MENGGUNAKAN ANALISIS KERAGAMAN
FENOTIPIK, DAERAH
D-LOOP
DNA MITOKONDRIA
DAN DNA MIKROSATELIT
MOHD. AGUS NASHRI ABDULLAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Saya menyatakan dengan sebenar-benarnya bahwa disertasi saya dengan judul Karakterisasi Genetik Sapi Aceh Menggunakan Analisis Keragaman Fenotipik, Daerah D-Loop DNA Mitokondria dan DNA Mikrosatelit adalah benar-benar asli karya saya dengan arahan komisi pembimbing, dan bukan jiplakan atau tiruan dari tulisan siapapun serta belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun.
Bogor, 10 Maret 2008
MOHD. AGUS NASHRI ABDULLAH. Genetics Characterization of Aceh Cattle Utilizing Phenotypic, Mitochondrial DNA of D-loop Region and Microsatellite DNA Analyses. Under the supervision of HARIMURTI MARTOJO, RONNY RACHMAN NOOR, and DEDY DURYADI SOLIHIN
The aims of this study were to describe the variation of body size measurement, D-loop of mtDNA and microsatellite alleles of Aceh cattle. The body measurement data were collected from 131 males and 269 females of Aceh cattle. The whole blood samples (8 samples for D-loop analyses and 160 samples for microsatellite genotyping) were collected from Aceh Besar, Pidie, North Aceh district and Banda Aceh city. For the out-group comparison, the whole blood samples were collected from 10 Bali cattle, two samples of each collected from Madura, PO and Pesisir cattle. The D-loop sequences of mtDNA amplification were done by using BIDLF and BIDLR primer with the PCR product of 980 bp. Sixteen markers were used for genotyping microsatellite DNA. Body size measurement data variation were analyzed using Minitab 14.13 software. The molecular data were analyzed using Minitab 14.13, Squint 1.02, Mega4 and Arlequin 3.11 and Excel software.
The result shows that the body weight and size of Aceh cattle were smaller than those of Bali, Madura and PO cattle. However the body size of Aceh cattle was larger when compared to that of Pesisir cattle. Most of Aceh cattle have red sand and light brown coat color. The horn shape of female cattle was bend slightly toward left or right and than bend to forward direction. The male has the same horn shape as females but at the tips of the horn goes upward. Most of Aceh cattle have concave face line and some of them have diametrical face line. The result of the D-loop mtDNA analyses showed that there were 27 site variation of D-loop mtDNA Aceh cattle. These specific markers can be used to differentiate and subdivide Indonesian domestic cattle. The Aceh cattle were in the same cluster with Pesisir cattle. However, the PO cattle were in a closer cluster with Bos indicus cattle, while Bali and Madura cattle were in the same cluster. The result of microsatellite analyses showed that the averages allele number per locus was 10,25 ± 2,07. The percentage of the heterozygosity of Aceh cattle was higher than those of Bali, but lower compared to those of Madura, PO and Pesisir cattle. Based on the microsatellite alleles analyses, the Aceh cattle were in the same cluster with PO cattle and were in the same branch of the phylogeny tree with Pesisir and Madura cattle.
MOHD. AGUS NASHRI ABDULLAH. Karakterisasi Genetik Sapi Aceh Menggunakan Analisis Keragaman Fenotipik, Daerah D-loop DNA Mitokondria dan DNA Mikrosatelit. Dibimbing oleh HARIMURTI MARTOJO, RONNY RACHMAN NOOR, dan DEDY DURYADI SOLIHIN
Sapi Aceh merupakan satu dari empat bangsa sapi asli Indonesia (Aceh, Bali, Madura, Pesisir). Sapi Sumba-Ongole (SO) dan Java-Ongole (PO) juga dianggap sebagai bangsa sapi lokal Indonesia. Ternak-ternak asli telah terbukti dapat beradaptasi dengan lingkungan lokal termasuk makanan, ketersediaan air, iklim dan penyakit. Dengan demikian, ternak-ternak inilah yang paling cocok untuk dipelihara dan dikembangkan di Indonesia, walaupun produksinya lebih rendah dari ternak impor.
Sapi Aceh diduga dimasukkan oleh pedagang-pedagang India yang membawa sapi-sapi dari India ke Aceh pada masa lampau dengan tujuan berdagang dan menguasai perekonomian di Aceh. Selanjutnya sapi ini diduga mengalami persilangan dengan banteng liar yang ada di Sumatera, namun belum pernah diverifikasi dan diungkapkan melalui analisis genom.
Eksploitasi sapi Aceh melalui persilangan yang semakin luas dengan bangsa sapi eksotik yang dilakukan selama ini dapat mengancam keberadaan sapi Aceh pada masa yang akan datang. Kepunahan yang dapat terjadi pada sapi Aceh yang telah teradaptasi lingkungan akan sulit bahkan tidak akan dapat digantikan. Hal ini akan berdampak pada kehidupan sosial, ekonomi, budaya masyarakat Aceh yang tidak bisa terlepas dari beternak dan mengkonsumsi daging sapi.
Oleh karena itu, dilakukan penelitian pada sapi Aceh yang mencakup inventarisasi sumber daya genetiknya melalui analisis fenotipik, DNA mitokondria pada daerah D-loop dan DNA mikrosatelit. Tujuan penelitian ini adalah melakukan karakterisasi terhadap keragaman fenotipik dan keragaman genetik, daerah D-loop mtDNA dan DNA mikrosatelit yang berguna sebagai database
dalam pelaksanaan program pelestarian plasma nutfah sapi Aceh,
pengembangan dan pemanfaatannya secara berkelanjutan.
Pengumpulan data fenotipik sapi Aceh (131 jantan dan 269 betina) dan sampel darah (8 sampel untuk analisisD-loop dan 160 sampel untuk genotiping mikrosatelit) dilakukan di Kota Banda Aceh, Kabupaten Aceh Besar, Pidie dan Aceh Utara. Sampel pembanding telah diambil 10 sampel sapi Bali (Pulau Bali), dua sampel masing-masing sapi Madura (Pulau Madura), PO (Jawa Barat), Pesisir (Sumatera Barat). Ekstraksi dan purifikasi DNA total telah dilakukan menurut metode Sambrook yang dikembangkan Duryadi.Primeryang digunakan untuk mengamplifikasi fragmen daerah D-loop mtDNA adalah pasangan primer
BIDLF dan BIDLR dengan panjang produk 980 bp. Bagian DNA mikrosatelit, telah digunakan adalah enambelas lokus (BM1818, INRA005, CSRM60, BM2113, HEL5, HEL9, HEL13, INRA63, INRA35, HEL1, ETH225, ETH10, CSSM66, BM1824, ILSTS006 dan ILSTS005) untuk genotiping mikrosatelit. Data fenotipik dianalisis secara deskriptif dengan menggunakan program Minitab
dalam paket MEGA. Data ukuran-ukuran alel mikrosatelit dianalisis dengan program Arlequin 3.11 dan dukungan Minitab 14.13 serta Excel. Jarak genetik yang diperoleh digunakan untuk membentuk pohon filogeni dengan program
Phylip (phylogeny Inference Package) versi 3.67.
Hasil penelitian menunjukkan bahwa ditinjau dari segi bobot badan dan ukuran tubuh, sapi Aceh mengalami penurunan bobot badan dan ukuran-ukuran tubuh dibanding hasil laporan tahun 1926. Apabila dibandingkan sapi Bali, Madura dan PO, maka sapi Aceh mempunyai ukuran-ukuran tubuh yang lebih kecil pada tingkat umur yang sama, namun berada di atas rataan sapi Pesisir. Secara kualitatif, sapi Aceh berwarna dominan merah bata dan cokelat muda serta pola warna beragam mulai warna gelap sampai terang. Bentuk pertumbuhan tanduk sapi betina mengarah ke samping melengkung ke atas kemudian ke depan dan pada jantan mengarah ke samping melengkung ke atas. Pada umumnya sapi Aceh mempunyai garis punggung yang cekung (89,25%), sebagian mempunyai garis punggung cembung (6,25%) dan sebagian kecil mempunyai garis punggung lurus (4,5%). Hasil analisis D-loop mtDNA dengan sekuen acuanBos indicus dari GenBank, ada 27 situs beragam pada sapi Aceh. Penanda ini dapat digunakan untuk pembeda dan pengelompokan sapi lokal Indonesia yaitu sapi Aceh satu klaster dengan sapi Pesisir dan PO, mempunyai jarak genetik yang lebih dekat dengan sapiBos indicus, sedangkan sapi Bali dan Madura membentuk klaster sendiri. Pengelompokan sapi Aceh dengan sapi Pesisir dan PO dalam klaster sapi Bos indicus menunjukkan bahwa sapi Aceh, Pesisir dan PO adalah dari maternal zebu, sedangkan pengelompokan sapi Madura dalam klaster sapi Bali (Bos javanicus) menunjukkan bahwa sapi Madura bukan dari maternal zebu tetapi dari maternal banteng. Hasil analisis DNA mikrosatelit, diperoleh rataan alel per lokus 10,25 ± 2,07. Sapi Aceh memiliki derajat heterozigositas yang tinggi dan berbeda genetik dengan sapi Bali, Madura, Pesisir dan PO. Urutan kedekatan genetik antara sapi Aceh dengan sapi pembanding berturut-turut yaitu: sapi PO, Pesisir, Madura dan Bali, dengan pohon filogeni yang menunjukkan sapi Aceh memiliki klaster yang sama dengan sapi PO, membentuk cabang dengan sapi Pesisir dan sapi Madura.
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
MENGGUNAKAN ANALISIS KERAGAMAN
FENOTIPIK, DAERAH
D-LOOP
DNA MITOKONDRIA
DAN DNA MIKROSATELIT
MOHD. AGUS NASHRI ABDULLAH
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Ilmu Ternak
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Terbuka : 1. Dr. Ir. Achmad Machmud Thohari, DEA
DNA Mikrosatelit
Nama : Mohd. Agus Nashri Abdullah
NRP : D061030081
Program Studi : Ilmu Ternak
Disetujui,
Komisi Pembimbing
Prof. Dr. Harimurti Martojo, M.Sc Ketua
Prof. Dr. Ir. Ronny Rachman Noor, MRur.Sc Dr. Ir. Dedy Duryadi Solihin, DEA
Anggota Anggota
Diketahui,
Ketua Departemen Ilmu Produksi Dekan Sekolah Pascasarjana dan Teknologi Peternakan
Dr. Ir. Cece Sumantri, M.Agr.Sc Prof. Dr. Ir. Khairil Anwar Notodiputro, MS
Sujud syukur Alhamdulillah dan segala puji penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Sasaran yang dipilih dalam penelitian yang dilaksanakan sejak Pebruari 2005
sampai dengan Juni 2007 adalah keragaman genetik, yang berjudul
Karakterisasi Genetik Sapi Aceh Menggunakan Analisis Keragaman Fenotipik, DaerahD-loopDNA Mitokondria dan DNA Mikrosatelit.
