INDUKSI MUTASI KALUS EMBRIOGENIK
MENGGUNAKAN
Ethyl Methane Sulphonate
UNTUK
MERAKIT TANAMAN GANDUM (
Triticum aestivum
L. )
TOLERAN PADA DATARAN RENDAH
LAELA SARI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi berjudul Induksi Mutasi Kalus Embriogenik Menggunakan Ethyl Methane Sulphonate untuk Merakit Tanaman Gandum (Triticum aestivum L.) Toleran pada Dataran Rendah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2015
RINGKASAN
LAELA SARI. Induksi Mutasi Kalus Embriogenik Menggunakan Ethyl Methane Sulphonate untuk Merakit Tanaman Gandum (Triticum aestivum L.) Toleran pada Dataran Rendah. Dibimbing oleh AGUS PURWITO, DIDY SOPANDIE, RAGAPADMI PURNAMANINGSIH, dan ENNY SUDARMONOWATI.
Indonesia merupakan negara pengimpor gandum terbesar ke-3 di dunia. Kebutuhan yang sangat besar menuntut percepatan pengembangan gandum di dalam negeri. Upaya pemenuhan kebutuhan gandum nasional dapat dilakukan dengan penanaman gandum di Indonesia. Sementara gandum di Indonesia daya adaptasinya masih sempit, sehingga perlu perbaikan genotipe untuk memperluas daya adaptasinya. Oleh karena itu penelitian ini perlu dilakukan untuk merakit tanaman gandum yang toleran terhadap kondisi iklim di Indonesia terutama pada dataran rendah.
Tujuan umum penelitian ini adalah untuk mendapatkan genotipe-genotipe mutan gandum yang dapat beradaptasi dan berproduksi pada dataran rendah melalui induksi mutasi. Seleksi awal dilakukan melalui seleksi in vitro menggunakan inkubator dengan 5 macam tingkatan suhu. Tujuan khususnya adalah 1) Mengetahui respons kalus terbaik dari empat genotipe (Alibey, Rabe, Oasis, HP1744) dan dua varietas (Dewata, Selayar) pada media MS + 2.4-D 3 mg/l; 2) Memperoleh keragaman genetik gandum dengan induksi mutasi menggunakan EMS pada kisaran LC20 pada beberapa konsentrasi EMS (0.1; 0.3;
0.5%) dengan waktu perendaman kalus 0; 30; 60; 120; 180 menit; 4) Memperoleh genotipe mutan dengan seleksi in vitro pada suhu 27, 29, 31, 33, dan 35°C; 4) Memperoleh formulasi media yang tepat untuk meregenerasikan dua genotipe kalus hasil induksi mutasi dan seleksi in vitro; 5) Memperoleh genotipe mutan gandum generasi M1 dan generasi M2 berdasarkan karakter agronomi sebagai kriteria seleksi; dan 6) Memperoleh genotipe mutan gandum generasi M3 yang toleran dataran rendah berdasarkan morfo-fisiologis sebagai kriteria seleksi. Analisis yang dilakukan meliputi ANOVA untuk induksi kalus, seleksi in vitro dan regenerasi kalus, analisis probit untuk mengetahui LC20- LC50, analisis korelasi
antara karakter tetua yang diamati dengan karakter genotipe mutan, analisis augmented design, kerapatan stomata, ketebalan daun, viabilitas polen, analisis prolin dan gula total mutan gandum.
Hasil yang diperoleh dalam penelitian ini adalah bahwa respon pertumbuhan kalus terbaik dan diameter tertinggi terdapat pada Dewata dan Selayar. Persentase pertumbuhan kalus menurun sesuai peningkatan konsentrasi EMS dan waktu perendaman. Nilai LC20-LC50 varietas Dewata adalah 0.3% EMS dan waktu
perendaman 30 menit, sedangkan LC20-LC50 varietas Selayar adalah EMS 0.1%
Biji yang dihasilkan dari setiap genotipe generasi M1 dipanen dan jumlah biji dihitung. Selanjutnya semua biji yang diamati ditanam kembali untuk dilanjutkan pada penanaman M2. Karakter yang diamati pada M1 dan M2 terdiri atas: umur berbunga, umur masak, umur panen, tinggi tanaman, jumlah anakan, panjang tangkai malai, panjang malai, jumlai malai, jumlah biji, dan bobot biji. Data dianalisis dengan menghitung rataan dari setiap karakter kuantitatif dibandingkan tanaman mutan dengan tanaman kontrol. Hasil yang diperoleh menunjukkan bahwa perlakuan perendaman EMS pada varietas Dewata, Selayar dan genotipe Alibey mempengaruhi keragaan karakter agronomi pada tanaman gandum generasi pertama (M1) dibandingkan tanaman kontrol. Perendaman EMS 0.1% dengan waktu 60 menit pada genotipe Selayar dan Alibey serta EMS 0.3% dengan waktu perendaman 30 menit pada varietas Dewata dapat meningkatkan jumlah malai, kecuali pada Selayar mempunyai nilai sama yaitu 3. Karakter yang mempunyai korelasi nyata berpotensi untuk digunakan sebagai karakter seleksi. Karakter-karakter seleksi yang diperoleh dari hasil analisis korelasi pada generasi M1 adalah jumlah total biji, jumlah malai dan panjang malai. Karakter-karakter tersebut digunakan sebagai karakter seleksi untuk tanaman gandum generasi M2. Pada generasi M2 dilakukan seleksi berdasarkan hasil rata-rata karakter agronomi yaitu pada jumlah biji per malai dan bobot biji
Analisis augmented design karakter agronomi menunjukkan adanya pengaruh nyata terhadap persentase tumbuh awal (15 mutan), persentase tumbuh panen (13 mutan), panjang tangkai malai (17 mutan), panjang malai (14 mutan), dan tinggi tanaman (16 mutan). Pengaruh tidak nyata terdapat pada karakter waktu berbunga, waktu masak dan waktu panen. Hasil penelitian ini menunjukkan bahwa genotipe mutan yang adaptif biasanya mempunyai kerapatan stomata rendah, kecuali pada genotipe 2Dw-2-17-3 mempunyai kerapatan sama dengan tanaman kontrol. Karakter fisiologi menunjukkan perbedaan nyata pada jumlah prolin yaitu varietas kontrol Dewata, Selayar, Alibey yaitu 79.29 μg/gBB, 201.53 μg/gBB, 4.15 μg/gBB dengan genotipe mutan teringgi Dewata 511.10 μg/gBB. Hal yang sama diperoleh pada hasil analisis gula total/glukosa terdapat perbedaan antara varietas kontrol Dewata, Selayar, Alibey yaitu 14.32 mg/gBB, 5.87 mg/gBB, 13.32 mg/gBB dibanding mutan tertinggi yaitu: Alibey 29.97 dan Dewata 29.06 mg/gBB. Mutan Dewata, Selayar dan Alibey dapat diseleksi berdasarkan karakter jumlah biji per malai dan bobot biji per pengamatan karena kedua karakter tersebut menghasilkan lebih banyak mutan dibandingkan karakter lainnya. Analisis korelasi antar karakter pertumbuhan dan komponen hasil gandum mutan menunjukkan bobot biji per genotipe berkorelasi positif terhadap persentase hasil panen, waktu masak, waktu panen, panjang malai, tinggi tanaman, jumlah anakan, jumlah malai, jumlah biji per malai, bobot 100 bulir, bobot biji per pengamatan, tetapi tidak berkorelasi positif terhadap waktu berbunga dan panjang tangkai malai. Diharapkan, beberapa mutan yang dihasilkan dapat beradaptasi pada dataran rendah tropis, sehingga menambah keragaman plasma nutfah gandum di Indonesia terutama di dataran rendah.
SUMMARY
LAELA SARI. Induced Mutation of Embriogenic Callus using Ethyl Methane Sulphonate for Assembling Wheat (Triticum aestivum L.) in Lowland Tolerance
by AGUS PURWITO, DIDY SOPANDIE, RAGAPADMI
PURNAMANINGSIH, dan ENNY SUDARMONOWATI.
Indonesia is the third wheat importer countries in theworld. High national demand requires the development of wheat in the country. Addressing the needs of the national wheat can be conducted by planting wheat in Indonesia. While the adaptability of wheat in Indonesia is still narrow, so it is necessary to expand the repair genotype adaptability. Therefore, this research needs to be
conducted to assemble the wheat crop is tolerant to climatic conditions in Indonesia, especially in the lowlands
. The general aims of this study were to obtain wheat mutant that can adapt in the lowlands through induced mutation. The initial selection was conducted through in vitro selection in incubator with five different temperature levels. The research specific goals were 1) To obtain best callus formation response of six genotypes (Dewata, Selayar, Alibey, Rabe, Oasis and HP1744) using of MS + 2,4-D 3 mg/l; 2) To obtain the wheat genetic diversity by induced mutation using EMS (0.1; 0.3; 0.5%) with a soaking time for 0; 30; 60; 120; 180 minutes; 3). To obtain in vitro selection at temperature 27, 29, 31, 33 and 35 °C; 4) To obtain the type and concentration of plant growth regulator for regeneration mutant and selection in vitro; 5) To obtain generation wheat mutant genotype M1 and M2 generations with selection criteria (agronomic characters); 6) To obtain the mutant wheat genotypes tolerant generation M3 lowland with the selection criteria (agronomic characters, anatomy, physiology). Analysis ANOVA for callus induction, in vitro selection and its regeneration, probit analysis to determine LC20-LC50, correlation analysis between the parent character with mutant,
augmented analysis design, stomata density, thickness of the leaves, pollen viability, analysis of proline, and total sugar of wheat mutant as well.
The results obtained in this study were the best response and diameter of callus growth were highest in the genotype of the Dewata and Selayar. The percentage of callus growth would be lower with increasing concentration of EMS and immersion time. LC20-LC50 genotype Dewata was 0.3% EMS with 30
minutes soaking, while LC20-LC50 genotype Selayar was 0.1% EMS with 60
minutes. Genotypes Dewata and Selayar have tolerant adaptation at a temperature of 27 °C with a value of 84% and 72% respectively. The level tolerant of plants and callus growth decreases with increasing temperature. At the higest temperature of 35 °C no callus growth or plant death. The highest shoot number of genotypes Selayar and Dewata was in RG2 media (MS + BA 0,1 ml/l + kinetin 2 mg/l + tyrosine 0.05 gr/l + sucrose 3% + sorbitol 6%) which were 36% and 44% respectively.
quantitative character compared to control plants. Results obtained showed that the EMS soaking treatment in genotype Dewata, Selayar and Alibey affected the performance of agronomic characters in wheat M1 compared to the control. Soaking of EMS 0.1% for 60 minutes on genotype Selayar and Alibey and EMS 0.3% for 30 minutes in the Dewata genotype may increase the the number of panicles, except in Selayar with have the same value, namely 3. Characters that have significant correlation can potentially be used character selection. Characters selection obtained from the correlation analysis of M1 were total number of seeds, number of panicles and panicle length. These characters will be used as the character selection for the wheat M2 generation. In the M2 the selection was based on the average yield of wheat agronomic characters which were the number of grains per panicle and grain weight.
