www.elsevier.com/locate/ibmb
Stress-reactivity and juvenile hormone degradation in
Drosophila
melanogaster
strains having stress-related mutations
N.E. Gruntenko
a,*, T.G. Wilson
b, M. Monastirioti
c, I.Y. Rauschenbach
aaInstitute of Cytology and Genetics, Russian Academy of Sciences, Siberian Division, Novosibirsk 630090, Russia bDepartment of Biology, Colorado State University, Fort Collins, CO 80523, USA
cInstitute of Molecular Biology and Biotechnology, FORTH, 711-10 Heraklion, Crete, Greece
Received 31 October 1999; received in revised form 31 December 1999; accepted 25 January 2000
Abstract
Juvenile hormone (JH) degradation was studied under normal and stress conditions in young and matured females ofDrosophila melanogaster strains having mutations in different genes involved in responses to stress It was shown that (1) the impairment in heat shock response elicits an alteration in stress-reactivity of the JH system; (2) the impairment JH reception causes a decrease of JH-hydrolysing activity and of stress-reactivity in young females, while in mature ones stress reactivity is completely absent; (3) the absence of octopamine results in higher JH-hydrolysis level under normal conditions and altered JH stress-reactivity; (4) the higher dopamine content elicits a dramatic decrease of JH degradation under normal conditions and of JH stress-reactivity. Thus, the impairments in any component of the Drosophila stress reaction result in changes in the reponse of JH degradation system to stress. The role of JH in the development of the insect stress reaction is discussed. 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Drosophila melanogaster;ts403;Met;Tbh;ebony; Juvenile hormone; Stress reactivity
1. Introduction
Juvenile hormone (JH), a sesquiterpenoid involved in the regulation of developmental transitions and repro-duction in insects (reviews: Riddiford and Ashburner, 1991; Nijhout, 1994; Wyatt and Davey, 1996), is well known to play a main role in the development of the insect stress reaction (reviews: Cymborowski, 1991 Rau-schenbach 1991, 1997). Two other important compo-nents of this multi-faceted response are the metabolism of biogenic amines, dopamine (DA) and octopamine (OA), and the heat shock response (HSR) (Orchard and Loughton, 1981; Davenport and Evans, 1984; Woodring et al., 1989; Hirashima et al., 1993, 1999; Rauschenbach et al. 1993, 1997; Rauschenbach, 1997; Sukhanova et al., 1997; Khlebodarova et al., 1998).
We have previously shown that the JH metabolic
sys-* Corresponding author. Tel.: +383-2-333-526; fax: + 383-2-331-278.
E-mail address:[email protected] (N.E. Gruntenko).
0965-1748/00/$ - see front matter2000 Elsevier Science Ltd. All rights reserved. PII: S 0 9 6 5 - 1 7 4 8 ( 0 0 ) 0 0 0 4 9 - 7
tem of wild type females of Drosophila melanogaster andD. virilisresponds to stress conditions (termed here stressors) with a decrease in JH-hydrolysing activity. Males do not respond to stressors in this manner (Rauschenbach et al. 1995, 1996). The metabolic sys-tems of DA and OA respond to stress, in both sexes, by an increase in the amine content and by a decrease in the activity of their synthetic enzymes (Rauschenbach et al., 1993; Hirashima et al., 1999). We have also demon-strated that a mutation disturbing the development of the stress reaction inD. virilisalso elicits the impairment of HSR (Khlebodarova et al., 1998).
In this work, we analysed the mutations ts403, Met, Tbh andebony (e) with respect to the response of JH-degradation system to stress. The recessive temperature sensitive lethal mutation l(l)ts403 results in the failure of heat shock protein (HSP)83 and HSP35 to be expressed, and a number of HSP70 proteins are only par-tially expressed (Evgen’ev and Denisenko, 1990). Met27
is a null allele of the Methoprene-tolerant gene that shows resistance to the toxic effects of both JH and a JH analog, methoprene. The mechanism of the resistance appears to be altered JH reception(Wilson and Fabian, 1986; Shemshedini and Wilson, 1990).Met27completely
lacks Met transcript and is clearly a null allele (Wilson and Ashok, 1998).TbhnM18is a null mutation at the Tyr-amine b-hydroxylase locus, which results in complete absence of the tyramine β-hydroxylase protein and blockage of octopamine biosynthesis (Monastirioti et al., 1996). eis postulated to be the mutation of N-b-alanyl dopamine synthetase gene, based on the fact that e has twice as much DA as normal (Hodgetts, 1972; Hodgetts and Konopka, 1973; Ramadan et al., 1993).
