menggunakan SAS, dilanjutkan uji Duncan taraf 5%.
Lahan pertanaman kedelai yang luasnya 600 m² dibagi tiga petak. Tanaman kedelai 18 buah dipilih dengan jarak 1 m dari titik pelepasan. Pias kepadatan telur berbeda (20, 40, 80, 160, 320, dan 640 butir) ditempelkan pada permukaan bawah daun.
Pelepasan parasitoid dilaksanakan pukul 07:00 WIB. Pelepasan dilakukan saat tanaman berumur 3 MST. Jumlah pelepasan parasitoid sebanyak lima kali dan jumlah total parasitoid sebanyak 6000 per pelepasan. Pias dipaparkan selama ± 8 jam dan diambil pukul ± 15:00 WIB.
Pemasangan pias tetap dilakukan untuk melihat kemapanan T. pretiosum
dengan cara sama seperti pelepasan dan dilakukan sebanyak 4 kali. Pengambilan contoh telur-telur serangga hama pada pertanaman dilakukan secara diagonal.
Tipe tanggap fungsional T. pretiosum pada pertanaman kedelai dapat berubah karena kondisi lingkungan. Grafik hubungan kepadatan telur dengan tingkat parasitisasi menunjukkan korelasi positif. Kemapanan parasitoid bertahan 2 minggu.
Parasitoid larva yang diidentifikasi ialah famili Ichneumonidae, Braconidae, dan Elasmidae. Parasitoid telur yang keluar adalah T. pretiosum dan
TANGGAP FUNGSIONAL PARASITOID TELUR
Trichogramma pretiosum
Riley terhadap TELUR INANG
Corcyra
cephalonica
Stainton pada PERTANAMAN KEDELAI
Skripsi
Sebagai salah satu syarat untuk memperoleh Gelar Sarjana Pertanian Pada Fakultas Pertanian,
Institut Pertanian Bogor
Oleh :
Mia Nuratni Yanti Rachman A44101051
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
Judul : TANGGAP FUNGSIONAL PARASITOID TELUR
Trichogramma pretiosum Riley terhadap TELUR INANG
Corcyra cephalonica Stainton pada PERTANAMAN KEDELAI
Nama : Mia Nuratni Yanti Rachman NRP : A44101051
Menyetujui,
Pembimbing
Dr. Ir. Pudjianto, MSi NIP. 131 475 578
Mengetahui,
Dekan Fakultas Pertanian
Prof. Dr. Ir. Supiandi Sabiham, MAgr.
NIP. 130 422 698
RIWAYAT HIDUP
Penulis dilahirkan di Bekasi pada tanggal 23 September 1982 sebagai anak pertama dari empat bersaudara, anak dari pasangan H. Taufik Rachman dan Hj. Fatmah AF.
PRAKATA
Puji syukur penulis panjatkan ke hadirat Allah Subhanahu wa Ta’ala yang memberikan rahmat dan hidayah-Nya, sehingga penulis dapat menyelesaikan skripsi yang berjudul “Tanggap Fungsional Parasitoid Telur Trichogramma pretiosum Riley terhadap Telur Inang Corcyra cephalonica Stainton pada Pertanaman Kedelai”.
Pada kesempatan ini penulis mengucapkan terima kasih kepada:
1. Dr. Ir. Pudjianto, MSi. sebagai pembimbing, yang senantiasa sabar dalam membimbing dan mengarahkan selama penelitian dan penulisan skripsi ini. 2. Dr. Ir. Widodo, MS., selaku penguji tamu yang telah memberikan saran dan
kritik dalam penyusunan laporan ini.
3. Keluarga (Mama, Papa, my sister Ika, my brothers Fadli dan Fauzi) di Bekasi yang telah memberikan kasih sayang, perhatian, nasehat dan doa.
4. Seluruh anggota ( mbak Nita (alm), mbak Ada, mbak Diana, mbak Atik, dan lain- lain) Laboratorium Bioekologi Parasitoid dan Predator Departemen Proteksi Tanaman, Fakultas Pertanian yang telah membantu dalam penelitian dan penulisan skripsi ini.
5. Seluruh staf pengajar Departemen Proteksi Tanaman, Fakultas Pertanian yang telah memberikan ilmu selama penulis kuliah.
6. My best friends Mu’min, Okti, Rosita, Lia, Asti, dan Desmi yang telah memberikan arti persahabatan yang sebenarnya.
7. Kakak kelas dan rekan-rekan HPT 38 dari Departemen Proteksi Tanaman. 8. Kristal crew yang telah membantu dalam penulisan skripsi ini.
9. Semua pihak terkait yang telah membantu dalam penyelesaian skripsi ini. Penulis menyadari bahwa dalam penulisan skripsi ini, masih ada kekurangan. Oleh karena itu, kritik dan saran sangat diharapkan oleh penulis untuk perbaikan kegiatan selanjutnya. Semoga skripsi ini dapat bermanfaat. Amin.
Bogor, Februari 2006
DAFTAR ISI
DAFTAR TABEL
Nomor Halaman
Teks
1. Jumlah telur terparasit pada beberapa pelepasan ... 22 2. Jumlah telur terparasit pada kemapanan hidup T. pretiosum ... 23
Lampiran
1. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 3 MST ... 36 2. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 4 MST ... 37 3. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 5 MST ... 38 4. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 6 MST ... 39 5. Hasil analisis data telur terparasit pada pelepasan parasitoid
DAFTAR GAMBAR
Nomor Halaman
Teks
1. Hubungan antara kepadatan telur inang C. cephalonica
dengan jumlah telur inang terparasit pada saat tanaman berumur 3 MST ... 18
2. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 4 MST ... 19
3. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 5 MST ... 19
4. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 6 MST ... 19
5. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 7 MST ... 20
6. Grafik jumlah hama yang didapatkan selama sampling ... 25
7. Jumlah parasitoid yang terkumpul dari sampling ... 26
Lampiran 1. Petak percobaan dengan 18 tanaman berjarak 1 m dari titik pelepasan ... 33
2. Penempelan pias pada permukaan bawah daun ... 33
3. Tabung reaksi berisi parasitoid dengan posisi tegak lurus saat pelepasan parasitoid ... 33
4. Kondisi lahan saat generatif akhir ... 34
5. Imago betina T. pretiosum ... 34
6. Parasitoid larva C. chalcites famili Ichneumonidae ... 34
7. Parasitoid larva C. chalcites famili Braconidae ... 35
8. Parasitoid telur Ooencyrtus sp. famili Encyrtidae ... 35
9. Parasitoid larva C. chalcites famili Elasmidae spesies A ... 35
PENDAHULUAN
Latar belakang
Trichogrammatidae merupakan famili yang terkenal sebagai agens pengendali hayati berbagai serangga hama. Genera yang terkenal sebagai parasitoid telur adalah Trichogramma dan Trichogrammatoidea. Pemanfaatan parasitoid telur ini telah lama dilakukan dalam bidang pertanian dan kehutanan. Pemanfaatan Trichogramma dalam skala besar difokuskan pada tanaman pangan dan tanaman industri (Li 1994). Trichogramma digunakan untuk mengendalikan serangga-serangga hama penting yang merugikan secara ekonomi, seperti Ostrinia
sp. (Lepidoptera: Pyralidae), Chilo sp. (Lepidoptera: Pyralidae), dan Helicoverpa
sp. (Lepidoptera: Noctuidae).
Faktor utama yang mene ntukan keefektifan pemanfaatan Trichogramma
adalah pemilihan spesies yang sesuai, kualitas parasitoid yang dibiakkan, jumlah parasitoid yang dilepaskan, metode pelepasan, teknik pelepasan, iklim, waktu, dan integrasi dengan metode pengendalian lain. Pemilihan spesies parasitoid yang sesuai penting untuk mengendalikan serangga hama sasaran. Sebagai contoh, pada lahan pertanaman padi di Cina, T. japonicum merupakan spesies yang dominan dan menunjukkan kemampuan parasitisasi yang tinggi pada telur-telur Lepidoptera dibandingkan T. chilonis dan T. dendrolimi (Li 1994). Kualitas parasitoid berhubungan dengan lamanya parasitoid dibiakkan di laboratorium. Semakin lama parasitoid dibiakkan di laboratorium maka semakin menurun kualitas parasitoid tersebut. Jumlah parasitoid yang dilepaskan berkaitan dengan populasi serangga hama sasaran. Metode pelepasan yang umum digunakan ada dua macam yaitu inundasi dan inokulasi. Teknik pelepasan terdiri dari cara manual dan mekanik. Iklim dan waktu berkaitan dengan perilaku parasitoid dalam pencarian inang. Integrasi dengan metode pengendalian lain dapat mendukung keefektifan Trichogramma misalnya penanaman tanaman berbunga sebagai tanaman pinggir yang dapat menyediakan makanan bagi parasitoid.
TANGGAP FUNGSIONAL PARASITOID TELUR
Trichogramma pretiosum
Riley terhadap TELUR INANG
Corcyra
cephalonica
Stainton pada PERTANAMAN KEDELAI
Oleh :
Mia Nuratni Yanti Rachman A44101051
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
ABSTRAK
MIA NURATNI YANTI RACHMAN. Tanggap Fungsional Parasitoid Telur
Trichogramma pretiosum Riley terhadap Telur Inang Corcyra cephalonica
Stainton pada Pertanaman Kedelai. Dibimbing oleh PUDJIANTO.