Augmented design analysis of agronomic characters showed significant effect on the percentage of early growth (15 mutants), percentage of plant harvested (13 mutants), stem length panicle (17 mutants), panicle length (14 mutants), and plant height (16 mutants). There was no significant effect on the character of days to flowering, days to mature and harvest. These results also showed that the adaptive mutant usually have a low stomata density, except in genotype 2Dw-2-17-3 which had the same density with control. While, the physiology character showed significant differences in the concentration of proline on wild type genotypes of Dewata, Selayar, Alibey which were 79.29 μg/gBW, 201.53 µg/gBW, 4.15 µg/gBW respectively with the highest Dewata mutant genotype was 511.10 µg/gBW. The same result has obtained for the analysis of total sugar / glucose were no difference between the control varieties Dewata, Selayar, Alibey were 14.32 mg/gBW, 5.87 mg/gBW, 13.32 mg/gBW respevtively comapred to the highest mutant Alibey 29.97 and 29.06 Dewata mg/gBW. The mutants of the Dewata, Selayar and Alibey can be selected based on the character of the number of seeds per panicle and grain weight per observation because both characters are producing more mutants than the other characters. Correlation analysis between the agronomical characters of growth and grain yield components of mutants, showed that seed weight per genotype had a positive correlation with the percentage of yield, days to mature, days to harvest, panicle length, plant height, number of tillers, number of panicles, number of grains per panicle, weight of 100 grains, grain weight per observation, but not positively correlated to the days to flowering and stem length panicle. Hopefully some mutants produced could adapt to the tropical lowlands, thus adding to the diversity of wheat germplasm in Indonesia.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
INDUKSI MUTASI KALUS EMBRIOGENIK
MENGGUNAKAN
Ethyl Methane Sulphonate
UNTUK
MERAKIT TANAMAN GANDUM (
Triticum aestivum
L.)
TOLERAN PADA DATARAN RENDAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup : 1. Dr. Amin Nur, S.P, M.Si.
2. Dr. Ir. Hajrial Aswidinnoor, M.Sc. Penguji Luar Komisi pada Sidang Promosi: 1. Dr. Ir. Hajrial Aswidinnoor, M.Sc.
PRAKATA
Puji syukur selalu penulis panjatkan kepada Allah SWT atas segala rizki-Nya sehingga penulisan disertasi dengan judul “Induksi mutasi kalus embriogenik menggunakan Ethyl Methane Sulphonate untuk merakit tanaman gandum (Triticum aestivum L) toleran pada dataran rendah” dapat diselesaikan. Topik penelitian yang dipilih untuk memenuhi sebagian kebutuhan gandum yang sangat besar dan kemampuan impor yang semakin terbatas. Semoga upaya ini dapat mengurangi ketergantungan impor gandum di Indonesia.
Penulisan disertasi ini tidak mungkin diselesaikan sendiri oleh penulis tanpa bantuan dari berbagai pihak. Oleh karena itu penulis menyampaikan rasa terima kasih yang tulus dan penghargaan setinggi-tingginya kepada:
1. Dr. Ir. Agus Purwito, M.Sc.Agr. selaku ketua komisi pembimbing yang banyak memberikan arahan penulisan disertasi ini.
2. Dr. Ir. Ragapadmi Purnamaningsih, M.Si., Prof. Dr. Ir. Didy Sopandie, M.Agr dan Prof. (R). Dr. Ir. Enny Sudarmonowati sebagai anggota komisi pembimbing yang banyak membantu dalam penyusunan konsep sampai akhir penelitian, memberikan masukan dan bimbingan dalam penyusunan sampai selesainya penulisan disertasi.
3. Dr. Dewi Sukma, SP. M.Si. dan Dr. Ir. Syarifah Iis Asyiah, M.Sc sebagai penguji luar komisi pada ujian prelim lisan, Dr. Amin Nur, S.P. M.Si dan Dr. Ir. Hajrial Aswidinnoor, M.Sc sebagai penguji luar komisi pada ujian tertutup, Dr. Tri Muji Ermayanti dan Dr. Ir. Hajrial Aswidinnoor, M.Sc sebagai penguji luar komisi pada sidang promosi program doktor.
4. Kepala Pusat Penelitian Bioteknologi LIPI atas izin untuk melanjutkan sekolah di IPB.
5. KemenRistek (sekarang KemenRistekdikti) atas beasiswa pendidikan program doktor yang diberikan dari tahun 2010 sampai 2014.
6. Dr. Karden Mulya (Kepala BB-BIOGEN) dan Dr. Bambang Purwantara (Direktur SEAMEO BIOTROP) atas izin dan fasilitas selama penelitian, Prof. (R). Dr. Ika Mariska, Prof. (R). Dr. Endang Gati dan Dr. Ika Rostika atas diskusinya selama di Laboratorium in vitro Kelti Biologi Sel dan Jaringan.
7. Dr. Ir. Trikoesoemaningtyas, M.Sc dan Dr. Ir. Yudiwanti Wahyu EK, MS. selaku Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman beserta staf atas semua pelayanan yang telah diberikan.
8. Pimpinan Sekolah Pascasarjana IPB beserta seluruh jajaran, atas semua pelayanan yang diberikan selama mengikuti pendidikan di jenjang S3 SPS IPB. 9. Maria Imelda M.Sc selaku kepala laboratorium BSJT periode 2000-2014. 10. Teman-teman di Laboratorium BSJT P2 Bioteknologi LIPI.
pendidikan di Sekolah Pascasarjana IPB tetap terjalin dengan baik sampai masa datang.
12. Djoko Tamami, Dadang Komarudin, Joko Mulyono, Irawan dan Imam Mawardi atas bantuan pengamatan dan dalam persiapan penelitian.
13. Ayahanda Soedjono (Alm.) dan Ibunda Hj. Ai Sutarsih, terima kasih atas semua pengorbanan dan doa yang tulus yang telah diberikan. Ayahanda mertua H. Slamet dan Hj. Miyem, terima kasih atas perhatian dan dorongan kepada penulis.
14. Saudara tercinta Soegianto, Herlina, Yulia Sekarsari, Tyas, Tomo, Eni, Agus (Alm.), Devi, dan seluruh keponakan, terima kasih atas kesabaran dan motivasi. 15. Terima kasih yang sebesar-besarnya untuk suami Budi Murdiyantoro S.E, kedua
anakku tercinta Aliefa Aviryalashra Az-Zahra dan Muhammad Savero Avtariansyah atas kesabaran, motivasi, diskusi dan kerelaan waktu yang tersita untuk studi dan penelitian.
Penulis berharap disertasi ini dapat menjadi sumber pengetahuan dan munculnya ide-ide baru dalam pengembangan ilmu serta penelitian selanjutnya, terutama untuk komoditas gandum dalam berbagai aspek kajian dan penelitian. Akhirnya, penulis menaruh harapan semoga disertasi ini dapat memberi manfaat bagi semua pihak.
Bogor, Juni 2015
DAFTAR ISI
DAFTAR TABEL xiii
DAFTAR GAMBAR xiv
DAFTAR LAMPIRAN xv
PENDAHULUAN 1
Latar Belakang 1
Kerangka Pemikiran 3
Tujuan Penelitian 4
Tujuan Khusus 4
Diagram Alir Tahapan Penelitian 5
TINJAUAN PUSTAKA 6 Klasifikasi dan Asal Tanaman Gandum 6
Morfologi dan Budidaya Tanaman Gandum 7
Karakter Morfo-Fisiologis 10
Adaptasi Tanaman terhadap Cekaman Suhu Tinggi 11
Pemulian Mutasi Tanaman Gandum melalui Mutagenesis menggunakan EMS 13
Seleksi In Vitro 14
Seleksi In Vivo 15
INDUKSI MUTASI DAN SELEKSI In vitro MUTAN PUTATIF GANDUM (Triticum aestivum L.) 17
Abstrak 17
Abstract 17
Pendahuluan 18
Metode Penelitian 19 Hasil dan Pembahasan 21 Simpulan 39 PERBANYAKAN BENIH DAN KARAKTERISASI AGRONOMI M1, M2 GANDUM (Triticum aestivum L.) YANG DITANAM DI DATARAN TINGGI (>1000 m dpl) 40 Abstrak 40
Abstract 41
Pendahuluan 41 Metode Penelitian 43
Hasil dan Pembahasan 45
Simpulan 55
KARAKTERISASI AGRONOMI, ANATOMI DAN FISIOLOGI MUTAN GANDUM (Triticum aestivum L.) GENERASI M3 YANG DITANAM DI DATARAN RENDAH (±250 m dpl) 56
Abstract 57
Pendahuluan 57
Metode Penelitian 59
Hasil dan Pembahasan 63
Simpulan 77
PEMBAHASAN UMUM 78
SIMPULAN UMUM 83
SARAN 83
DAFTAR PUSTAKA 84
LAMPIRAN 95
DAFTAR TABEL
Hal 1 Hasil skor dan warna kalus gandum pada media
MS+2.4D 3 mg/l.
26 2 Analisis probit LC20-LC50 pada varietas Selayar dan
Dewata
29 3
4
Jumlah akar varietas Dewata dan Selayar pada berbagai macam media
Persentase hidup tanaman setelah aklimatisasi
38 45 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Karakter morfologi pada tanaman gandum M1.
Analisis korelasi antar karakter agronomi tanaman gandum generasi M1
Hasil Rata-rata karakter agronomi pada mutan gandum generasi M2 yang di tanam di kebun percobaan Pacet (>1000 m dpl).
Rata-rata karakter agronomi pada mutan gandum generasi M2 yang di tanam di kebun percobaan Pacet (>1000 m dpl).
Persentase tanaman yang tumbuh pada generasi M2 Nilai pendugaan komponen ragam dan heritabilitas arti luas populasi M1 di Pacet (>1000 m dpl).
