PERAN FUNGI MIKORIZA ARBUSKULAR DAN BAKTERI
ENDOSIMBIOTIK MIKORIZA DALAM MENINGKATKAN
DAYA ADAPTASI BIBIT KELAPA SAWIT
(Elaeis guineensis
Jacq) TERHADAP CEKAMAN BIOTIK
Ganoderma boninense
Pat
YENNI BAKHTIAR
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
2011
Saya menyatakan dengan sebenar-benarnya, bahwa segala pernyataan dalam disertasi yang berjudul:
PERAN FUNGI MIKORIZA ARBUSKULAR DAN BAKTERI ENDOSIMBIOTIK MIKORIZA DALAM MENINGKATKAN DAYA ADAPTASI BIBIT KELAPA SAWIT (Elaeis guineensisJacq) TERHADAP
CEKAMAN BIOTIKGanoderma boninensePat
merupakan gagasan atau hasil penelitian karya saya sendiri dengan bimbingan dan arahan dari Komisi Pembimbing. Disertasi ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi lain. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di dalam disertasi ini.
Bogor, September 2011
YENNI BAKHTIAR A161060072
ABSTRACT
YENNI BAKHTIAR. Role of Arbusccular Mycorrhizal Fungi and Mycorrhizal Endosymbiotic Bacteria in Increasing Oil Palm (Elaeis guineensisJacq) Seedlings Adaptation Towards Biotic Stress of Fungal Pathogen Ganoderma boninensePat. Under the direction of SUDIRMAN YAHYA (chairman), WAHONO SUMARYONO, MEITY SURADJI SINAGA and SRI WILARSO BUDI (members).
Basal stem rot caused by Ganoderma boninense is the most serious disease of oil palm (Elaeis guineensis) in Indonesia and it has caused major loss in palm oil production. Under natural conditions, oil palm is often colonized by arbuscular mycorrhizal (AM) fungi and together with mycorrhizosphere bacteria offer possible advantages in increasing plant adaptation against biotic stress of pathogen.
This research comprised five interrelated experiments, namely: (1) Exploration, Isolation and Identification of Arbuscular Mycorrhizal Fungi (AMF) and Mycorrhizal Endosymbiotic Bacteria from Oil Palm Rhizosphere and Their Ability in Inducing Oil Palm Adaptation Towards Biotic Stress of Pathogen Ganoderma boninensePat; (2) Selection of Mycorrhizal Endosymbiotic Bacteria on Spores Germination of AMF Gigaspora margarita and Their Inhibition Towards Fungal Pathogen G. boninense Pat; (3) Identification of Active Compounds from Mycorrhizal Endosymbiotic Bacteria B. subtilisB10 that Inhibit the Growth of Fungal Pathogen G. boninense Pat; (4) Adaptation of Oil Palm Seedlings Inoculated with AMF and Mycorrhizal Endosymbiotic Bacteria B. subtilis B10 Towards Biotic Stress of Pathogen G. boninense Pat; (5) Analyzing the Response of Oil Palm Seedlings Inoculated with AMF and Mycorrhizal Endosymbiotic Bacteria in the Form of Active Compound Profile as Expression of Oil Palm Adaptation Towards Biotic Stress of PathogenG. boninensePat.
The soil sampels were used in this experiment came from Aek Pancur Plantation belong to Indonesian Oil Palm Research Institute, Medan. The results showed that twenty isolates of mycorrhizal endosymbiotic bacteria were obtained from spores of AMF isolated from oil palm rhizosphere and were dominated by genus of Bacillus sp. Among these isolates, isolate B10 identified based on 16S rDNA as Bacillus subtilis B10, had the highest activity against G. boninense in vitro by producing intracellular active compounds with molecular weight 255.39 and postulated as of 2-(4-aminophenoxy)-6-methyl-tetrahydro-2H-pyran-3,4,5-triol. Dual inoculation of AMF and B. subtilis B10 on oil palm seedlings resulted in the lowest basal stem rot disease incidence caused by G. boninese which indicated disease severity index was only 5% and necrotic area of roots just 10%. Application of AMF together with B. subtilis B10 also increased oil palm seedlings height, root dry weight and stem diameter at 52 weeks after planting. The adsorption of nitrogen (N), phosphorus (P), potassium (K) and magnesium (Mg) increased when seedlings were co-inoculated with AMF and bacteria B. subtilis B10 together. This finding is important in terms of recommendation for application of AMF and bacteria B. subtilis B10 as biocontrol of pathogen G. boninensecausal agent of basal stem rot in oil palm.
Keywords: Arbuscular mycorrhizal fungi, mycorrhizal endosymbiotic bacteria, oil palm seedlings, adaptation of biotic stress, pathogen Ganoderma boninense.
YENNI BAKHTIAR.
Peran Fungi Mikoriza Arbuskkular dan Bakteri Endosimbiotik Mikoriza dalam Meningkatkan Daya Adaptasi Bibit Kelapa sawit (Elaeis guineensisJacq) terhadap Cekaman Biotik Patogen Ganoderma boninense Pat. Di bawah bimbingan SUDIRMAN YAHYA (ketua), WAHONO SUMARYONO, MEITY SURADJI SINAGA dan SRI WILARSO BUDI (anggota).Penyakit busuk pangkal batang yang disebabkan oleh fungi Ganoderma boninense merupakan penyakit yang paling serius pada kelapa sawit (Elaeis guineensis) di Indonesia karena menyebabkan kerugian besar dalam produksi minyak kelapa sawit. Secara alami, tanaman kelapa sawit dikolonisasi oleh fungi mikoriza arbuskular (FMA) dan telah diketahui memberikan manfaat bagi kelapa sawit. Di daerah rizosfir berbagai jenis mikroorganisme termasuk bakteri hidup di sekitar mikoriza dan mengambil manfaat dari berbagai jenis senyawa organik yang dilepaskan oleh tanaman. Mikroorganisme ini memberikan manfaat bagi perkembangan dan stabilitas mikoriza maupun terhadap tanaman inangnya.
Penelitian ini terdiri dari lima percobaan yang saling terkait, yaitu: (1) Eksplorasi, isolasi dan identifikasi FMA dan bakteri endosimbiotik mikoriza dari rizosfir kelapa sawit. (2) Seleksi bakteri endosimbiotik mikoriza terhadap daya kecambah FMA dan daya hambatnya terhadap Ganoderma boninense. (3) Ekstraksi senyawa aktif dari bakteri endosimbiotik mikoriza yang menghambat pertumbuhan G. boninense dan bersifat sinergis dengan FMA (4) Daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense dengan inokulasi FMA dan bakteri endosimbiotik mikoriza. (5) Menganalisis respon inang dalam bentuk senyawa aktif sebagai ekspresi daya adaptasi kelapa sawit terhadap cekaman biotik G. boninense yang diinokulasi FMA dan bakteri endosimbiotik mikoriza.
Sampel tanah yang digunakan dalam percobaan ini diambil dari rizosfir empat varietas kelapa sawit yaitu Pisifera, Tenera, Dura Deli dan Dura Dumpy yang terdapat pada Kebun Percobaan Aek Pancur Pusat Penelitian Kelapa Sawit, Medan. Hasil penelitian menunjukkan bahwa rizosfir kelapa sawit varietas Dura
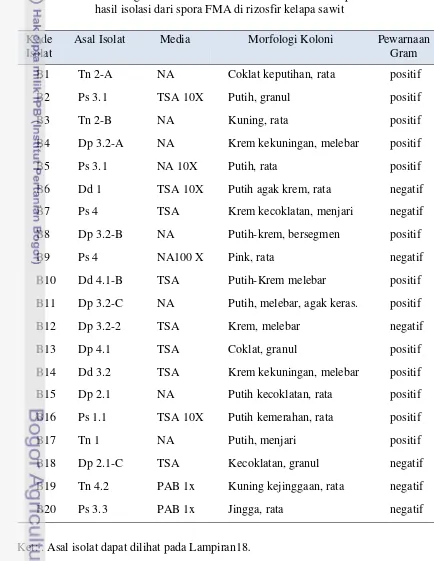
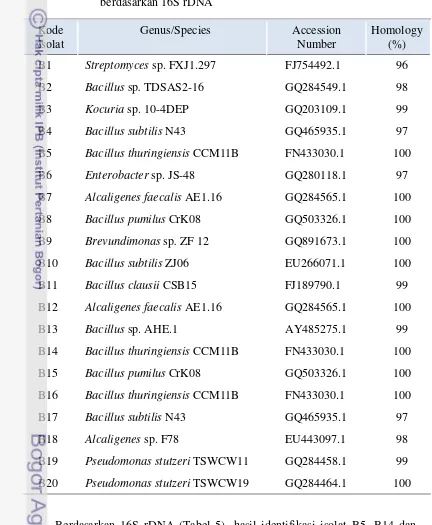
Dumpy memiliki jumlah spora FMA terbanyak berkisar antara 9-127 per 200 g sampel tanah dan jumlah isolat bakteri endosimbiotik mikoriza tertinggi yaitu 24 isolat. Berdasarkan morfologi dan warna koloni diperoleh 20 isolat bakteri endosimbiotik mikoriza dan hasil identifikasi berdasarkan 16S rDNA nya, isolat tersebut terdiri dari genus Bacillus, Streptomyces, Kocuria, Enterobacter, Alcaligenes, Brevundimonasdan Pseudomonas.
Hasil uji antagonis terhadap patogen G. boninensedan uji perkecambahan spora FMA Gigaspora margarita in vitro terhadap keduapuluh bakteri endosimbiotik mikoriza tersebut, bakteri B10 yang diidentifikasi sebagai Bacillus subtilis B10 memiliki kemampuan tertinggi dalam menghambat pertumbuhan patogen G. boninense dengan luas zona hambat mencapai 81,87 mm2. Isolat bakteri B. subtilis B10 tersebut juga memiliki kemampuan meningkatkan persentase berkecambah spora FMA dengan panjang hifa mencapai 1053,32 μm. Bakteri Bacillus subtilis B10 menghasilkan senyawa intra seluler yang dihasilkan secara in vitro, memiliki aktivitas menghambat pertumbuhan Ganoderma boninense pada uji in vitro dengan bobot molekul 255,39 g/mol dan dipostulasikan sebagai 2-(4-aminophenoxy)-6-methyl-tetrahydro-2H-pyran-3,4,5-triol. Senyawa tersebut termasuk kelompok senyawa yang memiliki kemampuan sebagai antifungi.