Here we asked whether these mutations would affect the decrease in JH degradation occurring in D. mel-anogaster when stressed. In order to answer this ques-tion, we studied the JH degradation in individuals of ts403,nMet27, TbhnM18andStestrains (carringl(l)ts403, Met27, Tbhandemutations, respectively), under normal and stress conditions, and compared their stress-reac-tivity (calculated as percent change in JH hydrolysis under stress compared to hydrolysis under normal conditions) with that in a number of wild type and lab-oratory strains.
We demonstrated (1) that ts403 females respond to stress by a decrease in JH degradation, as occurs in wild type females, but that their stress-reactivity significantly differs from that of wild type; (2) that in youngv Met27 females, similar to wild type flies, JH hydrolysis is decreased upon stress, but their stress-reactivity is sig-nificantly lower than in wild type; (3) that JH degra-dation is unaffected in older v Met27 females under stress; (4) that TbhnM18 females show a significantly higher JH-hydrolysis level and different stress-reactivity than does the wild type; and (5) that youngSte females demonstrate significantly lower JH-hydrolysis and stress-reactivity, compared to the wild type.
2. Materials and methods
2.1. Drosophila strains
The followingD. melanogasterstrains were used: the wild type laboratory strain Canton S; wild type iso-female strain 921500 from a natural population of Gorno-Altaisk; laboratory balancer strainFirst Multiple Seven (FM7); vermilion (n) strain from which the n
Met27 strain was derived; laboratory balancer strain
In(2LR)Cy/L; In(3LR)D/Sb, carrying morphological mutations with recessive lethal action Curly, Lobe (chromosome 2) andDichaete, Stubble(chromosome 3; hereafter termed CyLDSb); strain ts403 carrying the recessive temperature sensitive lethal mutationl(l)ts403 (Arking, 1975); strain n Met27 carrying a null allele of the Methoprene-tolerant gene (Wilson and Ashok, 1998); strainTbhnM18carrying a null mutation at the Tyr-amine b-hydroxylase locus (Monastirioti et al., 1996); and the laboratory Ste strain carrying the e mutation. Cultures were raised on standard medium (Rauschenbach et al., 1987) at 25°C, and adults were synchronized by eclosion. Flies were subjected to stress at 38°C for 3 h, and were subsequently frozen in liquid nitrogen and stored at220°C.
2.2. JH hydrolysis
JH hydrolysis was measured by the assay of Ham-mock and Sparks, 1977. A fly was homogenized on ice in 30µl of 0.1 M Na-phosphate buffer, pH 7.4, contain-ing 0.5 mM phenylthiourea. The homogenates were cen-trifuged for 5 min at 12,000 rpm, and samples of the supernatant (10µl) were utilized for the reaction. A mix-ture consisting of 0.1 µg unlabeled JH-III (Sigma) and 12,500 dpm 3H labeled JH-III (17.4 Ci/mmol at C-10,
NEN Research Products, Germany) was used as sub-strate. The reaction was carried out in siliconized tubes in 100 µl of incubation mixture for 3 h, and it was stopped by the addition of 250µl heptane and 50 µl of a solution containing 5% ammonia and 50% methanol (V/V). The tubes were shaken vigorously and centri-fuged at 12,000 rpm for 10 min. Samples (100 µl) of both aqueous and heptane phases were placed in vials containing dioxane scintillation fluid and counted. Con-trol experiments have shown a linear substrate–reaction relationship (Gruntenko et al., 1999), as well as the fact that measured activity is proportional to homogenate (i.e. enzyme) concentration (Rauschenbach, 1991; unpub-lished data).