Analisis korelasi antar karakter agronomi pertumbuhan dan komponen hasil tanaman gandum Dewata, Selayar dan Alibey generasi M2 di Kebun Percobaan Pacet, Tahun 2012.
Model analisis ragam Augmented Design
Karakter agronomi pertumbuhan pada gandum mutan Dewata, Selayar dan Alibey yang di tanam di kebun
percobaan SEAMEO-BIOTROP, Bogor, Tahun 2013 (± 250 m dpl).
Karakter agronomi komponen hasil pada gandum mutan Dewata, Selayar dan Alibey yang di tanam di SEAMEO-BIOTROP, Bogor (± 250 m dpl).
Analisis korelasi antar karakter agronomi pertumbuhan dan komponen hasil tanaman gandum Dewata, Selayar dan Alibey generasi M3 di Kebun Percobaan SEAMEO-BIOTROP, Bogor, Tahun 2013.
Keragaan tebal daun, kerapatan stomata dan kehijauan daun gandum kontrol dan mutan.
Jumlah kalus dan tunas yang terbentuk dari induksi kalus sampai regenerasi
Penurunan karakter jumlah biji per malai dan bobot biji per malai yang ditanam di dataran tinggi dan dataran rendah
Persentase tanaman yang tumbuh di M1, M2, M3 dan M4. Jumlah total tanaman yang ditanam di lapangan Jumlah total tanaman yang ditanam di lapangan.
DAFTAR GAMBAR
Hal
1. Diagram alir tahapan penelitian 5
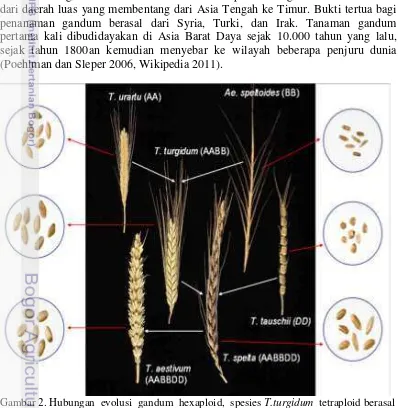
2 Hubungan evolusi gandum hexaploid, spesies T.turgidum tetraploid berasal dari T.uratu dan Ae.speloides (spesies liar), sedangkan T.aestivum berasal dari T.speta yang sudah dibudidaya.
6
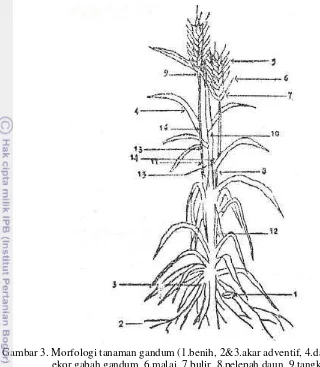
3 Morfologi Tanaman Gandum (1. benih, 2&3. akar adventif, 4. daun, 5. ekor gabah gandum, 6. malai, 7. bulir, 8. pelepah daun, 9. tangkai malai, 10 &11. batang utama, 12. anakan, 13. ruas, 14. daun telinga, 15. lidah daun,16. buku).
8
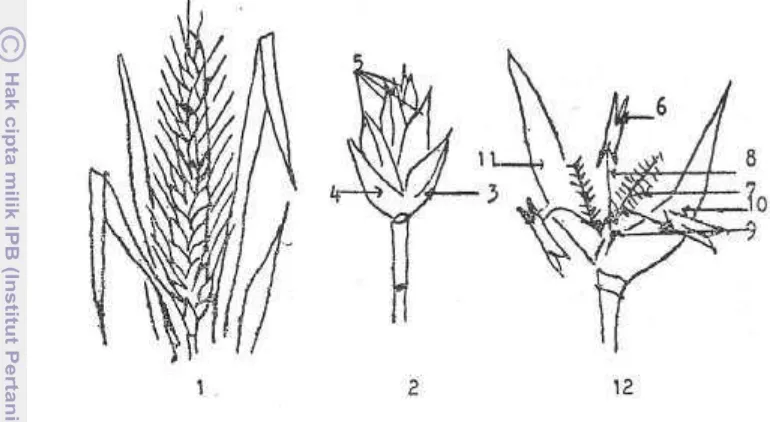
4 Malai, Bulir dan Bunga (1. malai, 2. bulir, 3. gabah atas, 4. gabah bawah, 5. bunga (2-5 buah), 6. kotak sari, 7. kepala putik, 8. tangkai sari, 9. putik, 10. palea, 11. lemma, 12. bunga gandum).
9
5 Struktur biji gandum 10
6 Keragaan biji gandum: A. Biji yang normal, B. Biji akibat cekaman suhu tinggi
12 7 Bahan tanaman enam genotipe yang digunakan untuk
induksi kalus
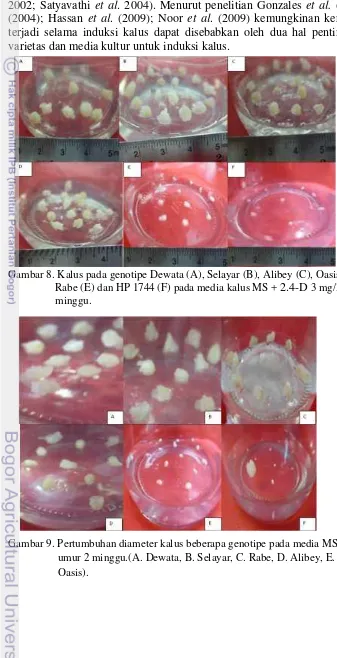
22 8 Induksi kalus pada genotipe Dewata(A), Selayar(B),
Alibey(C), Oasis(D), Rabe(E), HP1944(F) pada media MS+ 2.4-D 3 mg/l.
23
9 Pertimbuhan diameter kalus beberapa genotipe pada media MS+ 2.4-D 3 mg/l (A. Dewata, B. Selayar, C. Rabe, D. Alibey, E. HP1744, F. Oasis).
23
10 Persentase pertumbuhan kalus dari beberapa genotipe gandum di media MS+ 2.4-D 3 mg/l umur 2 minggu.
24 11 Diameter kalus dari beberapa genotipe gandum pada
media MS+ 2.4-D 3 mg/l umur 2 minggu.
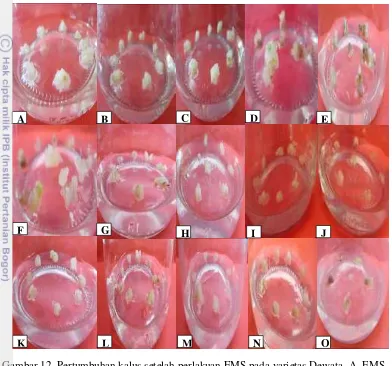
25 12 Pertumbuhan kalus setelah perlakuan EMS pada
varietas Dewata, A. EMS 0.1%; waktu 0’, B. EMS
0.1%; waktu 30’, C. EMS 0.1%; waktu 60’, D. EMS 0.1%; waktu 120’, E. EMS 0.1%; waktu 180’, F.
EMS 0.3%; waktu 0’, G.EMS 0.3%; waktu 30’, H.
EMS 0.3%; waktu 60’, I. EMS 0.3%; waktu 120’, J. EMS 0.3%; waktu 180’, K. EMS 0.5%; waktu 0’,
L. EMS 0.5%; waktu 30’, M. EMS 0.5%; waktu 60’,
N. EMS 0.5%; waktu 120’, O. EMS 0.5%; waktu 180’.
27
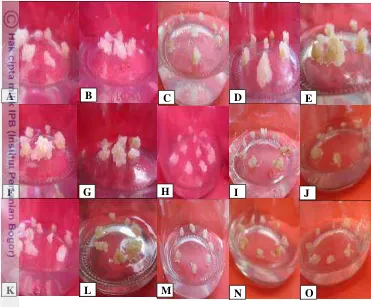
13 Pertumbuhan kalus setelah perlakuan EMS pada varietas Selayar, A. EMS 0.1%; waktu 0’, B. EMS 0.1%; waktu 30’, C. EMS 0.1%; waktu 60’, D. EMS
0.1%; waktu 120’, E. EMS 0.1%; waktu 180’, F. EMS 0.3%; waktu 0’, G.EMS 0.3%; waktu 30’, H.
EMS 0.3%; waktu 60’, I. EMS 0.3%; waktu 120’, J. EMS 0.3%; waktu 180’, K. EMS 0.5%; waktu 0’,
L. EMS 0.5%; waktu 30’, M. EMS 0.5%; waktu 60’,
N. EMS 0.5%; waktu 120’, O. EMS 0.5%; waktu 180’.
14 Persentase pertumbuhan kalus gandum Dewata (A) dan Selayar (B) pada konsentrasi EMS 0.1; 0.3 dan 0.5% dengan waktu perendaman 0; 30; 60; 120 dan 180 menit
29
15
16
17
(A) Persentase kalus Dewata yang hidup pada suhu 27°C - 35°C.
(B) Persentase kalus Selayar yang hidup pada suhu 27°C - 35°C
Pertumbuhan kalus setelah seleksi in vitro. A. Dewata (Dw) 27 °C, B. Dw 29 °C, C. Dw 31 °C, D.
Dw 33 °C, E. Dw 35°C. F. Selayar (Sl) 27°C, G. Sl 29 °C, H. Sl 31 °C, I. Sl 33 °C, J. Sl 35 °C.
Kalus embriogenik mutan varietas A. Dewata, B. Selayar, yang akan diregenerasikan pada berbagai media regenerasi.
31
33
33
18 Persentase tumbuh tunas dan tumbuh akar pada kalus Selayar di berbagai macam media umur 2 bulan.
34 19
20
Persentase tumbuh tunas dan tumbuh akar pada kalus Dewata di berbagai macam media umur 2 bulan. Rhizogenesis pada media regenerasi RG3
34 37 21 Pertumbuhan regenerasi kalus Selayar menjadi
planlet pada berbagai macam media. A) RG0. B) RG1. C) RG2. D) RG3. E) RG4. F) RG5.G) RG6
37
22 Pertumbuhan regenerasi kalus Dewata menjadi planlet pada berbagai macam media. A) RG0. B) RG1. C) RG2. D) RG3. E) RG4. F) RG5.G) RG6.