Hasil uji aplikasi di rumah kaca, dual inokulasi FMA dan bakteri B. subtilis B10 pada bibit kelapa sawit memberikan persentase penyakit busuk pangkal batang terendah dengan indeks keparahan penyakit (KP) tajuk 5% dan luas nekrotik akar 10%. Inokulasi FMA dan bakteri B. subtilisB10 secara tunggal dan bibit tanpa perlakuan (kontrol) memberikan nilai KP dan luas nekrotik akar yang nyata lebih tinggi. Dual inokulasi FMA dan B. subtilis B10 juga meningkatkan tinggi tanaman, berat kering akar dan diameter batang pada 52 minggu setelah tanam. Serapan hara nitrogen (N), fosfor (P), kalium (K) dan magnesium (Mg) juga meningkat ketika bibit di dual inokulasi dengan FMA dan B. subtilisbakteri B10.
Uji in vivo inokulasi fungi mikoriza arbuskular dikombinasikan dengan bakteri endosimbiotik mikoriza B. subtilis B10 pada bibit kelapa sawit menghasilkan kumpulan senyawa aktif yang berperan dalam meningkatkan daya
berfungsi menginduksi bibit kelapa sawit untuk meningkatkan produksi senyawa aktif yang dapat menghambat pertumbuhan patogen G. boninense. Meningkatnya daya adatasi bibit kelapa sawit terhadap cekaman biotik G. boninense dimungkinkan karena bibit kelapa sawit membentuk ketahanan yang dipicu atau di-trigger oleh FMA dan bakteri endosimbiotik mikoriza dengan meningkatkan lignifikasi akar, penyerapan hara yang lebih baik serta produksi senyawa-senyawa yang berperan dalam meningkatkan adaptasi bibit kelapa sawit terhadap cekaman biotik G. boninense.
Temuan dari penelitian ini penting artinya sebagai rekomendasi bagi praktisi perkebunan kelapa sawit untuk memanfaatkan FMA dan bakteri endosimbiotik mikoriza B. subtilis B10 dalam meningkatkan daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense penyebab penyakit busuk batang pada kelapa sawit sehingga dapat meminimalkan kerusakan yang ditimbulkan oleh patogen tersebut.
Kata kunci: Fungi mikoriza arbuskular, bakteri endosimbiotik mikoriza, bibit kelapa sawit, adaptasi cekaman biotik, patogen Ganoderma boninense
© Hak cipta milik Yenni Bakhtiar, tahun 2011 Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apapun, baik cetak,
fotocopy, microfilm dan sebagainya
DAYA ADAPTASI BIBIT KELAPA SAWIT
(Elaeis guineensis
Jacq) TERHADAP CEKAMAN BIOTIK
Ganoderma boninense
Pat
YENNI BAKHTIAR
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Sekolah Pascasarjana
Institut Pertanian Bogor
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011
Judul Disertasi : PERAN FUNGI MIKORIZA ARBUSKULAR DAN BAKTERI ENDOSIMBIOTIK MIKORIZA DALAM MENINGKATKAN DAYA ADAPTASI BIBIT
KELAPA SAWIT (Elaeis guineensisJacq) TERHADAP CEKAMAN BIOTIKGanoderma boninensePat
Nama Mahasiswa : Yenni Bakhtiar
Nomor Pokok : A161060072
Program Studi : Agronomi
Disetujui. Komisi Pembimbing
Prof. Dr. Ir. Sudirman Yahya, M.Sc Prof. Dr. Wahono Sumaryono, Apt
Ketua Anggota
Prof. Dr. Ir. Meity S. Sinaga, M.Sc Dr. Ir. Sri Wilarso Budi, MS
Anggota Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, MS Dr. Ir. Dahrul Syah, M.Sc.Agr
Tanggal Ujian: 28 September 2011 Tanggal Lulus:
Puji dan syukur penulis panjatkan ke hadirat Allah Yang Maha Kuasa atas segala karunia dan hidayahNya, sehingga penullis dapat menyelesaikan penelitian dan penulisan disertasi ini. Tema yang dipilih dalam penelitian yang dilaksanakan dari bulan Maret 2007 sampai bulan Desember 2010 ini adalah Pemanfaatan Fungi Mikoriza Arbuskular dan Bakteri Endosimbiotik Mikoriza pada Tanaman Kelapa Sawit, dengan judul “PERAN FUNGI MIKORIZA ARBUSKULAR
DAN BAKTERI ENDOSIMBIOTIK MIKORIZA DALAM
MENINGKATKAN DAYA ADAPTASI BIBIT KELAPA SAWIT (Elaeis guineensis Jacq) TERHADAP CEKAMAN BIOTIK Ganoderma boninense Pat”. Adapun penelitian ini dilakukan di Laboratorium dan Rumah Kaca di Balai Pengkajian Bioteknologi BPPT.
Selama melaksanakan penelitian ini, penulis banyak mendapatkan bantuan dan bimbingan dari berbagai pihak. Untuk itu pada kesempatan ini penulis ingin menyampaikan ucapan terima kasih kepada:
1. Prof. Dr. Ir. Sudirman Yahya, M.Sc, selaku ketua komisi pembimbing, Prof. Dr. Wahono Sumaryono, Prof. Dr. Ir. Meity Suradji Sinaga dan Dr. Ir. Sri Wilarso Budi, MS, selaku anggota pembimbing atas segala arahan dan bimbingan yang telah diberikan sejak penulisan proposal hingga disertasi ini selesai. Banyak ilmu dan pengalaman berharga yang penulis dapatkan selama bimbingan, yang sangat bermanfaat dalam mempertajam analisis dan sistematika berpikir dalam penyelesaian disertasi ini.
2. Kementrian Riset dan Teknologi dan seluruh Staf yang telah memberikan fasilitas beasiswa kepada penulis untuk melanjutkan Program Doktor. 3. Kepala BPPT dan Deputi Kepala BPPT Bidang Teknologi Agroindustri
dan Bioteknologi yang telah memberikan izin kepada penulis untuk melanjutkan studi ini.
4. Prof. Dr. Ir. Nadirman Haska, MS yang telah berkenan memberikan izin bagi penulis untuk mengikuti Program Doktor dan Dr. Bambang Marwoto, Apt., M.EngSc selaku Kepala Balai Pengkajian Bioteknologi BPPT atas sarana dan waktu yang diberikan dalam penyelesaian studi ini.
5. Pimpinan Institut Pertanian Bogor, Dekan Sekolah Pascasarjana, Dekan Fakultas Pertanian dan Ketua Program Studi Agronomi, yang telah memberikan penulis kesempatan untuk mengikuti Program Doktor ini. 6. Staf Pengajar dan pegawai di lingkungan Program Studi Agronomi,
Fakultas Pertanian untuk bantuan dan kemudahan administrasi.
7. Almarhum ayahanda tercinta Drs. H. Bakhtiar Buyung dan ibunda tercinta Hj. Asnidar serta ibu mertua Hj. Wara Supeni yang tidak pernah bosan memberikan doa, dorongan, semangat kepada penulis untuk bisa menjalani studi ini dengan baik dan lancar.
8. Suami tercinta Ir. Iman Seno Adji M.EngSc, ananda tercinta Iman Trilaksamana dan Shabrina Astrilia, untuk doa yang tulus, dukungan moril dan kesabaran serta kerelaan berbagi waktu dan perhatian dengan studi ini. 9. Taufik Firmansyah S.Si, Abdul Wahid SE dan rekan-rekan di
Laboratorium Agromikrobiologi, Analitik dan Rumah Kaca untuk semua bantuan dan dukungan dalam menyelesaikan studi ini.
10. Kakak dan adik: Yanti, Lily, Lita, Yessi, Ancan, Apit dan seluruh keponakan tercinta; Om dan tante Sumarsono; semua keluarga besar yang telah membantu memberikan doa dan dukungan yang tulus kepada penulis. 11. Rekan seperjuangan Erwahyuni Prabandari, M.Si, teman-teman di Balai Pengkajian Bioteknologi: Farida Rosana Mira, Bedah Rupaedah, Anis Matsunah, Syofie Rosmalawati dan teman-teman lainnya atas segala perhatian, diskusi, bantuan, saran, dorongan dan semangat hingga selesainya studi ini.
12. Teman-teman seperjuangan di Program Studi Agronomi yang selalu memberikan semangat dan pemicu untuk segera menyelesaikan studi ini. Besar harapan penulis, semoga disertasi ini dapat bermanfaat dalam pengembangan ilmu pengetahuan bagi semua pihak yang membutuhkannya.
Bogor, September 2011
YENNI BAKHTIAR
Penulis dilahirkan di Padang pada tanggal 26 Agustus 1966, sebagai anak ketiga dari pasangan Drs. H. Bakhtiar Buyung (almarhum) dan Hj. Asnidar. Pada tanggal 7 Agustus 1992 penulis menikah dengan Ir. Iman Seno Adji, M.EngSc, dan saat ini dikarunia dua orang anak yaitu Iman Trilaksamana (19 tahun) dan Shabrina Astrilia (17 tahun).
Penulis menyelesaikan pendidikan Sekolah Dasar Negeri No 27 Padang tahun 1979, Sekolah Menengah Pertama Negeri 1 Padang tahun 1982 dan Sekolah Menengah Atas Negeri 1 Padang tahun 1985.
Pendidikan sarjana ditempuh di Jurusan Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Padjadjaran Bandung dan lulus tahun 1990. Pada tahun 1998 penulis berkesempatan melanjutkan pendidikan master (S2) di Departemen Soil and Water, Faculty of Agriculture, University of Adelaide, South Australia dengan beasiswa dari AUSAID dan lulus tahun 2000. Selanjutnya pada tahun 2006, penullis mendapatkan kesempatan untuk melanjutkan pendidikan ke Program Doktor pada Program Studi Agronomi di Sekolah Pascasarjana Institut Pertanian Bogor dengan beasiswa dari Kementrian Riset dan Teknologi.