Penelitian bertujuan mengetahui hubungan antara tingkat parasitisasi
T. pretiosum pada kepadatan inang C. cephalonica berbeda dan mengetahui pengaruh pemanfaatan parasitoid terhadap serangga lain pada pertanaman kedelai. Penelitian dilaksanakan Desember 2004-Juli 2005 di Laboratorium Bioekologi Parasitoid dan Predator serta kebun percobaan Cikabayan. Penelitian menggunakan RAK dengan tiga ulangan. Data diolah menggunakan SAS, dilanjutkan uji Duncan taraf 5%.
Lahan pertanaman kedelai yang luasnya 600 m² dibagi tiga petak. Tanaman kedelai 18 buah dipilih dengan jarak 1 m dari titik pelepasan. Pias kepadatan telur berbeda (20, 40, 80, 160, 320, dan 640 butir) ditempelkan pada permukaan bawah daun.
Pelepasan parasitoid dilaksanakan pukul 07:00 WIB. Pelepasan dilakukan saat tanaman berumur 3 MST. Jumlah pelepasan parasitoid sebanyak lima kali dan jumlah total parasitoid sebanyak 6000 per pelepasan. Pias dipaparkan selama ± 8 jam dan diambil pukul ± 15:00 WIB.
Pemasangan pias tetap dilakukan untuk melihat kemapanan T. pretiosum
dengan cara sama seperti pelepasan dan dilakukan sebanyak 4 kali. Pengambilan contoh telur-telur serangga hama pada pertanaman dilakukan secara diagonal.
Tipe tanggap fungsional T. pretiosum pada pertanaman kedelai dapat berubah karena kondisi lingkungan. Grafik hubungan kepadatan telur dengan tingkat parasitisasi menunjukkan korelasi positif. Kemapanan parasitoid bertahan 2 minggu.
Parasitoid larva yang diidentifikasi ialah famili Ichneumonidae, Braconidae, dan Elasmidae. Parasitoid telur yang keluar adalah T. pretiosum dan
TANGGAP FUNGSIONAL PARASITOID TELUR
Trichogramma pretiosum
Riley terhadap TELUR INANG
Corcyra
cephalonica
Stainton pada PERTANAMAN KEDELAI
Skripsi
Sebagai salah satu syarat untuk memperoleh Gelar Sarjana Pertanian Pada Fakultas Pertanian,
Institut Pertanian Bogor
Oleh :
Mia Nuratni Yanti Rachman A44101051
PROGRAM STUDI HAMA DAN PENYAKIT TUMBUHAN
FAKULTAS PERTANIAN
Judul : TANGGAP FUNGSIONAL PARASITOID TELUR
Trichogramma pretiosum Riley terhadap TELUR INANG
Corcyra cephalonica Stainton pada PERTANAMAN KEDELAI
Nama : Mia Nuratni Yanti Rachman NRP : A44101051
Menyetujui,
Pembimbing
Dr. Ir. Pudjianto, MSi NIP. 131 475 578
Mengetahui,
Dekan Fakultas Pertanian
Prof. Dr. Ir. Supiandi Sabiham, MAgr.
NIP. 130 422 698
RIWAYAT HIDUP
Penulis dilahirkan di Bekasi pada tanggal 23 September 1982 sebagai anak pertama dari empat bersaudara, anak dari pasangan H. Taufik Rachman dan Hj. Fatmah AF.
PRAKATA
Puji syukur penulis panjatkan ke hadirat Allah Subhanahu wa Ta’ala yang memberikan rahmat dan hidayah-Nya, sehingga penulis dapat menyelesaikan skripsi yang berjudul “Tanggap Fungsional Parasitoid Telur Trichogramma pretiosum Riley terhadap Telur Inang Corcyra cephalonica Stainton pada Pertanaman Kedelai”.
Pada kesempatan ini penulis mengucapkan terima kasih kepada:
1. Dr. Ir. Pudjianto, MSi. sebagai pembimbing, yang senantiasa sabar dalam membimbing dan mengarahkan selama penelitian dan penulisan skripsi ini. 2. Dr. Ir. Widodo, MS., selaku penguji tamu yang telah memberikan saran dan
kritik dalam penyusunan laporan ini.
3. Keluarga (Mama, Papa, my sister Ika, my brothers Fadli dan Fauzi) di Bekasi yang telah memberikan kasih sayang, perhatian, nasehat dan doa.
4. Seluruh anggota ( mbak Nita (alm), mbak Ada, mbak Diana, mbak Atik, dan lain- lain) Laboratorium Bioekologi Parasitoid dan Predator Departemen Proteksi Tanaman, Fakultas Pertanian yang telah membantu dalam penelitian dan penulisan skripsi ini.
5. Seluruh staf pengajar Departemen Proteksi Tanaman, Fakultas Pertanian yang telah memberikan ilmu selama penulis kuliah.
6. My best friends Mu’min, Okti, Rosita, Lia, Asti, dan Desmi yang telah memberikan arti persahabatan yang sebenarnya.
7. Kakak kelas dan rekan-rekan HPT 38 dari Departemen Proteksi Tanaman. 8. Kristal crew yang telah membantu dalam penulisan skripsi ini.
9. Semua pihak terkait yang telah membantu dalam penyelesaian skripsi ini. Penulis menyadari bahwa dalam penulisan skripsi ini, masih ada kekurangan. Oleh karena itu, kritik dan saran sangat diharapkan oleh penulis untuk perbaikan kegiatan selanjutnya. Semoga skripsi ini dapat bermanfaat. Amin.
Bogor, Februari 2006
DAFTAR ISI
DAFTAR TABEL
Nomor Halaman
Teks
1. Jumlah telur terparasit pada beberapa pelepasan ... 22 2. Jumlah telur terparasit pada kemapanan hidup T. pretiosum ... 23
Lampiran
1. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 3 MST ... 36 2. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 4 MST ... 37 3. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 5 MST ... 38 4. Hasil analisis data telur terparasit pada pelepasan parasitoid
saat tanaman berumur 6 MST ... 39 5. Hasil analisis data telur terparasit pada pelepasan parasitoid
DAFTAR GAMBAR
Nomor Halaman
Teks
1. Hubungan antara kepadatan telur inang C. cephalonica
dengan jumlah telur inang terparasit pada saat tanaman berumur 3 MST ... 18
2. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 4 MST ... 19
3. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 5 MST ... 19
4. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 6 MST ... 19
5. Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 7 MST ... 20
6. Grafik jumlah hama yang didapatkan selama sampling ... 25
7. Jumlah parasitoid yang terkumpul dari sampling ... 26
Lampiran 1. Petak percobaan dengan 18 tanaman berjarak 1 m dari titik pelepasan ... 33
2. Penempelan pias pada permukaan bawah daun ... 33
3. Tabung reaksi berisi parasitoid dengan posisi tegak lurus saat pelepasan parasitoid ... 33
4. Kondisi lahan saat generatif akhir ... 34
5. Imago betina T. pretiosum ... 34
6. Parasitoid larva C. chalcites famili Ichneumonidae ... 34
7. Parasitoid larva C. chalcites famili Braconidae ... 35
8. Parasitoid telur Ooencyrtus sp. famili Encyrtidae ... 35
9. Parasitoid larva C. chalcites famili Elasmidae spesies A ... 35
PENDAHULUAN
Latar belakang
Trichogrammatidae merupakan famili yang terkenal sebagai agens pengendali hayati berbagai serangga hama. Genera yang terkenal sebagai parasitoid telur adalah Trichogramma dan Trichogrammatoidea. Pemanfaatan parasitoid telur ini telah lama dilakukan dalam bidang pertanian dan kehutanan. Pemanfaatan Trichogramma dalam skala besar difokuskan pada tanaman pangan dan tanaman industri (Li 1994). Trichogramma digunakan untuk mengendalikan serangga-serangga hama penting yang merugikan secara ekonomi, seperti Ostrinia
sp. (Lepidoptera: Pyralidae), Chilo sp. (Lepidoptera: Pyralidae), dan Helicoverpa
sp. (Lepidoptera: Noctuidae).