38
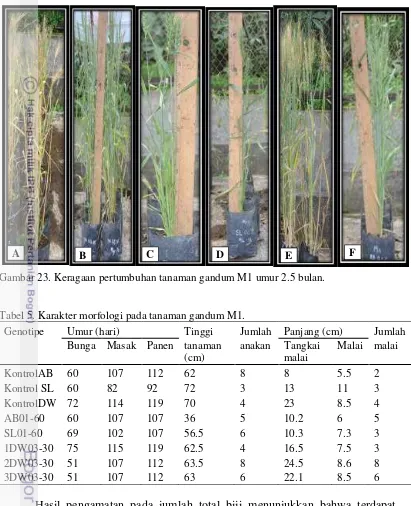
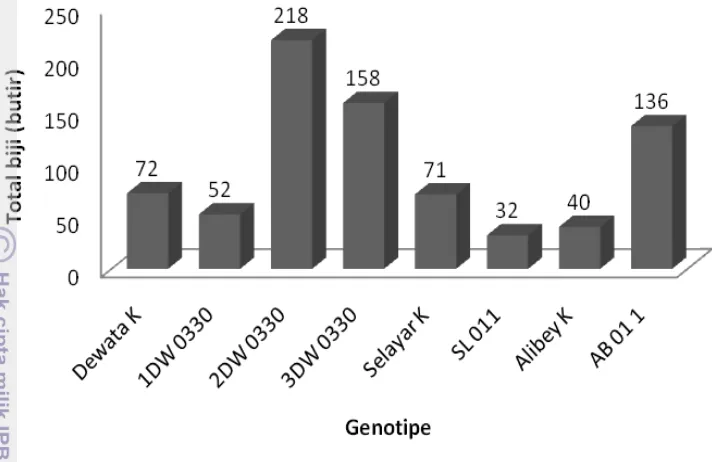
23 Keragaan pertumbuhan tanaman M1 umur 2.5 bulan. 46 24 Total biji yang dihasilkan pada tanaman gandum M1. 47 25 Keragaan biji gandum generasi M1 di Pacet (>1000
m dpl.). A.Dewata kontrol, B. Selayar kontrol, C. Alibey kontrol,D. Dewata mutan, E. Selayar mutan, F. Alibey mutan.
48
26 Keragaan biji gandum generasi M2 di Pacet ( > 1000 m dpl.) A.Dewata kontrol, B. Selayar kontrol, C. Alibey kontrol, D. Dewata mutan, E. Selayar mutan, F. Alibey mutan.
51
27 Keragaan Tanaman M2. A). Tanaman umur 2 bulan B). Panen di Pacet (> 1000 m dpl.).
53 28 Denah penanaman gandum generasi M3 di
SEAMEO-BIOTROP
29 Keragaan panjang malai dan kehijauan daun bendera tanaman M3 di BIOTROP (± 250 m dpl).
68 30 Keragaan biji gandum generasi M3 Dewata, Selayar,
dan Alibey di Biotrop (± 250 m dpl). A. Dewata
kontrol, B. 2Dw-6-17-1, C. Selayar kontrol, D. Sl-3-2-2, E. Alibey kontrol, F. Ab-2-14-1.
69
31 Fenomena yang terjadi di lapangan, A). Biji gandum tidak berkecambah B) Biji gandum berkecambah (toleran). C) Daun menggulung karena cekaman suhu tinggi (toleran). D) Daun tidak menggulung.
70
32 Keragaan stomata gandum generasi M3 Dewata, Selayar, dan Alibey di Biotrop (± 250 m dpl). A. Dewata kontrol, B. Dw-1-15-1, C. Selayar kontrol, D. Sl-3-2-2, E. Alibey kontrol, F. Ab-2-14-1.
71
33 Keragaan ketebalan daun pada gandum varietas Dewata, Selayar, Alibey dan mutan putatif. A. Dewata kontrol,B. Dw-1-15-1, C. Selayar kontrol,D. Sl-3-2-2, E. Alibey kontrol, F. Ab-2-14-1.
73
34 Persentase viabilitas polen pada tanaman gandum kontrol dan mutan M4
74 35 Keragaan jumlah polen antara tanaman mutan (atas)
dan kontrol (bawah) dengan Bar 20µm. A. Dw-1-15-1, B. Dewata kontrol, C. Sl-3-2-2 D. Selayar kontrol, E. Ab-2-14-1. F. Alibey kontrol
75
36 Keragaan bunga gandum yang terdiri atas 3 benang sari.
75 37 Hasil analisis prolin pada gandum varietas kontrol
dan genotipe mutanDewata, Selayar, dan Alibey
76 38
39
Hasil analisis glukosa pada gandum varietas kontrol dan genotipe mutan
Dewata, Selayar, dan Alibey.Persentase jumlah kalus dari induksi kalus sampai regenerasi.
DAFTAR LAMPIRAN
Hal 1. Hasil analisis contoh tanah di Pacet (>1000 m dpl). 95 2 Hasil analisis contoh tanah di Seameo-Biotrop Bogor
(±250 m dpl).
95 3
4 5 6
Data iklim bulanan tahun 2011-2012 di Pacet (>1000 m dpl).
Data iklim bulanan tahun 2013 di Seameo-Biotrop Bogor (±250 m dpl).
Deskripsi Gandum Dewata, Selayar dan Alibey yang ditanam di dataran tinggi >1000 m dpl.
Analisis Augmented design karakter pertumbuhan generasi M3
95 96 96 97 7 Analisis Augmented design karakter komposisi hasil
generasi M3
PENDAHULUAN
Latar Belakang
Perubahan iklim global berdampak secara langsung terhadap sektor pertanian yang menyebabkan beberapa kerugian antara lain gagal panen, karena keberhasilan produksi pertanian sangat tergantung pada kondisi iklim (Warren et al. 2006). Salah satu produksi pertanian yang tergantung pada iklim adalah tanaman gandum.
Gandum (Triticum aestivum L.) merupakan tanaman serealia subtropis. Program pengembangan varietas unggul gandum di Indonesia telah dilakukan sejak tahun 1985,tetapi sangat terbatas pada wilayah dataran tinggi (>1000 m dpl) yang mempunyai suhu optimum sekitar 15–25 o
C. Wilayah dataran tinggi (>1000 m dpl) mempunyai kendala yaitu persaingan dengan komoditas lain yang nilai ekonominya cukup tinggi seperti sayuran dan hortikultura lainnya. Laju peningkatan hasil gandum saat ini masih terlalu rendah untuk dapat memenuhi kebutuhan gandum di masa depan (Reynolds 2002). Kebutuhan yang sangat besar dan kemampuan impor yang semakin terbatas menuntut pengembangan gandum di dalam negeri. Usaha untuk memenuhi kebutuhan gandum dalam negeri dapat dilakukan dengan pengusahaan budidaya gandum di Indonesia. Upaya ini dapat mengurangi dan menekan ketergantungan impor gandum.
Kebutuhan gandum di Indonesia hampir 100% dipenuhi dari impor, sehingga Indonesia merupakan negara pengimpor gandum terbesar ke-3 dunia dengan total impor 7.43 juta ton pada tahun 2014 dan akan terus meningkat dengan laju 2% per tahun (Reynolds 2002; Aptindo 2014). Konsumsi terigu pada tahun 2013 mencapai 5.35 juta naik 4.2% dari tahun 2012 (Aptindo 2014). Menurut Wittenberg (2004) gandum memiliki peranan sebagai pendukung ketahanan pangan dunia karena secara global tanaman ini merupakan komoditas serealia yang banyak diusahakan di dunia dan dikonsumsi oleh sekitar 36% dari total penduduk dunia.
belum ada rekomendasi galur-galur gandum yang adaptif di dataran rendah di Indonesia.
Salah satu kendala yang dihadapi di Indonesia yaitu kurangnya varietas yang adaptif dan faktor lingkungan seperti cekaman suhu tinggi. Di antara berbagai faktor lingkungan, cekaman yang diakibatkan oleh suhu merupakan yang paling banyak dihadapi oleh tanaman (Iba 2002). Setiap jenis tanaman mempunyai suhu optimum masing-masing untuk tumbuh dan pola penyebarannya ditentukan oleh wilayah suhu tempat tumbuhnya. Suhu berubah lebih cepat dibandingkan penyebab cekaman lainnya. Suhu mempengaruhi komponen sel dan metabolisme sel dalam ruang yang luas (Sung 2003). Cekaman suhu tinggi merupakan kendala umum selama tahapan pengisian biji dan pembungaan pada banyak tanaman sereal pada daerah tropis. Cekaman suhu tinggi memperpanjang waktu pengisian biji, menurunkan berat kernel gandum hingga 7% pada musim semi. Pengurangan serupa terjadi pada pati, protein dan isi minyak kernel jagung dan kualitas biji-bijian sereal lainnya. Jumlah dan berat biji gandum juga peka terhadap cekaman suhu tinggi, seperti jumlah butir dan berat biji per malai menurun dengan meningkatnya suhu (Wahid et al. 2007). Cekaman suhu tinggi menjadi faktor pembatas dalam usaha pengembangan gandum di dataran rendah, karena pada dasarnya gandum merupakan tanaman subtropis yang menghendaki suhu yang rendah untuk proses pertumbuhan dan perkembangannya. Menurut Peet dan Willits (1998) cekaman suhu tinggi diartikan sebagai kenaikan suhu yang menyebabkan kerusakan irreversibel pada pertumbuhan dan perkembangan tanaman. Batasan kerusakan suhu tinggi untuk tiap tanaman bersifat relatif tergantung wilayah atau habitat asal tanaman.
Sifat toleransi gandum terhadap cekaman suhu tinggi dapat diperbaiki dan ditingkatkan melalui perakitan genetik dalam program pemuliaan tanaman. Keberhasilan perakitan varietas baru dengan sifat – sifat yang diinginkan sangat ditentukan oleh beberapa faktor yaitu ketersediaan sumber genetik, metode rekombinasi genetik (persilangan), kultur jaringan, rekayasa genetik, induksi mutasi dan metode seleksi yang tepat. Witjaksono (2003) berpendapat Induksi mutasi dan bioteknologi dapat meningkatan keragaman genetik dan perbaikan varietas untuk satu atau dua sifat pada tanaman tersebut.
Strategi adaptasi yang paling mendesak dilakukan untuk menanggulangi pemanasan suhu tinggi terhadap pertanian di Indonesia menurut World Development Report (2008) antara lain: menanam varietas yang memiliki daya adaptasi tinggi, mengubah masa tanam menyesuaikan iklim dan mempraktekkan pertanian dengan masa tanam yang lebih singkat. Induksi mutasi merupakan salah satu metode yang dapat digunakan untuk memperbaiki genetik untuk mengantisipasi menghadapi pemanasan global untuk merakit varietas gandum yang toleran pada dataran rendah.
kromosom sehingga sangat menguntungkan bagi kegiatan pemuliaan. EMS termasuk senyawa alkil yang banyak digunakan sebagai mutagen yang efisien pada tanaman tingkat tinggi. EMS sering digunakan karena mudah didapat, harganya murah dan tidak meninggalkan racun setelah hidrolisat (Van Harten 1998).