The significance of the differences between the data sets was tested by Student’s t-test. Sample size varied from 12 to 28 individuals for each measurement in all experiments.
3. Results
3.1. JH degradation in 1-day old ts403 and Canton S females under normal and heat stress conditions
Fig. 1. Hydrolysis of [3H]JH-III in 1-day-old females ofCanton S
andts403strains ofD. melanogasterunder normal and stress (38°C, 3 h) conditions. Means±SE.
does not differ from that in Canton Sones. The data of Fig. 1 also demonstrate that ts403 females respond to stress as well as Canton Sdo: exposure to 38°C evokes in females of both strains a significant (P,0.001) decrease in JH-hydrolysing activity compared to control females maintained at 25°C.
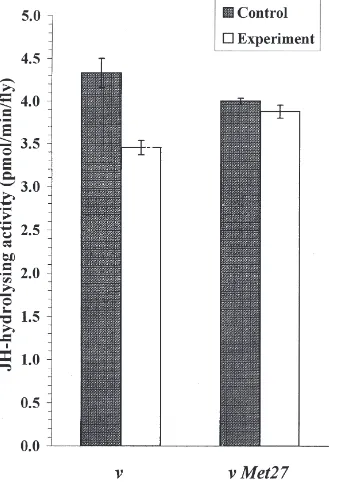
3.2. JH degradation in 1-day old n Met27 andn females under normal conditions and under heat stress
JH-hydrolysis in 1-day old females of both n and n
Met27 strains under normal and stress conditions are shown in Fig. 2. They indicate that under normal con-ditions n Met27 females show a significantly (P
,0.01)
Fig. 2. Hydrolysis of [3H]JH-III in 1-day-old females of n and n Met27strains ofD. melanogasterunder normal and stress (38°C 3 h)
conditions. Means±SE.
lower JH-hydrolysing activity than do n females. Exposure to 38°C causes females of both n and Met27 strains to show a significant (P,0.001) decrease in JH-hydrolysis level, compared to control females kept at 25°C.
3.3. JH degradation in 1-day-old TbhnM18 and Ste females under normal conditions and under heat stress
The levels of JH-hydrolysing activity in 1-day-old females of TbhnM18 and Ste strains under normal and stress conditions are shown in Fig. 3, together with that of Canton S. The data reveal that under normal con-ditions, the level of JH degradation in females ofTbhnM18 strain is significantly higher than that in Canton S (P,0.001). In contrast,Stefemales are distinguished by a lower level of JH-hydrolysing activity compared to Canton S (P,0.001). The data in Fig. 3 also show that TbhnM18 and
Ste females respond to heat stress as do Canton S: exposure to 38°C elicits in females of all three strains a decrease in the level of JH degradation com-pared to control females (P,0.001).
Fig. 3. Hydrolysis of [3H]JH-III in 1-day-old females ofCanton S, TbhnM18andSte strains ofD. melanogasterunder normal and stress
3.4. JH degradation under normal and stress conditions in l-day-old females of wild type (both Canton S and strain 921500) and laboratory (FM7, n
and CyLDSb) strains
It can be seen in Fig. 4 that 1-day-old females of Can-ton S strain are characterized by a level of JH degra-dation similar to that of the iso-female strain921500and of laboratory strains FM7, n and CyLDSb, which have no mutations relating with any components of the stress reaction (differences betweenCanton Sand other strains are insignificant). Heat treatment of females of all these strains results in a significant (P,0.001) lowering of the level of JH degradation (compared to control females kept at 25°C).
3.5. JH degradation in 6-day-old Canton S and ts403 females under normal conditions and under heat stress
Since JH degradation can control Drosophila repro-duction under normal and heat stress conditions (Rauschenbach et al., 1996), we further measured the level of JH-hydrolysing activity in 6-day-old females of Canton S and ts403 strains. As seen in Fig. 5, under normal conditions the JH-hydrolysing activity in mature ts403 females does not differ from that of Canton S. Females of both strains show lower JH degradation (0.01) after heat stress (38°C, 3 h).