Faktor utama yang mene ntukan keefektifan pemanfaatan Trichogramma
adalah pemilihan spesies yang sesuai, kualitas parasitoid yang dibiakkan, jumlah parasitoid yang dilepaskan, metode pelepasan, teknik pelepasan, iklim, waktu, dan integrasi dengan metode pengendalian lain. Pemilihan spesies parasitoid yang sesuai penting untuk mengendalikan serangga hama sasaran. Sebagai contoh, pada lahan pertanaman padi di Cina, T. japonicum merupakan spesies yang dominan dan menunjukkan kemampuan parasitisasi yang tinggi pada telur-telur Lepidoptera dibandingkan T. chilonis dan T. dendrolimi (Li 1994). Kualitas parasitoid berhubungan dengan lamanya parasitoid dibiakkan di laboratorium. Semakin lama parasitoid dibiakkan di laboratorium maka semakin menurun kualitas parasitoid tersebut. Jumlah parasitoid yang dilepaskan berkaitan dengan populasi serangga hama sasaran. Metode pelepasan yang umum digunakan ada dua macam yaitu inundasi dan inokulasi. Teknik pelepasan terdiri dari cara manual dan mekanik. Iklim dan waktu berkaitan dengan perilaku parasitoid dalam pencarian inang. Integrasi dengan metode pengendalian lain dapat mendukung keefektifan Trichogramma misalnya penanaman tanaman berbunga sebagai tanaman pinggir yang dapat menyediakan makanan bagi parasitoid.
mengendalikan penggerek batang kakao Conomorpha cramerella (Lepidoptera: Gracillariidae) dapat mencapai 63 - 86% di Sabah. Di Indonesia, Ramlan (2001) melaporkan bahwa pelepasan T. armigera populasi Cianjur pada pertanaman kedelai dapat menurunkan kerusakan polong kedelai sebesar 23,27% dan meningkatkan produksi sebesar 70 g biji kering dalam 10 rumpun, dengan tingkat parasitisasi sebesar 37,77%. Tingkat parasitisasi T. exiguum berkisar antara 67 - 83% dalam menekan Helicoverpa sp. pada pertanaman kapas di Carolina Utara (Suh 2000). Pemanfaatan Trichogramma sp. di Cina untuk mengendalikan
serangga hama Lepidoptera memiliki tingkat parasitisasi berkisar dari 6,5% - 98,9% (Li 1994).
Hubungan antara tingkat parasitisasi dengan kepadatan inang dikenal dengan istilah tanggap fungsional. Holling (1959 dalam Hassell 2000) menggolongkan tiga tipe tanggap fungsional, yaitu:
1. Tipe 1, hubungan bersifat konstan contohnya Encarsia citrina
(Hymenoptera: Aphelinidae) (Matadha et al. 2005), Daphnia magna
(Hassell 2000)
2. Tipe 2, hubungan yang bersifat logaritmik contoh Telenomus remus
(Hymenoptera: Scelionidae) (Widyarti 2003), T. ostriniae ( Hymenoptera: Trichogrammatidae) (Wang & Ferro 1998).
3. Tipe 3, hubungan dengan respon berbentuk sigmoid contoh Plea atomoria
(Hassell 2000).
Smith (1996) melaporkan bahwa pelepasan Trichogramma sp. mempengaruhi dinamika populasi inang dan komunitas serangga lain di sekitar lahan pelepasan. Akibat langsung pelepasan tersebut dapat dilihat pada serangga ordo Lepidoptera bukan sasaran di sekitar lahan pelepasan, sedangkan efek tidak langsung dapat dilihat pada kompleks musuh alami (Howarth 1991).
Buchori (2003) melaporkan bahwa tingkat parasitisasi T. pretiosum
berbeda pada suhu dan kepadatan inang yang berbeda dalam skala laboratorium. Studi tentang tanggap fungsional T. pretiosum di lapangan belum banyak dilakukan di Indonesia. Penelitian ini diharapkan dapat memberikan informasi mengenai kemampuan parasitisasi parasitoid T. pretiosum pada kepadatan inang
Tujuan
Penelitian ini bertujuan untuk mengetahui hubungan antara kemampuan parasitisasi parasitoid telur T. pretiosum dengan kepadatan inang C. cephalonica
TINJAUAN PUSTAKA
Parasitoid Telur Famili Trichogrammatidae
Famili Trichogrammatidae merupakan parasitoid telur yang bersifat generalis. Ciri khas Trichogrammatidae terletak pada tarsi yang beruas tiga (Doutt & Viggiani 1968; Grissell & Schauf 1990; Borror 1996). Struktur metasoma yang menempel pada mesosoma lebar dan fragma yang kedua menonjol jauh ke dalamnya (Borror 1996). Antena pendek dengan tujuh atau lebih ruas, menggada pada bagian pedicel, flagellum khas dengan satu atau dua segmen. Sayap depan memiliki rambut-rambut yang tersusun dalam baris (Doutt & Viggiani 1968; Borror 1996). Venasi sayap sigmoid, terdapat pola RS1
pada sayap depan, dan dorsal lamina berasosiasi dengan genitalia jantan. Famili Trichogrammatidae termasuk serangga yang sangat kecil dengan ukuran panjang tubuh berkisar antara 0,2 - 1,5 mm (Pinto & Stouthamer 1994).
Trichogramma dan Trichogrammatoidea sangat sulit diklasifikasikan karena perbedaan morfologi yang sangat kecil, memiliki sebaran yang luas dan strain yang bervariasi (Kalshoven 1981). Trichogramma memiliki morfologi sebagai berikut: seta fringe pada sayap lebih pendek, funikel dua ruas pada betina, gada antena jantan tidak beruas, ada venasi RS1 pada sayap depan, dan genitalia
jantan memiliki ‘dorsal gonobase expansion’. Morfologi khusus genus
Trichogrammatoidea adalah sebagai berikut: seta fringe pada sayap lebih panjang, funikel dua ruas pada jantan dan betina, antena jantan terdiri dari tiga ruas, tidak ada vein track RS1 pada sayap depan, dan genitalia jantan tanpa ‘dorsal gonobase
expansion’ (Buchori et al. 1998).
Penyebaran parasitoid ini sangat luas. Tiap negara memiliki jumlah spesies yang bervariasi. Pinto & Stouthamer (1994) melaporkan bahwa dari 145 spesies Trichogramma yang diakui, 114 spesies telah dideskripsikan. Beberapa spesies Trichogramma bahkan telah dibiakkan secara komersial, misalnya
T. pretiosum Riley, T. platneri Nagakarti, T. minutum Riley, T’toidea armigera
Nagaraja, T. ostriniae Pan & Chen, dan T. brassicae Bezd. (Losey & Calvin 1995; Vasquez et al. 1997)
Bioekologi Trichogramma pretiosum Riley
Trichogramma pretiosum adalah salah satu spesies parasitoid telur famili Trichogrammatidae yang berasal dari Amerika Utara (Smith 1996; Knutson 2002). Saat ini, distribusi parasitoid ini tersebar luas di dunia. Hal ini berkaitan dengan kemampuan yang baik parasitoid tersebut dalam beradaptasi terhadap lingkungan.
Di alam, T. pretiosum merupakan parasitoid alami pada telur Helicoverpa armigera. Parasitoid ini juga dapat memarasit telur-telur serangga hama lain seperti Sitotroga cerealella (Lepidoptera: Gelechiidae), Chilo spp. (Lepidoptera: Pyralidae), Manduca sexta (Lepidoptera: Sphingidae), Ephestia kuehniella (Lepidoptera: Pyralidae), Galleria mellonella (Lepidoptera: Pyralidae), Corcyra cephalonica (Lepidoptera: Pyralidae), Antheraea pernyi (Lepidoptera: Saturniidae), Samia cynthia (Lepidoptera: Saturniidae), Alabama argillacea (Lepidoptera: Noctuidae), Anticarsia spp. (Lepidoptera: Noctuidae),
Cadra cautella (Lepidoptera: Pyralidae), Diatraea spp. (Lepidoptera: Pyralidae),
Heliothis spp. (Lepidoptera: Noctuidae), Trichoplusia ni (Lepidoptera: Noctuidae), Plodia interpunctella (Lepidoptera: Pyralidae), dan
Scrobipalpula absoluta (Lepidoptera: Gelechiidae) (Li 1994; Greenberg et al.
1998; Monje et al. 1999).
imago betina memiliki rambut-rambut yang lebih sedikit dan pendek (Knutson 2002).
Tipe reproduksi T. pretiosum adalah arrhenotoky. Imago parasitoid betina yang berkopulasi akan memiliki keturunan jantan dan betina. Nisbah kelamin antara jantan dan betina adalah 1:2 (Kalshoven 1981).
Siklus hidup parasitoid berkisar antara 8 - 10 hari (Clausen 1940). Telur
T. pretiosum menetas 26 jam setelah telur inang diparasit. Pada hari kedua dan ketiga, telur inang yang terparasit berubah warna menjadi kehitaman karena larva parasitoid memakan isi telur inang secara internal. Selama instar ketiga, butiran melanin disimpan pada permukaan dalam korion telur inang. Hal tersebut menyebabkan telur inang berubah warna menjadi hitam (Knutson 2002). Hari keempat hingga ketujuh, telur inang akan menjadi hitam. Parasitoid berada pada tahap pupa yang berlangsung selama 108 jam (Strand 1985). Hari kedelapan, imago T. pretiosum akan keluar dari telur inang (Knutson 2002). Lama hidup imago berkisar antara 7 - 14 hari.
Tanggap Fungsional
Tanggap fungsional mengga mbarkan hubungan antara jumlah inang yang diparasit per parasitoid dan kepadatan inang (Wang & Ferro 1998). Tanggap fungsional merupakan komponen yang sangat esensial dari dinamika interaksi antara parasitoid dan inang serta sangat penting untuk determinasi stabilitas dari sistem yang dikelola (Oaten & Murdoch 1975 dalam Wang & Ferro 1998).