Induksi mutasi yang dilanjutkan dengan seleksi ketahanan terhadap suhu tinggi secara in vitro telah dilakukan pada tanaman kentang dan bawang putih dan telah berhasil memperoleh mutan toleran suhu tinggi (Das et al. 2000; Gosal et al. 2001; Zhen 2001). Induksi mutasi yang digabungkan dengan kultur in vitro merupakan metode yang menguntungkan menurut Svetleva dan Crino (2005) karena dapat meningkatkan frekuensi terbentuknya variasi baru. Secara teknis kultur in vitro dapat menghasilkan variasi somaklonal dan variasi ini dapat ditingkatkan dengan mutagen. Melalui teknik kultur in vitro ini akan didapatkan keuntungan bagi pemuliaan tanaman yaitu merangsang terbentuknya keragaman genetik.
Toleran suhu tinggi telah diteliti melalui induksi mutasi secara in vitro menggunakan EMS antara lain pada tanaman purwoceng (Ajijah 2009), dan ubi jalar (Luan et al. 2007). Perlakuan EMS dengan konsentrasi 0.25-0.5 % pada tanaman barley, menimbulkan mutasi dengan laju 5x lebih tinggi dibandingkan radiasi sinar x, terutama untuk mutasi klorofil (Van Harten 1998). Gandum yang diperlakukan dengan EMS lebih cepat berbunga dan matang bijinya (Viswanathan dan Reddy 1996). EMS juga telah digunakan secara in vitro untuk menginduksi keragaman pada tanaman alpukat (Yenisbar 2005) dan perbaikan sifat agronomi pada tanaman anggur (Khawale et al. 2007).
Kerangka Pemikiran
Dewasa ini pola pangan di Indonesia mulai bergeser tiap tahunnya dari pola konsumsi beras ke konsumsi gandum seperti roti, mie, sereal, dan terigu yang mencapai 17.7 kg/kapita/tahun (Aptindo 2012). Dengan harga gandum saat ini US$ 593/ton dibutuhkan devisa hampir US$ 2.4 miliar atau Rp 22.1 triliun. Tahun 2020 impor gandum diprediksi akan mencapai 8.5 juta ton sehingga memerlukan devisa yang sangat tinggi dan ketergantungan terhadap negara Amerika Serikat sebagai negara pengekspor. Kebutuhan gandum nasional hampir 100% diperoleh dari impor, sehingga Indonesia merupakan negara pengimpor gandum terbesar ke 3 dunia dengan total impor 4.8 juta ton/tahun dan akan terus meningkat dengan laju 2% pertahun (Aptindo 2012; Reynolds 2002). Gandum memiliki peranan sebagai pendukung ketahanan pangan dunia karena secara global tanaman ini merupakan komoditas serealia yang paling banyak diusahakan di dunia dan dikonsumsi sekitar 36% dari total penduduk dunia (Wittenberg 2004).
pada dataran tinggi. Dataran tinggi mempunyai kendala yaitu persaingan komoditas lain yang nilai ekonominya cukup tinggi seperti sayur-sayuran dan hortikultura lainnya. Para petani lebih memilih untuk menanam sayuran dibandingkan menanam gandum. Oleh karena itu wilayah alternatif lain untuk menanam gandum adalah di dataran rendah. Tanaman gandum di Indonesia belum ada yang beradaptasi pada dataran rendah, sehingga sampai saat ini belum ada informasi tentang perakitan gandum yang adaptif di dataran rendah. Strategi adaptasi yang paling efisien dilakukan untuk menanggulangi pemanasan suhu tinggi terhadap pertanian di Indonesia adalah dengan menanam varietas yang memiliki daya adaptasi luas, mengubah masa tanam menyesuaikan iklim dan menanam dengan masa tanam yang lebih singkat serta menggunakan mutagen untuk keragaman genetik. Penelitian ini memilih menggunakan mutagen kimia yaitu EMS untuk meningkatkan keragaman genetik yang diperlukan untuk menghasilkan galur-galur tanaman gandum untuk dataran rendah dan mempunyai hasil produksi yang tinggi. EMS diharapkan dapat menginduksi sifat ketahanan terhadap suhu tinggi pada tanaman gandum terutama di dataran rendah. Oleh karena itu, penelitian ini perlu dilakukan untuk merakit tanaman gandum yang toleran terhadap kondisi iklim di Indonesia terutama di dataran rendah.
Tujuan Penelitian
Tujuan penelitian ini adalah untuk memperoleh genotipe-genotipe tanaman gandum yang toleran suhu tinggi pada dataran rendah.
Tujuan Khusus
1) Mempelajari respon pembentukan kalus 4 genotipe dan 2 varietas dengan menggunakan media MS + 3 mg/l 2.4-D.
2) Memperoleh keragaman genetik gandum dengan induksi mutasi menggunakan EMS pada kisaran Lethal concentration 20 (LC20) – LC50
pada beberapa konsentrasi EMS dan waktu perendaman kalus.
3) Memperoleh genotipe mutan dengan seleksi in vitro pada suhu 27, 29, 31, 33 dan 35 °C.
4) Memperoleh formulasi media yang tepat untuk meregenerasikan dua genotipe kalus hasil induksi mutasi dan seleksi in vitro.
5) Memperoleh genotipe mutan gandum generasi M1 dan generasi M2 berdasarkan karakter agronomi sebagai kriteria seleksi.
6) Memperoleh genotipe mutan gandum generasi M3 yang toleran dataran rendah berdasarkan morfo-fisiologis sebagai kriteria seleksi.
Gambar 1. Diagram alir tahapan penelitian Induksi pembentukan kalus embriogenik
( 6 genotipe )
Perlakuan EMS (0.1; 0.3; 0.5 %) dan waktu perendaman (0, 30, 60, 120,180 menit)
Seleksi kalus mutan putatif hasil LC50 pada
2 genotipe dalam suhu yang berbeda (27; 29; 31; 33; 35 °C).
Pengujian genotipe generasi M3 dengan karakter agronomi, anatomi, dan fisiologi di dataran rendah ± 250 m dpl. (Bogor) dengan kisaran suhu 27-35°C
Perbanyakan dan pengujian generasi M2 dengan karakter agronomi di dataran tinggi >1000 m dpl. (Pacet) dengan kisaran suhu 20-27°C.
Regenerasi kalus mutan putatif hasil seleksi in vitro dalam berbagai media RG.
Dipilih 2 genotipe yang terbaik
Dipilih LC50 pada 2 genotipe
yang dipilih.
Dihasilkan kalus mutan putatif pada setiap suhu.
Planlet yang optimal pada media RG
Aklimatisasi M1 (Pacet)
Tanaman M2 (Biji M3)
Tanaman M3 (Biji M4) Tanaman M1 (Biji M2)
TINJAUAN PUSTAKA
Klasifikasi dan Asal Tanaman Gandum
[image:30.595.85.483.302.710.2]Gandum (Triticum aestivum L.) merupakan spesies yang berasal dari divisi Spermatophyta, kelas Angiospermae, subkelas Monocotyledone, famili Gramineae (Poaceae), genus Triticum. Genus Triticum dikelompokkan ke dalam tiga kelas ploidi berdasarkan jumlah kromosom yaitu diploid (2n=2x=14) terdapat 7 spesies, tetraploid (2n=4x= 28) terdapat 12 spesies dan heksaploid (2n=6x=42) terdapat 7 spesies. Spesies T. turgidum tetraploid berasal dari kombinasi genom A dari T. uratu dan genom B dari spesies liar (Ae. spetoides). T. aestivum hexaploid berasal dari kombinasi genom AB dari T. turgidum dam genom D dari T. tauschi (Fehr 1987, Wittenberg 2004, Poehlman dan Sleper 2006, Shewry PR. 2009, Carver 2009) (Gambar 2). Gandum adalah tanaman subtropis namun daerah asal tanaman gandum ini tidak diketahui secara pasti. Diduga tanaman ini berasal dari daerah luas yang membentang dari Asia Tengah ke Timur. Bukti tertua bagi penanaman gandum berasal dari Syria, Turki, dan Irak. Tanaman gandum pertama kali dibudidayakan di Asia Barat Daya sejak 10.000 tahun yang lalu, sejak tahun 1800an kemudian menyebar ke wilayah beberapa penjuru dunia (Poehlman dan Sleper 2006, Wikipedia 2011).
Pada tahun 1529 Spanyol memperkenalkan gandum di Amerika dan pada tahun 1966 Spanyol juga menanamnya di Filipina. Semenjak itu mulai dilakukan perbaikan kualitas bulir, peningkatan hasil, peningkatan ketahanan kekeringan, masa simpan, hama dan penyakit (Poehlman dan Sleper 2006). Tanaman gandum diintroduksi di Indonesia sekitar tahun 1784 ditanam secara terbatas di beberapa pegunungan di Jawa dan Timor. Pada tahun 1969 melaksanakan program kerjasama ekonomi antara Indonesia dengan Amerika Serikat dengan nama PL480. PL (Public Low) 480 adalah kebijakan pemerintah Amerika Serikat untuk memberikan produk pangan kepada negara-negara berkembang. Pada tahun yang sama dimulailah impor gandum untuk membantu pembangunan ekonomi dalam jangka panjang. Seiring dengan berjalannya waktu, Amerika menghentikan hibah gandum ke Indonesia. Penghentian tersebut telah sesuai dengan kesepakatan yang tercantum dalam PL 480. Untuk memenuhi kebutuhan gandum di dalam negeri dilakukan impor dari Australia dan Amerika (Nur et al. 2013).
Namun karena iklim di Indonesia tidak sesuai dengan pertumbuhan tanaman gandum dan belum adanya usaha pemerintah untuk mengembangkan budidaya gandum maka tanaman gandum tidak berjalan dengan baik (Wiyono 1980; Sastrosumarjo 1987). Saat ini, setelah uji coba adaptasi multilokasi di berbagai daerah, tanaman gandum dapat tumbuh dan berproduksi tinggi di Indonesia tetapi hanya terbatas pada dataran tinggi (>1000 m dpl).