Gabungan tiga bagian utama penelitian (fenotipik, daerah D-loop DNA mitokondria, dan DNA mikrosatelit) pada sapi Aceh dengan cakupan daerah pengambilan sampel yang luas dan mengetahui kedudukan sapi Aceh dalam pengelompokan sapi lokal lainnya di Indonesia belum pernah dilakukan sebelumnya. Penelitian ini memberikan informasi yang sangat penting sebagai
database sapi Aceh dan ini merupakan yang pertama dilakukan di Provinsi
Nanggroe Aceh Darussalam.
Pengamatan penulis diawali ketika masa kecil sekitar awal tahun 80-an pada beberapa ekor sapi Aceh milik seorang peternak Desa Tanjong Seulamat di depan rumah orang tua penulis di Kampus Universitas Syiah Kuala. Sapi tersebut sering digembalakan berhari-hari tanpa dipindahkan dengan cara diikat di lapangan rumput siang dan malam beserta panas dan hujan, hingga di sekitarnya tidak terlihat lagi rumput bahkan akarnya, namun sapi tersebut tetap tegar. Banyak sapi milik peternak lain di sekitar kampus digembalakan dengan cara dilepas siang dan malam, sehingga sering bergerombol masuk ke lapangan kampus untuk merumput. Keadaan demikian telah berlangsung sejak awal kampus berdiri di tahun 60-an hingga sekarang kadang masih juga terjadi. Jika dilakukan perjalanan darat di malam hari dari Banda Aceh menuju perbatasan dengan Sumatera Utara, maka akan ditemui sapi beristirahat sepanjang jalan negara lintas Sumatera, sehingga ada sebutan '“Aceh memiliki kandang sapi terpanjang di dunia”. Keadaan ini berlangsung hingga diberlakukan Operasi Jaring Merah (DOM) di Aceh karena konflik berkepanjangan. Betapa besar daya tahan hidup sapi Aceh dengan kondisi demikian.
Gagasan penelitian ini muncul setelah melihat kekhawatiran pada sapi Aceh yang ada di Provinsi Nanggroe Aceh Darussalam (NAD). Sejak ada pelaksanaan program Inseminasi Buatan (IB) dan semakin meluas persilangan yang dilakukan pada sapi Aceh di provinsi ini, peternak semakin sulit mendapatkan sapi Aceh murni sehingga kemungkinan inbreeding telah terjadi. Peternak di desa yang umumnya mempunyai tingkat pendidikan sangat rendah (tamat dan tidak tamat SD), hampir seluruhnya tertarik memelihara sapi yang lebih besar setelah melihat hasil-hasil persilangan yang telah dilakukan dengan bangsa sapi impor. Bahkan di Aceh, sebagian orang peternakan sendiri menyepelekan sapi Aceh dengan beberapa alasan seperti sapi kecil (bahasa
Acehleumó bukriėk), lambat dewasa, tidak efisien dan perlu diganti dengan sapi
lain, sehingga sapi ini semakin terancam keberadaannya. Padahal dari laporan Merkens tahun 1926 dan beberapa keterangan dari sesepuh pendahulu di Kabupaten Aceh Besar, Pidie dan Aceh Utara menjelaskan bahwa, sapi Aceh dahulu berukuran besar-besar dan tidak kecil seperti sekarang. Perayaan pesta perkawinan besar hanya cukup dipotong satu ekor sapi Aceh.
Pembimbing yang telah mendukung, meluangkan waktu, pikiran dan tenaganya dalam membimbing penulis sejak dalam perkuliahan, penulisan proposal sampai selesai penulisan karya ilmiah ini. Semoga Guru-guru penulis diberikan pahala oleh Allah SWT.
Penulis mengucapkan terima kasih kepada Rektor Universitas Syiah Kuala atas izin dan dukungannya. Terima kasih disampaikan kepada Dikti atas dukungan dana pendidikan BPPs. Ucapan terima kasih penulis ucapkan kepada Kepala Dispet TK. I Provinsi NAD dan Dispet TK. II Kota Banda Aceh, Kabupaten Aceh Besar, Pidie dan Aceh Utara beserta stafnya atas bantuan selama koleksi sampel di Aceh. Penulis mengucapkan terima kasih kepada Ilham, S.Si atas bantuan selama koleksi sampel darah sapi di Pulau Madura dan terima kasih kepada Dr. Jakaria, S.Pt, M.Si atas pemberian sampel darah sapi Bali dari P3Bali. Penulis mengucapkan terima kasih kepada Dr. Ir. Sarbaini Anwar, M.Sc atas pemberian sampel darah sapi Pesisir dari Sumatera Barat dan juga terima kasih kepada Prof. Dr. Eddie Gurnadi, M.Sc atas keizinan pengambilan sampel darah sapi PO di Laboratorium Ilmu Ternak Daging dan Kerja Fapet IPB. Penulis mengucapkan terima kasih kepada Kepala Laboratorium Biologi Molekuler Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) Institut Pertanian Bogor Dr. Ir. Dedy Duryadi Solihin, DEA, atas segala fasilitas alat dan bahan yang dapat penulis gunakan mulai dari isolasi, ekstraksi DNA total sampai pelaksanaan amplifikasi PCR daerah D-loop DNA Mitokondria. Ucapan terima kasih penulis sampaikan kepada Prof. Heriberto Rodriguez-Martinez, Ph.D atas dukungan dana penelitian DNA mikrosatelit dan terima kasih kepada Prof. Dr. Göran Andersson, Ph.D selaku Kepala Laboratorium Molekuler dan Genetika di Swedish University of Agricultural Sciences (SLU), Uppsala, Swedia atas segala fasilitas alat dan bahan yang dapat penulis gunakan selama penelitian DNA Mikrosatelit berlangsung, serta terima kasih disampaikan kepada Mia Ollson (mahasiswa program Ph.D di SLU) atas bantuannya selama penulis melakukan analisis DNA Mikrosatelit. Ucapan terima kasih penulis sampaik juga kepada seluruh teman dan kolega.
Akhirnya, ungkapan terima kasih penulis sampaikan kepada Drs. Abdullah Rayeuk, M.Si (Ayah) dan Ibu Salwiyah Abdul Wahab, isteri tercinta Sofia Kurnia, S.Ag serta ananda Mohd Anshar Anashri atas dukungan, pengertian dan do’anya.
Semoga karya ilmiah ini dapat bermanfaat bagi perkembangan kemajuan ilmu peternakan di Indonesia dan khususnya Nanggroe Aceh Darussalam serta pembaca.
Bogor, 10 Maret 2008
Penulis dilahirkan di Kopelma Darussalam pada tanggal 16 Agustus 1971 sebagai anak ketiga dari empat bersaudara dari pasangan Drs. Abdullah Rayeuk, M.Si dan Salwiyah Abdul Wahab. Pendidikan Sekolah Dasar Negeri No. 82 (SD Teladan Lamnyong) Banda Aceh ditamatkan pada tahun 1984, kemudian dilanjutkan pada SMPN 13 tamat tahun 1987 dan SMAN 6 tamat tahun 1990 yang keduanya berada di Kota Pelajar Mahasiswa (Kopelma) Universitas Syiah Kuala Darussalam Banda Aceh. Pendidikan sarjana dimulai pada tahun 1990 pada Program Studi Produksi Ternak Jurusan Peternakan Fakultas Pertanian Universitas Syiah Kuala, lulus pada tahun 1995. Kesempatan untuk melanjutkan ke program magister pada Program Studi Ilmu Ternak, Sekolah Pascasarjana IPB diperoleh pada tahun 2000 dan lulus 2003 dengan beasiswa pendidikan dari BPPs Dikti Jakarta. Pada tahun 2003 kembali penulis mendapat kesempatan melanjutkan pendidikan (Doktor) pada program studi yang sama Institut Pertanian Bogor dengan beasiswa dari BPPs Dikti Jakarta. Pada tahun 2005, penulis memperoleh dana penelitian dari Riset Unggulan Terpadu XII tahun I sebagai Ketua Peneliti. Selanjutnya, dalam tahun 2007, penulis memperoleh kesempatan melanjutkan penelitian ini pada Laboratorium Molekuler dan Genetika, Husdjursgenetik, University of Agricultural Sciences (SLU), Uppsala, Swedia atas kerja sama Pembimbing Anggota Prof. Dr. Ir. Ronny Rachman Noor, MRur.Sc dengan peneliti di SLU Swedia Prof. Heriberto Rodriguez-Martinez, Ph.D.