Penulis bekerja di Balai Pengkajian Bioteknologi, Badan Pengkajian dan Penerapan Teknologi sejak tahun 1991 sampai sekarang.
xii
DAFTAR ISI
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xx
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 6
Perumusan Masalah ... 6
Hipotesis ... 7
Strategi Penelitian ... 8
Manfaat Penelitian ... 9
Diagram Alir Penelitian ... 10
TINJAUAN PUSTAKA 11 Tanaman Kelapa Sawit ... 11
Arti Penting Tanaman Kelapa Sawit ... 11
Faktor Lingkungan Tumbuh Tanaman Kelapa Sawit ... 12
Pengaruh Faktor AbiotikTerhadap Pertumbuhan Kelapa Sawit ... 14
Penyakit Busuk Pangkal Batang pada Kelapa Sawit ... 16
Ganoderma boninensePat Penyebab Penyakit Busuk Pangkal Batamg .. 17
Fungi Mikoriza Arbuskular ... 19
Pengertian Umum Fungi Mikoriza Arbuskular (FMA) ... 19
Peranan FMA dalam Penyerapan Hara ... 20
Peranan FMA terhadap Cekaman Abiotik Kekeringan ... 22
Peranan FMA terhadap Cekaman Abiotik Toksisitas Logam Berat ... 23
Perana FMA terhadap Cekaman Biotik Patogen ... 25
Bakteri Endosimbiotik Mikoriza ... 27
Pengertian Umum Bakteri Endosimbiotik ikoriza ... 27
Interaksi Mutualisme FMA dan Bakteri Endosimbiotik Mikoriza ... 28
EKSPLORASI, ISOLASI DAN IDENTIFIKASI FUNGI MIKORIZA ARBUSKULAR DAN BAKTERI ENDOSIMBIOTIK MIKORIZA DARI RIZOSFIR KELAPA SAWIT ... 31
Abstrak ... 31
Abstract ... 31
Pendahuluan ... 32
Bahan dan Metode ... 35
Hasil dan Pembahasan ... 41
Hasil ... 41
Pembahasan ... 49
Simpulan ... 53
Daftar Pustaka ... 53
SELEKSI BAKTERI ENDOSIMBIOTIK MIKORIZA TERHADAP DAYA KECAMBAH SPORA FMA Gigaspora margarita DAN DAYA HAMBATNYA
TERHADAP PATOGEN Ganoderma boninensePat ... 57
Abstrak ... 57
Abstract ... 57
Pendahuluan ... 58
Bahan dan Metode ... 61
Hasil dan Pembahasan ... 65
Hasil ... 65
Pembahasan ... 69
Simpulan ... 74
Daftar Pustaka ... 74
IDENTIFIKASI SENYAWA AKTIF DARI BAKTERI ENDOSIMBIOTIK MIKORIZA Bacillus subtilis ZJ06 YANG MENGHAMBAT PERTUMBUHAN PATOGEN Ganoderma boninense Pat ... 79
Abstrak ... 79
Abstract ... 79
Pendahuluan ... 80
Bahan dan Metode ... 82
Hasil dan Pembahasan ... 85
Hasil ... 85
Pembahasan ... 96
Simpulan ... 98
Daftar Pustaka... 98
DAYA ADPTASI BIBIT KELAPA SAWIT DIINOKULASI FUNGI MIKORIZA ARBUSKULAR DAN BAKTERI ENDOSIMBIOTIK MIKORIZA TERHADAP CEKAMAN BIOTIK PATOGEN Ganoderma boninense Pat ... 101
Abstrak ... 101
Abstract ... 102
Pendahuluan ... 103
Bahan dan Metode ... 105
Hasil dan Pembahasan ... 110
Hasil ... 110
Pembahasan ... 128
Simpulan ... 132
Daftar Pustaka ... 133
ANALISIS RESPON INANG DALAM BENTUK PROFIL SENYAWA AKTIF SEBAGAI EKSPRESI DAYA ADAPTASI KELAPA SAWIT DIINOKULASI FMA DAN BAKTERI ENDOSIMBIOTIK MIKORIZA TERHADAP CEKAMAN BIOTIK PATOGEN Ganoderma boninense Pat ... 137
Abstrak ... 137
Abstract ... 137
Pendahuluan ... 138
Bahan dan Metode ... 140
Hasil dan Pembahasan ... 141
Hasil ... 141
xiv
Pembahasan ... 147
Simpulan ... 150
Daftar Pustaka ... 151
PEMBAHASAN UMUM ... 154
SIMPULAN UMUM DAN SARAN ... 172
Simpulan Umum ... 172
Saran ... 174
DAFTAR PUSTAKA ... 175
LAMPIRAN ... 192
No. Judul Halaman
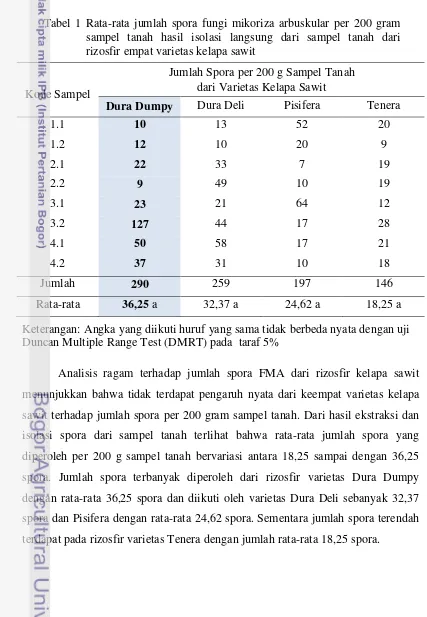
1. Rata-rata jumlah spora fungi mikoriza arbuskular per 200 gram sampel tanah hasil isolasi langsung dari sampel tanah dari rizosfir empat varietas kelapa
sawit ... 41
2. Jenis spora fungi mikoriza arbuskular hasil isolasi dari rizosfir kelapa sawit dari kebun Percobaan Aek Pancur Pusat Penelitian Kelapa Sawit ... 42
3. Keragaman isolat bakteri endosimbiotik mikoriza yang tumbuh pada media isolasi nutrient agar, tryptic soy agardan pseudomonas agar base ... 45
4. Morfologi koloni bakteri endosimbiotik mikoriza pada media hasil isolasi dari spora FMA dari rizosfir kelapa sawit pada media nutrient agar... 47
5. Hasil identifikasi bakteri endosimbiotik mikoriza terseleksi berdasarkan 16S rDNA ... 48
6. Rata-rata panjang hifa pada perkecambahan spora fungi mikoriza arbuskular Gigaspora margaritapada hari keempat belas dengan inokulasi bakteri endosimbiotik mikoriza ... 66
7. Rata-rata luas zona bening yang terbentuk sebagai aktivitas antagonis senyawa aktif Bacillus subtilisB10 pada masing-masing fraksi pelarut dan kontrol positif nystatin terhadap G. boninense ... 86
8. Rata-rata panjang hifa pada hari kelima sebagai aktivitas senyawa aktifBacillus subtilisB10 pada masing-masing fraksi dalam meningkatkan persentase berkecambah spora FMA Gigaspora margarita... 87
9. Uji setara aktivitas senyawa aktif pada beberapa konsentrasi dari bakteri endosimbiotik mikoriza Bacillus subtilisB10 terhadap pertumbuhan G. boninense dengan kontrol (+) antifungi nystatin ... 88
10. Penghitungan bobot molekul senyawa aktif Bacillus subtilisB10 ... 91
11. Hasil identifikasi gugus fungsi spektrum infra merah senyawa aktif dari bakteri endosimbiotik mikoriza Bacillus subtilisB10 ... 93
12. Rekapitulasi hasil sidik ragam peubah yang diamati terhadap inokulasi FMA, bakteri endosimbiotik mikoriza pada bibit kelapa sawit yang diberi cekaman biotik patogen G. boninense... 111
xvi
DAFTAR GAMBAR
No. Judul Halaman
1. Morfologi isolat murni bakteri endosimbiotik mikoriza hasil isolasi dari spora FMA dari rizosfir kelapa sawit pada media nutrient agar ...
47 2. Pohon filogenetik isolat bakteri endosimbiotik mikoriza terseleksi berdasarkan homologinya ... 49
3. Panjang hifa (garis hijau) dari perkecambahan spora FMA Gigaspora margaritadengan inokulasi bakteri B17 Bacillus subtilisB17 (kiri) dan tanpa inokulasi
bakteri sebagai kontrol (kanan), pada perbesaran 100x ... 67
4.
Grafik aktivitas bakteri endosimbiotik mikoriza pada hari keempat setelah inokulasi terhadap pertumbuhan fungi patogen G. boninenseberupa luas zona bening yang terbentuk di sekeliling kertas cakram. Terlihat bakteri B10 memiliki luas zona hambat terbesar . huruf yang berbeda menunjukkan beda nyata dengan uji Duncan pada taraf 5% ...
68
5.
Aktivitas bakteri endosimbiotik mikoriza B10 (kanan) terhadap pertumbuhan G. boninense in vitropada hari keempat. Zona bening yang terbentuk (tanda
panah) menunjukkan aktivitas penghambatan oleh bakteri B10 (kanan) dan
tanpa inokulasi bakteri endosimbiotik mikoriza (kiri) ... 69 6. Morfologi sel bakteri Bacillus subtilisB10 pada perbesaran 100x ... 85
7.
Uji setara aktivitas senyawa aktif dari bakteri Bacillus subtilis B10 terhadap pertumbuhan G. boninense in vitrohari ke empat (A) dan hari kedua belas (B). Senyawa aktif dari bakteri B10 memiliki daya hambat yang lebih besar
dibandingkan nystatin (kontrol positif) ... 89
8. Miselia fungi patogen G. boninenseakibat mengalami nekrotik oleh aktivitas senyawa aktif dari bakteri B. subtilisterlihat berwarna coklat (tanda panah)
B10 ... 90
9. Kromatogram senyawa aktif dari bakteri endosimbiotik mikoriza Bacillus subtilisB10 dengan LC-MS ... 91
10. Spektra spektroskopi massa senyawa aktif dari bakteri endosimbiotik mikoriza Bacillus subtilisB10 ...
92 11. Spektrum FT IR senyawa aktif dari bakteri endosimbiotik mikoriza subtilis B10 ...Bacillus 92
12. Spektra subtilisB10 ...1H-NMR senyawa aktif dari bakteri endosimbiotik mikoriza Bacillus
94 13. Posisi proton pada nilai geseran kimia (a) dan posisi gugus rhamnosida pada struktur senyawa postulat (b) ... 94 14. Kemiripan nilai geseran kimia 1Hsenyawa aktif hasil prediksi ... 95
15. Postulasi struktur kimia senyawa aktif dari bakteri Bacillus subtilisB10 ... 96
16.
