HUBUNGAN ANTARA STRUKTUR LANSKAP PERTANIAN
DENGAN KOMUNITAS LEPIDOPTERA DAN
HYMENOPTERA PARASITIKA
EVAWATY SRI ULINA
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi berjudul Hubungan antara Struktur Lanskap Pertanian dengan Komunitas Lepidoptera dan Hymenoptera Parasitika adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir Disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
EVAWATY SRI ULINA. Hubungan antara Struktur Lanskap Pertanian dengan Komunitas Lepidoptera dan Hymenoptera Parasitika. Dibimbing oleh DAMAYANTI BUCHORI, SJAFRIDA MANUWOTO dan PUDJIANTO.
Sektor pertanian masih dihadapkan pada permasalahan rendahnya produksi komoditas pertanian walaupun berbagai cara telah dilakukan baik melalui pendekatan intensifikasi maupun ekstensifikasi pertanian. Di sisi lain, pendekatan tersebut menyebabkan menurunnya keanekaragaman hayati yang diketahui memiliki kontribusi besar pada layanan ekosistem dalam usaha mewujudkan pertanian berkelanjutan. Oleh karena itu, lahan pertanian yang kurang sesuai bagi perkembangan serangga hama namun mendukung perkembangan musuh alami perlu dirancang. Secara umum penelitian ini bertujuan untuk mempelajari hubungan antara struktur lanskap, keanekaragaman spesies dan interaksi spesies yang terjadi di pertanaman, sehingga dapat dikembangkan menjadi landasan pengembangan pengendalian hama dalam rangka mewujudkan pertanian berkelanjutan. Secara khusus penelitian ini bertujuan untuk (1) mengetahui komposisi dan konfigurasi lanskap pertanian di kawasan Bogor, Sukabumi, dan Cianjur, Jawa Barat; (2) mempelajari praktik budi daya mentimun yang dilakukan oleh petani; (3) menjelaskan hubungan antara kompleksitas lanskap pertanian dengan komunitas hama Lepidopteradan Hymenoptera parasitika; (4) menjelaskan hubungan antara jarak dari habitat alami dengan komunitas Lepidoptera dan Hymenoptera parasitika; (5) mempelajari pergerakan parasitoid yang dominan memarasit serangga herbivora pada tanaman mentimun dan mengukur jarak terbang parasitoid tersebut.
Daerah penelitian adalah lanskap pertanian yang berada di kawasan Bogor, Sukabumi dan Cianjur, Jawa Barat. Pada daerah tersebut ditentukan 16 lokasi penelitian dengan kriteria memiliki lahan pertanaman mentimun, luas pertanaman mentimun 1 250 m2 (25 m x 50 m), jarak antar lokasi penelitian minimal 3 km dan memiliki habitat alami. Kuantifikasi lanskap dilakukan pada radius 500 m, dengan pertanaman mentimun sebagai titik pusatnya. Secara umum, 16 lokasi penelitian tersebut memiliki jenis penggunaan lahan yang sama, yaitu lahan pertanian, pepohonan, semak, perumahan, jalan dan badan air. Dalam radius 500 m (78.56 ha), kondisi lanskap di masing-masing lokasi penelitian tersebut sangat bervariasi. Proporsi lahan pertanian berkisar 16.20% – 68.56%, sedangkan area pepohonan berkisar antara 4.6% – 61.40%. Jenis tanaman yang dibudidayakan berjumlah 3 – 15 tanaman. Number of patch (NumP) berkisar antara 46 – 190, mean patch size (MPS) berkisar antara 0.41 – 1.72 ha, total edge (TE) berkisar antara 26 735 – 59 812 m. Matriks lokasi penelitian adalah lahan pertanian. Lokasi penelitian dapat dikelompokan menjadi empat tipe lanskap yaitu lanskap sangat sederhana (habitat alami 12%), sederhana (habitat alami 22%), kompleks (habitat alami 31%) dan sangat kompleks (habitat alami 35%). Berdasarkan jarak dari habitat alami, lanskap dikelompokkan menjadi dua, yaitu pertanaman dekat dari habitat alami (< 200 m) dan jauh dari habitat alami (> 400 m).
(Lepidoptera: Crambidae) merupakan hama yang ditemukan sepanjang masa pertumbuhan tanaman mentimun, sedangkan Aulacophora similis (Coleoptera: Chrysomelidae) dianggap sebagai hama utama pada masa awal pertumbuhan tanaman mentimun. Seluruh petani responden tidak mengetahui peran serangga sebagai musuh alami dan penyerbuk. Petani umumnya menggunakan pestisida untuk mengendalikan serangan hama. Perilaku petani dalam penggunaan pestisida relatif tidak berbeda antar tipe lanskap.
Hasil penelitian ini menunjukkan bahwa kompleksitas lanskap tidak memengaruhi komunitas Lepidoptera namun memengaruhi kelimpahan parasitoid, keanekaragaman fungsional Hymenoptera parasitika dan parasitisasinya. Rata-rata ukuran patch (MPS) dan jumlah patch (NumP) habitat pertanian memengaruhi parasitisasi dan keanekaragaman fungsional Hymenoptera parasitika. Peningkatan rata-rata ukuran patch habitat pertanian menyebabkan parasitisasi dan keanekaragaman fungsional menurun, dan sebaliknya pertambahan jumlah patch pertanian akan meningkatkan parasitisasi dan keanekaragaman fungsional Hymenoptera parasitika.