DAFTAR ISI
Halaman
DAFTAR TABEL... xiv
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xix
PENDAHULUAN... 1
Latar Belakang ... 1
Tujuan Penelitian... 5
Manfaat Penelitian ... 5
Hipotesis ... 5
TINJAUAN PUSTAKA ... 6
Gambaran Umum Nanggroe Aceh Darussalam ... 6
Keragaman Genetik Ternak... 6
Pelestarian Sumber Daya Genetik Ternak ... 9
Sifat Kuantitatif dan Kualitatif ... 11
Sumber Daya Genetik Ternak Lokal Indonesia ... 12
Karakteristik Sapi Aceh... 14
Penanda Molekuler ... 15
DNA Mitokondria ... 18
DNA Mikrosatelit... 20
Teknik Penelitian DNA... 23
MATERI DAN METODE ... 26
Penelitian Lapangan ... 26
Waktu dan Tempat Penelitian... 26
Materi Penelitian... 25
Peralatan yang Digunakan... 27
Pengambilan Sampel Data Kuantitatif dan Kualitatif ... 27
Analisis Data... 28
Penelitian Laboratorium... 30
DaerahD-loopDNA Mitokondria... 30
Waktu dan Tempat Penelitian... 30
Pelaksanaan Pengambilan Sampel Darah... 31
Bahan-bahan dan Peralatan ... 31
PerancanganPrimerDaerahD-loopDNA Mitokondria ... 32
Isolasi dan Purifikasi DNA Total ... 32
Elektroforesis untuk Visualisasi DNA Hasil Isolasi ... 33
Amplifikasi DaerahD-loopDNA Mitokondria ... 34
Penentuan Sekuen Nukleotida ... 34
Analisis Data... 35
DNA Mikrosatelit... 36
Waktu dan Tempat Penelitian... 37
Pelaksanaan Pengambilan Sampel Darah... 37
PrimerMikrosatelit ... 37
Amplifikasi Lokus Mikrosatelit ... 38
Elektroforesis Produk PCR ... 38
Analisis Data... 39
HASIL DAN PEMBAHASAN ... 42
Penelitian Lapang... 42
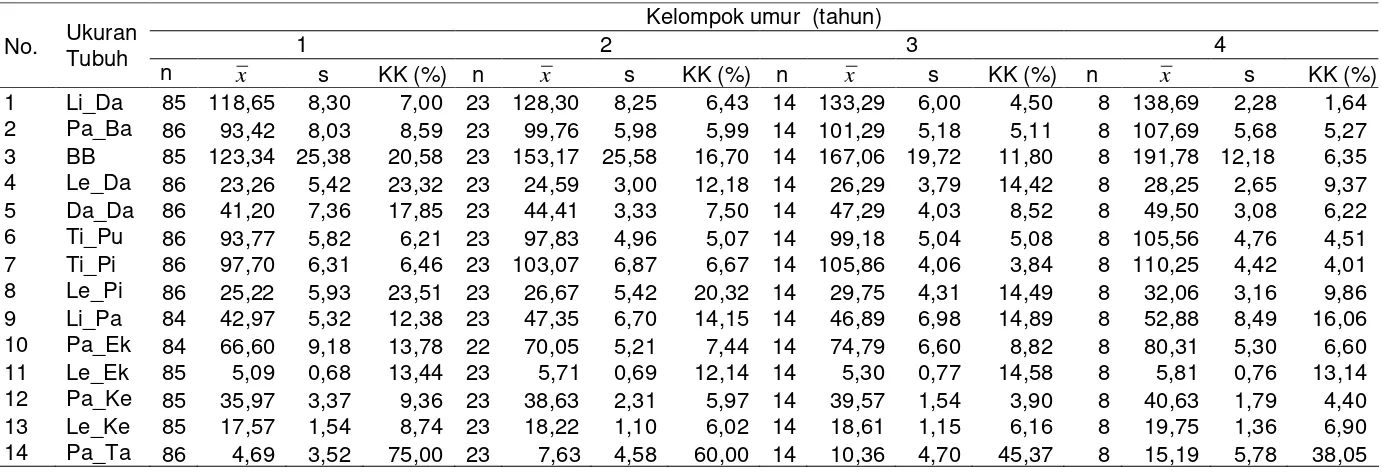
Profil Peternak ... 42
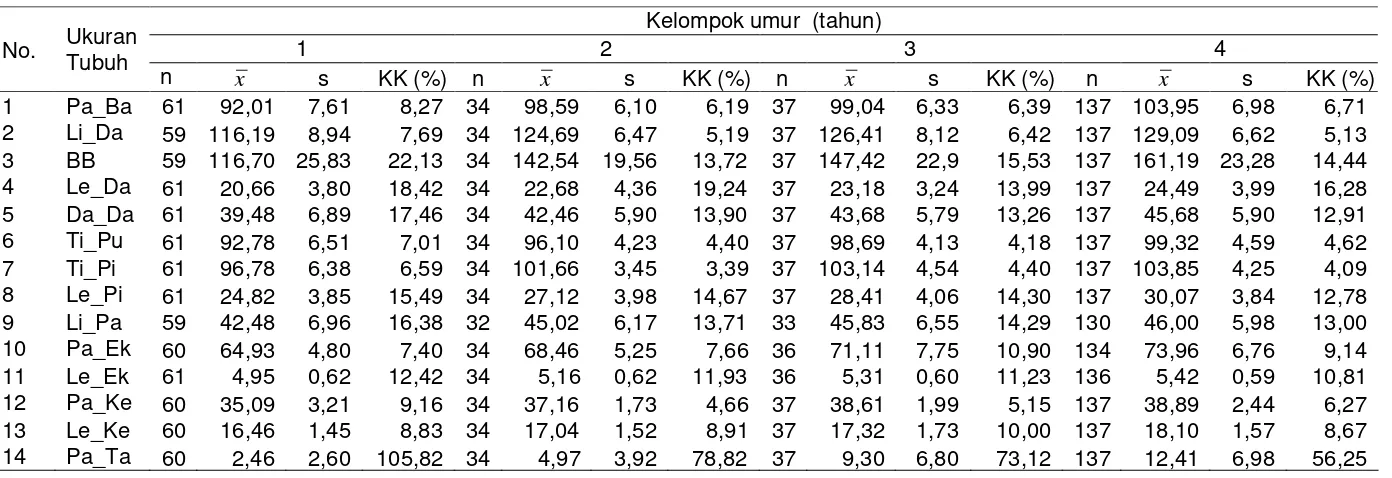
Ukuran-ukuran Tubuh ... 45
Bentuk Tubuh ... 50
Warna dan Pola Warna Tubuh... 51
Bentuk Tanduk... 55
Penelitian Laboratorium... 57
DaerahD-loopDNA Mitokondria... 57
DNA Total ... 57
Amplifikasi daerahD-loop... 57
Penentuan daerahD-loopParsial dan Keragaman Runutan Nukleotida... 58
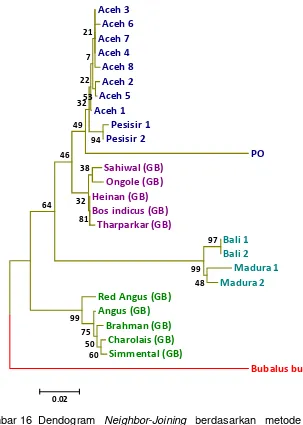
Jarak Genetik sapi Aceh dan Sapi Pembanding ... 63
Hubungan Kekerabatan Sapi Aceh... 64
DNA Mikrosatelit... 70
DNA Total ... 70
Amplifikasi Mikrosatelit... 71
Alel dan Lokus Polimorfik... 72
Distribusi dan Jumlah Genotipe ... 76
Distribusi Frekuensi Alel ... 77
Variasi Genetik dan Keseimbangan Hardy-Weinberg ... 93
Jarak Genetik Sapi Aceh dan SapiOutgroup... 96
Penelusuran Asal-usul Sapi Aceh... 98
Pembahasan Umum ... 104
SIMPULAN DAN SARAN... 111
Simpulan ... 111
Saran... 112
DAFTAR PUSTAKA... 113
DAFTAR TABEL
Halaman
1 Urutan basa dan suhu penempelanprimer untuk mengamplifikasi
daerahD-loopsapi penelitian ... 32
2 Profil peternak sapi Aceh... 43
3 Ukuran-ukuran tubuh dan bobot badan sapi Aceh jantan dengan
menggunakan rumus lingkar dada dan panjang badan ... 46
4 Ukuran-ukuran tubuh dan bobot badan sapi Aceh betina dengan
menggunakan rumus lingkar dada dan panjang badan ... 47
5 Ukuran-ukuran tubuh dan bobot badan sapi-sapi jantan lokal pada
umur berbeda... 50
6 Warna-warna tubuh sapi Aceh... 53
7 Frekuensi bentuk-bentuk pertumbuhan tanduk sapi Aceh ... 55
8 Jumlah insersi dan delesi basa-basa nukleotida pada sapi Aceh, Bali,
Madura, PO dan Pesisir dengan acuan Bos indicus... 59
9 Rataan komposisi nukleotida daerahD-loopparsial sapi Aceh, Bali, Madura, PO dan Pesisir setelah disejajarkan dengan komposisi nukleotida acuanBos indicusdariGenBank
(ukuran 479 bp) ... 61
10 Perbedaan susunan basa nukleotida sapi Aceh, Bali, Madura, PO,
Pesisir dan Bos indicusdariGenBank... 62
11 Jarak genetik berdasarkan metode 2 parameter Kimura pada sapi
Aceh, Bali, Madura, PO, Pesisir dan Bos indicusdariGenBank... 64
12 Panjang produk PCR dan selisih ukuran maksimum dan minimum
pada masing-masing lokus mikrosatelit sapi penelitian ... 71
13 Jumlah alel masing-masing lokus mikrosatelit pada sapi Aceh,
Bali, Madura, PO, dan Pesisir... 73
14 Perbandingan rataan jumlah alel per lokus pada berbagai ternak
ternak penelitian ... 74
15 Alel-alel pada sepuluh lokus mikrosatelit yang hanya ditemukan
pada sapi Bali, Madura dan Pesisir... 75
16 Kisaran ukuran alel dan jumlah genotipe sapi Aceh dan sapi
17 Heterozigositas masing-masing lokus mikrosatelit pada sapi Aceh
Bali, Madura, PO dan Pesisir... 94
18 Matriks jarak genetik Nei yang diperoleh dari frekuensi-frekuensi alel pada 16 lokus mikrosatelit sapi Aceh, Bali, Madura, PO dan
Pesisir... 96
19 Estimasi proporsi sumber gen-gen dariBos taurus,Bos indicus
danBos javanicus... 98
20 Perbedaanrange ukuran alel sapi Aceh dan sapioutgroup terhadap
DAFTAR GAMBAR
Halaman
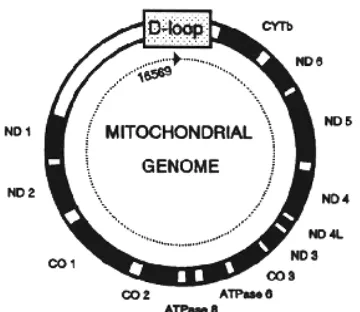
1 Skema genom mitokondria... 19
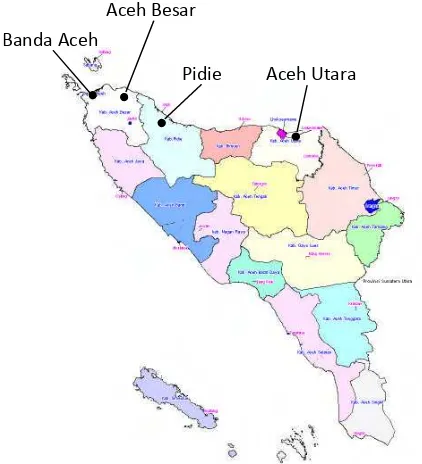
2 Lokasi pengambilan sampel data fenotipik sapi Aceh ... 26
3 Sketsa bagian-bagian permukaan tubuh sapi Aceh yang diukur ... 28
4 Lokasi pengambilan sampel darah sapi Aceh dan sapi outgroup
untuk analisis daerahD-loopDNA mitokondria ... 30
5 Lokasi pengambilan sampel darah sapi Aceh dan sapi outgroup
untuk analisis DNA mikrosatelit ... 36
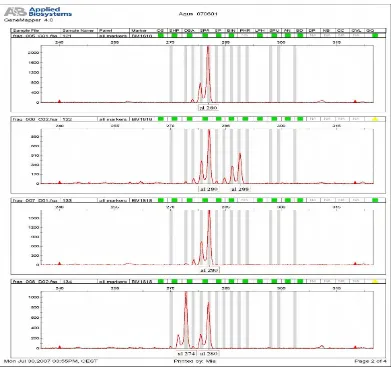
6 Sinyal fluoresen yang dihasilkan mesin ABI Prism 3100 DNA analyzer (Applied Biosystems) yang menunjukkan hasil amplifikasi
DNA mikrosatelit dengan menggunakan marker BM1818 ... 40
7 Perbedaan ukuran-ukuran tubuh sapi Aceh pada tahun berbeda.... 49
8 Warna-warna tubuh sapi Aceh ... 52
9 Sketsa bentuk-bentuk pertumbuhan tanduk sapi Aceh... 56
10 Spektrofotometer DNA total sapi penelitian setelah dimigrasikan
dalam gel agarose 1,2% pada tegangan 90 volt selama 30 menit... 57
11 Sketsa letak penempelanprimerBIDLF dan BIDLR untuk
mengamplifikasi fragmen daerahD-loopsapi Aceh, Bali, Madura,
PO dan Pesisir... 58
12 Hasil amplifikasi daerahD-loopdengan menggunakan pasangan
primerBIDLF dan BIDLR setelah dimigrasikan dalam gel agarose
1,2% pada tegangan 90 volt selama 45 menit ... 58
13 Sketsa daerahD-loopparsial hasil perunutan DNA (berukuran 479 bp) yang dipakai untuk analisis keragaman genetik pada
sapi Aceh... 59
14 Hasil sekuensing daerahD-loopparsial DNA mitokondria sapi Aceh
yang dianalisis ... 60
15 Frekuensi nukleotida daerahD-loopparsial berukuran 479 nt pada sapi Aceh, Bali, Madura, PO, Pesisir dan Bos indicus(Nellore) dari
16 DendogramNeighbor-Joining Neighbor-Joiningberdasarkan metode 2 parameter Kimura dari nukleotida daerahD-loopparsial (berukuran 479 nt) sapi Aceh, Bali, Madura, PO, Pesisir dan bangsa-bangsa sapi dariGenBankdengan pengolahanbootstrap
1000 ulangan... 66
17 Tampilan DNA total sapi penelitian setelah dimigrasikan dalam gel
agarose 1,2% pada tegangan 90 volt selama 30 menit ... 70
18 Perbandingan jumlah genotipe yang hanya terdapat pada sapi
Aceh, outgroupdan genotipe bersama... 77
19 Distribusi frekuensi alel lokus BM1818 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 78
20 Distribusi frekuensi alel lokus INRA005 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 79
21 Distribusi frekuensi alel lokus CSRM60 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 80
22 Distribusi frekuensi alel lokus BM2113 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 81
23 Distribusi frekuensi alel lokus HEL5 pada sapi Aceh, Bali, Madura,
PO dan Pesisir... 81
24 Distribusi frekuensi alel lokus HEL9 pada sapi Aceh, Bali, Madura,
PO dan Pesisir... 82
