Summary Concentrations of Pb in individual stem xylem rings of 5-year-old Norway spruce trees (Picea abies (L.) Karst.) were determined after one growing season in soil containing a low, medium or high concentration of Pb. In trees in the control and low-Pb soil treatments, Pb concentrations increased from the outer annual rings toward the stem center, whereas in trees grown in the soil treatment containing a medium Pb concentration, all of the four tree rings analyzed contained similar concentrations of Pb. Although trees grown in the high-Pb soil treatment had higher concentrations of Pb in the outer annual rings than both control trees and trees in the low-Pb soil treatment, the highest concentrations of Pb were never observed in the outermost rings, which were formed during the period of exposure to increased soil Pb. We conclude that radial distribution patterns of Pb in Norway spruce stems do not directly reflect changes in soil Pb concentration but depend on several internal, physiological factors and, there-fore, do not provide reliable information about past variations in Pb contamination of soil.
Keywords: Pb, Picea abies, tree rings, xylem.
Introduction
Trace element concentrations of tree rings have been used for retrospective biomonitoring of past environmental pollution (Ferretti et al. 1993, Momoshima and Bondietti 1994, Eklund 1995). The method, called dendroanalysis (Gilboy et al. 1976), is based on the assumption that element concentrations in tree rings are closely related to the environmental abundance of an element at the time the growth rings are formed. Thus, chang-ing element concentrations in the environment should be re-flected in changing concentrations of the same elements in tree rings. It is assumed that trees incorporate and store absorbed elements mainly in currently growing xylem rings. However, this assumption has not been validated, although dendroana-lytical techniques have been used in many investigations (Bur-ton 1985, Cutter and Guyette 1993, Hagemeyer 1993). Donnelly et al. (1990) conducted an experiment in which 2-year-old Picea rubens Sarg. trees were grown hydroponi-cally and supplied with Pb either in the first or second year of
growth and found that stem xylem formed in the absence of Pb contained significant amounts of Pb.
Hagemeyer and Lohrie (1995) studied uptake and incorpo-ration of Cd and Zn in annual xylem rings of young Picea abies (L.) Karst. trees and found that growth rings formed during treatment with high concentrations of Cd and Zn contained lower concentrations of these metals than older growth rings. However, the total amount of Cd and Zn incorporated into the annual xylem rings increased from the stem center toward the outer xylem rings. It was suggested that the lower metal con-centrations in outer rings than in inner rings could be explained by the greater wood dry matter of the outer rings compared with the rings at the stem center. In the present study, we examined the accumulation of the relatively less mobile ele-ment Pb in individual annual xylem rings and bark of 5-year-old Picea abies trees grown for one year in Pb-contaminated soil.
Materials and methods
Experimental conditions
Four-year-old Norway spruce trees (Picea abies (L.) Karst.) were obtained from a tree nursery and planted in pots contain-ing 5 liters of a mix of composted garden soil, peat and sand (23/17/10, v/v), to which fixed concentrations of Pb were added to provide four soil treatments (control (no Pb added), low-Pb, medium-Pb and high-Pb). Before planting, Pb was added as Pb(NO3)2 in aqueous solution to the soil treatments to adjust the Pb concentration of the soil to the extractable Pb concentrations (in 1 M ammonium acetate) given in Table 1. After adding Pb, the soils were stored for 2 months and fre-quently mixed to stabilize ion exchange equilibria. Both at planting and at the time of harvest, soil samples were taken from each pot in the four treatment groups and analyzed for extractable Pb (Table 1). Variations in extractable Pb concen-trations of the substrate during the experimental period are presumed to be caused by shifts in the exchange equilibrium between mobile and immobile Pb, as a result of pH changes, microbial degradation of organic matter, leaching of soluble substances from the containers, and plant uptake.