Holling (1959 dalam Hassell 2000) menggolongkan tanggap fungsional menjadi tiga tipe yaitu:
1. Tipe 1, hubungan bersifat konstan. Tipe 1 menggambarkan tingkat parasitisasi oleh parasitoid meningkat atau menurun sebanding dengan kepadatan inang sampai mencapai tahap kejenuhan. Serangga-serangga yang memiliki tanggap fungsional tipe 1 adalah Encarsia citrina (Matadha
et al. 2005), Daphnia magna (Hassell 2000).
kepadatan inang. Namun, dengan pertambahan kepadatan inang, tingkat parasitisasi mulai menurun. Tipe ini dimiliki oleh sebagian besar parasitoid seperti Telenomus remus (Widyarti 2003), T. ostriniae (Wang & Ferro 1998).
3. Tipe 3, hubungan dengan respon berbentuk sigmoid. Tipe ini menggambarkan tingkat parasitisasi pada awal peningkatan kepadatan inang lambat, terjadi peningkatan parasitisasi pada kepadatan inang yang makin meningkat serta tingkat parasitisasi berjalan lambat lagi pada kepadatan inang yang tinggi, contohnya Plea atomoria (Hassell 2000).
Holling (1959 dalam Hassell 2000) mengasumsikan bahwa tanggap fungsional tipe 2 merupakan khas invertebrata, sedangkan tanggap fungsional tipe 3 lebih cocok untuk vertebrata predator yang memiliki kemampuan untuk merespon peningkatan jumlah mangsa.
Tarumingkeng (1994) menyatakan bahwa tanggap fungsional tipe 1 dan 2 memiliki kestabilan yang rendah. Pada kedua tipe tersebut, tingkat parasitisasi semakin menurun dengan meningkatnya populasi inang. Tanggap fungsional tipe 3 lebih stabil jika dibandingkan tipe 1 dan 2 karena parasitoid memberikan pengaruh terpaut kerapatan bagi inang.
Studi-studi tanggap fungsional Trichogramma yang telah dilakukan di laboratorium dan di lapangan menunjukkan hasil yang tidak sama. Kfir (1983 dalam Wang & Ferro 1998) menunjukkan bahwa tanggap fungsional T. pretiosum
mengarah pada tipe 2. Di lapangan, tanggap fungsional parasitoid dapat berubah dari satu tipe ke tipe lainnya. Hal tersebut mungkin terjadi karena perubahan kondisi lingkungan terutama suhu (Kalyebi et al. 2005).
Ngengat Beras Corcyra cephalonica Stainton
gudang. Serangga ini dapat menyerang berbagai macam komoditas antara lain beras, tepung terigu, kopra, kacang-kacangan, kakao, bungkil dan lain- lain.
C. cephalonica memiliki metamorfosis yang lengkap (holometabola). Serangga ini meletakkan telur pada komoditas yang diserangnya. Telurnya berbentuk oval, licin, dan berwarna putih (Kalshoven 1981). Telur menetas dalam waktu 3 - 5 hari. Larva berwarna putih keabuan dengan panjang 17 mm (Kalshoven 1981). Larva terdiri dari 8 instar. Saat akan menjadi pupa, larva naik ke permukaan bahan makanan dan membuat kokon dengan menganyam benang sutra di antara butiran bahan makanan (Widodo 1987). Pupa berwarna coklat dan terbentuk dalam kokon berwarna putih yang kuat. Masa pupa berlangsung selama 12 hari. Imago berwarna abu-abu dengan panjang 7 - 12 mm. Ngengat bersifat nokturnal. Lama hidup imago berlangsung selama 10 hari. Nisbah kelamin
C. cephalonica adalah 1:1. Imago dapat menghasilkan 400 butir telur (Kalshoven 1981).
Parasitisasi Trichogramma pada Serangga Lain
Pemanfaatan Trichogramma sebagai agens pengendalian hayati telah lama dilakukan terutama dalam pelepasan secara inundasi (Orr et al. 2000). Li (1994) melaporkan bahwa 32 juta ha lahan pertanian dan kehutanan menggunakan
Trichogramma sebagai agens pengendali hayati di dalamnya.
Pemanfaatan Trichogramma yang luas dan dalam jumlah yang besar memiliki pengaruh potential terhadap serangga non sasaran di lahan pelepasannya. Dampak langsung dapat terlihat pada serangga ordo Lepidoptera. Hal ini terjadi pada pelepasan T. brassicae yang memarasit kupu-kupu Karner blue (Andow et al. 1995 dalam Orr etal. 2000).
Trichogramma merupakan parasitoid yang bersifat generalis. Parasitoid ini dapat memarasit 400 spesies serangga dalam 203 genera, 44 famili, dan 7 ordo ( Bao & Chen 1989 dalam Li 1994). Namun demikian, ada kecenderungan bahwa
Trichogramma memiliki preferensi yang kuat pada inang, tanaman, dan kondisi tertentu (Hassan and Gou 1991 dalam Orr et al. 2000). Trichogramma mungkin akan memiliki kisaran inang yang terbatas karena preferensi tersebut, sehingga dapat menurunkan potensi parasitisasi Trichogramma pada serangga non target.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan mulai bulan Desember 2004 - Juli 2005. Pelepasan parasitoid telur T. pretiosum dilakukan pada pertanaman kedelai di kebun percobaan IPB Cikabayan, Dramaga, Bogor. Pemeliharaan parasitoid telur
T. pretiosum dan identifikasi parasitoid lain yang tertangkap dari hasil pengambilan contoh dilakukan di Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Bahan dan Alat
Parasitoid yang digunakan adalah T. pretiosum yang berasal dari populasi Jambi. T. pretiosum yang digunakan berasal dari telur H. armigera pada tanaman kapas dan merupakan generasi ± ke - 94. Asumsi nisbah kelamin T. pretiosum
Peralatan yang digunakan dalam penelitian adalah bambu ajir sebagai penanda tanaman kedelai yang akan ditempelkan pias dan titik pelepasan parasitoid. Tabung reaksi ukuran 30 mm x 200 mm yang berguna sebagai tempat pemeliharaan parasitoid dan penyimpanan telur dan larva yang diambil dari lapangan. Cawan petri sebagai tempat penyimpanan pias yang disterilkan dan pias yang dibawa ke lapangan. Gunting digunakan untuk memotong kertas karton manila sesuai dengan yang diinginkan dan untuk memotong pita. Kuas berfungsi untuk mengambil parasitoid saat membedakan antara parasitoid betina dan jantan dan membersihkan tabung reaksi sebelum dicuci. Pinset digunakan untuk memasukkan pias dalam tabung reaksi dan mengambil pias yang telah berada dalam tabung reaksi selama 24 jam. Lemari pendingin (freezer) sebagai tempat mensterilkan telur C. cephalonica. Mikroskop untuk mengamati telur-telur yang terparasit. Jarum digunakan untuk menusuk telur yang tidak terparasit. Lampu duduk berfungsi sebagai penerangan saat pengamatan. Oven berfungsi untuk mensterilkan media hidup C. cephalonica.
Metode Pemeliharaan Serangga Inang C. cephalonica
Media campuran pakan ayam dan dedak disterilkan dengan oven untuk mematikan organisme kontaminan yang terbawa dalam media. Media dengan
ketebalan ± 3 cm kemudian dimasukkan dalam kotak pemeliharaan larva
C. cephalonica. Telur atau larva C. cephalonica dimasukkan ke dalam kotak pemeliharaan yang telah berisi media dengan kepadatan 2 - 3 telur atau ulat per cm². Kotak pemeliharaan ditutup dan disimpan pada rak penyimpanan dengan
suhu kamar sampai imago C. cephalonica muncul. Kemunculan imago
C. cephalonica memerlukan waktu selama 5 - 7 minggu. Imago yang muncul diambil dengan menggunakan tabung reaksi, lalu dikumpulkan dan dimasukkan ke dalam kotak peneluran. Kotak peneluran disimpan pada posisi tegak dan
dialasi dengan kertas buram. Selama dalam kotak peneluran, imago
C. cephalonica tidak memerlukan makanan. Telur-telur C. cephalonica akan
diletakkan pada bagian atas dan dasar kotak peneluran. Telur-telur
kassa disikat dengan kuas dan ditampung sementara di atas kertas buram. Setelah terkumpul, telur-telur tersebut dibersihkan dari sisa-sisa kotoran dan imago yang mati dengan cara mengalirkan telur-telur tersebut pada selembar kertas buram dan ditampung pada kertas buram lain. Hal tersebut dilakukan berulang kali hingga kotoran dan sisa-sisa imago tidak ada lagi. Telur-telur yang telah bersih dimasukkan dalam cawan petri, kemudian digunakan untuk pembiakan parasitoid dan sebagian lagi untuk pembiakkan C. cephalonica.