25 Distribusi frekuensi alel lokus HEL13 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 83
26 Distribusi frekuensi alel lokus INRA63 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 84
27 Distribusi frekuensi alel lokus INRA35 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 85
28 Distribusi frekuensi alel lokus HEL1 pada sapi Aceh, Bali, Madura,
PO dan Pesisir... 87
29 Distribusi frekuensi alel lokus ETH225 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 88
30 Distribusi frekuensi alel lokus ETH10 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 89
31 Distribusi frekuensi alel lokus CSSM66 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 89
32 Distribusi frekuensi alel lokus BM1824 pada sapi Aceh, Bali,
33 Distribusi frekuensi alel lokus ILSTS006 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 91
34 Distribusi frekuensi alel lokus ILSTS005 pada sapi Aceh, Bali,
Madura, PO dan Pesisir... 92
35 Kontruksi pohon filogeni berdasarkan metodeNeighbor-Joining
dari data jarak genetik Nei pada sapi Aceh, Bali, Madura, PO dan
Pesisir... 97
36 Spesies liar dan domestikasi antarsubfamiliBovinae... 100
37 DendogramNeighbor-Joiningberdasarkan metode 2 parameter Kimura dari nukleotida gencytochrome-bparsial (berukuran 420 nt)
Sapi dariGenBankdengan pengolahan bootstrap 1000 ulangan... 106
38 Kontruksi pohon filogeni berdasarkan metodeNeighbor-Joining
dari data jarak genetik Nei pada sapi Aceh, Bali, Madura, PO dan
DAFTAR LAMPIRAN
Halaman
1 Lokasi pengambilan sampel sapi Aceh di Nanggroe Aceh
Darussalam untuk analisis fenotipik ... 124
2 Lokasi pengambilan sampel darah sapi Aceh, Bali, Madura, PO
dan Pesisir untuk analisis daerahD-loopDNA mitokondria... 125
3 Lokasi pengambilan sampel darah dan nomor sapi di Provinsi
Nanggroe Aceh Darussalam untuk analisis DNA mikrosatelit ... 126
4 Lokasi pengambilan sampel darah sapi pembanding untuk analisis
DNA mikrosatelit... 128
5 Komposisi bahan pereaksi yang digunakan untuk isolasi DNA dari
sampel darah ... 129
6 Konsentrasi sampel DNA total hasil pemeriksaan dengan mesin
NanoDrop Spectrophotometer ... 130
7 Lokasi penempelan primerBIDL-F dan BIDL-R pada sekuen basa
nukleotida daerah D-loopsapiBos indicus... 132
8 Pensejajaran berganda nukleotida dari daerahD-loopparsial sapi
Aceh, Bali, Madura, PO, Pesisir dan bangsa ternak dariGenBank . 133
9 Situs-situs delesi dan insersi basa-basa nukleotida daerah D-loop
parsial sapi Aceh, Bali, Madura, PO dan Pesisir ... 140
10 Jumlah Nukleotida sapi Aceh, Bali, Madura, PO, dan Pesisir
setelah disejajarkan dengan nukleotidaBos indicus(GenBank) ... 147
11 Komposisi Nukleotida sapi Aceh dan sapi pembanding ... 148
12 Jarak Genetik sapi Aceh dengan sapi Bali, Madura, PO, Pesisir
dan bangsa-bangsa sapiBos indicus, Bos taurus dariGenBank .... 149
13 Hasilblastsekuen sapi Aceh (476 nt) pada situs NCBI... 150
14 Nomor akses sekuen daerahD-looputuhBos indicus, Bos taurus
danBubalus bubalis dariGenBankpada situs NCBI yang
digunakan untuk membentuk pohon filogeni ... 157
15 Sekuens nukleotidaprimermikrosatelit yang digunakan untuk
penelitian ... 152
16 Runutan indeks Garza-Williamson hasil uji heterozigositas hitung
(observed) dan heterozigositas harapan (expected) pada sapi
PENDAHULUAN
Latar Belakang
Sapi asli Indonesia secara genetik dan fenotipik umumnya merupakan: (1)
turunan dari Banteng (Bos javanicus) yang telah didomestikasi dan dapat pula (2)
berasal dari hasil silangan sapi asli Indonesia dengan sapi eksotik yang
kemudian mengalami domestikasi serta adaptasi lokal. Kelompok sapi yang
termasuk dalam kategori pertama adalah sapi Bali karena sapi Bali diketahui
merupakan hasil domestikasi langsung dari Banteng (MacHugh 1996; Martojo
2003; Hardjosubroto 2004) dan mempunyai ciri-ciri fisik yang hanya mengalami
perubahan kecil dibandingkan dengan moyangnya (Handiwirawan dan
Subandriyo 2004). Kelompok sapi yang kedua adalah sapi Madura karena
menurut Payne dan Rollinson (1976); Nijmanet al. (2003); Verkaar et al. (2003)
merupakan hasil silangan Banteng atau sapi Bali dengan sapi zebu yang telah
berlangsung kurang lebih 1.500 tahun yang lalu, walaupun hal tersebut tidak
terdokumentasi dengan baik secara prinsip pemuliaan (tanpa recording yang
jelas). Kedua pengelompokan itu secara genetik telah terbukti dengan
menggunakan marker genetik DNA mitokondria (D-loop dan cytochrome-b) dan
DNA inti (Mikrosatelit dan AFLP) (Nijmanet al. 2003). Kemungkinan yang ketiga
adalah sapi eksotik yang telah menetap di Indonesia dalam kurun waktu lama
(impor dan perdagangan) dan dapat berkembang biak dengan baik (mampu
beradaptasi pada lingkungan setempat), selanjutnya sapi-sapi tersebut
mengalami persilangan. Salah satu yang termasuk dalam kelompok ini adalah
sapi Ongole India yang masuk ke Pulau Sumba pada tahun 1905 yang kemudian
menjadi sapi Sumba Ongole (SO). Pada tahun 1915 sampai 1929 sapi Sumba
Ongole (SO) mulai disebarkan ke Pulau Jawa melalui program “Ongolisasi”
dengan sebutan “kontrak Sumba” (Hardjosubroto 2004). Dampak dari program
ini adalah terbentuknya sapi Peranakan Ongole (PO) dan hasil silangan lainnya,
bahkan program ini telah mempunyai kontribusi yang jelas terhadap hilangnya
sapi Jawa. Menurut Merkens (1926) di Jawa terdapat sapi Jawa dengan
karakteristik tertentu yang merupakan campuran berbagai bangsa sapi.
Sapi Aceh pada mulanya diduga dimasukkan oleh pedagang India pada
masa kerajaan Islam pertama di Peureulak yang terbentuk tahun 847 M (225 H),
karena pada masa itu sudah terjalin hubungan kerja sama antarnegara dan
pedagang-pedagang dari Mesir, Parsi, dan Gujarat (catatan sejarah Aceh,
catatan Marcopolo 1256 dan Ibnu Bathutah 1345; Mulyana 1968; Putra 2001).
Hal ini telah dijelaskan pula oleh Merkens (1926) bahwa, perdagangan yang
ramai sudah lama terjalin antara Aceh dengan Malaka. Pedagang Arab, Cina
serta India yang datang ke Aceh, mereka membawa barang-barang dagangan
dan khususnya imigran India ini sudah dikenal membawa sapi-sapi dari India ke
Aceh. Pada abad ke-19 telah menjadi kebiasaan mengimpor ternak melalui
Selat Malaka, khususnya ke Pidie dan Aceh Timur Laut (Peureulak).
Kemungkinan sapi-sapi di Aceh mengalami persilangan dengan Banteng
yang ada di Sumatera seperti dikemukakan Merkens (1926) dari hasil kumpulan
catatan, foto dan laporan singkatnya, namun belum pernah diverifikasi dan
diungkapkan melalui analisis genom. Beberapa sapi tersebut berkembang dan
menyebar ke pesisir barat Aceh hingga ke wilayah pantai Sumatera Barat.
Keadaan wilayah pesisir barat tersebut memiliki keadaan pakan terbatas dan
kualitas nutrisi rendah sehingga telah turut menyeleksi ragam sapi yang hidup di
daerah ini yaitu kebanyakan sapi berukuran kecil (±150 kg) yang dapat bertahan
hidup dengan baik (ILRI 1995). Disamping itu di daerah pesisir barat ini jauh dari
hewan buas pemangsa. Di daerah Aceh yang lain seperti Banda Aceh, Aceh
Besar, Pidie, Aceh Utara, dan Aceh Timur terdapat sapi-sapi yang beragam
ukurannya. Menurut Gunawan (1998), ada sapi Aceh di Aceh Besar dan Aceh
Utara yang hanya mempunyai bobot hidup dewasa 150 kg, namun ada pula sapi
Aceh yang ada di daerah ini mencapai bobot hidup dewasa 400 kg atau lebih.