Morfologi dan Budidaya Tanaman Gandum
Tanaman gandum termasuk tanaman herba setahun/semusim dengan karakteristik menyerbuk sendiri (self-polinated), penyerbukan silang hanya 1-4%. Gandum memiliki batang beruas sebanyak 6 ruas, berongga seperti tanaman padi. Semua spesies Triticum mempunyai morfologi yang hampir serupa, seperti pada Gambar 3. Daun pertama (koleoptil), berbentuk silinder, diselaputi plumula yang terdiri dari dua sampai tiga helai daun. Daun gandum terdiri dari tangkai pelepah, helai daun, dan ligula dengan dua pasang telinga pada dasar helai daun dengan tulang daun sejajar dan memanjang. Daun tanaman gandum tumbuh tegak/melengkung berbentuk pita dan daun yang sudah tua menjadi kuning dan melengkung ke bawah (Wiyono 1980).
Gambar 3. Morfologi tanaman gandum (1.benih, 2&3.akar adventif, 4.daun, 5. ekor gabah gandum, 6.malai, 7.bulir, 8.pelepah daun, 9.tangkai malai, 10 &11.batang utama, 12.anakan, 13.ruas, 14.daun telinga, 15.lidah daun,16.buku) (Wiyono 1980).
Pembungaan pada gandum bersifat majemuk dan berbentuk malai terdiri atas bulir-bulir. Pembungaan dimulai pada sepertiga bagian tengah malai kemudian menyebar secara bersamaan ke arah ujung dan pangkal malai. Bunganya bermekaran pada pertengahan pagi. Biasanya untuk varietas musim semi, digolongkan ke dalam varietas hari pendek, sedangkan varietas musim dingin digolongkan ke dalam varietas hari panjang. Kemampuan reseptif stigma berkisar antara 4-13 hari sedangkan viabilitas pollen sekitar 30 menit. Bulir yang berada pada bagian tengah malai dan bagian proksimal dari floret cenderung membesar. Kondisi masak fisiologis dicapai apabila kandungan kelembaban dari keseluruhan bulir yang terbentuk telah menurun antara 25-35%. Pada dasar spikelet terdapat glume yang halus dan beberapa varietas ada yang mempunyai glume berambut pendek. Tiap spikelet menghasilkan 2-3 biji (kernel), dan terdapat kulit ari (lemma dan palea) yang di dalamnya terdapat 3 anther dan 2 stigma dengan 1 ovarium. Lemma dan palea lazim disebut sekam (Gambar 4) (Stoskoff 1985; Phoelman dan Sleper 2006; Carver 2009).
bagian kulit ini dapat dipisahkan melalui proses penggilingan. Kulit merupakan bagian terluar dari biji gandum yang mempunyai fungsi untuk melindungi biji pada saat tumbuh dengan persentase terhadap biji gandum 15%. Endosperm adalah bagian terbesar dari biji sekitar 82.5% dan biasanya diproses menjadi tepung terigu. Germ adalah bakal biji gandum yang mempunyai persentase terhadap biji sekitar 2.5% (Gambar 5) (Haryati 2000 ;Welirang 2008).
Gambar 4. Malai, bulir dan bunga gandum (1. malai, 2. bulir, 3. gabah atas, 4. Gabah bawah, 5. bunga (2-5 buah), 6. kotak sari, 7. kepala putik, 8. tangkai sari, 9. putik, 10. palea, 11. lemma, 12. bunga gandum) (Wiyono 1980).
[image:33.595.111.496.188.399.2]Gandum dibudidayakan dengan cara menanam benihnya secara alur maupun tugal. Penanaman secara alur sering digunakan untuk menghindari kerebahan dan tujuan komersial. Sedangkan penanaman secara tugal digunakan untuk tujuan menghemat benih, memperoleh tingkat perkecambahan dan pertumbuhan yang seragam, dan biasanya menghasilkan angka produksi yang lebih tinggi. Gandum menghendaki suhu bagi pertumbuhannya, yaitu sekitar 15 – 25 oC. Untuk daerah tropis seperti Indonesia, suhu tersebut terdapat di dataran tinggi >800 m dpl (Wiyono 1980, Acquaah 2007). Gandum tidak toleran terhadap kekeringan, sensitif terhadap salinitas tanah, dan tidak dapat tumbuh pada daerah yang hangat dan suhu tinggi (Van Ginkel dan Villareal 1996). Pengaruh suhu tinggi terhadap perkembangan bulir pada serealia meliputi laju perkembangan bulir yang lebih cepat, penurunan berat bulir, biji keriput, berkurangnya laju akumulasi pati dan perubahan komposisi lipid dan polipeptida (Stone 2001).
Karakter Morfo-Fisiologis
Karakter morfo-fisiologis di dalam seleksi tanaman dapat digunakan untuk pemecahan masalah yang dihadapi pada setiap tanaman yang diteliti, seperti tanaman gandum yang dapat beradaptasi di dataran rendah. Analisis keragaman genetik menggunakan karakter morfologi dapat dipengaruhi oleh faktor lingkungan, oleh karena itu untuk menghasilkan data yang lebih baik harus di dukung dengan karakter fisiologi. Karakter morfologi juga dapat terjadi perubahan pada pertumbuhan tanaman bila terjadi cekaman lingkungan. Sebagai contoh cekaman suhu tinggi, dapat menyebabkan perubahan daun menjadi lebih tipis, dan keragaan biji menjadi keriput. Pengaruh cekaman lingkungan terhadap pertumbuhan tanaman sangat ditentukan oleh besarnya tingkatan cekaman yang dialami tanaman tersebut. Beberapa hasil penelitian menunjukkan bahwa akibat dari cekaman lingkungan menyebabkan penghambatan pertumbuhan tanaman dan penurunan laju fotosintesis. Penurunan laju fotosintesis berkaitan dengan beberapa faktor seperti penutupan stomata, dehidrasi kutikula dan membran sel mengurangi permiabilitasya terhadap CO2. Penurunan nyata biasanya terjadi pada tinggi
tanaman, luas tanaman, jumlah ruas pertanaman dan jumlah polong per ruas (Hoogenboom et al. 1987), penurunan jumlah bunga dan penurunan jumlah biji pertanaman (Korte et al. 1983). Respon pertama pada tanaman yang mendapat cekaman adalah terjadinya hambatan pembukaan stomata daun. Mekanisme pembukaan stomata daun pada tanaman dikontrol oleh terjadinya perubahan potensial air di daun (Pennypacker et al. 1990, Watanabe et al. 1991).
jumlah tertentu dan bervariasi tergantung pada varietas tanaman yang digunakan dan umur tanaman (Hamim 2004).
Adaptasi Tanaman terhadap Cekaman Suhu Tinggi
Adaptasi tanaman adalah kemampuan tanaman untuk menyesuaikan diri terhadap kondisi lingkungan yang spesifik seperti kondisi suhu, cahaya, dan ketersediaan mineral dan hara (Dubey 1996). Toleransi cekaman suhu didefinisikan sebagai kemampuan tanaman untuk tumbuh dan menghasilkan secara ekonomi pada suhu tinggi (Peet dan Willits 1998). Menurut Rao (2001) bahwa perbaikan adaptasi tanaman terhadap lingkungan dapat dicapai dengan perubahan lingkungan pertumbuhan dan pengembangan genotipe tanaman, tetapi yang paling efektif adalah gabungan antara keduanya. Peningkatan hasil panen disebabkan oleh perubahan agronomi melalui perbaikan adaptasi genetik untuk mengatasi kendala biotik dan abiotik dalam produksi tanaman serta meningkatkan potensial hasil genetik tetuanya dalam lingkungan yang sama. Untuk meningkatkan adaptasi tanaman terhadap cekaman abiotik yaitu dengan cara menyeleksi genotipe pada lingkungan cekaman suhu tinggi untuk mendapatkan galur-galur yang toleran dan karakter tanaman yang diinginkan seperti berproduksi tinggi (Blum 1983; Hall 1992). Indonesia sebagai daerah tropis yang mempunyai kisaran suhu dan kelembaban yang cukup bervariasi, yaitu sekitar 22-33 °C dan 55-97 % (BMKG 2013). Hal ini menjadi kekhawatiran apabila tanaman gandum ditanam pada dataran rendah sampai sedang, karena gandum membutuhkan suhu tidak lebih dari sekitar 15-25 °C untuk periode pembungaannya.
Stres suhu tinggi atau kombinasi dengan kekeringan, adalah kendala umum selama tahapan pengisian biji dan pembungaan pada banyak tanaman sereal pada daerah yang beriklim tropis. Sebagai contoh, stres suhu tinggi memperpanjang waktu pengisian biji-bijian, menurunkan berat kernel gandum hingga 7% pada musim semi (Gambar 6) (Wahid et al. 2007; Guilioni et al. 2003). Pengurangan serupa terjadi pada pati, protein dan isi minyak kernel jagung (Wilhelm et al. 1999) dan kualitas biji-bijian sereal lain (Maestri et al. 2002). Pada gandum, berat biji dan jumlah biji gandum tampaknya peka terhadap stres suhu tinggi, seperti jumlah butir per malai menurun dengan meningkatnya suhu (Ferris et al. 1999).
Gambar 6. Keragaan biji gandum: A. Biji yang normal, B. Biji akibat cekaman suhu tinggi.
Peningkatan suhu dapat menyebabkan perubahan distribusi geografis dan musim pertumbuhan tanaman pertanian dengan membiarkan ambang suhu untuk awal musim dan kematangan tanaman lebih awal (Porter 2005). Suhu yang sangat tinggi, menyebabkan kerusakan parah bahkan kematian sel dapat terjadi dalam beberapa menit, yang dapat dikaitkan sebagai keruntuhan organisasi selular (Schoffl et al. 1999). Menurut laporan dari Inter Panel Climate Change (IPCC), suhu global akan naik 0.3 °C per dekade (Jones et al. 1999) mencapai sekitar 1 dan 3 °C di atas nilai sekarang pada tahun 2025 dan 2100, dan menyebabkan pemanasan global.
Memahami mekanisme fisiologis tanaman dengan perubahan-perubahan kondisi lingkungan sangat penting untuk menciptakan strategi yang efisien untuk mengembangkan kultivar unggul tahan cekaman. Untuk memahaminya perlu adaptasi tanaman tersebut terhadap cekaman seperti perubahan suhu terutama pada suhu tinggi. Mekanisme respons tanaman terhadap cekaman juga bervariasi pada setiap jenis tanaman. Mekanisme respons tumbuhan terhadap cekaman dan peranan mekanisme toleransi terhadap cekaman merupakan informasi dasar dalam perbaikan tanaman (Wahid et al. 2007). Mekanisme pertahanan antioksidan adalah bagian dari adaptasi terhadap cekaman suhu tinggi, yang berkorelasi dengan akuisisi termotoleransi (Maestri et al. 2002).