Jarak pertanaman mentimun dari habitat alami tidak memengaruhi keanekaragaman dan kelimpahan komunitas Lepidoptera, parasitoid primer dan hiperparasitoid pada pertanaman tersebut. Namun, jarak pertanaman mentimun dari habitat alami memengaruhi persentase Diaphania indica terparasit. Keanekaragaman dan kelimpahan Hymenoptera parasitika cenderung meningkat dengan semakin dekatnya pertanaman mentimun dari habitat alami. Sejumlah 56 (43.1%) spesies Hymenoptera parasitika hanya ditemukan pada pertanaman dengan jarak tertentu saja dari habitat alami. Dari jumlah tersebut, sebanyak 39 spesies (69.6%) Hymenoptera parasitika ditemukan di pertanaman mentimun yang berjarak dekat dari habitat alami, sedangkan pada pertanaman yang berjarak sedang dari habitat alami ditemukan 17 spesies (13.1%). Keragaman fungsional Hymenoptera parasitika cenderung meningkat dengan semakin dekatnya lahan pertanaman ke habitat alami.
Perilaku pemencaran parasitoid merupakan informasi yang harus diketahui agar tindakan konservasi parasitoid berlangsung efektif. Pola pemencaran Apanteles taragame (Hymenoptera: Braconidae) dan inangnya adalah berkelompok. Apanteles taragamae mampu terbang hingga 53.6 m dari titik pelepasan. Pemencaran dan kemampuan terbang parasitoid dipengaruhi oleh keberadaan inang dan bukan oleh kondisi habitat sekitar pertanaman.
SUMMARY
EVAWATY SRI ULINA. The Relations between Landscape Structure with the Communities of Lepidoptera and Hymenopteran Parasitica. Supervised by DAMAYANTI BUCHORI, SJAFRIDA MANUWOTO and PUDJIANTO.
The agricultural sector has always been faced with the problem of low production of agricultural commodities although the agricultural intensification and extension have been done. On the other hand, the approach led to the decline of biodiversity known to have a major contribution to ecosystem services in any efforts towards sustainable agriculture. Therefore, we should design the agricultural land not suitable for insect pests but can conserve natural enemies. In general, this study aims to provide substantial information on the relationship between landscape structure, species diversity and species interactions that occur in the crop of cucumber, so it can be developed into a cornerstone of the development of pest control in order to realize sustainable agriculture. In particular, the objectives of study were (1) obtain the composition and configuration of agricultural landscapes in lowland and medium West Java; (2) an inventory of cucumber cultivation practices carried out by farmers; (3) describes the relationship between the complexity of the agricultural landscape with the communities of Lepidoptera and Hymenoptera parasitica; (4) explain the relationship between the distance of a natural habitat with Lepidoptera and Hymenoptera parasitica communities; (5) to study the movement of the dominant parasitoid in cucumber fields and measure the distance of the parasitoid fly.
Agricultural landscapes in 16 research sites are located in lowland and midland in West Java has the same type of landuse, such as agricultural land, trees, shrubs, housing, roads and water bodies. Within a radius of 500 m, the landscape is very greatly. The proportion of agricultural land ranges from 16.20% - 68.56%, while the area of trees ranging between 4.6% - 61.40%. Types of cultivated plants around 3-15 plants. The patch number (NumP) ranged between 46-190, MPS ranged from 0.41 - 1.72 ha, TE range between 26 735-59 812 m. For understanding the relationship between landscape composition and insect communities, landscapes are grouped based on the proportion of natural habitat into four types of landscape, the very simple landscape (natural habitat around 12%), simple landscape (natural habitat about 22%), complex landscape (natural habitat is 31%) and very complex landscape (natural habitats is 35%). Based on the distance to natural habitat, landscape grouped into two, namely the cropland close to natural habitats (<200 m) and the cropland far to natural habitats (> 400 m).
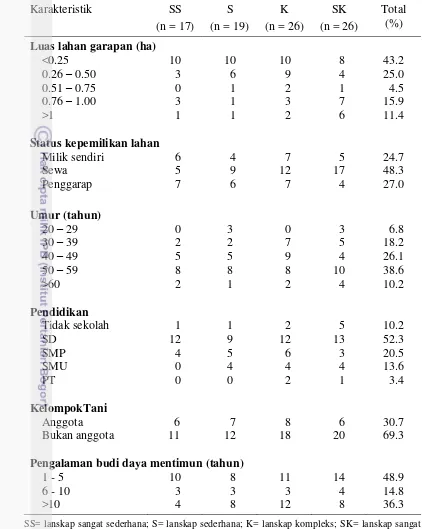
Results of interviews with farmers revealed that farmers generally have a productive age, land area used for cucumber cultivation less than 0.5 ha, low education and never contact with agricultural extension. Diaphania indica (Lepidoptera: Crambidae) reported as pests are found throughout the plant growth cucumber, while Aulacophora similis (Coleoptera: Chrysomelidae) considered a major pest in the early growth of cucumber plants. The whole farmer respondents did not know the role of insects as natural enemies and pollinators. Farmers are very dependent on the use of pesticides.
percentage. Mean patch size (MPS) and patch number (NumP) of farmland influenced percentage of parasitation and functional diversity of Hymenoptera parasitoid. Increasing the mean patch size of farmland patch decreases percentage of parasitation and functional diversity of Hymenoptera parasitoid, while the patch number of farmland will increase parasitation and functional diversity of Hymenoptera parasitoid.
Distance of cucumbers field to the natural habitat does not affect the diversity and abundance of Lepidoptera communities, primary parasitoid and hyperparasitoid. However, distance of cucumbers field to natural habitats affect the percentage of parasitized Diaphania indica. Diversity and abundance of Hymenoptera parasitica tends to increase on the cucumber field that closed to natural habitats. A number of 56 (43.1%) parasitica Hymenoptera species found only in cropping with a certain distance away from the natural habitat. Of these, as many as 39 species (69.6%) Hymenoptera parasitica found in cucumber cultivation within close natural habitats, while the crop is being of natural habitats found 17 species (13.1%). The functional diversity of Hymenoptera parasitica tends to increase on cropland closed to natural habitat.
The distribution pattern of Apanteles taragamae and its host are aggregate. Parasitoid A. taragamae (Hymenoptera: Braconidae) able to fly up to 53.6 m from the point of release. Dispersal of A. taragamae influenced by the presence of the host and is not affected by habitat conditions around the farmland.