Perbanyakan Parasitoid Telur Trichogramma pretiosum
Telur C. cephalonica direkatkan pada pias dengan cara menaburkan telur secara merata pada pias yang telah diberi gum arabic. Pias berukuran 2 cm x 2 cm dan dapat menampung ± 1000 butir telur. Telur C. cephalonica pada pias dimasukkan dalam lemari pendingin (freezer) selama ± 120 menit untuk mematikan telur tersebut. Bila telur tidak dimatikan, telur akan menetas dan larva dapat memakan telur-telur yang terparasit. Pias dengan telur steril dimasukkan dalam tabung reaksi (satu pias untuk satu tabung) yang di dalamnya telah dimasukkan imago parasitoid. Jumlah imago parasitoid ya ng berada pada tabung reaksi berjumlah ± 500 ekor. Tabung reaksi ditutup dengan tisu dan diikat dengan karet. Tabung reaksi disimpan pada rak dengan posisi mulut tabung menjauhi arah datangnya sinar. Setelah ± 24 jam, pias telur yang berada dalam tabung reaksi diambil dan dipindahkan ke tabung reaksi lain. Telur pada pias yang terparasit akan berubah warna menjadi hitam pada hari ke-3. Imago parasitoid akan muncul 8 - 10 hari setelah telur C. cephalonica terparasit. Setiap hari, imago parasitoid diberikan pakan madu pada sisi tabung reaksi dengan menggunakan jarum (cukup dengan sedikit olesan saja).
Persiapan Lahan Kedelai
pengolahan tanah, dengan dosis kapur dan pupuk kandang masing- masing sebesar 2.000 kg/ha dan 20.000 kg/ha.
Dua benih dimasukkan dalam satu lubang. Benih kedelai yang ditanam adalah varietas Willis yang berasal dari Balitro. Kedelai ditanam dengan jarak tanam 25 cm x 25 cm. Pemupukan pertama (urea, TSP, dan KCl) dilaksanakan secara bersamaan di sekitar lubang benih, dengan dosis berturut-turut 83,33 kg/ha, 150 kg/ha, dan 150 kg/ha. Aplikasi kedua dengan urea dilakukan menjelang tanaman berbunga (6 MST - 8 MST). Setelah seminggu, penyulaman dilakukan terhadap tanaman yang mati. Pengairan dilakukan secara manual. Penyiangan dilakukan sebanyak dua kali yaitu pada saat tanaman berumur 2 - 3 MST dan tanaman mulai berbunga (6 MST - 8 MST). Lahan pertanaman kedelai yang luasnya 600 m², dibagi menjadi tiga petak percobaan yang masing- masing berukuran 10 m x 20 m.
Studi Tanggap Fungsional T. pretiosum
Studi tanggap fungsional T. pretiosum dilakukan dengan memasang pias-pias yang berisi berbagai tingkat kepadatan telur C. cephalonica pada pertanaman kedelai, kemudian mengamati tingkat parasitisasi telur pada pias-pias tersebut oleh T. pretiosum. Untuk menambahkan populasi T. pretiosum di pertanaman kedelai, maka dilepaskan imago parasitoid pada setiap pemasangan pias.
Pemasangan pias-pias telur dilakukan pada pagi hari sekitar pukul 06:30 WIB. Pias dengan berbagai kepadatan telur yang berbeda dipasang pada tanaman kedelai yang dipilih secara melingkar dengan jarak 1 m dari titik pelepasan. Titik pelepasan parasitoid terletak pada bagian tengah setiap petak percobaan. Sebanyak 18 tanaman pada masing- masing petak yang terpilih ditandai dengan bambu ajir (Gambar lampiran 1). Pemilihan tanaman dilakukan saat tanaman berumur ± 2 MST.
Koleksi pias-pias telur perangkap dilakukan ± 8 jam setelah pelepasan parasitoid. Koleksi tersebut dibawa ke laboratorium dan disimpan dalam tabung reaksi pada suhu ruang. Pengamatan dilakukan setiap hari untuk melihat apakah ada telur yang terparasit atau telur ya ng menetas sebagai larva.
Pelepasan parasitoid dilaksanakan sejak tanaman berumur 3 MST. Pelepasan dilakukan sebanyak lima kali dengan selang waktu satu minggu. Jumlah parasitoid yang dilepas adalah 2.000 individu per petak. Tabung reaksi yang berisi parasitoid direkatkan pada bambu ajir dengan posisi tegak lurus (Gambar lampiran 3), lalu tutup tabung dibuka secara bersamaan antar petak. Pelepasan parasitoid dilaksanakan pada pagi hari sekitar pukul 07:00 WIB.
Kemapanan T. pretiosum dan Pengaruhnya pada Serangga Lain
Pelepasan parasitoid terakhir dilakukan ketika tanaman berumur 8 MST. Namun, pemasangan pias-pias perangkap tetap dilaksanakan untuk melihat apakah parasitoid yang dilepas mampu bertahan (mapan) atau tidak. Pemasangan pias-pias dilakukan sebanyak 4 kali dengan interval waktu satu minggu. Proses pemasangan pias dan pemaparannya di lapangan sama dengan proses saat pelepasan parasitoid.
Rancangan Percobaan
Penelitian dilakukan dengan menggunakan rancangan acak kelompok (RAK). Perlakuannya adalah kepadatan telur C. cephalonica yang berbeda (20, 40, 80, 160, 320, 640 butir) yang masing- masing terdiri dari tiga ulangan. Data yang diperoleh dianalisis menggunakan program SAS dan selanjutnya dilakukan uji Duncan pada taraf 5%. Analisis regresi sederhana digunakan untuk melihat hubungan antara jumlah telur terparasit dan kepadatan telur.
HASIL DAN PEMBAHASAN
Tanggap Fungsional Trichogramma pretiosum
Interaksi antara parasitoid- inang merupakan salah satu faktor penting dalam menentukan keberhasilan parasitoid sebagai agens hayati. Perubahan populasi inang akan mempengaruhi populasi parasitoid. Salah satu interaksi inang-parasitoid adalah tanggap fungsional. Tanggap fungsional pada intinya menggambarkan hubungan antara jumlah inang/mangsa yang diparasit/dimakan oleh parasitoid/predator (Hassell 2000).
Grafik-grafik tanggap fungsional parasitoid T. pretiosum pada inang
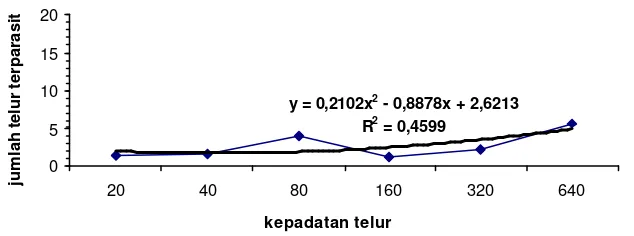
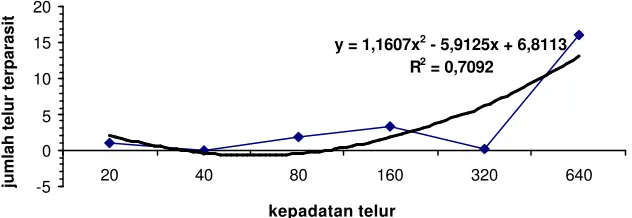
C .cephalonica menunjukkan bahwa kepadatan telur inang berpengaruh terhadap jumlah telur inang yang terparasit. Hal tersebut dapat dilihat pada gambar 1, 3, 4, dan 5, dimana pada kepadatan telur 640 jumlah telur terparasit yang tertinggi. Pada gambar 2, grafik hubungan antara kepadatan telur dengan jumlah telur yang terparasit cenderung menurun dengan nilai R2= 0.287. Meskipun nilainya kecil, hubungan antara kepadatan telur inang dengan jumlah telur terparasit berkorelasi positif.
y = -0,1162x2
Gambar 2 Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada tanaman berumur 4 MST
Gambar 3 Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 5 MST
y = 0,2102x2
y = 1,1607x2 - 5,9125x + 6,8113
Gambar 5 Hubungan antara kepadatan telur inang C. cephalonica dengan jumlah telur inang terparasit pada saat tanaman berumur 7 MST
Grafik 1,4,5 menunjukkan adanya kecenderungan tanggap fungsional tipe 3. Hal tersebut dapat dilihat dari peningkatan jumlah telur terparasit yang tinggi pada populasi inang yang makin tinggi. Peningkatan populasi inang yang cukup tinggi membuat jumlah inang yang diparasit pun makin meningkat. Oaten and Murdoch (1975 dalam Bernstein 2000) tanggap fungsional tipe 3 memiliki kestabilan sistem inang-parasitoid. Kestabilan dapat dicapai jika parasitoid yang digunakan adalah parasitoid polifag yang mampu beralih mangsa (Tarumingkeng 1994).
Gambar 2 menunjukkan bahwa T. pretiosum memiliki tanggap fungsional tipe 2. Tanggap fungsional tipe 2 memiliki ”handling time” yaitu interval waktu antara pencarian inang pertama dan keseluruhan pencarian inang-inang lainnya (masa penanganan inang). Waktu yang tersedia untuk pencarian inang berkurang karena handling time, akibatnya respon meningkat dan kemudian menurun sejumlah inang yang diparasit (kisaran inang tertentu). Parasitoid biasanya bergerak secara bebas diantara patch dalam lingkungan patch, sehingga penting mempertimbangkan waktu yang parasitoid habiskan dalam patch dengan kepadatan inang yang berbeda.
Berdasarkan model perilaku, spesies parasitoid yang memperlihatkan tanggap fungsional tipe 2 diperkirakan tidak mampu mengatur populasi hama yang menjadi inangnya (Hassell 2000). Berapapun jumlah inang maka laju parasitisasinya akan tetap sama, yang berarti tidak ada perubahan tanggap fungsional parasitoid terhadap kepadatan inang.
kepadatan telur kemudian mencapai titik maksimum, tingk at parasitisasi selanjutnya akan bersifat konstan.