Presentase indeks Keparahan Penyakit (KP) tajuk bibit kelapa sawit 52 MST. Inokulasi FMA dan bakteri Bacillus subtilisB10 (M1B1) memberikan
persentase KP yang paling rendah .Huruf yang sama pada grafik tidak berbeda
nyata dengan uji Duncan pada taraf 5% ... 113
17.
Pertumbuhan bibit kelapa sawit 52 MST. (A) Bibit tanpa inokulasi FMA, bakteri endosimbiotik mikoriza B. subtilisB10 dan G. boninense(M0B0G0). (B) Bibit tanpa inokulasi FMA dan bakteri endosimbiotik mikoriza B. subtilis B10 dengan inokulasi G. boninese(M0B0G1), terlihat tumbuh tubuh buah Ganoderma, daun dan batang bewarna coklat dan mengering. (C) Bibit dengan inokulasi bakteri endosimbiotik mikoriza B. subtilisB10 dan G. boninense, dimana daun terlihat coklat dan mengering (M0B1G1). (D) Bibit dengan inokulasi FMA, bakteri endosimbiotik mikoriza B. subtilisB10 dan patogen G.
boninense(M1B1G1) walapun ada patogen bibit tumbuh lebih sehat ... 113
18.
Bibit kelapa sawit pada 52 MST yang lebih tahan serangan G. boninense, terlihat tumbuh tubuh buah (tanda panah) tapi kondisi bibit masih terlihat sehat walaupun beberapa daun terlihat berwarna cokelat dan mengering (A). Kondisi bibit kelapa sawit yang tidak tahan serangan G. boninense, terlihat tumbuh tubuh buah (tanda panah), semua daun dan batang berwarna cokelat, mengering dan mati (B) ... 114
19.
Persentase luas nekrotik akar bibit kelapa sawit pada 52 MST, Inokulasi FMA dan bakteri Bacillus subtilisB10 (M1B1) memberikan persentase KP yang paling rendah. Huruf yang sama pada grafik tidak berbeda nyata dengan uji
Duncan pada taraf 5% ... 115
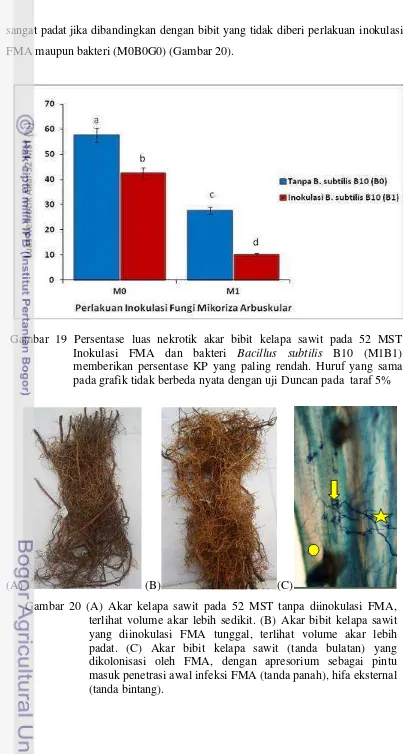
20.
(A) Akar kelapa sawit pada 52 MST tanpa diinokulasi FMA, terlihat volume dan akar lebih sedikit. (B) Akar bibit kelapa sawit yang diinokulasi FMA tunggal, terlihat volume akar lebih padat. (C) Akar bibit kelapa sawit (tanda bulatan) yang diinokulasi oleh FMA, dengan apresorium sebagai pintu masuk
penetrasi awal infeksi FMA (tanda panah), hifa eksternal (tanda bintang) ... 115
21.
(A) Akar bibit kelapa sawit 52 MST yang diberi perlakuan inokulasi bakteri B. subtilisB10 dan diberi cekaman biotik G. boninense, terlihat volume akar lebih sedikit, akar berwarna coklat kehitaman (tanda panah) karena nekrotik. (B) Akar yang didual inokulasi FMA dan bakteri B. substilis B10 dengan inokulasi G. boninense, volume akar sehat lebih banyak dengan nekrotik akar lebih
sedikit ... 116
22.
Persentase kolonisasi FMA akar bibit kelapa sawit pada 52 MST. Inokulasi patogen G. boninense menurunkan persentase kolonisasi akar oleh FMA. Huruf yang sama pada grafik tidak berbeda nyata dengan uji Duncan pada taraf
5% .... ... 117 23. Kolonisasi FMA pada akar bibit kelapa sawit. Terlihat adanya struktur vesikel (V), arbuskular (A) dan hifa internal (tanda panah) ...
118
24. Jumlah spora FMA per 100 gram media tanam pada 52 MST. Perlakuan patogen G. boninensesedikit menurunkan jumlah spora FMA. Dual inokulasi FMA dan bakteri B. subtilisB10 memberikan jumlah spora tertinggi. Huruf
xviii
yang sama pada grafik tidak berbeda nyata dengan uji Duncan pada taraf 5%
... 119
25. Spora fungi mikoriza arbuskular yang diisolasi dari rizosfir kelapa sawit. Spora yang diperoleh didominasi oleh genus Glomus (A dan B) danGigaspora(C dan
D) ... 119
26.
Rata-rata tinggi bibit kelapa sawit 52 MST. Tanaman yang didual inokulasi FMA dan bakteri B. subtilisB10 memiliki tinggi tanaman yang lebih baik daripada yang diinokulasi bakteri B. subtilisB10 saja. Huruf yang sama pada
grafik tidak berbeda nyata dengan uji Duncan pada taraf 5% ... 120
27.
Rata-rata jumlah daun bibit kelapa sawit 52 MST. Cekaman biotik patogen G. boninensemenurunkan jumlah daun bibit kelapa sawit. Inokulasi FMA secara tunggal maupun kombinasi dengan bakteri B. subtilisB10 memberikan jumlah daun paling tinggi. Huruf yang sama pada grafik tidak berbeda nyata dengan uji Duncan pada taraf 5% ... 121
28.
Rata-rata diameter batang bibit kelapa sawit 52 MST. Inokulasi FMA secara tunggal maupun kombinasi dengan bakteri B. subtilisB10 memberikan diameter batang lebih besar Huruf yang sama pada grafik tidak berbeda nyata
dengan uji Duncan pada taraf 5% ... 122
29.
Rata-rata bobot kering akar bibit kelapa sawit 52 MST. Cekaman biotik patogen G. boninensemenurunkan bobot kering akar bibit kelapa sawit. Dual inokulasi FMA dan bakteri B. subtilisB10 memberikan bobot kering akar paling tinggi. Huruf yang sama pada grafik tidak berbeda nyata dengan uji
Duncan pada taraf 5% ... 123
30.
Rata-rata serapan hara fosfor (P) daun bibit kelapa sawit 52 MST. Inokulasi bakteri B. subtilisB10 memberikan serapan hara P daun paling tinggi. Inokulasi FMA tidak meningkatkan serapan hara P daun. Huruf yang sama pada grafik
tidak berbeda nyata dengan uji Duncan pada taraf 5% ... 124
31.
Rata-rata serapan hara nitrogen (N) daun bibit kelapa sawit 52 MST. Inokulasi bakteri B. subtilisB10 secara tunggal maupun kombinasi dengan FMA
memberikan serapan hara N daun paling tinggi. Huruf yang sama pada grafik
tidak berbeda nyata dengan uji Duncan pada taraf 5% ... 125
32.
Rata-rata serapan hara kalium (K) daun bibit kelapa sawit 52 MST. Inokulasi bakteri B. subtilisB10 secara tunggal maupun kombinasi dengan FMA
meningkatkan serapan hara K daun. Huruf yang sama pada grafik tidak berbeda
nyata dengan uji Duncan pada taraf 5% ... 126
33.
Rata-rata serapan hara magnesium (Mg) daun bibit kelapa sawit 52 MST. Inokulasi bakteri B. subtilisB10 secara tunggal maupun kombinasi dengan FMA memberikan serapan hara Mg daun paling tinggi. Huruf yang sama pada
grafik tidak berbeda nyata dengan uji Duncan pada taraf 5% ... 127 34. Profil kromatogram metanol yang digunakan sebagai fasa gerak dalam analisis ekstrak akar menggunakan HPLC. AU = Absorbance Unit ...
141 35. Profil kromatogram ekstrak akar bibit kelapa sawit yang tidak dinokulasikan
FMA, bakteri B. subtilis B10 dan G. boninense (M0B0G0) hasil analisis
menggunakan HPLC ... 142
36. Profil kromatogram ekstrak akar bibit kelapa sawit yang tidak dinokulasikan FMA, bakteri B. subtilis B10 akan tetapi diinfeksi patogen G. boninense
(M0B0G1) hasil analisis menggunakan HPLC ... 143 37. Profil kromatogram ekstrak akar bibit kelapa sawit yang dinokulasikan FMA dan diinfeksi G. boninense (M1B0G1) hasil analisis menggunakan HPLC ...
144
38. Profil kromatogram ekstrak akar bibit kelapa sawit yang dinokulasikan bakteri B. subtilis B10 dan diinfeksi G. boninense (M0B1G1) hasil analisis
menggunakan HPLC ... 145
39. Profil kromatogram ekstrak akar bibit kelapa sawit yang dinokulasikan FMA, bakteri B. subtilis ZJ06 dan diinfeksi G. boninense (M1B1G1) hasil analisis
menggunakan HPLC ... 146
40.