Sapi ini mempunyai daya tahan terhadap lingkungan yang buruk dan sistem
pemeliharaan ekstensif tradisional. Laporan Merkens (1926), menyebutkan
bahwa kepala sapi Aceh berwarna antara cokelat merah sampai cokelat
abu-abu, bahkan di Aceh Utara dan Aceh Timur ditemukan sapi yang warna
kepalanya lebih gelap sampai hitam. Ciri tersebut merupakan salah satu karakter
dari sapi India. Namikawa et al. (1982a) menambahkan bahwa, sapi Sumatera
(Aceh dan Pesisir) memiliki macam-macam warna yaitu hitam, cokelat
kehitaman, cokelat kuning, dan abu-abu putih yang didominasi oleh warna
cokelat kuning. Dibandingkan dengan warna sapi Bali (Banteng), menurut Payne
dan Rollinson (1973); NRC (1983), sapi Bali terdapat warna putih pada bagian
belakang paha (pantat), bagian bawah (perut), keempat kaki bawah (white
stocking) sampai di atas kuku, bagian dalam telinga, dan pada pinggiran bibir
dengan garis hitam tipis terdapat di sepanjang tengah punggung dan jantan
berwarna cokelat. Pada saat umur 12-18 bulan, warna sapi Bali jantan berubah
menjadi agak gelap sampai mendekati hitam pada saat dewasa. Tipe warna sapi
Bali (Banteng) yang demikian diduga juga ada kemiripan dengan pola warna
yang dimiliki sapi Aceh, namun hal ini belum pernah dilaporkan.
Sapi asli Indonesia telah mengalami seleksi alam dengan berbagai tekanan
iklim tropis basah dan ketersediaan pakan yang sesuai daerah di mana sapi-sapi
tersebut hidup. Dengan demikian dalam jangka waktu yang cukup lama telah
terjadi interaksi genetik dan lingkungan sehingga menghasilkan sapi-sapi Aceh
yang telah teradaptasi terhadap wilayah tersebut, dengan penampilan
fenotipiknya beragam. Ketahanan ternak lokal terhadap lingkungan yang ekstrim
telah diuji melalui hewan percobaan mencit (Mus musculus) oleh Abdullah et al.
(2005) bahwa, mencit liar yang telah teradaptasi lingkungan dengan segala
perubahan yang ada mempunyai gen pengatur daya produksi dan reproduksi
yang lebih unggul terhadap stres lingkungan dibanding mencit laboratorium.
Pengujian tersebut mendukung pendapat Noor (2008) bahwa, ternak-ternak asli
telah terbukti dapat beradaptasi dengan lingkungan lokal termasuk makanan,
ketersediaan air, iklim dan penyakit. Dengan demikian, ternak-ternak inilah yang
paling cocok untuk dipelihara dan dikembangkan di Indonesia, walaupun
produksinya lebih rendah dari ternak impor, tetapi pengelolaannya lebih efisien.
Eksploitasi sapi Aceh melalui persilangan yang semakin luas dan tidak
terkontrol dengan bangsa sapi eksotik akan memberikan dampak yang kurang
baik terhadap sapi-sapi Aceh yang telah teradaptasi pada lingkungan setempat.
Kekhawatiran ini telah terjadi pada sapi asli di Lithuania (Eropa Timur) yang
terancam punah (Malevičiūtė et al. 2002) akibat persilangan yang disengaja
tetapi tidak terstruktur. Bahkan beberapa sapi asli di negara India telah punah
sebelum sapi ini diidentifikasi dan dimanfaatkan akibat persilangan yang meluas
dan tidak terkontrol (Sodhi et al. 2006). Hal demikian ini juga ditegaskan oleh
FAO (2000) bahwa, sumber daya genetik ternak asli akan cenderung punah
akibat permintaan pasar yang baru (eksploitasi besar-besaran), persilangan yang
tidak terkendali, pergantian breed (penggantian bangsa sapi yang sudah ada
dengan bangsa sapi baru) dan kegiatan mekanisasi pertanian (penggantian
penggunaan tenaga sapi dengan tenaga mesin untuk mengolah lahan
Sehubungan dengan hal tersebut di negara berkembang, banyak peneliti
sedang melakukan karakterisasi ternak asli/lokal secara fenotipik dan juga pada
tingkat molekuler untuk digunakan dalam dokumentasi plasma nutfah yang ada
serta prospek pemanfaatannya di masa yang akan datang (Sodhi et al. 2006).
Karakter fenotipe ternak dapat menunjukkan ciri khas bangsa ternak tertentu.
Ternak sapi merupakan hewan peliharaan sangat penting di Aceh secara
turun-temurun sampai sekarang. Ancaman kepunahan sapi Aceh akibat
persilangan yang tidak terkendali akan berdampak sangat luas bagi kehidupan
sosial ekonomi masyarakat Aceh. Kehilangan sapi Aceh yang telah eksis selama
ratusan tahun akan mengurangi pemenuhan kebutuhan protein hewani dan
penyediaan daging meugang (hari adat pemotongan dan makan daging
bersama) serta hewan kurban dalam perayaan keagamaan di Aceh, sehingga
Aceh akan bergantung pada distribusi daging dari daerah lain atau impor.
Disamping itu, hal ini akan mematikan perekonomian peternak yang merangkap
petani dan merupakan bagian terbesar dari mata pencaharian penduduk Aceh.
Lemahnya perekonomian peternak di Aceh akan menimbulkan gejolak sosial dan
akan berakibat pada naiknya tingkat kemiskinan. Selain itu, FAO sebagai badan
dunia sudah menganjurkan bahwa sedapat mungkin sumber daya genetik ternak
lokal harus dipertahankan.
Oleh karena itu, perlu dilakukan penelitian pada sapi Aceh mencakup
inventarisasi sumber daya genetiknya melalui analisis fenotipik, DNA mitokondria
pada daerah D-loop dan DNA mikrosatelit. Metode ini dapat digunakan karena
tingkat akurasi sangat tinggi dalam menggambarkan keragaman genetik
sehingga hasil penelitian ini dapat digunakan sebagai dasar pengambilan
kebijakan guna menerapkan keputusan yang lebih tepat dan terarah dalam
program pelestarian plasma nutfah sapi Aceh, pengembangan dan
pemanfaatannya secara berkelanjutan.
Berdasarkan laporan terdahulu, penentuan daerah D-loop mtDNA pada
sapi dapat menunjukkan sejarah sapi (Nijman et al. 2003; Edwards et al. 2007)
dan hibridisasi yang terjadi pada Banteng dan sapi Madura (Nijman et al. 2003).
DNA mitokondria terutama daerah D-loop, sangat baik digunakan untuk analisis
keragaman hewan, baik intraspesies maupun antarspesies (Muladno 2006).
DNA mitokondria sudah terbukti suatu alat yang tangguh dalam analisis variasi
dalam dan antarspesies, struktur populasi dan filogeni (Patricia et al. 2002).
molekuler yang digunakan untuk mengkarakterisasi genetik, mikrosatelit
merupakan penanda yang paling disukai. Hal ini karena penanda tersebut
bersifat polimorfik dan sangat informatif, kelimpahannya di dalam genom inti
relatif besar, dan dapat diamplifikasi melalui PCR. Penanda ini telah digunakan
untuk menjelaskan pola migrasi dan domestikasi pada sapi eropa (Loftus et al.
1994; Bruford et al. 2003) dan untuk karakterisasi populasi-populasi ternak sapi
dari turunan Bos indicus danBos taurus (Moore et al. 1992; Beja-Pereira et al.
2003). Machado et al. (2003) menggunakan lokus-lokus mikrosatelit untuk
mengevaluasi keanekaragaman genetik dalam masing-masing bangsa sapi dan
perbedaan genetik di antara setiap bangsa. Penanda genetik mikrosatelit dapat
memberikan informasi-informasi penting sehingga dapat dibuat keputusan
mengenai konservasi pada ternak sapi (Sunnucks 2000; Sodhiet al. 2006).
Tujuan Penelitian
1. Menginventarisasi sifat-sifat fenotipe kualitatif (warna dan pola warna, bentuk
tanduk, garis muka dan punggung) dan kuantitatif (ukuran-ukuran tubuh dan
bobot badan) sapi Aceh sebagai ciri-ciri sapi lokal.
2. Mengkaji keragaman genetik daerahD-loopDNA mitokondria pada sapi Aceh
untuk mengetahui asal-usulnya.
3. Mengkaji keragaman DNA mikrosatelit populasi sapi Aceh dan asal-usulnya.
Manfaat Penelitian
1. Dapat memberi informasi keragaman fenotipik dan genetik sapi Aceh dalam
pengelompokan sapi lokal di Indonesia.
2. Karakteristik sumber daya genetik sapi Aceh sebagai pedoman dalam
menerapkan kebijakan dalam program pelestarian plasma nutfah,
pengembangan dan pemanfaatannya secara berkelanjutan.
Hipotesis
1. Penanda genetik daerah D-loop DNA mitokondria dapat mengelompokkan
sapi Aceh terhadap sapi lokal Indonesia dan sapi luar Indonesia.
2. Alel-alel mikrosatelit sapi Aceh bersifat polimorfik.
3. Asal-usul sapi Aceh adalah dari sapi-sapi india (Bos indicus) yang mengalami
TINJAUAN PUSTAKA
Gambaran Umum Nanggroe Aceh Darussalam
Provinsi Nanggroe Aceh Darussalam terletak di bagian paling barat dari
gugusan kepulauan Nusantara. Ibukota Provinsi Nanggroe Aceh Darussalam
adalah Banda Aceh, mempunyai luas daratan 2,75% terhadap luas daratan
Indonesia yaitu 5.193.700 ha dengan 17 kabupaten, 227 kecamatan dan 5.947
desa (Departemen Kehutanan 2004).
Secara geografis Provinsi Nanggroe Aceh Darussalam terletak di antara
2-6oLU dan 95-98 oBT, termasuk 119 pulau. Rataan tinggi daratan 125 m di
atas permukaan laut (Sujitno dan Achmad 1995). Suhu udara maksimum 32,4oC
pada bulan Nopember dan minimum 24,2oC pada bulan Juni, curah hujan
maksimum 243,4 mm pada bulan Januari dan curah hujan minimum 22,2 mm
pada bulan Juni (Departemen Kehutanan 2004). Udara di Aceh mempunyai
kelembaban tinggi terutama di wilayah pesisir barat sangat lembab dan basah,
sedangkan di wilayah timur Aceh mempunyai udara kering. Batas di sebelah
barat adalah Samudera Indonesia, dan di sebelah utara dan timur adalah Selat
Malaka, sedangkan di sebelah utara mengikuti sungai Simpang Kiri di sebelah
timur dan sungai Tamiang di sebelah barat bagian selatan (Sujitno dan Achmad
1995; PEMDA NAD1997). Jumlah penduduk 4.240.000 jiwa (estimasi Juni 2003)
dengan laju pertumbuhan 2,57% dan kepadatan penduduk 81/km2(Departemen
Kehutanan 2004).
Sebagian besar penduduk Aceh hidup dari pertanian (peternakan), ladang
padi, palawija dan hortikultura (Sujitno dan Achmad 1995). Daerah Aceh
(Nanggroe Aceh Darussalam) memiliki kekayaan ternak, terutama ternak sapi.
Keadaan ini didukung oleh adanya padang gembalaan yang dapat menampung
ratusan ribu sapi merumput bebas dan pada saat tertentu dipanen (Gunawan
1998). Pada tahun 2002 terdapat 701.356 ekor sapi di Nanggroe Aceh
Darussalam (Direktorat Jenderal Bina Produksi Peternakan 2003).