© Hak Cipta Milik IPB, Tahun 2017
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi
HUBUNGAN ANTARA STRUKTUR LANSKAP PERTANIAN
DENGAN KOMUNITAS LEPIDOPTERA DAN
HYMENOPTERA PARASITIKA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup: 1. Dr. Ir. Purnama Hidayat, MSc
(Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor)
2. Dr. Akhmad Rizali, SP., MSi
(Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Brawijaya)
Penguji Luar Komisi pada Sidang Promosi: 1. Dr. Ir. Purnama Hidayat, MSc
(Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor)
2. Prof. (Riset) Dr. Ir. Deciyanto Soetopo, MS
PRAKATA
Alhamdulillahirabbil ‘alamin. Puji syukur atas segala rahmat dan karunia yang diberikan Allah subhanahu wa ta’alasehingga disertasi berjudul “Hubungan antara Struktur Lanskap Pertanian dengan Komunitas Lepidoptera dan Hymenoptera parasitika” berhasil penulis selesaikan. Disertasi ini disusun untuk memenuhi persyaratan memperoleh gelar Doktor pada Program Studi Entomologi, Sekolah Pascasarjana, Institut Pertanian Bogor (IPB).
Penghargaan dan ucapan terima kasih penulis sampaikan kepada ketua komisi pembimbing Prof. Dr. Ir. Damayanti Buchori, MSc., untuk waktu, arahan, bimbingan dan memperkenalkan saya pada ilmu dan seni melakukan penelitian lapangan dalam skala luas, dan juga menginspirasi saya untuk bekerja lebih keras. Terima kasih saya ucapkan pada anggota komisi pembimbing, Prof. Dr. Ir Sjafrida Manuwoto, MSc dan Dr. Ir. Pudjianto, MSi untuk waktu, arahan, bimbingan dan saran yang diberikan dengan tulus dan penuh kesabaran sehingga penulis dapat menyelesaikan penelitian dan penulisan disertasi ini. Pengalaman dan wawasan berfikir yang diberikan merupakan hal yang sangat berharga bagi penulis dalam menyusun disertasi ini serta pembelajaran dalam kehidupan penulis. Terima kasih diucapkan kepada Dr. Ir. Purnama Hidayat, MSc, Dr. Akhmad Rizali, SP, MSi dan Prof. (Riset) Dr. Ir. Deciyanto Soetopo, MSi sebagai penguji Luar Komisi.
Terima kasih penulis ucapkan kepada Kepala Badan Penelitian dan Pengembangan Pertanian Kementerian Pertanian Republik Indonesia, Kepala Balai Besar Pengkajian dan Pengembangan Teknologi Pertanian, dan Kepala Balai Pengkajian Teknologi Pertanian (BPTP) Sumatera Utara, atas kesempatan, izin, serta bantuan materil yang diberikan selama penulis melaksanakan tugas belajar di Program Doktor, Sekolah Pascasarjana IPB. Ucapan terimakasih disampaikan kepada Pimpinan dan staf Pusat Aplikasi Teknologi Isotop dan Radiasi (PAIR) Badan Tenaga Nuklir Nasional (BATAN) yang telah memfasilitasi penyediaan radioaktif 32P dan proses penandaan dan deteksi 32P pada serangga. Terima kasih juga penulis sampaikan kepada Rektor IPB, Ketua Departemen Proteksi Tanaman (PTN), Ketua Program Studi Entomologi, staf pengajar Program Studi Entomologi, serta staf administrasi Pascasarjana dan PTN IPB atas bantuan yang diberikan selama penulis menempuh pendidikan.
Kepada kedua orangtua terkasih, H. Sjamsuddin Sembiring (alm) dan Hj. Mariany Surbakti, yang telah mencurahkan kasih sayang, memberikan pendidikan terbaik, serta doa yang tidak pernah putus sehingga penulis dapat menggapai cita-cita, semoga Allah melimpahkan rahmat dan karuniaNya. Terima kasih juga penulis ucapkan kepada Ayah mertua H. Asmen Moechtar (alm), ibu mertua Hj. Yuwisma (alm), adik, kakak ipar, adik ipar serta keponakan atas dorongan semangat dan doa yang diberikan.
Kepada suamiku tercinta, Herman Asmen, terima kasih atas seluruh doa, cinta, kesabaran, dorongan semangat serta selalu berada di samping penulis dalam setiap tawa dan tangis selama menempuh pendidikan ini. Untuk permata hatiku, Abiyyu Denvisevian AAPS, terima kasih atas pengertian dan pengorbanan yang telah diberikan selama bunda menempuh pendidikan ini. Semoga ini dapat memberikan contoh yang baik bagi ananda untuk senantiasa belajar dan menuntut ilmu sampai kapanpun.