Ekstrak akar bibit kelapa sawit yang sudah dibuat dalam konsentrasi yang sama untuk dianalisis dengan HPLC. (kiri) Ekstrak akar dari perlakuan inokulasi FMA dengan cekaman biotik patogen G. boninense (M1B0G1). (kanan) Ekstrak akar dari perlakuan dual inokulasi FMA dan bakteri B. subtilis B10 dengan
cekaman biotik patogen G. boninense(M1B1G1) ... 147
xx
DAFTAR LAMPIRAN
No. Judul Halaman
1. Ekstraksi spora dengan teknik tuang saring basah ... 191
2. Pengamatan kolonisasi FMA pada akar tanaman ... 192
3. Analisis sidik ragam luas nekrotik akar bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza ... 193
4. Analisis sidik ragam indeks keparahan penyakit bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza ... 193
5. Analisis sidik ragam bobot kering tajuk (g) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza diinvasi patogen G. boninense ... 194
6. Analisis sidik ragam bobot kering akar (g) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza diinvasi patogen G. boninense ... 194
7. Analisis sidik ragam tinggi tanaman (cm) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense... 195
8. Analisis sidik ragam jumlah daun (helai) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 195
9. Analisis sidik ragam diameter batang (cm) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 196
10. Analisis sidik ragam kolonisasi FMA pada akar bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 196
11. Analisis sidik ragam jumlah spora FMA/10 g media bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 197
12. Analisis sidik ragam serapan hara nitrogen pada daun (g/tan) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 197
13. Analisis sidik ragam serapan hara fosfor pada daun (g/tan) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 198
14. Analisis sidik ragam serapan hara kalium pada daun (g/tan) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan diinvasi patogen G. boninense ... 198
15.
Analisis sidik ragam serapan hara magnesium pada daun (g/tan) bibit kelapa sawit yang diinokulasi dengan FMA dan bakteri endosimbiotik mikoriza dan
diinvasi patogen G. boninense ... 199 16. Analisis sifat-sifat tanah dan media percobaan ... 200 17. Rekomendasi pemupukan bibit kelapa sawit yang dilaksanakan oleh Pusat Penelitian Kelapa Sawit (PPKS) Medan ... 201 18. Isolat bakteri endosimbiotik mikoriza hasil isolasi dari spora FMA dari empat varietas kelapa sawit pada media agar yang berbeda ... 202
PENDAHULUAN
Latar Belakang
Kelapa sawit (Elaeis guineensis Jacq.) merupakan tanaman perkebunan yang penting di Indonesia sebagai penghasil minyak nabati beserta beberapa produk turunan lainnya. Pada saat Indonesia mengalami krisis ekonomi, industri kelapa sawit merupakan salah satu agroindustri andalan yang menghasilkan devisa bagi negara. Perkembangan industri kelapa sawit pada dekade terakhir ini berkembang sangat pesat sehingga menempatkan Indonesia sebagai produsen minyak kelapa sawit terbesar di dunia. Setelah tahun 2005, produksi minyak sawit dunia menjadi penyumbang terbesar dalam produksi minyak sayur dunia menggantikan minyak kedelai. Disamping itu, krisis energi yang melanda dunia membuat orang berusaha untuk mencari energi alternatif yang dapat diperbaharui (renewable energy).
Luas perkebunan kelapa sawit di Indonesia pada tahun 2010 mencapai lebih dari 8,04 juta ha dengan produksi minyak sawit (CPO) sekitar 19,76 juta ton (Ditjenbun 2011). Tahun 2010 nilai ekspor minyak kelapa sawit Indonesia dan turunannya mencapai 17,1 juta ton CPO dengan nilai 14,1 miliar US$ (Ditjenbun 2011). Sejak tahun 2007, Indonesia telah menjadi negara penghasil minyak kelapa sawit terbesar dunia menggeser kedudukan Malaysia, akan tetapi secara umum produktivitas tanaman kelapa sawit Indonesia masih kalah dibandingkan dengan Malaysia. Produktivitas tanaman kelapa sawit di Indonesia 2,3 ton – 3,4 ton CPO/ha/tahun (Ditjenbun 2010) sementara Malaysia mencapai 3,8 ton – 4,1 tonCPO/ha/tahun (MPOB, 2010). Sebagai negara tropis yang masih memiliki lahan cukup luas, Indonesia berpeluang besar untuk mengembangkan perkebunan kelapa sawit dan meningkatkan produktivitasnya sehingga dapat menggeser posisi Malaysia, baik melalui penanaman modal asing maupun skala perkebunan rakyat (Sastrosayono 2003).
Salah satu hambatan utama dalam budidaya kelapa sawit ialah adanya cekaman biotik berupa serangan penyakit. Di antara penyakit yang menyerang tanaman kelapa sawit, penyakit busuk pangkal batang yang disebabkan oleh fungi Ganoderma boninense Pat. merupakan penyakit yang paling merugikan
(Semangun 1990; Treu 1998). Saat ini penyakit busuk pangkal batang kelapa sawit menjadi penyakit terpenting pada perkebunan kelapa sawit di Indonesia (Turner 1981; Darmono 2000). Pada beberapa kebun kelapa sawit di Indonesia, penyakit ini telah menimbulkan kematian sampai 80% atau lebih dari seluruh populasi tanaman kelapa sawit, sehingga mengakibatkan penurunan produksi kelapa sawit per satuan luas (Susanto et al. 2003). Dahulu diyakini bahwa G. boninense hanya menyerang tanaman tua, tetapi pada saat ini G. boninense diketahui juga menyerang tanaman belum menghasilkan (TBM) yang berumur 1 tahun. Kejadian penyakit meningkat sejalan dengan replanting kebun kelapa sawit. Gejala penyakit akan lebih cepat muncul dan serangannya lebih berat pada tanaman replanting kedua atau ketiga. Kejadian penyakit pada tanaman TBM pada replantingsatu, dua, tiga dan empat masing-masing sebesar 0, 4, 7, dan 11%. Sementara itu pada tanaman produktif pada replantingsatu, dua, dan tiga masing-masing sebesar 17, 18, dan 75% (Susanto & Sudartho 2003).
Beberapa faktor yang mempengaruhi perkembangan penyakit BPB di lapangan adalah umur tanaman, jenis tanah, status hara dan teknik replanting (Arifin et al. 2000). Infeksi oleh fungi G. boninese dimulai ketika G. boninense kontak dengan akar kelapa sawit dan dengan cepat mendegradasi pati, lignin dan selulosa dari akar kelapa sawit. Penghancuran yang intensif akan menyebabkan pecahnya dinding sel korteks akar kelapa sawit, sehingga terjadi kebocoran sel dan lama kelamaan tanaman akan mati (Reeset al. 2009).
Sampai saat ini sudah banyak upaya dilakukan untuk mengendalikan penyakit tersebut yang meliputi pengendalian kultur teknis, mekanis dan kimiawi. Semua usaha pengendalian tersebut di atas belum memberikan hasil yang memuaskan (Susanto 2002). Berdasarkan kegagalan pengendalian yang tidak terpadu tersebut dan sifat Ganoderma yang tular tanah (soil borne), nekrotropik serta mempunyai alat pertahanan diri yang bermacam-macam (Abadi 1987; Hadiwiyono et al. 1997) maka pengendalian penyakit BPB harus bersifat terpadu antara pemanfaatan tanaman kelapa sawit tahan, penggunaan agen pengendalian hayati superior dan tindakan kultur teknis yang benar. Salah satu alternatif pencegahan dan pegendalian yang dapat dilakukan adalah dengan pemanfaatan mikroba tanah yang dapat bersimbiosis dengan akar kelapa sawit, seperti Fungi
3
Mikoriza Arbuskular (FMA) dan pemanfaatan bakteri endosimbiotik mikoriza yang hidup bersama mikoriza.
Interaksi menguntungkan antara tanaman dengan mikroorganisme di daerah rizosfer diyakini banyak memberikan manfaat bagi tanaman. Interaksi yang terjadi antara akar tanaman, mikroba tanah dan partikel dalam tanah ikut berperan dalam meningkatkan pertumbuhan dan produktivitas tanaman. Interaksi tersebut umumnya terjadi antara bakteri tanah dan fungi yang hidup secara kooperatif di antara atau di dalam akar-akar tanaman. Bakteri tanah seperti rhizobium, pseudomonas memberikan kontribusi pada kesehatan tanaman dengan memfasilitasi fiksasi hara, penyediaan zat-zat yang dibutuhkan untuk pertumbuhan atau mengurangi serangan patogen. Fungi juga memberikan sumbangan yang bermanfaat di dalam rizosfer, termasuk pendistribusian air dan pembebasan hara dari ikatan logam dalam tanah. Interaksi akar tanaman dan fungi yang saling menguntungkan yang umum dijumpai adalah mikoriza.
Fungi mikoriza arbuskular (FMA) adalah fungi yang bersimbiosis secara mutualisme dengan akar tanaman yang berperan penting dalam siklus hara dalam ekosistem. Fungi mikoriza arbuskular ini sangat bermanfaat bagi tanaman terutama dalam meningkatkan penyerapan unsur hara, meningkatkan ketahanan tanaman terhadap serangan patogen, meningkatkan ketahanan terhadap kekeringan dan meningkatkan stabilitas agregat tanah (Menge 1984). Secara umum, FMA tidak banyak menyebabkan perubahan morfologi akar tanaman inang, akan tetapi secara fisiologi terjadi perubahan yang nyata, seperti perubahan konsentrasi zat pengatur tumbuh pada jaringan, meningkatnya aktivitas fotosintesis dan perubahan penyebaran hasil fotosintesis pada akar dan pucuk (Linderman 1994). Peningkatan penyerapan unsur hara dari tanah menyebabkan perubahan pada status hara jaringan tanaman inang yang pada akhirnya akan mengubah struktur dan aspek biokimia sel-sel akar. Perubahan ini pada akhirnya akan membuat tanaman lebih sehat, dapat bertahan pada cekaman abiotik dan biotik (Linderman 1994).
Perubahan proses fisiologi pada tanaman inang dan interaksi biologis di daerah lingkungan tanah yang dipengaruhi oleh mikoriza, diyakini juga akan mempengaruhi kejadian penyakit pada tanaman. Peranan FMA dalam
pengendalian hayati penyakit tanaman telah menjadi pembicaraan bagi para ahli, dengan berbagai ragam respon dan interpretasi (Jalali & Jalali 1991).