Interaksi inang-parasitoid pada tanggap fungsional tipe 1 dan 2 menunjukkan kestabilan yang rendah, karena pada kedua tipe tersebut tingkat parasitisasi menurun dengan meningkatnya populasi inang (Tarumingkeng 1994). Tanggap fungsional tipe 3 memiliki interaksi inang-parasitoid yang lebih stabil dibandingkan tipe 1 dan 2. Pada tipe 3, terdapat kecenderungan parasitoid memarasit meningkat pada populasi inang yang lebih banyak, maka peluang tingkat parasitisasi akan meningkat. Parasitoid yang memiliki tanggap fungsional tipe 3 di alam menunjukkan bahwa parasitoid tersebut secara alami dapat mengendalikan inangnya.
Tipe tanggap fungsional yang dapat berubah pada parasitoid telur
T. pretiosum menunjukkan bahwa pemanfaatan parasitoid tersebut akan memerlukan bantuan manusia untuk mendukung keefektifannya. Perencanaan yang matang seperti waktu pelepasan, tipe tanaman, jenis hama dan lain- lain diperlukan untuk mendukung keberhasilan pemanfaatan T. pretiosum.
Perubahan tipe tanggap fungsional dari tipe satu ke lainnya dapat disebabkan oleh perubahan perilaku parasitoid dalam penerimaan inang pada suhu yang berbeda, kemampuan reproduksi, dan lama hidup parasitoid betina. Perbedaan tipe tanggap fungsional tiap pelepasan dapat terjadi karena kondisi lapangan yang terus berubah (Wang & Ferro 1998). Fluktuasi hujan dan intensitas cahaya di lapangan dapat mempengaruhi aktifitas dan perilaku parasitoid yang kemudian juga mempengaruhi tingkat parasitisasi (Wang & Ferro 1998).
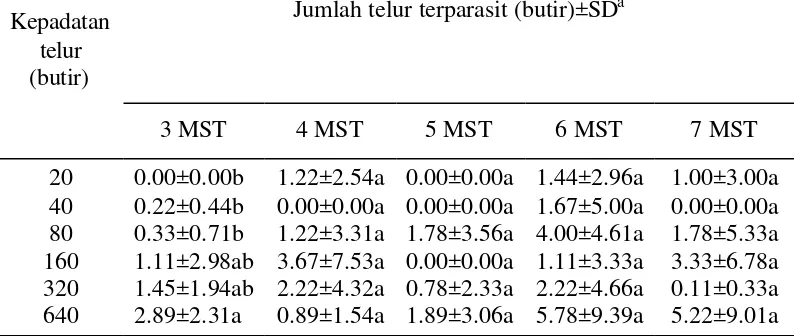
Jumlah telur terparasit pada pelepasan ke-2, 3, 4, dan 5 tidak berbeda nyata
pada kepadatan telur inang yang berbeda (P2 = 0.42; Tabel lampiran 2), (P3 = 0.11; Tabel lampiran 3), (P4 = 0.43 ; Tabel lampiran 4), (P5 = 0.21; Tabel
lampiran 5). Rata-rata pelepasan menunjukkan tingkat parasitisasi yang tertinggi pada kepadatan telur 640 butir.
Tabel 1 Jumlah telur terparasit pada berbagai kepadatan telur perangkap oleh T. pretiosum yang dilepaskan pada berbagai umur tanaman kedelai Kepadatan
telur (butir)
Jumlah telur terparasit (butir)±SDa
3 MST 4 MST 5 MST 6 MST 7 MST
20 0.00±0.00b 1.22±2.54a 0.00±0.00a 1.44±2.96a 1.00±3.00a 40 0.22±0.44b 0.00±0.00a 0.00±0.00a 1.67±5.00a 0.00±0.00a 80 0.33±0.71b 1.22±3.31a 1.78±3.56a 4.00±4.61a 1.78±5.33a 160 1.11±2.98ab 3.67±7.53a 0.00±0.00a 1.11±3.33a 3.33±6.78a 320 1.45±1.94ab 2.22±4.32a 0.78±2.33a 2.22±4.66a 0.11±0.33a 640 2.89±2.31a 0.89±1.54a 1.89±3.06a 5.78±9.39a 5.22±9.01a
a
Rataan pada lajur yang sama yang diikuti oleh huruf yang sama menunjukkan tidak berbeda nyata (uji duncan, a=0,05)
Rendahnya tingkat parasitisasi pada tabel 1 dapat disebabkan oleh berbagai faktor seperti faktor biologi, fisik lingkungan, dan kimia. Berbagai faktor tersebut di atas mempengaruhi perilaku parasitoid dalam tahap penemuan inang, yang berpengaruh terhadap tingkat parasitisasi parasitoid.
Faktor biologi, fisik lingkungan, dan kimia sangat mempengaruhi parasitoid dalam menemukan inang. Faktor biologi seperti lama hidup, kebugaran, lama generasi, ukuran tubuh, dan lain- lain mempengaruhi aktifitas parasitoid di lapangan. Faktor fisik lingkungan seperti cahaya, embun, kelembaban, suhu, warna dan bentuk tanaman dapat dijadikan sinyal oleh parasitoid untuk menemukan habitat inangnya. Kalyebi et al. (2005) menyatakan bahwa suhu memiliki pengaruh yang signifikan terhadap tanggap fungsional.
Wang and Ferro (1998) melaporkan bahwa tanggap fungsional T. ostriniae
oleh parasitoid. Perubahan suhu mempengaruhi aktifitas perilaku parasitoid misalnya dalam penanganan dan laju pencarian inang. Peningkatan suhu sampai pada suhu tertentu dapat menurunkan masa penanganan inang. Laju parasitisasi akan meningkat dengan meningkatnya suhu sampai batas tertentu karena interval waktu parasitisasi menjadi lebih pendek. Faktor kimia seperti senyawa kimia yang dikeluarkan tanaman dapat menjadi tanda bagi parasitoid dalam menemukan inangnya.
Arsitektur lahan (ukuran, bentuk, kepadatan) dan morfologi tanaman (tulang daun, rambut, bentuk jaringan) juga mempengaruhi perilaku parasitoid dalam pencarian inang dan keberhasilannya dalam parasitisasi (Bigler et al. 1997). Keragaman fisik dari suatu habitat mempengaruhi tanggap fungsional parasitoid terhadap kepadatan inang. Hal ini berkaitan dengan habitat sebagai tempat berlindung dan sumber makanan alternatif parasitoid. Perubahan bentuk dan ukuran tanaman dapat mempengaruhi laju pencarian dan kemampuan bertahan parasitoid di tanaman tersebut.
Kemapanan Parasitoid T. pretiosum
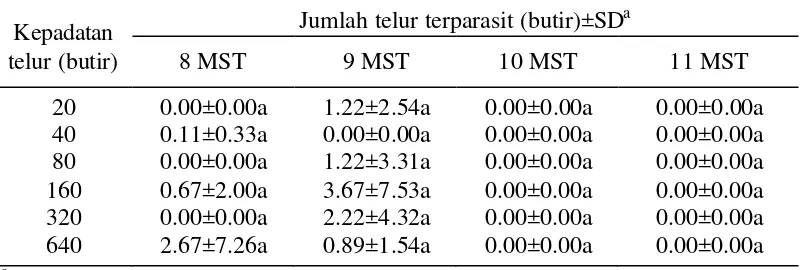
Parasitisasi oleh parasitoid yang dilepas pada telur perangkap setelah pelepasan terakhir dapat menunjukkan kemapanan parasitoid yang dilepaskan. Tabel 2 memperlihatkan bahwa tidak ada perbedaan yang nyata dalam jumlah telur yang terparasit pada telur perangkap dengan kepadatan yang berbeda.
Tabel 2 Jumlah telur terparasit pada berbagai kepadatan telur perangkap pada berbagai umur tanaman kedelai (tanpa pelepasan parasitoid)
Kepadatan telur (butir)
Jumlah telur terparasit (butir)±SDa
8 MST 9 MST 10 MST 11 MST
20 0.00±0.00a 1.22±2.54a 0.00±0.00a 0.00±0.00a 40 0.11±0.33a 0.00±0.00a 0.00±0.00a 0.00±0.00a 80 0.00±0.00a 1.22±3.31a 0.00±0.00a 0.00±0.00a 160 0.67±2.00a 3.67±7.53a 0.00±0.00a 0.00±0.00a 320 0.00±0.00a 2.22±4.32a 0.00±0.00a 0.00±0.00a 640 2.67±7.26a 0.89±1.54a 0.00±0.00a 0.00±0.00a
a
Kemapanan parasitoid di lapangan dipengaruhi oleh kemampuan pencarian inang, faktor lingkungan, kualitas parasitoid dan keberadaan nutrisi (Hassan 1994). Tabel 2 menunjukkan bahwa parasitoid yang dilepaskan hanya bertahan pada saat tanaman berumur 8 MST dan 9 MST atau dua minggu setelah pelepasan parasitoid terakhir. Hal tersebut sesuai dengan lama hidup parasitoid yang berkisar antara 7 - 14 hari.