Kepada semua pihak yang telah membantu dalam penyelesaian studi ini, penulis ucapkan terima kasih semoga Allah SWT berkenan membalas semua kebaikan yang telah diberikan. Semoga karya ilmiah ini bermanfaat bagi perkembangan ilmu pengetahuan. Aamiin
Bogor, Februari 2017
DAFTAR ISI
DAFTAR TABEL xix
DAFTAR GAMBAR xx
DAFTAR LAMPIRAN xxii
1 PENDAHULUAN 1
Latar Belakang ... 1
Perumusan Masalah ... 3
Tujuan Penelitian ... 4
Hipotesis Penelitian ... 4
Manfaat Penelitian ... 6
2 TINJAUAN PUSTAKA 7 3 KOMPOSISI DAN KONFIGURASI LANSKAP PERTANIAN DI KAWASAN BOGOR, SUKABUMI DAN CIANJUR JAWA BARAT 16 Pendahuluan ... 16
Bahan dan Metode... 17
Hasil ... 20
Pembahasan ... 26
Simpulan ... 29
4 PENGETAHUAN DAN PERILAKU PETANI DALAM BUDI DAYA DAN PENGENDALIAN HAMA MENTIMUN 30 Pendahuluan ... 30
Bahan dan Metode... 31
Hasil ... 32
Pembahasan ... 37
Simpulan ... 38
5 HUBUNGAN KOMPLEKSITAS LANSKAP DENGAN KOMUNITAS LEPIDOPTERA DAN HYMENOPTERA PARASITIKA PADA PERTANAMAN MENTIMUN 39 Pendahuluan ... 39
Bahan dan Metode... 40
Hasil ... 44
Pembahasan ... 61
Simpulan ... 65
6 HUBUNGAN JARAK PERTANAMAN DARI HABITAT ALAMI DENGAN KOMUNITAS LEPIDOPTERA DAN HYMENOPTERA PARASITIKA 66 Pendahuluan ... 66
Hasil ... 70
Pembahasan ... 78
Simpulan ... 80
7 POLA PEMENCARAN Apanteles taragamae Viereck (HYMENOPTERA: BRACONIDAE) PADA LANSKAP PERTANIAN 81 Pendahuluan ... 81
Bahan dan Metode ... 82
Hasil ... 85
Pembahasan ... 89
Simpulan ... 91
8 PEMBAHASAN UMUM 92
9 SIMPULAN DAN SARAN 96
DAFTAR PUSTAKA 97
LAMPIRAN 113
DAFTAR TABEL
3.1 Letak administratif, koordinat dan ketinggian lokasi penelitian 18 3.2 Skor dan bobot parameter lanskap yang digunakan pada
pengelompokan lanskap 20
3.3 Proporsi penggunaan lahan dalam radius 500 m (78.56 ha) di 16
lanskap pertanian 22
3.4 Luas area pertanian dalam radius 500 m dan jumlah jenis tanaman
yang ditanami pada area tersebut 24
3.5 Nilai parameter lanskap 16 lokasi penelitian 25 3.6 Rata-rata (+SD) nilai parameter lanskap 16 lokasi penelitian pada
level kelas 27
3.7 Nilai CA, MPS dan NumP pada area pepohonan dan semak serta tipe
lanskap di 16 lokasi penelitian 28
3.8 Luas lahan dan proporsi penggunaan lahan pada lanskap sangat sederhana (SS), sederhana (S), kompleks (K), sangat kompleks (SK) 29 4.1 Karakteristik petani mentimun di empat tipe lanskap pertanian Jawa
Barat 33
4.2 Perilaku petani dalam budi daya mentimun di empat tipe lanskap
pertanian Jawa Barat 34
5.1 Serangga Lepidoptera yang berasosiasi dengan tanaman mentimun,
jumlah individu dan kategori dominasi 44
5.2 Spesies inang, stadia inang serta spesies parasitoid yang berasosiasi dengan serangga Lepidoptera pada tanaman mentimun 46 5.3 Spesies hiperparasitoid dan hiperparasitisasi pada A. taragamae 49 5.4 Kekayaan spesies dan kelimpahan hama Lepidoptera dan
parasitoidnya pada beberapa tipe lanskap 50
5.5 Hubungan antara parameter lanskap dengan kelimpahan, kekayaan
spesies serangga dan parasitisasi 54
5.6 Keanekaragaman dan kelimpahan Hymenoptera parasitika di pertanaman mentimun pada beberapa tipe lanskap 55 5.7 Hubungan antara parameter lanskap dengan community weighted
mean (CWM) panjang tubuh, panjang sayap depan, panjang sayap belakang, panjang tibia belakang dan lebar kepala Hymenoptera
parasitika 62
6.1 Kekayaan spesies dan kelimpahan hama Lepidoptera dan parasitoidnya pada pertanaman mentimun yang dekat dan sedang
dari habitat alami 71
6.2 Keanekaragaman dan kelimpahan Hymenoptera parasitika di pertanaman mentimun yang berjarak dekat dan sedang dari habitat
alami 75
7.1 Lokasi administratif dan koordinat lokasi pelepasan parasitoid 84 7.2 Pola distribusi larva D. indica dan A. taragamae pada pertanaman
mentimun 86
7.3 Jumlah inang dan parasitisasinya oleh parasitoid yang dilepas pada
pertanaman mentimun 88
DAFTAR GAMBAR
1.1 Peta jalan penelitian hubungan struktur lanskap pertanian dengan
komunitas Lepidoptera dan Hymenoptera parasitika 5
2.1 Komponen penyusun lanskap 7
2.2 Tipe patch berdasarkan cara terbentuknya 8
2.3 Ilustrasi pola pemencaran serangga 14
3.1 Lokasi penelitian di kawasan Bogor, Sukabumi dan Cianjur 17 3.2 Foto udara lokasi penelitian yang digunakan sebagai pedoman
dalam melakukan groundcheck 19
3.3 Peta penggunaan lahan dalam radius 500 m di 16 lanskap pertanian 21 3.4 Hubungan proporsi luas area pepohonan dan pertanian dalam
radius 500 m di 16 lanskap pertanian 22
3.5 Komposisi tanaman pertanian pada radius 500 m di 16 lanskap
pertanian 23
4.1 Peta lokasi penelitian wawancara petani mentimun 32 4.2 Hama utama tanaman mentimun pada lanskap sangat sederhana,
sederhana, kompleks dan sangat kompleks 35
4.3 Jenis pestisida yang umum digunakan petani untuk mengendalikan hama tanaman mentimun di lanskap sangat sederhana (SS),
sederhana (S), kompleks (K) dan sangat kompleks (SK) 36 4.4 Interval waktu aplikasi pestisida pada pertanaman mentimun
dalam satu musim tanam 36
4.5 Jumlah aplikasi pestisida pada pertanaman mentimun dalam satu
musim tanam 37
5.1 Peta lokasi penelitian berdasarkan kompleksitas lanskap 41 5.2 Desain plot pengambilan serangga Lepidoptera pradewasa dan
Hymenoptera parasitika 42
5.3 Serangga Lepidoptera yang menyerang tanaman mentimun 44 5.4 Komposisi hama Lepidoptera pada berbagai umur tanaman
mentimun 45
5.5 Diaphania indica (A) imago, (B) Telur, (C) Larva dan (D) Pupa 45 5.6 Parasitoid yang memarasit D. indica, C. chalcites dan S. litura