Penggunaan FMA pada tanaman pertanian, kehutanan dan perkebunan telah dikenal secara luas. Lebih dari 80 % tanaman dapat berasosiasi dengan FMA ini (Sieverding 1991) termasuk tanaman kelapa sawit. Kesesuaian tanaman inang dengan isolat FMA menentukan keberhasilan simbiosis antara tanaman dengan FMA (Menge 1984). Telah banyak ditemukan hasil-hasil penelitian yang menyebutkan manfaat inokulasi FMA terhadap peningkatan pertumbuhan tanaman kelapa sawit. Blal et al.(1990) menyatakan bahwa tanaman kelapa sawit yang diinokulasi dengan FMA menyerap fosfor lebih banyak dibandingkan tanaman yang tidak diinokulasi dan merupakan faktor yang penting bagi optimasi penyerapan P dalam produksi bibit tanaman kelapa sawit di daerah tropis sehingga dapat mengurangi pemupukan. Pernyataan tersebut dikuatkan oleh Bakhtiar et al. (2002) yang menyimpulkan bahwa aplikasi FMA pada tanaman kelapa sawit mengurangi penggunaan pupuk fosfat sebanyak 50%.
Hasil penelitian Hashim (2004) menunjukkan bahwa inokulasi FMA pada bibit kelapa sawit yang diikuti dengan inokulasi fungi patogen Ganoderma, mampu memperpanjang masa inkubasi patogen dalam menyebabkan infeksi ataupun menyebabkan kematian pada bibit. Setelah 9 bulan, semua bibit kelapa sawit yang tidak diinokulasi FMA menunjukkan gejala penyakit yang disebabkan oleh Ganoderma. Sementara itu hanya 20% bibit yang diinokulasi mikoriza menunjukkan gejala penyakit tersebut dan hanya 10% yang menyebabkan kematian pada bibit kelapa sawit.
Fungi mikoriza arbuskular berinteraksi dengan bakteri tanah yang mempengaruhi perkembangan dan kestabilan simbiosisnya, yang dapat bersifat positif, negatif maupun netral. Interaksi antara FMA dan Plant Growth Promoting Bacteria (PGPB) seperti bakteri penambat nitrogen, Pseudomonads golongan fluoresen merupakan contoh sinergis interaksi positif antara FMA dengan bakteri tanah (Hameeda et al. 2007). Meyer & Linderman (1986) melaporkan bahwa PGPB seperti Pseudomonas putida meningkatkan kolonisasi FMA pada tanaman subterranean clover. Sementara Azcon (1987) melaporkan bahwa pertumbuhan miselium dari spora Glomus mosseae meningkat dengan adanya PGPB.
5
bakteri pembentuk nodul seperti Frankia, Rhizobium dan Bradyrhizobium umumnya membentuk interaksi yang sinergis dengan FMA. Simbiosis FMA menurunkan cekaman fosfat bagi tanaman. Kondisi tersebut menguntungkan bagi sistem nitrogenase fiksasi-N2 bakteri karena meningkatkan fiksasi dan status N
dari tanaman, yang akan meningkatkan pertumbuhan tanaman dan perkembangan FMA (Bethlenfalvay 1992). Bentuk interaksi antara FMA dan bakteri tanah ditentukan oleh faktor lingkungan, jenis bakteri, jenis FMA dan jenis tanaman.
Walaupun FMA telah dikenal dapat meningkatkan penyerapan berbagai macam unsur hara dari tanah terutama fosfor (Marschner 1995; Lange-Ness & Vlek 2000), beberapa studi membuktikan bahwa inokulasi dengan beberapa PGPB dapat meningkatkan hubungan simbiosis antara tanaman dengan fungi simbion. Hasil penelitian Ratti et al. (2001) menyimpulkan bahwa kombinasi FMA Glomus aggregatum dan PGPB Bacillus polymyxa dan Azospirillum brasilense memaksimalkan produksi biomasa dan kadar fosfor (P) dari Cymbopogon martinii ketika ditanam pada media dengan fosfat anorganik yang tidak larut. Sementara Toro et al. (1997) menemukan bahwa Enterobacter sp dan Bacillus subtillismerangsang kestabilan pembentukan FMA, Glomus intraradices serta meningkatkan biomassa tanaman dan kadar N dan P dalam jaringan. Kim et al. (1998) menemukan bahwa kadar P pada tanaman tomat meningkat dengan inokulasi baik itu oleh FMA, Glomus etunicatum ataupun dengan bakteri pelarut fosfat PGPR, Enterobacter agglomerans. Akan tetapi penyerapan P dan N tertinggi diperoleh ketika tanaman tomat diinokulasi dengan kedua mikroorganisme tersebut (FMA dan PGPB). Camprubi et al. (1996) menyimpulkan bahwa Trichoderma aureoviride Rifai yang diinokulasi dengan FMA Glomus intraradices meningkatkan pertumbuhan Citrus reshni dibandingkan tanaman yang hanya diinokulasi oleh G. intraradicessaja.
Salah satu alasan utama kurang efektifnya pengendalian hayati penyakit di bidang pertanian adalah tidak konsistennya hasil serta kurangnya aktivitas antagonis pengendalian hayati yang diaplikasikan secara tunggal di lapangan. Untuk itu, kombinasi beberapa agen pengendalian hayati dengan peranannya masing-masing terhadap tanaman dapat menjadi alternatif yang menjanjikan dalam manajemen hama terpadu di masa yang akan datang. Kombinasi FMA
dengan bakteri endosimbiotik mikoriza yang berasosiasi dengan akar tanaman kelapa sawit merupakan suatu alternatif solusi yang tepat sebagai pengendali hayati terhadap serangan patogen akar. Untuk itu perlu dilakukan suatu penelitian untuk melihat bagaimana simbiosis FMA dan bakteri endosimbiotik mikoriza dengan tanaman inang dapat menngkatkan daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense.
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk:
1. Mengeksplorasi dan mengidentifikasi isolat fungi mikoriza arbuskular (FMA) dan bakteri endosimbiotik mikoriza yang diisolasi dari rizosfir kelapa sawit yang berpotensi meningkatkan daya adaptasi tanaman kelapa sawit terhadap cekaman biotik G. boninensepenyebab penyakit busuk pangkal batang. 2. Mengisolasi dan mengidentifikasi bakteri endosimbiotik mikoriza yang
bekerja sinergis dengan FMA dan bersifat antagonis terhadap G. boninense 3. Mengidentifikasi senyawa aktif yang dikeluarkan oleh bakteri endosimbiotik
mikoriza in vitro yang memiliki kemampuan menghambat pertumbuhan G. boninensepada ujiin vitro.
4. Menguji keefektifan dual inokulasi isolat FMA dan bakteri endosimbiotik mikoriza dalam menginduksi daya adaptasi bibit kelapa sawit terhadap cekaman biotik G. boninense.
5. Menganalisis respon bibit kelapa sawit (uji in vivo) dalam bentuk profil senyawa yang dihasilkan oleh bibit kelapa sawit yang diinduki oleh inokulasi FMA dan bakteri endosimbiotik mikoriza sebagai ekspresi daya adaptasi bibit kelapa sawit terhadap cekaman biotik G. boninense.
Perumusan Masalah
Penyakit busuk pangkal batang (BPB) yang disebabkan oleh fungi Ganoderma boninense pada tanaman kelapa sawit termasuk penyakit yang mematikan. Serangan penyakit BPB ini pada perkebunan kelapa sawit
7
mengurangi populasi kelapa sawit sehingga menurunkan produksinya. Cara pengendalian yang efektif dan efisien sampai saat ini belum ditemukan sehingga para pengusaha kebun kelapa sawit masih berusaha mencari alternatif pengendalian yang efektif. Oleh karena itu perlu dilakukan pengembangan teknik pencegahan dini dengan memanfaatkan mikroba tanah yang memiliki potensi dalam mengurangi serangan G. boninense pada kelapa sawit. Hingga saat ini belum ada populasi kelapa sawit yang tahan terhadap serangan G. boninense, akan tetapi di areal pertanaman yang terserang berat penyakit tersebut ada beberapa tanaman yang tetap sehat seakan-akan mengekspresikan tahan terhadap G. boninense. Di duga pada populasi tanaman tersebut terbentuk sistem ketahanan karena adanya induksi yang dipicu oleh keberadaan komunitas mikroba tanah di daerah rizosfir tanaman kelapa sawit.
Inokulasi beberapa jenis mikroba tanah seperti fungi mikoriza arbuskular (FMA) dan bakteri endosimbiotik mikoriza diketahui mampu meningkatkan toleransi tanaman terhadap serangan penyakit. Eksplorasi dan identifikasi FMA dan bakteri endosimbiotik mikoriza dari rizosfir kelapa sawit sehat di daerah serangan Ganoderma belum dilakukan, padahal sering ditemukan tanaman yang mampu bertahan hidup yang diduga diakibatkan oleh adanya mikoriza arbuskular yang berkembang secara alami. Inokulasi FMA dan bakteri endosimbiotik mikoriza yang mampu meningkatkan efisiensi penyerapan unsur hara dari tanah, meningkatkan lignifikasi tanaman, meningkatkan aktivitas enzim kitinase, β-1,3 glukanase, β-1,4 glukosidase, peroksidase, serta meningkatkan konsentrasi senyawa golongan fitoaleksin dapat dimanfaatkan untuk menghambat serangan G. boninensepenyebab penyakit busuk pangkal batang pada tanaman kelapa sawit.
Evaluasi pengaruh FMA dan bakteri endosimbiotik mikoriza terhadap kejadian dan perkembangan penyakit, harus memperhitungkan beberapa faktor, seperti patogen tanaman, asosiasi fungi dan bakteri endosimbiotik mikoriza dengan kondisi lingkungan. Oleh karena interaksi terjadi antara patogen, simbion dan bakteri endosimbiotik mikoriza, maka harus dipahami hubungan antara FMA dengan bakteri endosimbiotik mikoriza, FMA dengan tanaman dan penyakit dengan tanaman, sehingga akan diperoleh pengendalian yang efektif.
Hipotesis
Berdasarkan permasalahan yang telah dikemukan di atas, rumusan hipotesis penelitian ini adalah sebagai berikut:
1. Pada rizosfir kelapa sawit terdapat isolat FMA dan bakteri endosimbiotik mikoriza yang berpotensi menginduksi daya adaptasi tanaman kelapa sawit terhadap cekaman biotik Ganoderma boninense.
2. Pada spora FMA terdapat isolat bakteri endosimbiotik mikoriza yang mampu menghambat pertumbuhanG. boninense in vitro.
3. Terdapat senyawa aktif yang dikeluarkan oleh bakteri endosimbiotik mikoriza secara in vitro yang dapat menghambat pertumbuhan patogen G. boninense in vitro.