Keragaman Genetik Ternak
Keragaman genetik terjadi tidak hanya antarbangsa tetapi juga di dalam
satu bangsa yang sama, antarpopulasi maupun di dalam populasi, atau di antara
individu dalam populasi. Pada spesies domestik suatu identifikasi tingkat
mempunyai keterkaitan dengan seleksi dalam program pemuliaan (Handiwirawan
dan Subandriyo 2004). Salah satu ’alat bantu’ yang dapat digunakan untuk
mendeteksi keragaman populasi adalah DNA mitokondria dan DNA mikrosatelit
(Muladno 2006).
Keragaman genetik dalam populasi merupakan modal dasar aplikasi
teknologi pemuliaan dalam pemanfaatan hewan. Keragaman genetik populasi
yang digambarkan dalam keragaman penampilan hewan adalah refleksi
informasi genetik yang dimilikinya. Sebagai ilustrasi sapi Bali yang hidup di pulau
Bali memiliki kontruksi gen-gen yang berbeda dengan populasi sapi pesisir yang
ada di Sumatera Barat. Perbedaan-perbedaan tersebut dapat dinyatakan dalam
kemampuan adaptasi, besarnya tubuh, dan ketahanan penyakit. Komponen ini
sangat berpengaruh terhadap kemampuan menyesuaikan diri (adaptasi)
terhadap perubahan lingkungan seperti degradasi kualitas lingkungan sebagai
media tumbuh hewan. Perbedaan penampilan ini disebabkan selama
domestifikasi, tipe-tipe atau bangsa-bangsa hewan terpisah secara genetik
karena adanya penyesuaian (adaptasi) dengan masing-masing lingkungan lokal
dan kebutuhan komunitas lokal sehingga dihasilkan bangsa berbeda (Muladno
2006). Adanya kemampuan adaptasi hewan disebabkan hewan memiliki
kemampuan menghasilkan lebih dari satu alternatif bentuk morfologi, status
fisiologi, dan atau tingkah laku sebagai reaksi terhadap perubahan lingkungan
(pengaturan ekspresi gen) (Noor 2008).
Lebih dari 12.000 tahun yang lalu terdapat 14 spesies ternak telah
didomestikasikan dan berevolusi sehingga menjadi rumpun (breed) yang secara
genetika unik dan berbeda, beradaptasi terhadap lingkungan dan komunitas
setempat. Saat ini terdapat sekitar 6.000-7.000 rumpun ternak domestik dari
spesies yang telah terdomestikasi, bersama dengan lebih dari 80 spesies kerabat
liarnya yang merupakan sumber daya genetik ternak di bumi ini yang berperanan
penting untuk pangan dan produksi pertanian (Subandriyo dan Setiadi 2003).
Beja-Pereiraet al. (2006) menyatakan bahwa, penjinakan sapi (Bos taurus
danBos indicus) dariaurochsen liar (Bos primigenius) adalah satu yang penting
dalam sejarah manusia, mendorong ke arah modifikasi-modifikasi pangan yang
meluas, tingkah laku dan struktur sosial ekonomi dari banyak populasi: (1)
proses ini dimulai sekitar 11.000 tahun yang lalu; (2) penyimpangan genetik yang
besar antara bangsa-bangsa sapi taurine (Bos taurus) dan zebu (Bos indicus),
merupakan kejadian dari dua kelompokaurochsen yang jelas terpisah; (3). data
arkeologis menyatakan bahwa penjinakan sapi zebu terjadi mungkin di Indus
Valley (sekarang Pakistan); (4) dengan suatu difusi utama bangsa-bangsa sapi
ini di India dan hanya yang terakhir (<3.000 tahun) pengenalan sebagai jantan
sekunder di Afrika; dan (5) sebaliknya, lokasi penjinakan yang hampir bisa
dipastikan untuk bangsa-bangsa taurine dipertimbangkan yaitu di suatu areal
paling barat di Near East, Fertile Crescent (FC), meskipun suatu peristiwa
penjinakan yang berdiri sendiri mungkin telah terjadi di Afrika.
Berbagai bangsa ternak yang telah berkembang dalam berbagai sistem
dan lingkungan yang ada saat ini telah menghasilkan berbagai kombinasi gen
yang unik. Gen-gen ini tidak hanya menentukan kualitas sifat produksi dari
masing-masing bangsa, tetapi juga terhadap kemampuan adaptasinya pada
kondisi lokal termasuk makanan, ketersediaan air, iklim dan hama penyakit (FAO
2001).
Berbagai macam kebutuhan manusia sehari-hari dipenuhi dari spesies
ternak, dalam bentuk pangan maupun kebutuhan lainnya. Namun hanya
sebagian kecil dari total keragaman genetik ternak dan kerabat liarnya, yakni
sekitar 40 spesies yang memenuhi sebagian besar proporsi dari produksi ternak
global. Keragaman genetik di dalam spesies ternak dan beberapa kerabat
liarnya telah menjadi sumber keragaman dari rumpun dan populasi ternak.
Keragaman genetik ini penting dalam pembentukan rumpun ternak modern dan
akan terus berkelanjutan di masa mendatang (Subandriyo dan Setiadi 2003).
Sebagai contoh, negara Lithuania memiliki dua bangsa sapi utama yaitu
Lithuanian Black dan White dan Lithuanian Red, masing-masing 62% dan 32%
dari populasi ternak dalam negeri. Bangsa ternak Lithuanian Black dan White
ditemukan di Barat Daya Lithuania dan dikembangkan dari sapi lokal Lithuania
berwarna putih dan hitam melalui persilangan sesama dengan bangsa sapi hitam
dan putih Holstein-Friesian dari Belanda dan bangsa sapi hitam dan putih dari
Jerman. Bangsa sapi Lithuanian Black dan Black sebagai bangsa sapi perah
yang menghasilkan susu berkualitas dan telah teradaptasi dengan iklim di
tempatnya. Bangsa sapiLithuanian Red ditemukan di Timur Laut Lithuania dan
dibentuk pada awal abad ini dari bangsaLithuanian Redlokal melalui assortative
mating, seleksi masal dan persilangan dengan bangsa-bangsa: Ayrshire, Angeln,
Dutch, Danish Red, Swiss Brown dan Shorthorn. Bangsa-bangsa sapi unggul
bangsa sapi perah (FAO 2000). Di Barat Daya, bagian Tenggara dan sebagian
Lithuania bagian Tengah, sapi Lithuania paling asli yaitu memiliki warna tubuh
putih atau abu-abu terang. Dalam beberapa dekade terakhir, sapi Lithuania yang
asli, seperti Lithuanian Light Grey dan Lithuanian White, hampir seluruhnya
digantikan olehLithuanian BlackdanWhitedanLithuanian Red yang merupakan
sapi produksi tinggi. Hal ini diketahui bahwa sapi Lithuanian Light Grey dan
Lithuanian White-Backed dipelihara di Lithuania mulai zaman lampau dan
dikhususkan untuk negara. Sekarang ini, Lithuanian White-Backed, Lithuanian
Grey dan bangsa sapi Lithuanian Black danWhite tua populasinya sangat kecil
dan mempunyai status dari suatu bangsa sapi yang terancam kepunahan
(Malevičiūtė et al. 2002).
Keanekaragaman genetik ternak, sedikitnya memiliki empat manfaat, yaitu
(1) keberlanjutan dan peningkatan produksi pangan; (2) memaksimumkan
produktivitas lahan dan sumber daya pertanian; (3) pencapaian pertanian
berkelanjutan untuk memberikan keuntungan masa kini dan generasi yang akan
datang; (4) pemenuhan keanekaragaman baik yang telah maupun yang belum
diketahui manfaatnya bagi kehidupan sosial masyarakat. Ketersediaan
keanekaragaman genetik ternak, termasuk sapi akan mempengaruhi
keberhasilan strategi pemuliaan untuk masa yang akan datang (FAO-AAAS
1994).
Pelestarian Sumber Daya Genetik Ternak
FAO memprediksi bahwa paling sedikit satu bangsa ternak tradisional
punah setiap minggu dan lebih dari 30% ternak di Eropa sekarang ini
diperkirakan dalam keadaan terancam kepunahan (FAO 1995). Banyak bangsa
ternak tradisional sudah menghilang karena para petani lebih fokus pada bangsa
sapi baru. Sekitar 16% dari bangsa sapi tradisional telah punah dan kurang dari
15 % bersifat jarang (FAO 2000). Keadaan ini dapat dilihat pada bangsa-bangsa
sapi zebu di India yang secara signifikan telah kehilangan ekonomi yang sangat
penting dan penurunan ukuran populasi terutama karena persilangan secara
meluas (Sodhiet al. 2006).
Semakin berkembangnya ilmu pengetahuan dan teknologi pemuliaan
ternak, bioteknologi, permintaan pasar, mekanisasi pertanian dan produksi
ternak, akan mendorong eksploitasi ternak melalui persilangan, penggantian
pengurasan stock secara berlebihan, dan pada gilirannya akan mengancam
keragaman genetik ternak. Di lain pihak pelestarian keragaman genetik ternak
akan selalu diperlukan dalam pemuliaan di masa mendatang, karena tanpa
adanya keragaman genetik, pemuliaan ternak tidak mungkin dilaksanakan untuk
mengantisipasi keperluan di masa mendatang (Subandriyo dan Setiadi 2003)
Melalui literatur diketahui bahwa sudah sejak lama, ternak asli mempunyai
suatu arti penting yang sangat tinggi di dalam kegiatan pertanian dan kehidupan
sosial dari masyarakat pedesaan. Diketahui pula bahwa bangsa-bangsa sapi
lokal yang dipelihara petani di dalam areal pertanian menjadi titik rujukan dari
tradisi-tradisi lokal pada masa lalu, yang berhubungan dengan produksi pertanian
dan sektor-sektor lain, seperti makanan (Malevičiūtėet al. 2002).
Pelestarian terhadap sumber daya genetik ternak lokal sebagai bagian dari
komponen keanekaragaman hayati adalah penting untuk memenuhi kebutuhan
pagan, pertanian dan perkembangan sosial masyarakat di masa yang akan
datang. Ada beberapa alasan untuk ini, antara lain (1) lebih dari 60 p ersen dari
bangsa-bangsa ternak di dunia berada di negara-negara sedang berkembang;
(2) konservasi bangsa ternak lokal tidak menarik bagi petani; (3) secara umum
tidak ada program monitoring yang sistematis dan tidak tersedianya informasi
deskriptif dasar sebagian besar sumber daya genetik hewan ternak; serta (4)
sedikit sekali bangsa-bangsa ternak asli yang telah digunakan dan
dikembangkan secara aktif (FAO 2001).
Kesadaran dari pentingnya memelihara sumber daya genetik hewan sudah
meningkat dalam beberapa tahun terakhir. Banyak negara Eropa mempunyai
atau sedang mengembangkan program-program nasional untuk pemeliharaan
dan konservasi keanekaragaman hayati hewan (Malevičiūtė et al. 2002).