3.6. JH degradation under normal and stress conditions in 5-day-old n Met27and nfemales
Under normal conditions, the level of JH degradation in 5-day-old n Met27 females is the same as that in n
Fig. 4. Hydrolysis of [3H]JH-III in l-day-old females of921500,Canton S,FM7,nandCyLDSbstrains ofD. melanogasterunder normal and
stress (38°C, 3 h) conditions. Means±SE.
Fig. 5. Effect of short term heat stress (38°C, 3 h) on JH-hydrolysing activity in 6-day-old females ofts403andCanton Sstrains ofD. mel-anogaster. Means±SE.
females. After heat stress, maturenMet27females show no changes in the level of JH metabolism compared with mature nfemales (Fig. 6) which respond to stress with a significant decrease in JH-hydrolysing activity (P,0.001).
3.7. JH degradation in 6-day old TbhnM18 and Canton S females under normal conditions and under heat stress
Fig. 6. Effect of short term heat stress (38°C 3 h) on JH-hydrolyzing activity in 5-day-old females ofnandnMet27strains ofD. melanogas-ter. Means±SE.
(P,0.001) than that ofCanton S(Fig. 7). It is clear that matureTbhnM18females respond to heat stress by a sharp decrease in JH-hydrolysing activity (P,0.001).
3.8. The stress-reactivity of the JH degradation system in young and mature D. melanogaster females
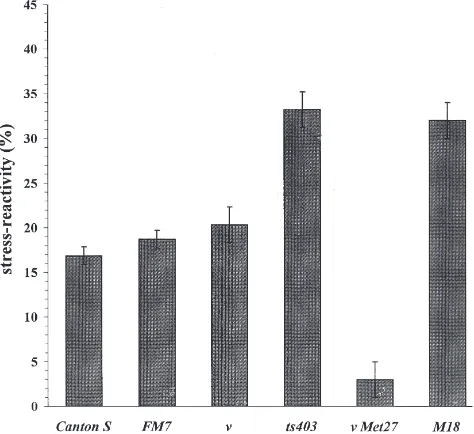
To characterize the stress-reactivity of the JH degra-dation system, we calculated the percent decrease of JH-hydrolysing activity for each stressed female relative to the value under normal conditions (every experiment value was related to the average value for the control group, since it is impossible to determine the JH-hydrolysing activity of the same individual under both control and stress conditions). As seen in Fig. 8, 1-day-old females of wild type (921500 and Canton S) and laboratory (FM7, n and CyLDSb) strains have similar stress-reactivity (the differences between strains are not significant). On the other hand, it is also apparent from the data of Fig. 8, that 1-day-old females having stress-related mutations (ts403,nMet27,TbhnM18andSte) have lower levels of stress-reactivity (P,0.05 for ts403, P,0.01 fornMet27andP,0.001 forTbhnM18andSte). We further analysed the stress-reactivity in mature females (6-day-old Canton S, ts403andTbhnM18strains and 5-day-old FM7, n and n Met27 strains). It is clear from the data in Fig. 9 that the stress-reactivity of mature TbhnM18 and ts403 females is significantly higher than that of wild type (Canton S) and laboratory (FM7 and n) strains (P,0.001 forts403andP,0.05 forTbhnM18). The stress-reactivity of n Met27females is insignificant.
Fig. 7. Effect of short term heat stress (38°C, 3 h) on JH-hydrolysing activity in 6-day-old females ofTbhnM18and Canton Sstrains ofD. melanogaster. Means±SE.
4. Discussion
In adult female insects, JH controls reproduction by regulation of the growth of previtellogenic and/or vitel-logenic follicles, maturation of ovaries, stimulation and maintenance of vitellogenesis, uptake of vitellogenins from hemolymph to oocytes, and oviposition (Shapiro et al., 1986; Roe et al., 1987; Adams and Filipi, 1988; Bownes 1989, 1994; Khlebodarova et al., 1996; Rausch-enbach et al., 1996; Soller et al., 1999). JH must be present at high levels to initiate maturation of ovaries and stimulate vitellogenesis, and then at lower levels to maintain vitellogenesis (Jowett and Postlethwait, 1981; Raikhel and Lea, 1985; Postlethwait and Parker, 1987; Bownes, 1989; Soller et al., 1999).