pada pertanaman mentimun 47
5.7 Komposisi parasitoid yang memarasit hama Lepidoptera pada
berbagai umur tanaman mentimun 48
5.8 Hubungan kelimpahan larva D. indica dan jumlah D. indica
terparasit pada tanaman mentimun 49
5.9 Hubungan kompleksitas lanskap dengan (A) kelimpahan larva D.
indica dan (B) persentase kerusakan tanaman 51 5.10 Interaksi tritropik hama, parasitoid dan hiperparasitoid pada
pertanaman mentimun di lanskap (A) sangat sederhana, (B)
sederhana, (C) kompleks dan (D) sangat kompleks 52 5.11 Efek kompleksitas lanskap terhadap parasitisasi 53 5.12 Jumlah spesies famili Hymenoptera parasitika di empat tipe
5.13 Kelimpahan famili Hymenoptera parasitika yang diperoleh dari (A) transek dan perangkap nampan kuning serta (B) perangkap nampan
kuning di empat tipe lanskap 52
5.14 Kelimpahan spesies Hymenoptera parasitika pada pertanaman
mentimun di empat tipe lanskap 57
5.15 Diagram Venn jumlah spesies Hymenoptera parasitika di empat
tipe lanskap 58
5.16 Keragaman ukuran tubuh Hymenoptera parasitika pada berbagai
kompleksitas lanskap 59
5.17 Hubungan kompleksitas lanskap dengan community weighted mean (CWM) karakteristik morfologi Hymenoptera parasitika 60 6.1 Peta lokasi penelitian berdasarkan jarak dari habitat alami 67 6.2 Desain plot pengambilan serangga Lepidoptera pradewasa dan
Hymenoptera parasitika 68
6.3 Hubungan jarak lokasi pertanaman mentimun dari habitat alami dengan (A) proporsi kelimpahan dan (B) distribusi spesies hama Lepidoptera yang menyerang tanaman mentimun 71 6.4 Hubungan jarak lokasi pertanaman mentimun dari habitat alami
dengan persentase tanaman mentimun terserang hama Lepidoptera 72 6.5 Interaksi tritrofik hama, parasitoid dan hiperparasitoid pada
pertanaman mentimun di berbagai jarak pertanaman dari habitat
alami 73
6.6 Hubungan jarak pertanaman mentimun dari habitat alami dengan (A) kekayaan spesies dan (B) kelimpahan parasitoid yang
memarasit D. indica 74
6.7 Hubungan antara persentase D. indica terparasit dengan (A) kekayaan spesies dan (B) kelimpahan parasitoid yang memarasit D.
indica 75
6.8 Jumlah spesies famili Hymenoptera parasitika di pertanaman mentimun yang berjarak dekat dan sedang dari habitat alami 76 6.9 Kelimpahan famili Hymenoptera parasitika di pertanaman
mentimun yang berjarak dekat dan sedang dari habitat alami 76 6.10 Diagram Venn jumlah spesies Hymenoptera parasitika di berbagai
jarak pertanaman dari habitat alami 77
6.11 Famili Hymenoptera parasitika yang ditemukan pada keseluruhan
jarak dari habitat alami 78
6.12 Hubungan jarak pertanaman mentimun dari habitat alami dengan community weighted mean (CWM) karakteristik morfologi
Hymenoptera parasitika 79
7.1 Peta lokasi penelitian pola pemencaran serangga 82 7.2 Lanskap pelepasan parasitoid dalam radius 500 m dengan titik
pelepasan pada areal pertanaman mentimun (titk tengah lanskap) 83
7.3 Titik pelepasan parasitoid A. taragamae 84
7.4 Denah plot pengamatan (pengumpulan) larva D. indica 84 7.5 Pola keberadaan D. indica dan A. taragamae pada lahan
pertanaman mentimun di Bantarjaya 86
7.6 Pola keberadaan D. indica dan A. taragamae pada lahan
7.7 Pola keberadaan D. indica dan A. taragamae pada lahan
pertanaman mentimun di Cihideung Udik 87
7.8 Pola keberadaan D. indica dan A. taragamae pada lahan
pertanaman mentimun di Mekarjaya 88
7.9 Pola pemencaran A. taragamae 89
DAFTAR LAMPIRAN
1 Daftar pertanyaan wawancara kepada petani mentimun 115 2 Jenis Lepidoptera yang berasosiasi dengan pertanaman mentimun di
empat tipe lanskap 121
3 Spesies Hymenoptera parasitika yang ditemukan di pertanaman
mentimun 122
4 Foto spesimen Hymenoptera parasitika yang ditemukan di empat tipe
1
PENDAHULUAN
Latar Belakang
Tantangan yang dihadapi sektor pertanian di masa yang akan datang adalah meningkatkan produktivitas tanaman dan di sisi lain menjaga keanekaragaman hayati pada ekosistem pertanian. Ini merupakan dua hal yang tidak mudah untuk dilaksanakan karena seringkali menimbulkan konflik kepentingan. Peningkatan produksi pertanian umumnya dilakukan melalui pendekatan intensifikasi maupun ekstensifikasi pertanian. Intensifikasi pertanian terjadi melalui peningkatan input seperti penggunaan pestisida, pupuk kimia, varietas unggul, benih rekayasa genetik, mesin pertanian untuk mengolah tanah serta rotasi tanaman yang singkat (Benton et al. 2003; Tscharntke et al. 2005). Sedangkan ekstensifikasi pertanian terjadi melalui aktifitas konversi ekosistem alami menjadi ekosistem pertanian, fragmentasi habitat alami dan regionalisasi komoditas pertanian (Tscharntke et al. 2005). Hal ini menyebabkan rendahnya keanekaragaman hayati pada ekosistem pertanian selain karena perubahan iklim, polusi dan invasi biotik (Hooper et al. 2005; MA 2005). Keanekaragaman hayati merupakan komponen yang sangat penting untuk mengoptimalkan jasa ekosistem (Hoehn et al. 2008; Bennett et al. 2012; Cardinale et al. 2012). Kontribusi jasa ekosistem yang tidak optimal akan menyebabkan terjadinya perbedaan produksi yang diperoleh petani dengan potensi produksi komoditas tersebut (Boomarco et al. 2013).