Ada tiga metode utama program pelestarian plasma nutfah ternak yang
telah dilaksanakan masyarakat atau pemulia: (1) mempertahankan populasi
ternak hidup, (2) penyimpanan beku materi genetik berupa haploid (n) seperti
gamet yakni semen dan oocyteatau berupa diploid (2n) seperti embrio, dan (3)
penyimpanan DNA (deoxyrybonucleic acid). Metode bioteknologi dapat
digunakan untuk mengkarakterisasi gen-gen ternak dan plasma nutfah suatu
populasi. Metode ini akan membantu dalam pembuatan keputusan tentang
pelestarian plasma nutfah yang unik. Studi mengenai struktur dan fungsi
gen-gen pada tingkat molekuler suatu populasi ternak dapat membantu menentukan
keragaman genetik dalam populasi ternak yang diamati. Identifikasi gen-gen dari
individu ternak akan membantu program pemuliaan (genetik) ternak, yang
membedakan dari penampilan (fenotipe) yang tampak, yang dapat menentukan
proses pemilihan tetua untuk generasi yang akan datang (seleksi buatan). Jika
gen-gen untuk sifat produksi dapat diidentifikasi, ternak-ternak tersebut dapat
diseleksi walaupun tidak diekspresikan oleh individu ternak yang bersangkutan.
Sebagai alternatif, jika mereka dapat diikatkan dengan gen-gen yang diketahui
lokasinya dalam kromosom (marker lokus-lokusi), seleksi dapat dilaksanakan
berdasarkan acuan tersebut (Subandriyo dan Setiadi 2003).
Negara Indonesia mempunyai undang-undang dan peraturan menteri
pertanian menyangkut pelestarian sumber daya genetik ternak yaitu
Undang-Undang Republik Indonesia Nomor 5 tahun 1990 tentang Konservasi Sumber
Daya Alam Hayati dan Ekosistemnya dan Undang-Undang Republik Indonesia
Nomor 5 tahun 1994 tentang Pengesahan Konvensi Perserikatan
Bangsa-Bangsa Mengenai Keanekaragaman Hayati serta Peraturan Menteri Pertanian
Nomor 35/Permentan/OT.140/8/2006 tentang Pedoman Pelestarian dan
Pemanfaatan Sumberdaya Genetik Ternak dan Peraturan Menteri Pertanian
Nomor 36/Permentan/OT.140/8/2006 tentang Sistem Pembibitan Ternak
Nasional.
Sifat Kuantitatif dan Kualitatif
Setiap sifat yang diekspresikan seekor hewan disebut fenotipe. Seekor
hewan atau ternak menunjukkan fenotipenya (P) sebagai hasil
pengaruh-pengaruh seluruh gen atau genotipenya (G), lingkungan (E) dan interaksi antara
genotipe dan lingkungan (IGE) (Martojo 1992; Hardjosubroto 1994). Sifat
kuantitatif dan kualitatif pada hewan atau ternak merupakan fenotipe.
Pada program pemuliaan, prediksi perbedaan genetik di antara hewan
dapat berdasarkan observasi fenotipe yang bergantung pada faktor genetik dan
lingkungan (Muladno 2006). Fenotipe ternak dapat diketahui melalui
ukuran-ukuran tubuh (Otsukaet al. 1980; Surjoatmodjo 1993; Karthickeyanet al. 2006),
warna dan pola warna tubuh, pertumbuhan tanduk (Wiley 1981; Warwick et al.
1990; Handiwirawan 2003; Riwantoro 2005), tekstur dan panjang rambut
(Handiwirawan dan Subandriyo 2004).
Otsuka et al. (1980) telah menggunakan ukuran-ukuran tubuh hewan
serta hubungannya dengan berbagai bangsa sapi lain di Asia. Warna termasuk
sifat kualitatif seekor ternak (Warwicket al. 1990). Warna tubuh ternak dianggap
sebagaicharacter displacementuntuk membedakan satu bangsa dengan bangsa
lainnya (Baker dan Manwell 1991).
Sifat kuantitatif adalah ciri-ciri dari makhluk hidup yang dapat diukur,
dihitung atau diskors. Karakter ini ditentukan oleh banyak pasang gen (poligenik)
dan sangat dipengaruhi oleh lingkungan (Wiley 1981), sedangkan sifat kualitatif
seperti warna, pola warna, sifat bertanduk atau tidak bertanduk dapat dibedakan
tanpa harus mengukurnya. Sifat kualitatif biasanya hanya dikontrol oleh
sepasang gen (Noor 2008).
Membedakan warna tubuh merupakan salah satu cara mengidentifikasi
ternak. Pelacakan pada sapi asli yang hampir punah di Lithuania untuk upaya
konservasi, salah satunya dilakukan melalui ciri-ciri umum pada breed asli
Lithuanian Black danWhite danLithuanian Red yaitu berdasarkan warna tubuh
yang khas, konstitusi tubuh, perlawanan tubuh sapi terhadap penyakit-penyakit
lokal, dan produksi umum sebagai sapi tipe perah dan sapi dwifungsi sebagai
penghasil susu dan daging (Malevičiūtė et al. 2002).
Sifat-sifat kualitatif adalah sifat-sifat yang pada umumnya dijelaskan
dengan kata-kata atau gambar (Warwick et al. 1990). Spesies-spesies sering
ditandai oleh warna atau pola warna tertentu. Pola warna apabila ada
kemungkinan lebih berguna daripada warna itu sendiri. Hal ini biasanya dianggap
sebagaicharacter displacementuntuk menghindari kesalahan saat membedakan
bangsa ternak (Wiley 1981).
Sumber Daya Genetik Ternak Lokal Indonesia
Sapi Bali merupakan satu dari empat bangsa sapi asli Indonesia (Aceh,
Pesisir, Madura dan Bali). Sapi Sumba-Ongole dan Java-Ongole (PO) juga
dianggap sebagai bangsa sapi lokal Indonesia (Martojo 2003; Dahlanuddinet al.
2003).
Ternak sapi sebagaimana jenis ternak lain, dikenali sebagai
komponen-komponen penting dari keanekaragaman hayati dunia, karena gen dan
kombinasi gen-gen yang terdapat pada ternak ini berguna bagi pertanian di masa
yang akan datang (Beja-Pereiraet al. 2003). Sumber daya genetik ternak adalah
populasi pada masing-masing spesies yang secara genetik unik, terbentuk dalam
termasuk kerabat dekat populasi tersebut yang masih liar. Istilah ini juga
digunakan semua spesies dan bangsa-bangsa hewan yang mempunyai nilai
ekonomis, ilmu pengetahuan dan sosial budaya untuk pertanian, atau
mempunyai kepentingan untuk masa depan (FAO-AAAS 1994).
Bangsa-bangsa ternak tradisional dan asli yang multifungsi telah
teradaptasi dengan kondisi-kondisi lokal, iklim, penyakit-penyakit dan nutrisi
lingkungan sepanjang tahun. Bangsa-bangsa ternak seperti itu juga dengan baik
telah beradaptasi dengan sumber pakan lokal, atau lebih resistan terhadap
patogen-patogen dan berbagai penyakit ternak yang ada di daerah di mana
ternak itu berada. Bangsa-bangsa ternak asli telah juga diseleksi dengan
berbagai tujuan, tergantung pada sifat-sifatnya yang penting bagi
masyarakat-masyarakat lokal. Bangsa ternak asli merupakan bangsa ternak yang dapat
menunjukkan hasil dari pengembangan sosial ekonomi masyarakat dan tidak mungkin bertahan di luar sistem pertanian yang telah terbentuk (Malevičiūtėet al. 2002).
Sumber daya ternak sapi di Indonesia saat ini terdiri atas tiga kelompok,
yakni (1) ternak asli; (2) ternak impor; dan (3) ternak yang telah beradaptasi.
Sehubungan pentingnya nilai konservasi pada kelompok ternak ini, beberapa
bangsa sapi menjadi target konservasi sekaligus pemanfaatannya (Utoyo 2002.
Beberapa di antara sumber daya ternak sapi tersebut ialah sapi Aceh, Bali,
turunan Ongole, Sumba Ongole, Madura, Jawa, Pesisir, dan Grati. Selanjutnya
Utoyo (2002) menjelaskan keanekaragaman sapi di Indonesia, terbentuk dari
sumber daya genetik asli dan impor. Importasi Bos indicus Ongole dari India
telah dimulai pada awal abad ke-20 dan kemudian bangsa sapi ini memegang
peranan penting dalam program pengembangan peternakan di Indonesia. Sapi
Ongole murni pertama dibawa ke pulau Sumba yang kemudian disebut sebagai
Sumba Ongole, kemudian sapi ini dibawa ke tepat-tempat lain untuk disilangkan
dengan sapi asli Jawa dan membentuk Peranakan Ongole (Ongole-cross) dan
sapi Madura.
Pada tahun 1976 terdapat 6.194.000 ekor sapi di Indonesia, umumnya
sapi-sapi tersebut ada di Pulau Jawa, Madura, Bali dan Nusa Tenggara dan
sekitar 650.00 ekor ada di Sumatera. Bangsa-bangsa sapi utama atau sapi lokal
Indonesia adalah Peranakan Ongole (Filial Ongole), Bali, Madura, Aceh dan
sapi terbesar. Sapi Bali, Madura dan Aceh diketahui secara luas sebagai jenis
sapi lokal khas atau bangsa ternak asli (Otsukaet al. 1980).
Menurut ILRI (1995), terdapat banyak ternak asli di Indonesia: tujuh bangsa
sapi asli (Sumba Ongole, Ongole cross, Bali, Madura, Aceh, Pesisir dan Grati);
kerbau (Sungai, Murrah, Toraja dan Kalang); domba (Domba Ekor Gemuk dan
Domba Ekor Kurus); kambing (Kacang, Etawah cross, Gembrong dan Kosta);
ayam (Kampung, Kedu, Pelung, Bekisar dan Nunukan); itik (Tegal, Modjosari,
Alabio dan Bali); serta babi (Java, Bali dan Nias).
Hubungan genetik antara sapi-sapi Indonesia sangat menarik karena
variasi sapi-sapi tersebut yang luas (Namikawaet al. 1982b). Ada sejumlah gen
pool ternak asli Indonesia yang telah beradaptasi dengan lingkungannya dan
belum secara penuh dimanfaatkan dalam kaitan dengan laju pertumbuhan atau
reproduksi. Bangsa-bangsa ternak asli Indonesia dipelihara secara sistem
tradisional dengan input yang rendah. Perusahaan ternak komersial tidak
mempercayai bangsa-bangsa ternak asli tersebut karena hal ini diperhitungkan
bahwa ternak asli mempunyai produktivitas yang rendah. Lebih lanjut bahwa
tidak ada jaminan tentang perolehan suatu penyediaan ternak dalam jumlah
besar secara reguler. Namun, di Indonesia telah timbul suatu keinginan akan
keuntungan dari keunggulan genetik bangsa-bangsa ternak asli (ILRI 1995).
Karakteristik Sapi Aceh
Sapi Aceh merupakan sapi lokal yang terdapat di Aceh Sumatera (Merkens
1926; Namikawa et al. 1982a) dan diminati sebagai ternak potong (Merkens
1926). Sapi tersebut masih terdapat beberapa variasi warna tubuh (Namikawaet
al. 1982a). Sapi kecil yang banyak ditemukan di bagian barat, lebih mendekati
jenis Sumatera biasa (sapi Pesisir). Sapi yang lebih baik dan lebih besar yang
diminati sebagai komoditas ekspor ke kawasan budaya Deli dan Medan, berasal
dari Aceh. Sebagian sapi Aceh digunakan sebagai alat transportasi pada
beberapa perusahaan berlokasi dekat rel kereta di Deli dan Medan karena lebih
baik dan lebih besar (Merkens 1926).