Fig. 8. Stress-reactivity of JH-degradation system in young females of 921500, Canton S,FM7,n, CyLDSb, ts403,nMet27,TbhnM18andSte
strains ofD. melanogaster. Means±SE.
Fig. 9. Stress-reactivity of JH degradation system in matured females of Canton S,FM7,n,ts403,nMet27andTbhnM18strains ofD. mel-anogaster. Means±SE.
Shapiro et al., 1986; Khlebodarova et al., 1996; Rausch-enbach et al., 1996; Soller et al., 1999). The n Met27 characteristics revealed earlier and in the present study are consistent with this requirement. Indeed,nMet27flies have reduced oogenesis (Wilson and Ashok, 1998) and decreased fertility (Gruntenko et al., 2000) under normal
conditions. It is possible that n Met27 females have an elevated JH level resulting from decreased JH-hydrolys-ing activity (see Figs. 2 and 6) or that the impaired JH reception in this strain may prevent JH-titre-mediated regulation of both the JH degradation system and oogen-esis. In both cases fertility would be disturbed.
the other hand, it is also possible that altered JH metab-olism in these flies is related to a hither to unidentified phenotype/process.
Evidence supporting the regulation of JH metabolism by biogenic amines in Drosophila comes from our data on Ste strain. The flies carrying e mutation are known to have twice as much DA as wild type (Hodgetts, 1972; Hodgetts and Konopka, 1973; Ramadan et al., 1993). Because DA can influence JH secretion (Piulachs and Belles, 1989; Granger et al., 1996) we might expect the JH titre to be affected, with consequent changes in JH-hydrolysing activity in females ofStestrain. Indeed,Ste females have a level of JH degradation almost a half that of Canton S(see Fig. 3).
We cannot exclude the possibility that the differences between Ste,TbhnM18andCanton Sfemales in the level of JH-hydrolysis are the result of strain polymorphism. However, four strains without any stress-related mutations (921500, FM7, n and CyLDSb) were exam-ined and were found to have JH-hydrolysing activity similar to that of Canton S, under normal conditions. Hence, the high JH-hydrolysis level in TbhnM18 females and low level in Ste apparently result from the corre-sponding mutations.
The reason to investigate JH metabolism in the ts403 strain, with its impairment of HSR, was the existing evi-dence on the regulation of the expression of heat shock proteins (hsp) genes by the combined action of the hor-mones JH and 20-OH-ecdysone. Inhibition of the ecdys-teroid peak at pupariation by a temperature shift of the conditionally ecdysteroid-deficient D. melanogaster strain ecd-1 results in a block of hsp26 RNA and a decline in hsp83 RNA level; subsequent addition of exogenous 20-OH-ecdysone restores expression of both genes (Thomas and Lengyel, 1986). JH was reported to inhibit in a dose-dependent manner the ecdysterone induction of the small hsp genes of Drosophila, expressed in cultured cells (Berger et al., 1992). Our data revealed that the block in HSR expression does not affect JH degradation under normal conditions (see Figs. 1 and 5).
How do the mutations in the different components of the D. melanogaster reaction effect the stress-reactivity of the JH degradation system? We have pre-viously demonstrated that exposure of Drosophila females to stress results in a sharp decrease of the JH-hydrolysing activity and as a consequence, the onset of oviposition by young females is delayed 24 h, while mature females cease oviposition for two days (Rauschenbach et al. 1995, 1996). We have also shown that in certainD. virilisandD. melanogasterstrains that do not respond to stress, the level of JH-hydrolysis in females was significantly lower compared to that of wild type. This level does not alter upon heat stress (Rauschenbach et al. 1995, 1996; Gruntenko et al., 1999).