4. Kombinasi inokulasi FMA dan bakteri endosimbiotik mikoriza meningkatkan daya adaptasi kelapa sawit terhadap cekaman biotik patogenG. boninense. 5. Interaksi FMA dan bakteri endosimbiotik mikoriza dapat meningkatkan
keefektifan pengendalian hayati terhadap penyakit busuk pangkal batang pada tanaman kelapa sawit yang dicirikan oleh ekspresi profil senyawa aktif dihasilkan secara in vivooleh bibit kelapa sawit yang diinduksi oleh inokulasi FMA dan bakteri endosimbiotik mikoriza.
Strategi Penelitian
Untuk menjawab hipotesis-hipotesis pada penelitian ini dilakukan lima percobaan yang saling berkaitan seperti tercantum dalam Diagram Alir Penelitian. Topik Penelitian 1 bertujuan untuk mendapatkan isolat FMA dan bakteri endosimbiotik mikoriza dari rizosfir kelapa sawit dengan judul “Eksplorasi, isolasi dan identifikasi FMA dan bakteri endosimbiotik mikoriza dari rizosfir kelapa sawit dan potensinya sebagai penginduksi ketahanan kelapa sawit terhadap cekaman biotik Ganoderma boninense”. Isolat bakteri endosimbiotik mikoriza yang diperoleh kemudian diseleksi kemampuannya dalam mempercepat perkecambahan spora FMA dan menghambat pertumbuhan patogen G. boninense secara in vitro pada Penelitian 2 yang berjudul “Seleksi bakteri endosimbiotik mikoriza terhadap daya kecambah FMA dan daya hambatnya terhadap Ganoderma boninense”. Isolat bakteri endosimbiotik mikoriza yang memiliki
9
kemampuan menghambat pertumbuhan G. boninense dan memiliki kemampuan mempercepat perkecambahan spora FMA dipilih untuk mendapatkan dan mengidentifikasi senyawa aktif yang dikeluarkan oleh bakteri tersebut yang dilakkan pada Penelitian 3 yang berjudul “Identifikasi senyawa aktif dari bakteri endosimbiotik mikoriza yang menghambat pertumbuhan G. boninense dan bersifat sinergis dengan FMA”. Isolat FMA dan bakteri endosimbiotik mikoriza yang diperoleh pada Penelitian 1 kemudian diuji kemampuannya dalam meningkatkan daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense yang dilakukan pada Penelitian 4 yang berjudul “Daya Adaptasi bibit kelapa sawit diinokulasi FMA dan bakteri endosimbiotik mikoriza terhadap cekaman biotik patogen G. boninense”. Akar kelapa sawit yang telah diinokulasi FMA dan bakteri endosimbiotik mikoriza kemudian dianalisis untuk melihat profil kromatogram senyawa yang dikeluarkan oleh dual inokulasi tersebut yang mampu menghambat pertumbuhan G. boninense yang dilakukan pada Penelitian 5 dengan judul “Analisis respon inang dalam bentuk senyawa aktif sebagai ekspresi daya adaptasi kelapa sawit terhadap cekaman biotik G. boninense yang diinokulasi FMA dan bakteri endosimbiotik mikoriza”.
Manfaat Penelitian
Penelitian yang dilakukan ini secara ilmiah diharapkan dapat memperoleh isolat fungi mikoriza dan bakteri endosimbiotik mikoriza bekerja secara sinergis untuk meningkatkan daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense berupa peningkatan serapan hara, pertumbuhan dan ketahanan terhadap penyakit. Dari penelitian ini juga diharapkan dapat mengetahui senyawa yang dikeluarkan oleh bakteri endosimbiotik mikoriza yang bermanfaat dalam meningkatkan daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense serta mendapatkan informasi profil kromatogram senyawa aktif dari akar kelapa sawit yang telah diinokulasi FMA dan bakteri endosimbiotik mikoriza sebagai ekspresi daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen G. boninense. Pemahaman ini diharapkan dapat menjadi landasan dalam mengendalikan penyakit busuk pangkal batang pada kelapa sawit yang efektif.
?
DIAGRAM ALIR PENELITIAN
= Hasil penelitian 3 belum dapat mengkonfirmasikan apakah senyawa tersebut juga dihasilkan oleh bibit kelapa sawit secara in vivo dengan inokulasi FMA dan bakteri endosimbiotik mikoriza, sehingga perlu penelitian lebih lanjut.
Penelitian 1
Eksplorasi, Isolasi dan Identifikasi Fungi Mikoriza Arbuskular dan Bakteri Endosimbiotik Mikoriza dari Rizosfir Kelapa Sawit
Hasil yang Diharapkan: Diperolehnya isolat FMA dan bakteri endosimbiotik mikoriza dari beberapa varietas kelapa sawit yang berpotensi meningkatkan daya adaptasi kelapa sawit terhadap cekaman biotik G. boninense.
Penelitian 2
Seleksi Bakteri Endosimbiotik Mikoriza yang Mempercepat Perkecambahan
Spora FMA dan Menghambat
PertumbuhanG.boninense
Hasil yang Diharapkan: Diperoleh isolat bakteri endosimbiotik mikoriza yang efektif dalam meningkatkan persentase
berkecambah spora FMA dan
menghambat pertumbuhanG. boninense
Penelitian 3
Identifikasi Senyawa Aktif dari Bakteri Endosimbiotik Mikoriza yang
Menghambat PertumbuhanG.boninense
dan Bersifat Sinergis dengan FMA
Hasil yang Diharapkan: Diperoleh senyawa aktif dari bakteri endosimbiotik mikoriza
yang menghambat pertumbuhan G.
boninense in vitro
Penelitian 4
Daya Adaptasi Bibit Kelapa Sawit Diinokulasi Fungi Mikoriza Arbuskular dan Bakteri
Endosimbiotik Mikoriza terhadap Cekaman Biotik Patogen G. boninense
Hasil yang Diharapkan: Diperoleh kombinasi isolat FMA dan bakteri endosimbiotik mikoriza yang efektif dalam meningkatkan daya adaptasi kelapa sawit terhadap cekaman biotik G. boninense.
Penelitian 5
Menganalisis Respon Inang dalam Bentuk Senyawa Aktif sebagai Ekspresi Daya Adaptasi
Kelapa Sawit terhadap Cekaman Biotik G. boninensedengan Inokulasi FMA dan Bakteri
Endosimbiotik Mikoriza
Hasil yang Diharapkan: Diperoleh profil kromatogram senyawa aktif yang dihasilkan oleh
bibit kelapa sawit sebagai ekspresi daya adaptasi kelapa sawit terhadap cekaman biotik G.
boninensedengan adanya inokulasi FMA dan bakteri endosimbiotik mikoriza
Hasil Akhir:
Diperoleh gambaran tentang daya adaptasi bibit kelapa sawit terhadap cekaman biotik patogen Ganoderma boninensedengan inokulasi FMA & bakteri
endosimbiotik mikoriza
11
TINJAUAN PUSTAKA
Tanaman Kelapa Sawit
Arti Penting Tanaman Kelapa Sawit
Tanaman kelapa sawit yang menghasilkan minyak sawit merupakan
tanaman tropis yang berasal dari Afrika Barat yang tumbuh sebagai tanaman
hibrida di berbagai tempat di dunia, termasuk Asia Tenggara dan Amerika Tengah.
Walaupun ditanam di luar daerah asalnya, tanaman kelapa sawit dapat tumbuh
dan berkembang dengan baik termasuk di Indonesia. Minyak sawit dengan harga
yang relatif lebih rendah dibandingkan dengan minyak nabati lainnya ini
digunakan untuk berbagai keperluan industri seperti industri minyak goreng,
industri sabun, kosmetik, lilin, makanan dan untuk bahan baku biodiesel.
Negara-negara di Asia Tenggara merupakan penghasil minyak sawit
terbesar di dunia termasuk Indonesia. Total produksi minyak sawit dunia
meningkat hampir tiga kali lipat selama tiga dasawarsa hingga tahun 2009. Pada
tahun 2009/2010, total produksi minyak sawit diperkirakan 45,1 juta ton, dengan
Indonesia dan Malaysia mencapai lebih dari 85 persen total dunia. Indonesia dan
Malaysia masing-masing memproduksi lebih dari 18 juta ton minyak sawit.
Minyak sawit produksi Indonesia terutama diekspor ke India, Cina dan Eropa
Barat masing-masing 6,7 juta ton, 6,3 juta ton dan 4,6 juta ton (World Growth
2011). Berdasarkan prospek ekonominya yang besar, industri minyak sawit ini
menjadi subsektor yang paling dinamis dan diminati, sehingga perkembanganya
menarik banyak perhatian pelaku bisnis.
Sejarah kelapa sawit di Indonesia dimulai tahun 1915 ketika turunan
kelapa sawit hasil introduksi yang berada di Kebun Raya Bogor ditanam di
Sumatera Utara (Lubis 1992). Di daerah tersebut, kelapa sawit kemudian
berkembang dan selanjutnya dibudidayakan secara komersial. Sejak dua dekade
terakhir terjadi pengembangan areal kelapa sawit yang sangat pesat.
Pengembangan kelapa sawit tidak hanya di Sumatera (69%), tetapi meluas sampai
di Kalimantan (26%), Sulawesi (3%), Papua (1%), dan Jawa (1%) (Tryfino 2006).
Perluasan ini tidak hanya membuka hutan baru tetapi termasuk juga konversi dari
merupakan tanaman komoditas perkebunan yang penting di Indonesia sebagai
penghasil minyak nabati beserta beberapa produk turunan lainnya. Komoditas
kelapa sawit, baik berupa bahan mentah maupun hasil olahannya menduduki
peringkat ketiga penyumbang devisa nonmigas terbesar bagi negara setelah karet
dan kopi (Sastrosayono 2003).
Pada saat Indonesia mengalami krisis ekonomi, industri kelapa sawit
merupakan salah satu agroindustri andalan yang menghasilkan devisa bagi negara.