Sapi Aceh mempunyai daya tahan terhadap lingkungan yang buruk seperti
krisis pakan, air dan pakan berserat tinggi, penyakit parasit, temperatur panas
dan sistem pemeliharaan ekstensif tradisional (Gunawan 1998). Berdasarkan
bahwa sapi Aceh merupakan hasil persilangan Banteng dan zebu (Merkens
1926).
Pada tahun 1926, di Aceh terdapat sekitar 150 ribu ekor sapi. Sejauh ini
tidak banyak literatur mengenai jenis sapi ini yang berperanan penting di seluruh
kawasan Sumatera Utara. Sapi Aceh jantan mempunyai tinggi pundak 115,5 cm;
tinggi pinggul 115,0 cm; panjang badan 126,0 cm; lebar dada 35,5 cm; dalam
dada 62,8 cm; lebar pinggul 42,2 cm; dan lingkar dada 160,8 cm (Merkens 1926).
Hasil laporan Otsuka et al. (1980; 1982) yang melakukan survei terbatas di
SNAKMA Saree dan Rumah Potong Hewan Banda Aceh, sapi Aceh betina
mempunyai tinggi pundak 105,0 cm; tinggi pinggul 108,2 cm; panjang badan
118,8 cm; lebar dada 22,0 cm; dalam dada 52,9 cm; lebar pinggul 34,4 cm;
lingkar dada 131,0 cm; panjang kepala 41,7 cm; dan lebar kepala 14,0 cm.
Sapi Aceh jantan mempunyai gumba berukuran sedang (medium),
biasanya mempunyai warna cokelat yang lebih gelap pada tubuh bagian
depannya dibanding bagian belakang (Otsuka et al. 1980). Sapi Aceh,
mempunyai lebar dan tinggi tengkorak lebih pendek dibanding panjangnya dan
bagiancerebraltengkorak lebih besar dari bagian mukanya (Hayashiet al. 1982).
Otsukaet al. (1980; 1982) telah menggunakan ukuran-ukuran tubuh hewan
dalam melakukan perbandingan antara sapi Aceh, Padang, Madura, Bali dan
Grati, serta hubungannya dengan berbagai bangsa sapi lain di Asia (Thai,
Batangas, Taiwan, Iloilo, Ilocos, Kedah-Kelantan, Palawan, Hainan, Cebu,
Kuchinoshima, Mishima dan Sindhi). Menurut Dwiyanto 1982), penggunaan
ukuran tubuh selain untuk menaksir bobot badan dan karkas, dapat digunakan
pula untuk memberikan gambaran bentuk tubuh hewan sebagai ciri khas bangsa
ternak tertentu.
Penanda Molekuler
Keragaman suatu spesies dapat didekati dengan menggunakan penanda
morfologi dan penanda molekuler (Hillis et al. 1996). Informasi mutu genetik
hewan dapat diperoleh dengan penafsiran melalui pencatatan performa produksi
dan reproduksi (penanda morfologi) hewan. Penanda morfologi merupakan
penanda yang telah banyak digunakan dalam program genetika dasar maupun
dalam program praktis pemuliaan, karena penanda ini paling mudah untuk
diamati dan dibedakan. Namun demikian, penanda morfologi memiliki beberapa
mutu genetik hewan dengan penanda morfologi sangat rendah, proses seleksi
berjalan lambat dan respons yang diperoleh sangat kecil, walaupun dilakukan
secara terus-menerus serta terdapat kesalahan dalam pengambilan keputusan
dalam penentuan strategi perkawinan. Dengan demikian untuk kegiatan
pemuliaan tidak cukup hanya berdasarkan pada informasi karakter morfologi
saja, tetapi dengan kemajuan ilmu pengetahuan dan teknologi saat ini, maka
dapat digunakan alternatif penanda lain yaitu penanda molekuler yang telah
relatif mudah untuk dikerjakan (Muladno 2006).
Akhir-akhir ini dengan berkembangnya ilmu biologi molekuler dan lebih
khusus pada genetika molekuler, studi tentang hubungan kekerabatan atau
variasi genetik bangsa ternak sering dilakukan dengan teknik molekuler (Muladno
2002). Penanda molekuler mampu mengidentifikasi perbedaan genetik langsung
pada level DNA sebagai komponen genetik. Semua karakter yang ditampilkan
baik secara nyata atau tidak oleh satu individu hewan tidak lain adalah
pencerminan karakter gen yang dimiliki oleh individu hewan tersebut, atau dapat
disebut bahwa semua informasi yang dapat diamati pada suatu individu hewan
adalah penanda genetik dari individu tersebut. Karakteristik penanda molekuler
ini dapat menanggulangi keterbatasan penggunaan penanda morfologi dapat
diminimalisasi dengan menggunakan penanda molekuler dalam aplikasi program
pemuliaan hewan karena penanda ini bebas dari pengaruh-pengaruh epistasis,
lingkungan dan fenotipe, sehingga dapat menyediakan informasi genetik yang
lebih akurat dalam mempelajari genotipe hewan, mempelajari sifat-sifat genetik
yang kompleks dan keragaman genetik hewan (Muladno 2006).
Sekarang telah ada beberapa penanda DNA untuk menganalisis latar
belakang genetik hibrida pada sapi (Nijman et al. 2003). Penanda DNA
mitokondria (mtDNA) menunjukkan introgresi melalui silsilah maternal (Loftus et
al. 1994; Ward et al. 1999; Mezzadraet al. 2005), yang pada sapi menunjukkan
sejarah kawanannya (Nijman et al. 2003; Ascunce et al. 2007).
Carvajal-Carmonaet al. (2003) melakukan penelitian dengan menguji keragaman sekuens
daerah kontrol DNA mitokondria dari bangsa sapi Criollo pada tujuh wilayah di
Colombia. Penemuan dalam bangsa sapi Criollo disimpulkan bahwa ada
perbedaan ancestor untuk beberapa bangsa sapi yang konsisten dengan pusat
domestikasi sapi di Afrika Utara.
Penanda mikrosatelit digunakan secara luas sebagai penanda genetik di
Erhardt 2005; Cervini et al. 2006), terutama karena mikrosatelit mengandung
informasi polimorfisme yang tinggi, tersebar luas di dalam genom eukariot (Tautz
dan Renz 1984; Cervini et al. 2006). Mikrosatelit sangat efektif dalam
mengevaluasi perbedaan di dalam bangsa sapi dan dalam menentukan
substruktur populasi (MacHugh et al. 1998). Lebih dari 1.400 mikrosatelit telah
dipetakan dalam genom sapi (Luikart et al. 1999; Cervini et al. 2006) dan
beberapa di antara mikrosatelit tersebut digunakan untuk studi genetik populasi
serta verifikasi asal-usul (Cervini et al. 2006). Mikrosatelit digunakan Mukesh
(2004) untuk mengkaji keragaman genetik dan menetapkan hubungan di antara
tiga bangsa sapi zebu India (Sahiwal, Hariana dan Deoni).
Sumber spesies genom inti dapat diduga dari alel-alel mikrosatelit khusus
spesies (MacHugh et al. 1997). Mikrosatelit bersifat polymorphic tinggi atau
hyperpolymorphic dan sangat informatif pada tingkat famili atau hewan
persilangan. Oleh karena itu, mikrosatelit sering digunakan sebagai tool’ yang
potensial dalam pemetaan pautan gen (linkage gene) pada organisme yang
berbeda. Sifat polimorfisme yang tinggi ini memungkinkan individu-individu akan
menjadi heterozigot dan karenanya akan lebih mudah untuk menelusuri
pewarisan suatu penanda pada penelusuran keluarga atau famili. Sifat yang
polimorfisme ini dan terletak sepanjang genom, maka mikrosatelit merupakan
sumber data yang ideal untuk determinasi jarak genetik (Nicholas 1996).
DNA adalah polimer dari nukleotida, terdiri atas molekul yang panjang
berisi beberapa rangkaian monomer nukleotida yang dirangkai dalam satu urutan
berseri dan diorganisasikan dalam bentuk suatu heliks. Setiap nukleotida itu
sendiri adalah suatu molekul yang kompleks terdiri tiga komponen yaitu (1) gula,
(2) basa nitrogen; dan (3) asam phosphat. Gula pada DNA adalah suatu pentosa
(dengan lima atom carbon) yang berbentuk cincin dikenal sebagai
2’-deoxyribose. Basa-basa nitrogen pada DNA adalah berstruktur satu ataupun
dua cincin. Basa-basa tersebut terdiri atas purine (Adenine dan Guanine) dan
pyrimidine(Thymine dan Cytosine) (Duryadi 2005).
Hewan memiliki total untaian DNA dengan ukuran kira-kira sebesar 3x109
basa nukleotida, dengan 10% dari untaian DNA merupakan gen sedangkan
sisanya bersifat sebagai DNA pengisi. Struktur DNA pengisi ini terdiri atas
ulangan yang bersifat khas dan unik. Tingginya persentase urutan berulang pada
DNA sebagai unit keturunan terkecil terdapat pada semua makhluk hidup
mulai dari mikroorganisme sampai organisme tingkat tinggi seperti manusia,
hewan dan tanaman. DNA terdapat dalam sel terdiri atas DNA inti sel dan DNA
sitoplasma. DNA sitoplasma berupa DNA mitokondria dan DNA kloroplast
(Muladno 2002). Genom inti dan sitoplasmik ini menjadi karakter organisme
sehingga dapat dijadikan sebagai acuan dalam perbandingan ciri baik kesamaan
maupun perbedaannya. Semakin dekat kekerabatan pada tingkat takson maupun
tingkatan keturunan (lineage) dari organisme tersebut, maka semakin besar
kesamaan pada tingkatan molekulnya. Hal demikian ini yang menjadi dasar
perunutan hubungan evolusi dan asal-usul organisme (Duryadi 2005).
DNA Mitokondria
Organisme eukariot termasuk ternak domestik, sumber DNA dapat
diperoleh dari organel-organel sitoplasmik antara lain DNA mitokondria. DNA
mitokondria memiliki karakteristik sebagai molekul DNA yang diturunkan secara
utuh tanpa adanya rekombinasi, memiliki molekul dengan ukuran kecil/pendek
yang susunannya berbeda dengan DNA inti, dan memiliki variasi basa nukleotida
yang lebih tinggi dibandingkan DNA inti. Tingginya variasi basa nukleotida
disebabkan DNA mitokondria memiliki laju perubahan 5-10 kali lebih tinggi
dibandingkan DNA inti (Muladno 2006).
Mitokondria memiliki molekul DNA tersendiri dengan ukuran kecil yang
susunannya berbeda dengan DNA inti (Duryadi 1994; Lewin 2000). Setiap sel
mengandung satu hingga ratusan DNA mitokondria. DNA mitokondria
merupakan DN