Kemapanan parasitoid di lapangan juga berhubungan dengan kondisi tanaman yang telah berada pada masa generatif akhir dan tidak tersedianya inang dan pakan sehingga parasitoid diperkirakan terbang menuju tempat dimana terdapat inang, pakan, dan perlindungan untuk dapat bertahan (Hassan 1994). Saat tanaman memasuki masa generatif akhir (Gambar lampiran 4), terdapat pertanaman jagung ya ng berdekatan dengan lahan pelepasan. Tanaman jagung merupakan habitat yang sesuai bagi perkembangan serangga-serangga hama Lepidoptera seperti Helicoverpa armigera dan Ostrinia sp. Serangga-serangga hama tersebut merupakan inang Trichogramma, sehingga ada dugaan bahwa parasitoid akan terbang ke pertanaman jagung.
Perpindahan parasitoid di pertanaman banyak dipengaruhi oleh pergerakan angin dan kemampuan terbang. Penerbangan Trichogramma dapat dibagi tiga macam yaitu jarak pendek, perpindahan lokal, dan penerbangan jarak jauh (Bigler et al. 1997).
Serangga parasitoid mampu melakukan penerbangan rata-rata 5 - 8 kali/menit bahkan dapat meningkat sampai 10 kali atau lebih pada saat angin tenang (Speight et al. 1999 dalam Usyati 2003). Tingkat parasitisasi parasitoid telur T. pretiosum pada inang C. cephalonica di lapangan pada arah yang berlawanan dengan arah angin sebesar 76,97%, sedangkan yang menyebar searah dengan arah angin hanya 23,03% (Usyati 2003). Penyebaran serangga tidak selamanya searah dengan arah angin, karena ada faktor lain yang lebih menarik untuk serangga yaitu adanya sumber makanan yang terletak berlawanan arah dengan arah angin.
Faktor-faktor eksternal dan internal mempengaruhi keberhasilan pelepasan parasitoid di lapangan. Faktor eksternal yang berpengaruh adalah faktor lingkungan (temperatur, angin, curah hujan, kelembaban, dan intensitas cahaya) dan faktor musuh alami, sementara faktor internal adalah kebugaran dan pemilihan spesies parasitoid yang dilepas. Predator utama Trichogramma sp. adalah semut, larva Chrysopid, larva dan imago Coccinellid ( Usyati 2003).
Parasitisasi T. pretiosum pada Serangga Lain
Pelepasan parasitoid T. pretiosum mempunyai pengaruh terhadap serangga lain terutama ordo Lepidoptera yang berada dalam pertanaman kedelai. Hal ini berkaitan dengan sifat generalis dari parasitoid T. pretiosum.
0
Gambar 6 Grafik jumlah hama yang didapatkan selama sampling
Gambar 6 memperlihatkan bahwa jumlah serangga hama yang berada di lahan relatif sedikit. Pada sampling pertama saat tanaman berumur 8 MST telah didapatkan kelompok telur famili Limacodidae dan ulat penjalin daun
Lamprosema indicata (Lepidoptera: Pyralidae) terparasit. Kelompok telur tersebut berjumlah 10 butir, namun hanya 2 butir telur yang terparasit. Sampling ke-2 saat tanaman berumur 9 MST menemukan satu telur Helicoverpa armigera
berumur 11 MST hanya didapatkan dua telur R. linearis terparasit dan kutu daun
Aphid craccivora (Hemiptera: Aphididae) yang terparasit.
Pada masa vegetatif, hama-hama yang dominan ditemukan adalah
L. indicata, A. craccivora, kumbang daun Phaedonia inclusia (Coleoptera: Chrysomelidae), belalang Valanga spp. (Orthoptera: Acrididae), wereng hijau
Empoasca sp.(Homoptera: Cicadellidae), thrips Frankliniella (Thysanoptera: Terebrantia), dan tungau (Acarina: Tetranychidae). Kepik R. linearis dan Nezara viridula (Hemiptera: Pentatomidae) merupakan hama-hama yang mendominasi saat fase generatif.
Sampling dilakukan pada saat tanaman berada pada fase generatif, sehingga hama-hama yang terkoleksi pun sebagian besar hama yang berada pada fase tersebut. Namun pada fase tersebut terdapat hama yang dapat dijumpai pada fase vegetatif seperti L. indicata. Keberadaan L. indicata didukung oleh ketersediaan makanannya, karena L. indicata menyerang hanya bagian daun saja (Kalshoven 1981). Pada awalnya sampling hanya dilakukan untuk mengambil telur-telur hama yang berada pada pertanaman kedelai, akan tetapi dalam sampling ditemukan L. indicata yang terparasit sehingga ikut serta dalam sampling.
Telur-telur hama yang dapat dikoleksi yaitu: telur ordo Lepidoptera dari famili Limacodidae, famili Geometridae dan famili Noctuidae (H. armigera), ordo
Hemiptera dari famili Alydidae (R. linearis) dan famili Pentatomidae (N. viridula). Telur-telur ordo Lepidoptera yang dikoleksi dan terparasit sebagian
besar diparasit oleh parasitoid telur T. pretiosum. Sedangkan telur-telur ordo Hemiptera terparasit, akan muncul parasitoid telur Ooencyrtus sp (Hymenoptera: Encyrtidae).
0
Gambar 7 Jumlah parasitoid yang terkumpul dari sampling
Keragaman jenis parasitoid yang berada di pertanaman kedelai relatif tinggi. Ada dua jenis parasitoid yang berada dalam lahan yaitu parasitoid telur dan parasitoid larva. Gambar 7 memperlihatkan jenis dan jumlah parasitoid yang ditemukan pada pertanaman kedelai. Parasitoid yang ditemukan berasal dari famili Ichneumonidae (Gambar lampiran 6), Braconidae (Gambar lampiran 7), Elasmidae, Trichogrammatidae dan Encyrtidae (Gambar lampiran 8). Parasitoid larva tergolong dalam famili Ichneumonidae, Braconidae, dan Elasmidae. Parasitoid telur yang muncul tergolong dalam famili Trichogrammatidae dan Encyrtidae.
Identitifikasi hingga tingkat famili pada parasitoid larva L. indicata
menemukan parasitoid dari famili Ichneumonidae, Braconidae, dan Elasmidae. Pada famili Elasmidae terdapat dua spesies yang berbeda (Gambar lampiran 9 dan 10). Tingkat parasitisasi parasitoid larva oleh Ichneumonidae, Braconidae, Elasmidae spesies A dan Elasmidae spesies B berturut-turut adalah 12,5%; 12,5%; 25% dan 50%.
Parasitoid telur yang keluar berhasil diidentifikasi hingga tingkat spesies yaitu T. pretiosum dan Ooencyrtus sp. Telur R. linearis terparasit oleh parasitoid
telur Ooencyrtus sp. dengan tingkat parasitisasi 66,67%. Parasitoid telur
T. pretiosum muncul dari telur-telur ordo Lepidoptera.
yang terparasit adalah T. pretiosum sesuai dengan parasitoid telur yang dilepaskan. Tingkat parasitisasi T. pretiosum pada telur-telur famili Limacodidae dan H. armigera berturut-turut 20% dan 100%.
Kemunculan T. pretiosum pada telur-telur ordo Lepidoptera berkaitan dengan preferensi Trichogramma yang lebih menyukai inang telur-telur ordo Lepidoptera. Komponen sex feromon yang dikeluarkan ngengat memiliki pengaruh terhadap kemampuan pencarian inang T. pretiosum, T. brassicae, dan
T. evanescens (Lewis et al. 1982; Kaiser et al. 1989; Noldus 1989 dalam Nordlund 1994).
Pelepasan parasitoid memiliki pengaruh terhadap dinamika populasi inang yang terdapat di pertanaman kedelai dimana parasitoid dilepas. Populasi inang
T. pretiosum yang utama di lahan pertanaman kedelai adalah H. armigera. Dari gambar 6 dapat dilihat bahwa jumlah telur-telur H. armigera yang terkoleksi relatif sedikit, tetapi tingkat parasitisasinya mencapai 100%. Hal yang sama juga dilaporkan oleh Llewellyn (2002) bahwa pelepasan T. pretiosum di pertanaman kedelai memiliki tingkat parasitisasi yang cukup tinggi yaitu 81%. Pelepasan T. pretiosum ternyata mempengaruhi jumlah H. armigera yang berada di lahan.
KESIMPULAN DAN SARAN
Kesimpulan
Tipe tanggap fungsional T. pretiosum dapat berubah dari tipe satu ke tipe lainnya. Perubahan ini dapat disebabkan oleh kondisi lingkungan pertanaman
kedelai yang berubah. Tanggap fungsional tipe 3 memiliki interaksi inang-parasitoid yang lebih stabil dibandingkan tipe 1 dan 2.
Kemapanan T. pretiosum pada pertanaman kedelai berlangsung selama 2 minggu. Ketersediaan inang dan pakan mempengaruhi kemapanan T. pretiosum
di pertanaman kedelai. Penyebaran parasitoid pada pertanaman kedelai dipengaruhi oleh pergerakan angin dan kemampuan terbang.