Perkembangan industri kelapa sawit pada dekade terakhir ini berkembang sangat
pesat sehingga menempatkan Indonesia sebagai produsen minyak kelapa sawit
terbesar kedua di dunia setelah Malaysia. Setelah tahun 2005, produksi minyak
sawit dunia diharapkan menjadi penyumbang terbesar dalam produksi minyak
sayur dunia menggantikan minyak kedelai. Disamping itu, krisis energi yang
melanda dunia membuat orang berusaha untuk mencari energi alternatif yang
dapat diperbaharui (renewable energy) menggunakan bahan baku minyak sawit
mentah (crude palm oil) yang direaksikan secara kimiawi untuk memenuhi
spesifikasi teknis sebagai bahan bakar nabati (biodiesel). Potensi minyak sawit
sebagai salah satu bahan baku biodiesel menggantikan bahan bakar minyak bumi
atau fosil membuat permintaan akan minyak sawit dunia semakin tinggi.
Faktor Lingkungan Tumbuh Tanaman Kelapa Sawit
Tanaman kelapa sawit merupakan tipikal tanaman tropis yang dapat
tumbuh dengan baik pada ketinggian kurang dari 400 m di atas permukaan laut
(dpl) dengan kemiringan lereng antara 0- 8%. Topografi datar dan berombak
sampai bergelombang. Suhu udara optimum adalah 27oC dengan rentang suhu
22oC - 33oC sepanjang tahun. Rentang curah hujan ratarata tahunan antara 1250
-3000 mm/tahun dengan curah hujan yang optimal 1750 - 2500 mm/tahun. Lama
penyinaran matahari rata-rata 5 - 7 jam/hari. Kecepatan angin 5-6 km/jam untuk
membantu proses penyerbukan. Tanah yang baik sebagai media tanam
mengandung banyak lempung, beraerasi baik dan subur. berdrainase baik,
permukaan air tanah cukup dalam, solum cukup dalam (80 cm), pH tanah 4 - 6,
dan tanah tidak berbatu. Secara umum kelapa sawit dapat tumbuh dan berproduksi
baik pada jenis-jenis tanah ultisols, entisols, inceptisols, andisols dan histosols
13
Dalam bertanam kelapa sawit, pola tanam dapat monokultur ataupun
tumpangsari. Pada masa tanaman kelapa sawit belum menghasilkan (0 - 3 tahun),
kanopi dan perakaran tanaman masih relatif belum berkembang. Sebagian besar
lahan tersebut akan terbuka dan memperoleh cahaya matahari secara penuh
sehingga dapat dimanfaatkan untuk budidaya tanaman sela dalam pola
tumpangsari. Pola ini memungkinkan pendapatan tambahan bagi petani selama
tanaman kelapa sawit belum berproduksi. Ketika tajuk belum saling menutup,
kelapa sawit dapat ditumpang sari dengan segala jenis tanaman pangan seperti
kedelai atau tanaman buah-buahan seperti nanas. Tetapi jika tajuk telah saling
menutup, hanya tanaman yang naungannya sedikit yang dapat ditanam diantara
barisan tanaman kelapa sawit (PPKS 2007). Tanaman penutup tanah (legume
cover crop) seperti tanaman kacang-kacangan pada areal tanaman kelapa sawit
sangat penting karena dapat memperbaiki sifat-sifat fisika, kimia dan biologi
tanah, mencegah erosi, mempertahankan kelembaban tanah dan menekan
pertumbuhan tanaman pengganggu (gulma). Penanaman tanaman
kacang-kacangan sebaiknya dilaksanakan segera setelah persiapan lahan selesai.
Tanah tropis kekurangan unsur hara nitrogen (N), fosfat (P) dan kalium
(K) sehingga ketiga unsur hara tersebut harus ditambah melalui pemupukan
anorganik yang terdiri dari 1,3 kg N; 0,2 kg P dan 1,8 kg K untuk setiap tanaman
selama satu tahun. Kekurangan unsur N, P, K dan Mg menghambat pertumbuhan
kelapa sawit sehingga tanaman menjadi kerdil, sementara kekurangan boron (B)
pada tanaman muda dapat mematikan tanaman. Hara K berperan dalam aktifitas
pembukaan dan penutupan stomata, aktifitas enzim dan sintesa minyak dan
meningkatkan ketahanan terhadap penyakit. Kekurangan K menyebabkan bercak
kuning/transparan, white stripe, daun tua kering dan mati. Kekurangan K
berasosiasi dengan munculnya penyakit seperti Ganoderma(Liang 2008).
Peremajaan kebun kelapa sawit biasanya dilakukan setelah umur tanaman
mencapai 25 tahun (Arifin et al. 2000). Beberapa perkebunan kelapa sawit di
Indonesia telah berumur 3-4 generasi. Pada kondisi tersebut kondisi tanah sudah
kurang mampu mendukung usaha yang efisien karena telah mengalami degradasi
sehingga kandungan unsur hara yang ada pada tanah tidak lagi mencukupi untuk
terus menerus pada beberapa generasi tersebut. Pemakaian bahan kimia dalam
bentuk pupuk dan pestisida yang berlebih secara terus menerus juga dapat
menyebabkan punahnya atau tidak berkembangnya biota tanah yang
menguntungkan bagi tanaman. Dengan kondisi tanah yang miskin unsur hara,
tentu tidaklah mudah untuk melaksanakan budidaya kelapa sawit karena
banyaknya masalah yang akan muncul seperti pertumbuhan tanaman yang
terhambat akibat kekurangan hara atau hara terbatas sehingga tanaman tidak dapat
tumbuh dengan baik dan menghasilkan produktivitas yang maksimal. Pada tanah
yang miskin unsur hara akan membuat tanaman kelapa sawit menjadi rentan
terhadap serangan penyakit sehingga tanaman menjadi sakit bahkan mati
(Darmono 2000).
Pengaruh Faktor Abiotik terhadap Pertumbuhan Kelapa Sawit
Kerusakan pada tanaman seringkali tidak hanya disebabkan oleh adanya
serangan hama dan penyakit. Tidak jarang kematian tanaman disebabkan oleh
faktor-faktor lingkungan seperti kelebihan atau kekurangan air, ketinggian yang
ekstrim, pH tanah yang tidak sesuai, suhu yang terlalu ekstrim serta kelebihan
atau kekurangan unsur hara mikro. Tanaman kelapa sawit memiliki perakaran
yang dangkal sehingga mudah mengalami cekaman kekeringan yang dapat
menurunkan pertumbuhan dan produksi. Cekaman kekeringan yang berlangsung
lama dapat menghambat pembukaan pelepah daun muda, daun bagian bawah
cepat mengering, merusak hijau daun, tandan buah mengering dan patah pucuk,
bahkan tanaman mati jika kondisi ekstrim kering terjadi (Caliman & Southworth
1998). Pada fase reproduktif cekaman kekeringan menyebabkan perubahan nisbah
kelamin bunga, bunga dan buah muda gugur, dan tandan buah gagal masak
(Caliman & Southworth, 1998), sehingga menurunkan produksi tandan buah segar
10 % – 40 % dan minyak sawit 21 % – 65 % (Subronto et al.2000).
Hasil kajian yang dilakukan oleh Pusat Penelitian Kelapa Sawit (PPKS)
menunjukkan bahwa ketinggian tempat (altitude) berpengaruh terhadap
pertumbuhan vegetatif kelapa sawit, baik pada tanaman menghasilkan (TM)
maupun tanaman belum menghasilkan (TBM). Pada tahap awal, terlihat adanya
perbedaan panjang rachis tanaman pada berbagai ketinggian tempat yang
15
menunjukkan bahwa rachis pelepah kelapa sawit pada altitude 0 - 250 m dpl (di
atas permukaan laut) nyata lebih panjang dibandingkan dengan rachis pelepah
kelapa sawit pada altitude 251 - 500 m dpl, 501 - 750 m dpl dan 751 - 1000 m dpl.
Untuk kelapa sawit tanaman yang telah menghasilkan (TM), panjang rachis tidak
lagi menunjukkan perbedaan nyata sebagai akibat pertumbuhan tanaman yang
sudah stabil (PPKS 2007).
Kemasaman (pH) tanah mempengaruhi pertumbuhan kelapa sawit. Tanah
mineral masam di daerah tropika yang tidak subur merupakan faktor pembatas
utama terhadap pertumbuhan dan produksi tanaman kelapa sawit. Untuk
menyatakan ketidaksuburan tanah ini umumnya dapat diamati dari adanya
masalah defisiensi unsur hara, terutama fosfat yang disebabkan besarnya jumlah
fosfat yang terfiksasi di permukaan koloid-koloid liat. Diantara beberapa kendala
yang ada pada tanah ultisol, kekahatan P merupakan kendala yang penting dan
utama, Kekahatan P tidak hanya disebabkan oleh kandungan P tanah yang rendah
akan tetapi juga karena sebagian besar P terikat oleh unsur-unsur logam seperti Al
dan Fe sehingga P tidak tersedia di dalam tanah untuk pertumbuhan tanaman.
Kendala lain yang tidak kalah pentingnya adalah rendahnya kandungan bahan
organik dan muatan-muatan negatif yang rendah pada tanah ultisol. Usaha-usaha
yang dilakukan untuk meningkatkan ketersediaan P tanah ultisol adalah dengan
cara merubah bentuk P dari bentuk P yang terikat pada fase padat menjadi bentuk
P yang dapat tersedia di dalam tanah dengan cara menaikkan pH tanah,
mineralisasi bahan-bahan organik yang menghasilkan asam-asam organik serta
memanfaatkan mikroba tanah (Amiruddin 2008).
Faktor-faktor abiotik secara tidak langsung juga akan mempengaruhi
kemampuan daya adaptasi tanaman terhadap cekaman biotik patogen. Kondisi
tercekam karena faktor-faktor abiotik seperti cekaman kekeringan, keracunan
logam berat, kemasaman tanah yang tinggi, akan menghambat pertumbuhan
tanaman sehingga tanaman menjadi tidak sehat dan rentan terhadap serangan
penyakit. Pengelolaan faktor-faktor abiotik pada budidaya kelapa sawit perlu
dilakukan agar tanaman memiliki daya adaptasi yang lebih tinggi pada saat
Penyakit Busuk Pangkal Batang pada Kelapa Sawit
Salah satu hambatan utama dalam budidaya kelapa sawit ialah adanya
serangan patogen. Di antara penyakit yang ada pada tanaman kelapa sawit,
penyakit busuk pangkal