As the present data show, each of the mutations stud-ied elicits some alteration in the stress reactivity of the JH degradation system. Mature females of wild type and laboratory strains without any disturbances in their reaction demonstrate significantly lower stress-reactivity than younger ones (see Figs. 8 and 9). In con-trast, matureTbhnM18females demonstrate higher stress-reactivity than younger ones. Moreover, this response differs from that in wild type: in mature females it is higher (see Fig. 9) and in young ones, lower (see Fig. 8). The stress-reactivity in ts403 females does not change with age in contrast to wild-type (see Figs. 8 and 9). YoungStefemales demonstrate the lowest stress-reactivity (see Fig. 8). YoungnMet27females also have decreased stress-reactivity compared toMet27flies of the same age (see Fig. 8). MaturenMet27females show the most essential differencies from wild type: their stress-reactivtity is insignificant (see Fig. 9).
In summary this work suggests that JH may play the key role in the development of the insect stress-reaction.
Acknowledgements
This study was supported by grants from the Russian Fundamental Research Foundation and the Siberian Branch of the Russian Academy of Sciences for Young Prominent Scientists. Dr. Gruntenko was the recipient of a travel award from the Organizing Committee of the Seventh International Conference on the Juvenile Hor-mones.
References
Adams, T.S., Filipi, P.A., 1988. Interaction between juvenile hormone, 20-hydroxyecdysone, thecorpus cardiacum–allatumcomplex, and the ovaries in regulating vitellogenin levels in the housefly,Musca domestica. Journal of Insect Physiology 34, 11–19.
Arking, R., 1975. Temperature sensitive cell-lethal mutants of Droso-phila: Isolation and characterisation. Genetics 80, 519–523. Berger, E.M., Goudie, K., Klieger, L., Berger, M., DeCato, R., 1992.
The juvenile hormone analogue, methoprene, inhibits ecdisterone induction of small heat shock protein gene expression. Develop-mental Biology 151, 410–418.
Bownes, M., 1989. The roles of juvenile hormone, ecdysone and the ovary in the control ofDrosophila vitellogenesis. Journal of Insect Physiology 35, 409–413.
Bownes, M., 1994. The regulation of yolk-protein genes, a family of sex differentiation genes in Drosophila melanogaster. BioEssays 16, 745–752.
Cymborowski, B., 1991. Effects of cold stress on endocrice system in
Galleria melonella. In: Ivanovic, J., Jankovic-Hladni, M. (Eds.), Hormones and Metabolism in Insect Stress. CRC Press, Boca Raton, FL, pp. 99–114.
Davenport, A.K., Evans, P.D., 1984. Stress-induced changes in octopa-mine levels of insect haemolymph. Insect Biochemistry 14, 135– 143.
Expression of HSP70 cognate proteins. Russian Journal of Genetics 26, 266–271.
Granger, N.A., Sturgis, S.L., Ebersohl, R., Geng, C., Sparks, T.C., 1996. Dopaminergic control of corpora allata activity in the larval tobacco hornworm,Manduca sexta. Archives of Insect Biochemis-try and Physiology 32, 449–466.
Gruntenko, N.E., Khlebodarova, T.M., Sukhanova, M.JH., Vasenkova, I.A., Kaidanov, L.Z., Rauschenbach, I.YU., 1999. Prolonged nega-tive selection ofDrosophila melanogasterfor a character of adapt-ive significance disturbs stress reactivity. Insect Biochemistry and Molecular Biology 29, 445–452.
Gruntenko, N.E., Khlebodarova, T.M., Vasenkova, I.A., Sukhanova, M.J., Wilson, T.G., Rauschenbach, I.Y., 2000. Stress-reactivity of aDrosophila melanogasterstrain with impaired juvenile hormone action. Insect Physiology 46, 451–456.
Hammock, B.D., Sparks, T.C., 1977. A rapid assay for insect juvenile hormone esterase activity. Analytical Biochemistry 82, 573–579. Hirashima, A., Nagano, T., Takeya, R., Eto, M., 1993. Effect of larval
density on whole-body biogenic amine levels of Tribolium free-mani Hinton. Comparative Biochemical Physiology 106C, 457– 461.
Hirashima, A., Sukhanova, M.JH., Kuwano, E., Rauschenbach, I.YU., 1999. Alteration of biogenic amines in Drosophila virilis under stress. Drosophila Information Service 82, 30–31.