Pengendalian populasi hama oleh musuh alami merupakan salah satu jasa ekosistem yang tidak saja bermanfaat bagi lingkungan tetapi juga menguntungkan dari sisi ekonomi, karena tindakan tersebut mampu mengurangi kehilangan hasil tanpa menimbulkan kerusakan lingkungan yang disebabkan oleh penggunaan pestisida (Ȍstman et al. 2003; Boomarco et al. 2013). Di Amerika, nilai ekonomi pengendalian hayati hama adalah sebesar US$ 4.5 milyar per tahun (Losey dan Vaughan 2006), sedangkan di dunia diperkirakan mencapai US$ 417 milyar per tahun (Costanza et al. 1997).
Keberhasilan pengendalian hayati dipengaruhi oleh berbagai komponen pada skala petakan maupun lanskap (Gurr et al. 2003; Tscharntke et al. 2007). Pada skala petakan, teknologi pengelolaan tanaman seperti penggunaan pestisida, pupuk kimia, varietas unggul, benih rekayasa genetik dan rotasi tanaman dapat memengaruhi komposisi, kelimpahan dan sebaran spesies serangga pada agroekosistem (Booij dan Noorlander 1992; Cárcamo 1995; Bengtsson et al. 2005). Sedangkan pada skala lanskap, struktur komunitas, kekayaan dan kelimpahan spesies, dinamika populasi dan interaksi antar trofik dipengaruhi ukuran patch, konfigurasi spasial, komposisi lanskap, konektivitas habitat dan kompleksitas habitat (Kareiva 1987; Marino dan Landis 1996; Zabel dan Tscharntke 1998; Tscharntke dan Brandl 2004; Bianci et al. 2006; Finke and Denno, 2006; Woodcock et al. 2007).
dipengaruhi oleh keberadaan musuh alami, tetapi juga turut dipengaruhi oleh vegetasi yang ada pada ekosistem tersebut. Menurut Root (1973), dalam teori resource concentration hypothesis kelimpahan hama rendah pada ekosistem dengan vegetasi yang lebih beragam karena hama spesialis akan sulit untuk menemukan inangnya.
Struktur komunitas dapat digambarkan melalui tiga pendekatan yaitu keanekaragaman spesies (kekayaan dan kelimpahan relatif spesies), interaksi trofik dan kelompok fungsional (Schowalter 2011). Keanekaragaman spesies merupakan pendekatan yang umum digunakan untuk mempelajari struktur komunitas serangga. Pendekatan yang sama juga digunakan dalam penelitian ini. Menurut Whittaker (1972), terdapat tiga tingkat keanekaragaman spesies berdasarkan skala geografi, yaitu keanekaragaman alfa, beta dan gamma. Keanekaragaman alfa adalah keanekaragaman spesies dalam suatu habitat. Keanekaragaman beta adalah ukuran keanekaragaman yang dihitung dengan membandingkan keanekaragaman spesies antar ekosistem atau berdasarkan gradien lingkungan. Sedangkan keanekaragaman gamma yaitu keanekaragaman spesies di seluruh habitat yang dibandingkan.
Komponen keanekaragaman spesies yang menentukan karakter spesies tersebut terdiri dari kekayaan spesies, kemerataan spesies, komposisi serta interaksi spesies (Chapin et al. 2000). Namun, peran kekayaan spesies musuh alami dalam pengendalian hayati tidak konsisten. Finke dan Denno (2004, 2005) melaporkan bahwa kemampuan komunitas musuh alami yang sederhana dalam mengendalikan populasi hama adalah sama atau lebih baik dibandingkan dengan komunitas musuh alami yang kompleks. Beberapa penelitian lain justru menunjukkan sebaliknya, dimana komunitas musuh alami yang beragam lebih efektif mengendalikan populasi serangga hama dibandingkan dengan komunitas musuh alami yang relatif kurang beragam (Cardinale et al. 2003; Aquilino et al. 2005; Wilby et al. 2005; Straub 2006; Snyder et al 2006). Hal ini menunjukkan bahwa pengaruh komunitas musuh alami tidak berlaku umum karena ada faktor-faktor lain yang turut memengaruhi hasil akhir dari interaksi hama dan musuh alami.
Beberapa penelitian menunjukkan bahwa keanekaragaman serangga dipengaruhi oleh kompleksitas lanskap. Keanekaragaman serangga herbivor akan meningkat dengan bertambah luasnya habitat alami (Clough et al. 2007). Menurut
Plećaš et al. (2014) kepadatan populasi kutudaun, parasitisasi dan keanekaragaman parasitoid kutudaun dan hiperparasitoid akan meningkat dengan bertambah kompleksnya suatu lanskap. Penelitian lainnya menunjukkan bahwa kompleksitas lanskap tidak memengaruhi keanekaragaman dan kelimpahan parasitoid. Kepadatan kutudaun dan parasitoidnya tidak dipengaruhi oleh kompleksitas lanskap (Vollhardt et al. 2008; Caballero-Lopez et al. 2012). Hal ini menunjukkan bahwa pengetahuan mengenai pengaruh kompleksitas lanskap terhadap struktur dan komposisi serangga serta kemampuannya untuk menekan populasi hama masih belum menjadi pengetahuan yang berlaku umum.
alternatif, tempat berlindung dan iklim mikro lahan pertanian yang sesuai bagi perkembangan musuh alami (Landis et al. 2000; Gagic et al. 2011).