Radial distributions of Pb in stems of young Norway spruce trees
grown in Pb-contaminated soil
JÜRGEN HAGEMEYER and THOMAS WEINAND
Bielefeld University, Faculty of Biology, Department of Ecology, D-33501 Bielefeld, Germany
Received August 11, 1995
Tree Physiology 16, 591--594
At the end of January 1993, the Norway spruce trees were planted in pots containing the Pb-treated soils. Each treatment group included more than 30 plants. The potted trees were kept in an unheated greenhouse until mid March when they were moved outdoors until the time of harvest in November 1993. During dry weather the trees were irrigated with tap water.
Harvest procedure
After the end of the growing season, November 1993, 12 trees per treatment were randomly chosen for harvest. A 5-cm-long section was cut from the basal part of the stem directly above the soil surface. Adhering dust and epiphytes were removed from the bark with a brush. The surfaces at both ends of each stem section were cut with a microtome to improve the visibil-ity of growth rings. The stem section was then clamped upright in a holder and the bark was removed with a scalpel. For analysis, the bark was divided into the outer dead part and the inner live part. The wood was then dissected with a scalpel and divided into single annual growth increments, starting from the outermost ring. Thin layers near ring boundaries were dis-carded in order to obtain a clear separation of tissue of adjacent rings. Because the innermost 1989 ring was usually very nar-row and did not yield sufficient material for a separate analysis, it was combined with the 1990 growth ring. To minimize sample contamination, scalpel blades were cleaned with iso-propanol between different harvest steps. Bark and wood sam-ples were collected in paper bags and dried at 105 °C to constant weight.
Analytical procedures
Samples of wood and bark were wet ashed in Teflon pressure vessels with concentrated HNO3. Whenever sufficient sample material was available, analyses were carried out in replicates. Concentrations of Pb in wood and bark were determined by graphite furnace atomic absorption spectrophotometry (Per-kin-Elmer 5100). For all samples, the method of standard additions calibration was applied including a Pd-Mg(NO3)2 matrix modifier.
To determine exchangeable Pb concentrations in soil sam-ples, 5 g of air-dried soil was extracted with 50 ml of 1 M ammonium acetate. The extracts were stirred for 2 h and then filtered. The filtrates were analyzed by flame atomic absorp-tion spectrophotometry (Perkin-Elmer 380). Soil pH (H2O)
was measured in 10 g of air-dried soil in 25 ml of demineral-ized water after stirring for 15 min.
The quality of the analytical results was checked with the certified reference material Tea (GBW 08505) of the National Research Centre for Certified Reference Material, People’s Republic of China. Based on 43 determinations, a mean Pb concentration of 1.03 ± 0.15 ppm was found which was in good agreement with the certified value of 1.06 ± 0.1 ppm.
Statistical analysis
Lead concentrations of different annual rings within a treat-ment group were compared with the Wilcoxon matched pairs signed rank test (Sachs 1992). An analysis of variance includ-ing the Scheffé test was conducted to compare data between treatment groups.
Results
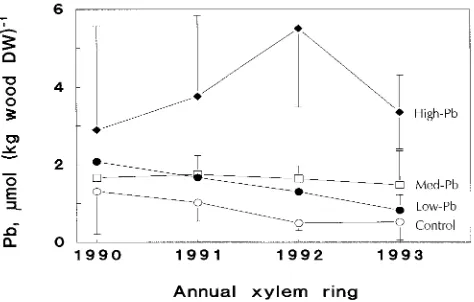
Concentrations of Pb in stem wood increased with increasing Pb concentration in the soil (Figure 1). Radial distribution patterns of Pb varied among the soil treatments. In control trees, Pb concentrations increased from the outer rings toward the stem center. This pattern was also observed in the low-Pb treatment, although at a higher range of concentrations. In trees in the medium-Pb soil treatment, Pb concentrations increased relative to that in control trees in all annual rings, but particu-larly in the outermost xylem ring (P < 0.01). As a consequence, these trees had similar Pb concentrations in all four of the xylem rings analyzed. For trees in the high-Pb soil treatment, highest Pb concentrations were found in the 1992 xylem ring, which was formed before the experiment began. Lead concen-trations in the outermost ring, which developed during the period of exposure to Pb-contaminated soil, were significantly lower (P < 0.01) than in the 1992 xylem ring.