Hodgetts, R.B., 1972. Biochemical characterization of mutants affect-ing the metabolism of β-alanin in Drosophila. Journal of Insect Physiology 18, 937–947.
Hodgetts, R.B., Konopka, R.J., 1973. Tyrosine and catecholamine metabolism in wild-typeDrosophila melanogasterand a mutant,
ebony. Journal of Insect Physiology 19, 1211–1220.
Jowett, T., Postlethwait, J., 1981. Hormonal regulation of synthesis of yolk proteins and a larvae serum protein (LSP) inDrosophila. Nat-ure 292, 633–635.
Kalogianni, E., Theophilidis, G., 1993. Centrally generatedrhythmic activity and modulatory function of the oviductal dorsal unpaired median (DUM) neurones in two orthopteran species (Calliptamus
SP. andDecticus albifrons). Journal of Experimental Biology 174, 123–138.
Khlebodarova, T.M., Ankilova, I.A., Gruntenko, N.E., Sukhanova, M.ZH., Rauschenbach, I.YU., 1998. The impairments in Droso-phila stress reaction correlates with the alterations in heat shock response. Dokl. Akademia Nauk (Russia) 361, 184–l86.
Khlebodarova, T.M., Gruntenko, N.E., Grenback, L.G., Sukhanova, M.Z., Mazurov, M.M., Tomas, B.A., Hammock, B.D., Rauschen-bach, I.Y., 1996. A comparative analysis of juvenile hormone meta-bolyzing enzymes in two species of Drosophiladuring develop-ment. Insect Biochemistry and Molecular Biology 26, 829–835. Monastirioti, M., Linn, C.E. Jr., White, K., 1996. Characterization of
Drosophila Tyramineb-hydroxylasegene and isolation of mutant flies lacking octopamine. Journal of Neuroscience 16, 3900–3911. Nijhout, F., 1994. Insect Hormones. Princeton University Press,
Prin-ceton, NJ.
Orchard, I., Loughton, B.G., 1981. Octopamine and short-term hyperli-paemia in the locust. Genetic and Comparative Endocrinology 45, 175–180.
Piulachs, M.D., Belles, H., 1989. Stimulatory activity of cycteamine on juvenile hormone release in adult females of the cockroach,
Blattela germanica. Comparative Biochemistry and Physiology 94A, 795–798.
Postlethwait, J.H., Parker, J., 1987. Regulation of vitellogenesis in
Drosophila. In: O’Connor, J.D., Alan, R. (Eds.), Molecular Biology of Invertebrate Development. Liss Inc, New York, pp. 29–42. Raikhel, A.S., Lea, A.O., 1985. Hormone-mediated formation of the
endocytic complex in mosquito oocytes. Genetics and Comparative Endocrinology 57, 422–423.
Ramadan, H., Alawi, A.A., Alawi, M.A., 1993. Catecholamines in
Drosophila melanogaster(wild type and ebony mutant) decuticala-rized retinas and brains. Cell Biology International 17, 765–771. Rauschenbach, I.Y., 1991. Changes in ecdysteroid and juvenile
hor-mone under heat stress. In: Ivanovic, J., Jankovic-Hladni, M. (Eds.) Hormones and Metabolism in Insect Stress. CRC Press, Boca Raton, FL, pp. 115–148.
Rauschenbach, I.YU., 1997. Stress response in insects: mechanism, genetic control, and role in adaptation. Russian Journal of Genetics 33, 942–949.
Rauschenbach, I.Y., Lukashina, N.S., Maksimovsky, L.F., Korochkin, L.I., 1987. Stress-like reaction ofDrosophilato adverse environ-mental factors. Journal of Comparative Physiology 157, 519–531. Rauschenbach, I.Y., Serova, L.I., Timochina, I.S., Chentsova, N.A., Schumnaja, L.V., 1993. Analysis of differences in dopamine con-tent between two lines of Drosophila virilisin response to heat stress. Journal of Insect Physiology 39, 761–767.
Rauschenbach, I.Y., Khlebodarova, T.M., Chentsova, N.A., Grun-tenko, N.E., Grenback, L.G., Yantsen, E.I., Filipenko, M.L., 1995. etabolism of the juvenile hormone inDrosophilaadults under nor-mal conditions and heat stress. Journal of Insect Physiology 41, 179–189.