Mekanisme termotoleransi yang paling sering diteliti adalah induksi HSPs (Heat Shock Proteins). Pada kondisi stres panas (suhu tinggi), tanaman memberikan respon dengan cara mensintesa satu set protein pelindung yaitu HSPs. Mekanisme toleransi terhadap suhu tinggi terkait ketahanan sitoplasma dengan adanya HSPs, perubahan komposisi lipid bilayer pada membran biologis, dan regulasi hormon. HSPs merupakan kelompok – kelompok protein yang toleran cekaman panas. Protein ini melibatkan beberapa famili protein yang terkonservasi dan memiliki mekanisme aktifitas penghantaran yang khas (Wahid et al. 2007). Struktur primer HSPs terkonservasi hampir pada seluruh organisme mulai dari prokariot sampai eukariot tingkat tinggi seperti hewan dan tanaman. HSPs berperan mencegah agregasi protein yang terdenaturasi oleh suhu tinggi dan mendorong renaturasi molekul protein yang sudah teragregasi (Iba 2002).
Pemulian Mutasi Tanaman Gandum melalui Mutagenesis Menggunakan EMS
Pemuliaan secara mutasi dapat diinduksi dengan mutagen fisik dan mutagen kimia. Umumnya mutagen fisik dapat menyebabkan mutasi pada tahap kromosom, sedangkan mutagen kimia umumnya menyebabkan mutasi pada tahapan gen atau basa nitrogen (Aisyah 2006). Awalnya, para pemulia tanaman menganggap bahwa induksi mutasi merupakan suatu teknik pemuliaan yang kurang meyakinkan. Namun, seiring dengan berkembangnya bioteknologi, keberhasilan regenerasi sel berdasarkan teori totipotensi, dan terbentuknya variasi somaklonal, maka induksi mutasi merupakan teknik dalam pemuliaan tanaman yang diharapkan. Teknik tersebut dapat menunjang perolehan varietas mutan baru yang bermanfaat bagi perkembangan gandum di Indonesia.
Pemuliaan tanaman dengan induksi mutasi bertujuan untuk perbaikan tanaman sehingga menghasilkan varietas unggul dengan hasil produksi lebih tinggi dari tetuanya dan mempunyai bentuk yang berbeda pula. Menurut Chopra (2005) perbaikan varietas tanaman dapat dilakukan dengan menggunakan mutagen fisik atau kimia untuk meningkatkan keragaman genetik. Mutasi adalah suatu perubahan baik terhadap gen tunggal, terhadap sejumlah gen dan terhadap susunan kromosom atau perubahan urutan (sequence) nukleotida DNA kromosom, yang mengakibatkan terjadinya perubahan pada protein yang dihasilkan. Mutasi dapat terjadi pada setiap bagian tanaman dan fase pertumbuhan tanaman, namun lebih banyak terjadi pada bagian yang sedang aktif mengadakan pembelahan sel seperti meristem.
Gandum bukan merupakan tanaman asli Indoneia maka keragaman genetik gandum yang terdapat di Indonesia masih sangat terbatas. Beberapa varietas gandum di introduksi dari International Maize and Wheat Improvement Center, Mexico (CIMMYT) dan dari beberapa negara seperti India, Pakistan dan Turki yang kondisi iklimnya kurang lebih sama dengan Indonesia dibandingkan dengan varietas asal Eropa. Setelah melalui proses pengujian adaptasi dan daya hasil selama beberapa generasi kemudian beberapa varietas introduksi dilepas oleh KEMTAN menjadi varietas unggul nasional seperti Dewata (galur DWR 162 dari India) dan Selayar (galur HHAHN/2 dari CIMMYT) (Dahlan et al. 2003). Varietas – varietas tersebut memiliki potensi untuk dikembangkan pada lahan pertanian di Indonesia hanya saja terbatas pada dataran tinggi. Sedangkan dataran tinggi di Indonesia umumnya telah ditanami dengan sayur mayur dan hortikultura lainnya yang mempunyai nilai ekonomis lebih tinggi. Oleh karena itu program pemuliaan tanaman gandum di Indonesia diarahkan untuk merakit varietas unggul yang dapat beradaptasi pada dataran rendah (< 400 m dpl). Poehlman dan Sleper (2006) menyatakan bahwa kegiatan pemuliaan gandum ditujukan untuk memperoleh varietas unggul berproduksi tinggi, meningkatkan kualitas produk, meningkatkan stabilitas hasil berkaitan dengan upaya untuk mendapatkan varietas yang seragam, dan mampu beradaptasi pada berbagai kondisi lingkungan.
meristematik (Maluszynski 1995; Ahloowalia 1995). Kalus bersifat meristematik sehingga lebih responsif terhadap mutagen dibandingkan dengan sel-sel dewasa.
Penggunaan mutagen dalam kultur in vitro dapat meningkatkan keragaman genetik sehingga dapat menghasilkan mutan baru. Proses mutasi alami (spontan) biasanya sangat lambat (10-6 s/d 10-7) sehingga perlu mutagen untuk menginduksi frekuensi, dan kecepatan mutasi tanaman. Salah satu mutagen yang paling potensial (Chopra 2005), paling efektif dan banyak digunakan (Medina et al. 2005) serta digunakan pada berbagai jenis organisme mulai dari virus sampai mamalia (Sega 1984) adalah mutagen kimia yaitu EMS. EMS sering digunakan dalam penelitian karena mudah diperoleh, murah dan tidak bersifat mutagenik setelah terhidrolisis (Van Harten 1998; Natarajan 2005). Mutagen kimia dapat diintroduksi ke dalam jaringan tanaman dan bahkan sel sehingga dapat menyebabkan jumlah mutasi tinggi dibandingkan dengan cara lain tetapi tergantung dari konsentrasi bahan kimia, waktu perlakuan, suhu, dan kadar air bahan eksplan (Nasir 2002).
EMS pada umumnya menyebabkan mutasi titik yaitu terjadinya delesi pasangan basa tertentu dalam kromosom. Senyawa EMS merupakan senyawa alkali yang efektif sebagai mutagen untuk tanaman tingkat tinggi (Van Harten 1998; Greene et al. 2003) dan dapat mengubah lokus tertentu tanpa menginduksi sejumlah besar mutasi yang terpaut dekat dengan lokus tersebut. Gugus alkil bereaksi dengan DNA dengan cara mengalkilasi basa purin dan pirimidin. Alkil dapat terjadi pada atom O-6 dari basa guanin sehingga berakibat perubahan pasangan dari sitosin menjadi berpasangan dengan timin yang mengakibatkan perubahan kode genetik dari GC menjadi AT pada generasi sel berikutnya (Sega 1984). Penggunaan EMS untuk meningkatkan terjadinya mutasi telah banyak dilaporkan, diantaranya untuk menghasilkan mutan pada tanaman kedelai melalui mutagenesis in vitro (Grabau et al. 1995), menghasilkan tanaman gandum yang cepat berbunga dan masaknya biji (Vismanathan dan Reddy 1996), dan memperoleh tanaman cabai tahan ChiVMV (Manzila et al. 2010).
Kultivar mutan gandum yang dilepas di berbagai negara, adalah 1 mutan (Argentina, Chili, Finlandia, Hongaria, dam Swiss), 2 mutan ( Brazil, Bulgaria, Jepang, Jerman dan Itali), 3 mutan ( Amerika dan Mongolia), 4 mutan ( India ), 6 mutan ( Pakistan), 36 mutan (Rusia), 60 mutan (Irak), 124 mutan (Cina) (Cheng et al. 1990 dalam Soedjono 2003).
Seleksi in vitro
2009). Seleksi ketahanan terhadap suhu tinggi pada tanaman kentang lebih tinggi 8 °C dari suhu optimum untuk pembentukan umbi mikro yaitu 20 °C menjadi 28 °C kemudian di uji lapang beriklim panas. Hasil dari evaluasi tersebut diperoleh beberapa mutan toleran suhu tinggi. Sedangkan pada bawang putih mendapatkan umbi mutan secara in vitro pada suhu 32 °C.
Seleksi in vitro sangat efisien karena tempat yang dibutuhkan relatif sedikit, kondisi seleksi dapat homogen dan terkendali serta menghasilkan efektifitas seleksi tinggi. Penggunaan seleksi in vitro pada tingkat sel dengan menggunakan agen penyeleksi diharapkan dapat dihasilkan karakter yang diinginkan. Menurut Ali (2006) teknik induksi mutasi dikombinasikan dengan seleksi in vitro seringkali digunakan untuk membentuk keragaman genetik baru dan meningkatkan kemungkinan diperolehnya varian baru yang beradaptasi dengan lingkungan tertentu. Tanaman hasil regenerasi dari jaringan yang dapat mengatasi kondisi seleksi in vitro, diharapkan mempunyai fenotipe toleran suhu tinggi dan dapat beradaptasi pada dataran rendah. Beberapa hasil penelitian seleksi in vitro untuk mendapatkan galur baru antara lain pada tanaman gandum tahan kekeringan (Bajji et al. 2004) dan tanaman barley tahan suhu beku (Tantau et al. 2004).
Seleksi in vivo
Induksi mutasi dan seleksi in vitro (seleksi di laboratorium) diharapkan dapat menghasilkan keragaman genetik yang diinginkan. Oleh karena itu untuk memperoleh genotipe-genotipe dengan karakter yang diinginkan maka perlu dilakukan seleksi in vivo (seleksi di lapangan). Seleksi di lapangan merupakan salah satu kegiatan utama dalam pemuliaan tanaman yaitu menyeleksi sejumlah individu dari suatu populasi dan membiarkannya membentuk generasi baru. Menurut Falconer dan Mackay (1996) seleksi ini merupakan salah satu cara untuk mengubah frekuensi gen dengan memilih karakter yang diinginkan dan tidak mengambil karakter yang tidak diinginkan.