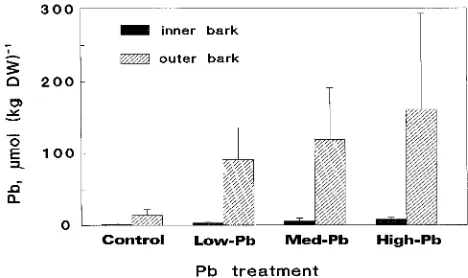
Concentrations of Pb in bark were higher than in stem wood (Figure 2) and higher in outer bark than in inner bark. In both outer and inner bark, Pb concentrations increased with increas-Table 1. Soil concentrations of Pb (1 M ammonium acetate extracts)
and pH (H2O) at the beginning and at the end of a pot culture experiment with young Norway spruce trees. Values are means ± SD of 11--12 containers, otherwise values are means of two samples.
Treatment Pb (µmol kgDW−1) Soil pH
Before After Before After
Control 6 4.7 ± 1.7 6.6 7.3
Low-Pb 74 74.9 ± 4.9 6.5 7.3
Medium-Pb 127 147.0 ± 41.2 6.5 7.3
High-Pb 646 575.0 ± 106 6.6 7.2
Figure 1. Concentrations of Pb in annual xylem rings of stems of 5-year-old Norway spruce trees. Plants were cultivated for one grow-ing season (1993) in soil containgrow-ing a low, medium or high concentra-tion of Pb. Values are means ± SD of 9--11 plants. Some error bars have been omitted for clarity.
ing soil Pb concentration. Lead concentrations of the outer-most annual xylem rings (1993) were significantly correlated with those of the outer bark (r = 0.570, n = 38, P < 0.001) and the inner bark (r = 0.595, n = 37, P < 0.001).
Discussion
Although the Pb concentration of the soil was increased sharply in the 1993 growing season, the highest Pb concentra-tions were not found in the growth ring formed during that period. Under conditions of increased Pb availability in the soil, the overall Pb uptake of the Norway spruce trees was enhanced. However, the additional quantities of absorbed Pb were deposited in both currently growing wood and in older growth rings. Therefore, Pb concentrations in annual xylem rings do not represent a chronological record of variations in substrate Pb concentrations.
Deposition of Pb in the inner xylem rings can be explained by the pattern of sap flow in the stem. In coniferous trees the xylem stream passes through several annual rings at the same time. Èermák et al. (1992) found that, in Picea abies trees with 18 xylem rings, between 10 and 12 rings conduct the xylem flow. Thus, there is a direct supply of absorbed metals from the root system via the xylem sap stream to several xylem rings at the same time. It can reasonably be assumed that, in the five-year-old Norway spruce trees studied, all xylem rings were part of the conducting system and were supplied with Pb. Control trees and trees in the low-Pb soil treatment had similar radial distribution patterns of Pb, though Pb concentra-tions in all of the annual growth rings were higher in trees in the low-Pb soil treatment than in control trees. Lead concen-trations increased from the outermost growth rings toward the stem center. Similar distribution patterns have also been found for Cd and Zn (Hagemeyer and Lohrie 1995). Such patterns may be related to spatial differences in the cation binding capacity of the wood tissue. In mature red spruce trees (Picea rubens), the cation binding capacity of xylem tissue increased from the outer growth rings to the stem center (Momoshima and Bondietti 1990). Because the Ca concentration of the
xylem tissue was highest at the stem center, Momoshima and Bondietti (1990) concluded that xylem Ca concentration in-creased with increasing cation binding capacity of the tissue. A similar explanation may account for the distribution of Pb in xylem tissue of the Norway spruce trees used in the present investigation.