Rauschenbach, I.YU., Gruntenko, N.E., Khlebodarova, T.M., Mazurov, M.M., Grenback, L.G., Sukhanova, M.JH., Shumnaja, L.V., Zakharov, I.K., Hammock, B.D., 1996. The role of the degra-dation system of the juvenile hormone in the reproduction of Dro-sophilaunder stress. Journal of Insect Physiology 42, 735–742. Rauschenbach, I.Y., Sukhanova, M.Z., Shumnaya, L.V., Gruntenko,
N.E., Grenback, L.G., Khlebodarova, T.M., Chentsova, N.A., 1997. Role of dopa decarboxylase and N-acetyltransferase in regulation of dopamine content inDrosophila virilisunder normal and heat stress conditions. Insect Biochemistry and Molecular Biology 27, 829–835.
Riddiford, L.M., 1970. Effects of juvenile hormone on the program-ming of postembryonic development in eggs of the silkworm, Hyla-phora cecropia. Developmental Biology 22, 249–263.
Riddiford, L.M., Ashburner, M., 1991. Role of juvenile hormone in larval development and metamorphosis inDrosophila melanogas-ter. Genetics and Comparative Endocrinology 82, 172–183. Roe, R.M., Crawford, C.L., Clifford, C.W., Woodring, J.P., Sparks,
T.C., Hammock, B.D., 1987. Role of juvenile hormone metabolism during embryogenesis of the house cricket, Acheta domesticus. Insect Biochemistry 17, 1023–1026.
Shapiro, A.B., Wheelock, G.D., Hagedorn, H.H., Baker, F.C., Tsai, L.W., Schooly, D.A., 1986. Juvenile hormone and juvenile hor-mone esterase in adult females of the mosquitoAedes aegypti. Jour-nal of Insect Physiology 32, 867–885.
Shemshedini, L., Wilson, T.G., 1990. Resistance to juvenile hormone and in insect growth regulator inDrosophilais associated with an altered cytosolic juvenile hormone binding protein. Proceedings of the National Academy of Science USA 87, 2072–2076.
Soller, M., Bownes, M., Kubli, E., 1999. Control of oocyte maturation in sexually mature Drosophila females. Developmental Biology 208, 337–351.
Sukhanova, M.JH., Shumnaya, L.V., Grenback, L.G., Gruntenko, N.E., Khlebodarova, T.M., Rauschenbach, I.Y., 1997. Tyrosine decar-boxylase and dopa decardecar-boxylase inDrosophila virilisunder heat stress. Biochemical Genetics 35, 91–103.
Temin, G., Zander, M., Roussel, J.-P., 1986. Physico-chemical (GC– MS) measurements of juvenile hormone III titres during embryogenesis of Locusta migratoria. International Journal of Invertebrate Reproductive Development 9, 105–112.
Thomas, S.R., Lengyel, J.A., 1986. Ecdysteroid-regulated heat-shock gene expression during Drosophila melanogaster development. Developmental Biology 115, 434–438.
Diploptera punctata. Journal of Comparative Physiology B 160, 241–249.
Wilson, T.G., Ashok, M., 1998. Insecticide resistance resulting from an absence of target-sitegene product. Proceedings of the National Academy of Science USA 95, 14040–14044.
Wilson, T.G., Fabian, G., 1986. A Drosophila melanogester mutant resistant to a chemical analog of juvenile hormone. Developmental Biology 118, 190–201.
Woodring, J.P., McBride, L.A., Fields, P., 1989. The role of octopam-ine in handling and exercise-induced hyperglycaemia and hyperli-paemia in Acheta domesticus. Journal of Insect Physiology 41, 613–617.
![Fig. 1.Hydrolysis of [3H]JH-III in 1-day-old females of Canton Sand ts403 strains of D](https://thumb-ap.123doks.com/thumbv2/123dok/3120453.1379263/3.598.86.248.73.268/fig-hydrolysis-jh-iii-females-canton-sand-strains.webp)