Seleksi pada tanaman menyerbuk sendiri seperti pada tanaman gandum dapat dibagi menjadi tiga metode, yaitu seleksi silsilah (pedigree), Seleksi bulk dan seleksi turunan biji tunggal (single seed descent/SSD) (Chahal dan Gosal 2003; Poehlman dan Sleper 2006). Dalam penelitian ini metode yang digunakan adalah pedigree. Prinsip seleksi pedigree adalah dilakukan pada generasi awal dengan tingkat segregasi tinggi (M2 atau M3), berulang dilaksanakan terhadap individu terbaik dari famili terbaik pula sampai tercapai tingkat homozigositas tinggi dan turunan setiap genotipe di catat secara lengkap. Syarat yang berlaku pada metode pedigree adalah mempunyai karakter kuantitatif dengan nilai heritabilitas yang tinggi (> 50%).
generasi awal, dan penggaluran baru dilakukan setelah mencapai tingkat homozigositas tinggi (M6). Seleksi efektif jika sifat yang dikehendaki dapat diwariskan. Efisensi seleksi sangat ditentukan oleh karakter seleksi yang digunakan sehingga sebelum melakukan seleksi perlu terlebih dahulu ditentukan kriteria seleksi. Karakter yang dapat digunakan sebagai kriteria seleksi adalah karakter yang memiliki nilai heritabilitas tinggi (Roy 2000; Chahal dan Gosal 2003).
INDUKSI MUTASI DAN SELEKSI
IN VITRO
TANAMAN
GANDUM (
Triticum aestivum
L.)
Abstrak
Perbaikan genetik tanaman gandum toleran suhu tinggi pada dataran rendah dapat dilakukan dengan peningkatan keragaman genetik dari genotipe/galur yang memiliki toleran suhu tinggi. Peningkatan keragaman genetik hanya mengandalkan pada introduksi galur-galur gandum toleran suhu panas di negara asalnya seperti Turki, India dan Meksiko. Tanaman gandum merupakan tanaman subtropik yang diupayakan dapat dikembangkan di Indonesia. Tujuan penelitian ini adalah untuk memperoleh genotipe-genotipe tanaman gandum yang toleran suhu tinggi pada dataran rendah. Penelitian ini dilaksanakan di Laboratorium Kultur Jaringan BB-Biogen dari bulan Juli 2011 sampai Desember 2013. Materi genetik yang digunakan adalah dua varietas yaitu Dewata, Selayar, dan empat genotipe yaitu Alibey, Oasis, Rabe dan HP-1744. Penelitian dilakukan terdiri atas empat tahap yaitu 1. Induksi pembentukan kalus terbaik menggunakan media MS+ 2.4-D 3 mg/l (dipilih dua genotipe yang terbaik), 2. Induksi mutasi kalus embriogenik menggunakan EMS, 3. Seleksi kalus in vitro pada suhu 27– 35°C dan 4. Regenerasi kalus hasil seleksi in vitro. Penelitian menunjukkan bahwa hasil induksi kalus yang mempunyai nilai tertinggi diperoleh pada genotipe Dewata 76% dan Selayar 70%. Semakin tinggi konsentrasi EMS dan semakin lama waktu perendaman menurunkan persentase pertumbuhan kalus. LC20 -LC50
genotipe Dewata adalah EMS 0.3% waktu 30 menit sedangkan LC20 -LC50
genotipe Selayar adalah EMS 0.1% waktu 60 menit. Genotipe Dewata dan Selayar mempunyai toleran suhu tinggi pada suhu 27°C dengan nilai 84 dan 72%. Semakin tinggi suhunya maka semakin menurun toleransi tanaman tersebut. Suhu semakin tinggi, pertumbuhan kalus semakin sedikit dan pada suhu tertinggi yaitu suhu 35°C tidak ada pertumbuhan kalus atau mengalami kematian. Regenerasi jumlah tunas genotipe Selayar dan Dewata yang tertinggi dihasilkan pada media RG2 (MS + BA 0.1 ml/l + kinetin 2 mg/l + tyrosin 0.05 gr/l + sorbitol 6% + sukrosa 3%) yaitu 36 dan 44%.
Kata Kunci: Induksi mutasi, gandum, EMS, seleksi in vitro, regenerasi.
Abstract
2.4-D (the best two genotypes was choose). The second stage was induced mutation of embryogenic callus using EMS. The third stage was in vitro selection at temperature 27–35°C. The last stage was callus regeneration from in vitro selection. The best result for callus production was 76% for Dewata and 70% for Selayar genotypes. The higher concentration of EMS and the longer the soaking time used decreased callus growth percentage. LC20 -LC50 of Dewata genotype
was 0.3% EMS at 30 minutes and LC20 -LC50 of Selayar genotype was 0.1% EMS
at 60 minutes. Dewata and Selayar genotypes were tolerant to temperature 27°C with value of 84 and 72%. The higher the temperature the more diminished the tolerant adaptation of the plants. At higher temperature, callus growth was inhibited and at the highest temperature (35 °C) callus did not grow and die. Highest number of regenerated shoots produced on media RG2 (medium MS containing 0.1 mg/l BA, 2 mg/l kinetin, 0.05 g/l tyrosine, 6% sorbitol and 3% sucrosa) at 36 and 44%.
Keywords: induced mutation, wheat (Triticum aestivum), EMS, in vitro selection, regeneration.
PENDAHULUAN
Perbaikan genetik gandum perlu dilakukan karena terbatasnya sumber daya genetik gandum asal daerah subtropis. Teknik in vitro, seperti mutasi dan seleksi in vitro merupakan solusi alternatif dalam memperluas keragaman genetik. Kombinasi pemuliaan mutasi dan seleksi in vitro dapat meningkatkan keragaman genetik secara lebih efektif dan efisien sehingga dihasilkan varietas unggul (Maluszynski et al. 1995; Jain 2010). Penggunaan bahan tanaman berupa kalus, bersifat meristematik sehingga lebih responsif terhadap radioaktif dan zat kimia dibandingkan dengan sel-sel dewasa. Pembentukan kalus pada tanaman secara umum tergantung pada genotipe, tipe jaringan, ZPT dan media yang digunakan (Bahieldin et al. 2000, Rashid et al. 2002). Menurut Sarker dan Biswas (2002) faktor lain yang mempengaruhi pembentukan kalus adalah pemilihan eksplan seperti embrio dewasa, embrio muda, biji, endosperm, tunas, dan ujung akar. Penggunaan eksplan yang tepat juga mempengaruhi regenerasi tanaman gandum secara in vitro.
Beberapa studi menunjukkan bahwa dosis optimum dalam induksi mutasi yang dapat menghasilkan mutan terbanyak umumnya diperoleh di sekitar lethal dose (LD50 ) atau lethal concentration (LC50). Variabilitas mutan tertinggi terdapat
pada mutan hasil mutagen kimia di sekitar LC20 dan LC50 (Datta 2001). Induksi
mutasi yang dilanjutkan dengan seleksi ketahanan terhadap suhu tinggi secara in vitro telah dilakukan pada tanaman kentang dan bawang putih dan telah berhasil memperoleh mutan toleran suhu tinggi (Das et al. 2000).
Cekaman suhu tinggi menjadi faktor pembatas dalam usaha perluasan gandum di dataran rendah, karena pada dasarnya gandum merupakan tanaman subtropis yang menghendaki suhu rendah untuk proses pertumbuhan dan perkembangannya. Cekaman suhu tinggi artinya terjadi kenaikan suhu di luar batas selama jangka waktu yang cukup untuk menyebabkan kerusakan pertumbuhan dan perkembangan tanaman (Sopandie 2013).
Penelitian ini perlu dilakukan untuk induksi mutasi kalus embriogenik menggunakan EMS dan seleksi in vitro untuk ketahanan terhadap suhu tinggi, sehingga teknik tersebut dapat menunjang perolehan genotipe mutan baru yang toleran dataran rendah tropis. Tujuan penelitian ini adalah untuk mendapatkan genotipe mutan unggul hasil seleksi in vitro yang toleran pada dataran rendah.
METODE PENELITIAN
Penelitian dilakukan di Laboratorium Biologi Sel dan Jaringan Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB-BIOGEN), dimulai bulan Juli 2011 s/d Desember 2013. Bahan tanaman yang digunakan dalam penelitian ini adalah immature embryo yang diisolasi dari biji 6 materi genetik yaitu empat genotipe introduksi HP 1744, Alibey, Oasis, Rabe dan dua varietas nasional yaitu Dewata dan Selayar. Media yang digunakan berupa media dasar MS yang ditambahkan zat pengatur tumbuh (ZPT). Penelitian dilakukan dalam 4 tahap yaitu: 1) Induksi pembentukan kalus embriogenik. 2) Induksi mutasi pada kalus embriogenik menggunakan EMS. 3) Seleksi in vitro pada suhu 27 – 35 °C. 4) Regenerasi kalus mutan putatif hasil seleksi in vitro. 1. Induksi Pembentukan Kalus Embriogenik.
Oasis dan Rabe. Perlakuan terdiri atas 10 ulangan, masing-masing ulangan terdiri atas satu botol dengan 10 eksplan embrio. Data dianalisis menggunakan program SAS 9.1 dan jika terdapat perbedaan nyata maka analisis dilanjutkan menggunakan DMRT (Duncan Multiple Range Test) pada taraf 5%. Pengamatan dilakukan setelah kalus berumur 2 minggu meliputi persentase pertumbuhan kalus embriogenik terbaik, dan diameter kalus. Penentuan warna kalus dengan skoring 1 = putih, 2= putih kekuningan dan 3= kuning. Dari 6 genotipe yang diuji selanjutnya dipilih 2 genotipe yang menghasilkan kalus embriogenik terbaik. Genotipe terpilih tersebut digunakan sebagai materi uji penelitian selanjutnya. 2. Induksi Mutasi pada Kalus Embriogenik Menggunakan EMS.
Tujuan: Mendapatkan keragaman genetik gandum dengan induksi mutasi menggunakan EMS pada kisaran Lethal concentration 20 (LC20) – LC50 pada
beberapa konsentrasi dan waktu perendaman eksplan. Eksplan yang digunakan adalah kalus embriogenik dari 2 genotipe terpilih hasil penelitian 1. Penelitian dilakukan dengan rancangan acak lengkap faktorial, faktor pertama yaitu konsentrasi EMS 3 taraf (0.1, 0.3, dan 0.5 % ) dan faktor kedua yaitu waktu perendaman 5 taraf (0, 30, 60, 120 dan 180 menit) untuk setiap genotipe. Perlakuan terdiri atas 5 ulangan, masing-masing ulangan terdiri satu botol dengan 5 kalus. Pengamatan dilakukan setelah kalus berumur 1 minggu meliputi jum