Concentrations of Pb were higher in bark than in stem wood. These tissues are connected by rays crossing the cambial zone (Van Bel 1990), which suggests that there may be radial trans-port of elements, including Pb, between them. In suptrans-port of this hypothesis, we found close correlations between Pb con-centrations in xylem and bark. The transport of toxic sub-stances, such as Pb, to the outer bark, where they will become incorporated in tissues that die and are eventually shed by the tree, may constitute a detoxification mechanism.
Because elements can move through rays in the radial direc-tion (Ziegler 1964), radial transport processes could affect Pb distribution patterns within the xylem. Furthermore, an ex-change of minerals between tissues of adjacent annual rings is possible through the pits, which connect tracheids of different rings (Bosshard 1976). Radial transport of mineral elements has been described for mature trees (Ziegler 1968, Bamber 1976, Daube 1883). The radial transport of toxic substances to be deposited in inner stem parts was suggested as part of a detoxification mechanism (Stewart 1966). However, the extent of such radial movements of Pb in stems is not known.
Our study provided no indication of the stability of the observed Pb distribution patterns. However, because an appre-ciable part of Pb in wood of coniferous and of ring-porous trees appears to be mobile (Balk and Hagemeyer 1994, Hagemeyer and Shin 1995), it seems possible that the radial distribution pattern of Pb will change, perhaps in response to variations in xylem sap composition. Seasonal changes in pH (Glavac et al. 1990) and in the concentrations of cation-complexing organic molecules (e.g., amino acids or citrate) in xylem sap have been described in various tree species (Sauter 1981, Rennenberg et al. 1994, Schneider et al. 1994). Such changes could give rise to remobilization of exchangeably bound Pb in the xylem. Hagemeyer and Shin (1995) investigated the potential mobil-ity and binding strength of metal cations in wood of mature Pinus sylvestris L. trees and found that, depending on the radial position in the stem, only about 15--40% of the total Pb was immobilized. The major portion of Pb was exchangeably bound. Thus, the possibility of changes in the radial tion of an element should be considered when radial distribu-tions are evaluated in terms of pollution records.
We conclude that the radial distribution of Pb in Norway spruce stems depends more on internal physiological factors than on variations in the concentration of the external Pb supply and, therefore, Pb concentrations in annual xylem rings do not represent a chronological record of variations in sub-strate Pb concentrations (cf. Donnelly et al. 1990). Some of the factors affecting Pb distributions in xylem are: (1) retention of Pb in the root system, which can result in a delayed transport to the stem (Donnelly et al. 1990); (2) the amount of xylem sap flow through individual rings that determines Pb input; (3) the cation binding capacity of growth ring tissue; and (4) the Figure 2. Concentrations of Pb in inner live and outer dead stem bark
of 5-year-old Norway spruce trees after cultivation for one growing season in soil containing a low, medium or high concentration of Pb. Values are means ± SD of 8--11 plants.
potential for remobilization of deposited Pb caused by vari-ations in xylem sap composition. Thus, the observed distribu-tion patterns of Pb represent the integrated results of effects of several factors, which can vary with time, making it difficult to interpret the physiological significance of element concentra-tions in tree rings.
Acknowledgment
We thank Prof. Dr. S.W. Breckle, Bielefeld University, for his help and continued interest in the study.
References
Balk, G. and J. Hagemeyer. 1994. Extraction of Cd and Pb from stem wood of oak trees (Quercus petraea (Matt.) Liebl.) with various solutions. Angew. Bot. 68:191--194.
Bamber, R.K. 1976. Heartwood, its function and formation. Wood Sci. Technol. 10:1--8.
Bosshard, H.H. 1976. Jahrringe und Jahrringbrücken. Schweiz. Z. Forstwes. 127:675--693.
Burton, M.A.S. 1985. Tree rings. In Historical Monitoring. A Techni-cal Report. Monitoring and Assessment Research Centre, Univ. London, pp 175--202.