Pada pertanaman kedelai ditemukan parasitoid larva dan parasitoid telur. Parasitoid larva L. indicata tergolong dalam famili Ichneumonidae, Braconidae,
dan Elasmidae. Parasitoid telur yang muncul adalah Ooencyrtus sp. dari telur
R. linearis dan T. pretiosum. Pengaruh pemanfaatan T. pretiosum secara langsung dapat dilihat pada telur-telur ordo Lepidoptera.
Saran
Penelitian ini dapat dilanjutkan dengan pemasangan inang utama
DAFTAR PUSTAKA
Bernstein C. 2000. Host-parasitoid models: the story of a successful failure. Di dalam: Hochberg ME, Ives AR, editor. Parasitoid Population Biology. New Jersey: Princeton University Press. hlm 41-57.
Bigler F, Suverkropp BP, Cerutti F. 1997. Host searching by Trichogramma and its implications for quality control and release techniques. Di dalam: Andow DA, Ragsdale DW, Nyvall RF. editor. Ecological Interactions and Biological Control. USA: Westview Press. hlm 240-251.
Borror DJ, Triplehorn CA, Johnson NF. 1996. Pengenalan Pelajaran Serangga. Penerjemah: Partosoedjono S. Terjemahan dari: An Introduction To The Study Of Insects. Yogyakarta: Gadjah Mada University Press.
Buchori D. 2003. Tanggap fungsional, fluktuasi asimetri, jumlah betina penemu dan ketersediaan pakan: kajian ekologi reproduksi serta implikasinya bagi keberhasilan pelepasan parasitoid telur Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Laporan penelitian Hibah Bersaing Perguruan Tinggi Tahun Anggaran 2003/2004. Bogor: IPB.
Buchori D, Hidayat P, Kartosuwondo U, Harahap IS, Nurmansyah A. 1998. Dinamika interaksi antara parasitoid Trichogramma (Hymenoptera: Trichogrammatidae) dan inangnya: faktor-faktor yang berpengaruh terhadap kualitas Trichogramma sebagai agens pengendalian hayati. Laporan Kemajuan Hibah Bersaing Perguruan Tinggi VII/I Tahun Anggaran 1998/1999. Direktorat Jenderal Pendidikan Tinggi. Depdikbud. Bogor: IPB.
Clausen CP. 1940. Entomophagous Insects. New York: McGraw Hill Book Company.
Doutt RL, Viggiani G. 1968. The Classification of the Trichogrammatidae (Hymenoptera: Chalcidoidea). Proceedings of the California. Academy of Sciences ed ke-4 35(20): 477-586.
Greenberg SM, Nordlund DA, Wu Z. 1998. Influence of rearing host on adult size and ovipositional behavior of mass produced female Trichogramma minutum Riley and Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Biol Con 11: 43-48.
Hassan SA. 1994. Strategies to select Trichogramma species for use in biological control. Di dalam: Wajnberg E, Hassan SA, editor. Biological Control With Egg Parasitoids. UK: CAB International. hlm 55-72.
Hassell, Michael P. 2000. The Spatial and Temporal Dynamics of Host-Parasitoid Interactions. New York: Oxford University Press.
Howarth FG. 1991. Environmental impacts of classical biological control. Annu Rev Entomol. 36: 485-509.
Kalshoven LGE. 1981. The Pest of Crops in Indonesia. Penerjemah: PA van der Laan. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesie. Jakarta: Ichtiar Baru-Van Hoeve.
Kalyebi A, Overholt WA, Schulthess F, Mueke JM, Hassan SA, Sithanantham S.
2005. Functional response of six indigenous trichogrammatid egg parasitoids (Hymenoptera: Trichogrammatidae) in Kenya: influence of temperature and relative humidity. Biol Con 32: 164-171.
Knutson A. 2002. The Trichogramma Manual. The Texas Agricultural University System. (http://www.insects.tamu.edu/extension/bulletins/b-6071 html) [31 Juli 2005]
Li YL. 1994. Worldwide use of Trichogramma for biological control on different crops: a survey. Di dalam: Wajnberg E, Hassan SA, editor. Biological Control With Egg Parasitoids. UK: CAB International. hlm 37-54.
Liau SS. 1991. The IPM experience in plantation crops. Di dalam: Integrated Pest Management in The Asia Pasific Region. Proceeding of the Conference on Integrated Pest Management in The Asia Pasific Region; Kualalumpur, 23-27 September 1991. Malaysia: CAB International. Asian Development Bank.
Llewellyn R. 2002. Releases of Trichogramma pretiosum in soybeans. (http://www.bioresources.com.au/pdf downloads/ trichs%20 in%20 soybean. Pdf) [17 Juli 2005]
Losey JE, Calvin DD. 1995. Quality assesment of four commercially available species of Trichogramma (Hymenoptera: Trichogrammatidae). J Econ. Entomol 88(5): 1243-1251.
Matadha D, Hamilton GC, Lashomb JH, Zhang J. 2005. Ovipositional preferences and functional response of parasitoids of euonymus scale, Unapis euonymi
Monje JC, Zebitz CPW, Ohnesorge B. 1999. Host and host age preference of
Trichogramma galloi and T. pretiosum (Hymenoptera: Trichogrammatidae) reared on different hosts. Ann of Entomol Soc Amer 92(1): 97-103.
Nordlund DA. 1994. Habitat location by Trichogramma. Di dalam: Wajnberg E, Hassan SA, editor. Biological Control With Egg Parasitoids. UK: CAB International. hlm 155-164.
Orr DB, Salazar CG, Landis DA. 2000. Trichogramma nontarget impacts: a method for biological control risk assessment. Di dalam: Follet, Peter A. & Duan JJ, editor. Nontarget Effects of Biological Control. hlm 111-124.
Pinto JD, Stouthamer R. 1994. Systematics of the Trichogrammatidae with emphasis on Trichogramma. Di dalam: Wajnberg E, Hassan SA, editor. Biological Control With Egg Parasitoids. UK: CAB International. hlm 1-36
Ramlan. 2001. Kajian pelepasan populasi parasitoid Trichogrammatidae untuk pengendalian Helicoverpa armigera (Hubner) dan dampaknya terhadap komunitas arthropoda pada pertanaman kedelai [tesis]. Bogor: Program Pascasarjana. IPB.
Smith SM. 1996. Biological control with Trichogramma; advances, successes, and potential of their use. Annu Rev Entomol 41: 375-406.
Strand MR. 1985. The physiological interactions of parasitoids with their hosts and their influence on reproductive strategies. Di dalam: Waage J and Greathead D, editor. Insect Parasitoid. 13 th Symposium of The Royal Entomological Society of London. Department of Physics Lecture Theatre Imperial College; London, 18-19 Sept 1985. London: Academic Press London. hlm 97-136
Suh CPC, Orr DB, Duyn JWV, Borchert DM. 2000. Trichogramma exiguum
(Hymenoptera: Trichogrammatidae) releases in North Carolina cotton: evaluation of Heliothine pest supression. Ann of Entomol Soc Amer 93(4): 1127-1136.
Tarumingkeng RC. 1994. Dinamika Populasi Kajian Ekologi Kuantitatif. Jakarta: Pustaka Sinar Harapan dan Universitas Kristen Krida Wacana.
Usyati N. 2003. Hubungan antara ciri kebugaran Trichogrammatoidea armigera
Nagaraja (Hymenoptera: Trichogrammatidae) di laboratorium dan keberhasilan parasitisasi di lapangan dengan teknik spot release [tesis]. Bogor: Program Pascasarjana. IPB.
Wang B & D. Ferro. 1998. Functional reponse of Trichogramma ostriniae (Hymenoptera: Trichogrammatidae) to Ostrinia nubilalis (Lepidoptera: Pyralidae) under laboratory and field conditions. Environ Entomol 27(3): 752-758.
Widodo D. 1987. Hama dan Penyakit Padi. Jakarta: CV Pustaka Buana.
Widyarti NAP. 2003. Tanggap fungsional Telenomus remus (Hymenoptera: Scelionidae) pada suhu yang berbeda [skripsi]. Bogor: Jurusan Hama dan Penyakit Tumbuhan. Fakultas Pertanian. Bogor: IPB.
Gambar Lampiran 1 Petak dengan 18 tanaman contoh berjarak 1 m dari titik pelepasan
Gambar Lampiran 2 Penempelan pias pada permukaan bawah daun
Gambar Lampiran 4 Kondisi lahan saat fase generatif akhir
Gambar Lampiran 5 Imago betina T. pretiosum
Gambar Lampiran 7 Parasitoid larva L. indicata famili Braconidae
. Gambar Lampiran 8 Parasitoid telur Ooencyrtus sp famili Encyrtidae
Gambar Lampiran 9 Parasitoid larva L. indicata famili Elasmidae spesies a
Ge n e r a l Li n e a r Mo d e l s Pr o c e d u r e
Du n c a n Gr o u p i n g Me a n N TELUR
A 5 . 2 2 2 9 6 4 0 A
A 3 . 3 3 3 9 1 6 0 A
A 1 . 7 7 8 9 8 0 A
A 1 . 0 0 0 9 2 0 A
A 0 . 1 1 1 9 3 2 0 A