Èermák, J., E. Cienciala, J. Kuèera and J.-E. Hällgren. 1992. Radial velocity profiles of water flow in trunks of Norway spruce and oak and the response of spruce to severing. Tree Physiol. 10:367--380. Cutter, B.E. and R.P. Guyette. 1993. Anatomical, chemical, and
eco-logical factors affecting tree species choice in dendrochemistry studies. J. Environ. Qual. 22:611--619.
Daube, W. 1883. Chemische Analysen des Kern- und Splintholzes wichtiger Waldbäume. Forstl. Blätter 20:177--192.
Donnelly, J.R., J.B. Shane and P.G. Schaberg. 1990. Lead mobility within the xylem of red spruce seedlings: implications for the development of pollution histories. J. Environ. Qual. 19:268--271. Eklund, M. 1995. Cadmium and lead deposition around a Swedish
battery plant as recorded in oak tree rings. J. Environ. Qual. 24:126--131.
Ferretti, M., R. Udisti and E. Barbolani. 1993. Mineral nutrients and trace metals in tree rings of Pinus sp. J. Anal. Chem. 347:467--470.
Gilboy, W.B., R.E. Tout and N.M. Spyrou. 1976. Dendroanalysis: the study of trace elements in tree rings. Proc. ERDA, X- and Gamma-Ray Symp., Ann-Arbor, MI, pp 164--165.
Glavac, V., H. Koenies and U. Ebben. 1990. Seasonal variation of calcium, magnesium, potassium, and manganese contents in xylem sap of beech (Fagus sylvatica L.) in a 35-year-old limestone beech forest stand. Trees 4:75--80.
Hagemeyer, J. 1993. Monitoring trace metal pollution with tree rings: a critical reassessment. In Plants as Biomonitors. Ed. B. Markert. VCH, Weinheim, New York, pp 541--563.
Hagemeyer, J. and K. Lohrie. 1995. Distribution of Cd and Zn in annual xylem rings of young spruce trees (Picea abies (L.) Karst.) grown in contaminated soil. Trees 9:195--199.
Hagemeyer, J. and K. Shin. 1995. Extraction of Cd and Pb from stem wood of pine (Pinus sylvestris L.) trees with various solutions. Angew. Bot. 69:55--59.
Momoshima, N. and E.A. Bondietti. 1990. Cation binding in wood: applications to understanding historical changes in divalent cation availability to red spruce. Can. J. For. Res. 20:1840--1849. Momoshima, N. and E.A. Bondietti. 1994. The radial distribution of
90Sr and 137Cs in trees. J. Environ. Radioact. 22:93--109.
Rennenberg, H., R. Schupp, V. Glavac and H. Jochheim. 1994. Xylem sap composition of beech (Fagus sylvatica L.) trees: seasonal changes in the axial distribution of sulfur compounds. Tree Physiol. 14:541--548.
Sachs, L. 1992. Angewandte Statistik, 7th Edn. Springer-Verlag, Ber-lin, pp 410--413.
Sauter, J.J. 1981. Seasonal variation of amino acids and amides in the xylem sap of Salix. Z. Pflanzenphysiol. 101:399--411.
Schneider, A., J. Kreuzwieser, R. Schupp, J.J. Sauter and H. Rennen-berg. 1994. Thiol and amino acid composition of the xylem sap of poplar trees (Populus × canadensis ‘robusta’). Can. J. Bot. 72:347--351.
Stewart, C.M. 1966. Excretion and heartwood formation in living trees. Science 153:1068--1074.
Van Bel, A.J.E. 1990. Xylem--phloem exchange via the rays: The undervalued route of transport. J. Exp. Bot. 41:631--644.
Ziegler, H. 1964. Storage, mobilization and distribution of reserve material in trees. In The Formation of Wood in Forest Trees. Ed. M.H. Zimmermann. Academic Press, New York, pp 303--320. Ziegler, H. 1968. Biologische Aspekte der Kernholzbildung. Holz
Roh- Werkstoff 26:61--68.