Mentimun (Cucumis sativus L) merupakan salah satu komoditas hortikultura komersial yang dibudidayakan hampir di seluruh wilayah Indonesia mulai dari dataran rendah hingga tinggi. Pada tahun 2013, luas areal panen mentimun mencapai 49 296 ha dengan produksi 491 636 ton dan rata-rata hasil 9.97 t/ha (Kementerian Pertanian 2015). Luas areal panen dan produksi mentimun tersebut jauh lebih rendah dibandingkan lima tahun yang lalu. Penurunan luas lahan tersebut disebabkan oleh perubahan komoditas yang ditanam oleh petani atau konversi lahan pertanian menjadi nonpertanian. Sedangkan angka rata-rata produktivitas 9.97 t/ha tergolong belum optimal, karena menurut hasil penelitian AVNET (Asean Vegetable Network) potensi hasil mentimun berkisar antara 12 sampai 19 t/ha (Sumpena dan Permadi 2005) dan potensi hasil tiga varietas mentimun yang dihasilkan oleh Balai Penelitian Tanaman Sayuran berkisar antara 21 sampai 35 t/ha (Sumpena dan Permadi 1999). Di Jawa Barat, tanaman mentimun dapat ditemukan sepanjang tahun pada lanskap sederhana maupun kompleks serta pada berbagai ketinggian daerah. Kabupaten Bogor, Sukabumi dan Cianjur merupakan salah satu sentra pertanaman mentimun di Jawa Barat.
Pengaruh lanskap terhadap serangga herbivor dan parasitoidnya di daerah tropis khususnya Indonesia perlu diteliti karena lanskap pertanian Indonesia juga banyak mengalami perubahan sejak terjadinya revolusi hijau. Sampai saat ini penelitian pengendalian hayati lebih ditekankan pada pendekatan individu dan skala lokal sedangkan praktek di lapangan lebih banyak difokuskan pada pelepasan (baik secara inokulasi dan inundasi musuh alami). Upaya untuk melakukan konservasi terhadap musuh alami yang telah ada di lapangan masih belum banyak dilakukan, padahal untuk mencapai tingkat keberhasilan yang tinggi dalam pengendalian hayati. Konservasi musuh alami mutlak perlu dilakukan setelah tahapan inokulasi dan inundasi dilaksanakan. Oleh karena itu, kajian ini dianggap penting sebab berpengaruh terhadap strategi pengendalian hama guna mendukung pertanian berkelanjutan.
Perumusan Masalah
berpindah serta menempati habitat lain di saat tanaman utama sudah dipanen atau tidak ada di lapangan. Penelitian ini dilaksanakan untuk menjawab pertanyaan-pertanyaan sebagai berikut:
1. Bagaimana tipe lanskap pertanian di kawasan Bogor, Sukabumi dan Cianjur?
2. Bagaimana praktik-praktik budi daya yang dilakukan oleh petani?
3. Apakah kompleksitas lanskap memiliki hubungan dengan komunitas serangga Lepidoptera dan Hymenoptera parasitika?
4. Apakah jarak pertanaman dari habitat alami memengaruhi komunitas serangga Lepidoptera dan Hymenoptera parasitika?
5. Bagaimana perilaku dan pola pemencaran parasitoid?
Peta jalan penelitian disajikan dalam Gambar 1.1. Peta jalan ini menggambarkan arah penelitian hubungan antara struktur lanskap pertanian dengan komunitas Lepidoptera dan Hymenoptera parasitika di pertanaman mentimun.
Tujuan Penelitian
Tujuan Umum
Secara umum penelitian ini bertujuan untuk mengetahui hubungan antara struktur lanskap, keanekaragaman spesies serangga dan interaksi spesies serangga yang terjadi di pertanaman mentimun, sehingga dapat dikembangkan menjadi landasan pengembangan pengendalian hama dalam rangka mewujudkan pertanian berkelanjutan.
Tujuan Khusus
Secara khusus penelitian ini bertujuan sebagai berikut:
1. Mengetahui komposisi dan konfigurasi lanskap pertanian di kawasan Bogor, Cianjur dan Sukabumi, Jawa Barat.
2. Mempelajari praktik budi daya yang dilakukan oleh petani dan implikasinya terhadap keanekaragaman serangga
3. Menjelaskan hubungan antara kompleksitas lanskap pertanian dengan komunitas Lepidopteradan Hymenoptera parasitika
4. Menjelaskan hubungan antara jarak dari habitat alami dengan komunitas Lepidopteradan Hymenoptera parasitika
5. Mempelajari pergerakan parasitoid yang dominan memarasit serangga herbivora pada tanaman mentimun dan mengukur jarak terbang parasitoid tersebut.
Hipotesis Penelitian
Berdasarkan latar belakang dan tujuan penelitian yang telah dikemukakan, hipotesis penelitian ini disusun sebagai berikut:
1. Komposisi dan konfigurasi lanskap pertanian di kawasan Bogor, Cianjur dan Sukabumi, Jawa Barat tidak berbeda.
3. Lanskap pertanian sangat kompleks tidak berpengaruh terhadap kelimpahan, keanekaragaman, komposisi serta interaksi antara serangga Lepidopteradan musuh alaminya
4. Jarak dari habitat alami tidak berpengaruh terhadap kelimpahan, keanekaragaman, komposisi serta interaksi antara serangga Lepidoptera dan musuh alaminya
5. Pola sebaran inang tidak berpengaruh terhadap pola sebaran parasitoid Apanteles taragamae
Manfaat Penelitian
2
TINJAUAN PUSTAKA
Ekologi Lanskap
Istilah ‘ekologi lanskap’ pertama kali diperkenalkan oleh Carl Troll pada
tahun 1939. Troll mendefinisikan ekologi lanskap sebagai ilmu yang mempelajari hubungan antara susunan bidang lahan dalam suatu lanskap dengan spesies yang hidup di dalamnya (Forman dan Godron 1986). Ekologi lanskap berbeda dengan cabang ilmu ekologi lainnya, seperti ekologi populasi, komunitas dan ekosistem. Ekologi lanskap menekankan skala spasial yang luas dan dampak ekologi dari pola spasial ekosistem. Secara khusus, ekologi lanskap dianggap sebagai pengembangan dan dinamika heterogenitas spasial, interaksi dan pertukaran di seluruh lanskap yang heterogen, pengaruh heterogenitas spasial pada proses biotik dan abiotik, dan pengelolaan heterogenitas spasial. Dengan demikian, fokus utama dari ekologi lanskap adalah pada (a) wilayah geografis spasial yang heterogen, (b) pengaturan kembali elemen lanskap, dan (c) tindakan manusia sebagai respon terhadap proses ekologi dan pengaruh timbal balik manusia pada proses ekologi (Risser et al. 1984).
Lanskap adalah suatu lahan heterogen yang tersusun dari ekosistem yang saling berinteraksi dan memiliki semacam pola yang berulang-ulang (Forman dan Godron 1986). Lanskap fokus pada tiga karakteristik, yaitu struktur, fungsi dan perubahan.
Struktur Lanskap
Struktur lanskap mengacu pada pola spasial elemen lanskap dan hubungan antara ekosistem yang beda. Struktur lanskap menilai hubungan antara ekosistem sebagai ukuran, bentuk dan jumlah (Forman dan Godron 1986). Menurut Farina (2000), struktur lanskap memiliki dua kualitas, yaitu komposisi dan konfigurasi. Komposisi lanskap didefinisikan sebagai jenis dan sebaran patch dalam lanskap. Komposisi lanskap merupakan indikator yang baik untuk menilai kesesuaian lingkungan terhadap suatu spesies. Konfigurasi mengacu pada karakteristik spasial atau dengan kata lain mengacu pada distribusi spasial tutupan lahan. Struktur dasar lanskap terdiri dari 3 komponen yaitu matriks, patch dan koridor (Gambar 2.1).
Matriks adalah komponen lanskap yang ukurannya paling luas dan terkoneksi serta memegang peranan penting dalam fungsi lanskap (Forman dan Godron 1986). Lebih lanjut (Samways 1995) menjelaskan bahwa matriks merupakan elemen lanskap yang berada di sekitar patch. Menurut Forman (1995) konektivitas adalah ukuran bagaimana suatu koridor, jaringan (network), atau matriks terhubung atau berkesinambungan. Sebagai contoh, suatu lanskap dengan matriksnya adalah hutan, memiliki sedikit celah (gap) dalam tutupan hutannya berarti memiliki konektifitas lebih tinggi.
Patch merupakan suatu area berbentuk poligon yang relatif homogen yang berbeda dari sekitarnya (Forman dan Godron 1986). Patch dapat dibedakan menjadi 5 tipe berdasarkan cara terbentuknya (Gambar 2.2), yaitu: 1) disturbance patch, patch yang terbentuk oleh karena adanya gangguan pada sebagian kecil area matriks, misalnya patch yang terbentuk oleh karena adanya kebakaran hutan; 2) remnant patch, patch yang terbentuk oleh karena adanya gangguan dalam skala luas, misalnya pulau/daratan yang ditumbuhi semak-semak oleh karena lembahnya tergenang oleh banjir; 3) environmental resource patch, patch yang mencerminkan sumberdaya terdistribusi normal pada suatu lingkungan dan merupakan hasil dari sumberdaya yang permanen dan relatif terpisah dari daerah sekitarnya (patch ini tidak disebabkan oleh kerusakan), misalnya terkonsentrasinya amfibi dan reptil di gurun pasir; 4) introduced patch, patch yang didominasi oleh individu hasil introduksi manusia, misalnya perkebunan kelapa sawit; 5) ephemeral patch, spesies teragregasi sementara oleh karena adanya perubahan lingkungan biotik atau abiotik dengan intensitas rendah sehingga akhirnya spesies tersebut beradaptasi dengan perubahan yang terjadi. Perbedaan yang terjadi pada cara terbentuknya patch ini menentukan dinamika spesies di dalamnya (Forman dan Godron 1981).
Gambar 2.2 Tipe patch berdasarkan cara terbentuknya (Forman dan Godron 1981)
patch yang terisolasi dapat memengaruhi kekayaan fauna khususnya serangga. Kekayaan spesies serangga pemakan biji pada tanaman Vicia sepium meningkat pada area yang terdapat tanaman inang, dan menurun akibat terdapatnya isolasi pada patch (Kruess dan Tscharntke 2000).
Koridor merupakan patch yang bentuknya memanjang (Forman dan Godron 1986). Menurut McGarigal et al. (2014), berdasarkan fungsinya dikenal tiga macam bentuk koridor ini yaitu koridor habitat, koridor perpindahan (movement corridor), dan koridor perintang (barrier corridor). Koridor habitat merupakan koridor yang berfungsi sebagai tempat untuk hidup bagi suatu organisme baik sementara maupun permanen. Koridor perpindahan berfungsi sebagai jalur penghubung perpindahan suatu spesies dari satu habitat ke habitat lainnya. Koridor perintang merupakan koridor yang dapat menghambat aliran energi, nutrisi mineral, dan atau perpindahan spesies ke habitat lain. Koridor habitat dan koridor perpindahan berturut-turut secara pasif dan aktif dapat meningkatkan keterhubungan lanskap dengan organisme, sebaliknya koridor perintang dapat menurunkan keterhubungan lanskap dengan organisme.
Fungsi Lanskap
Fungsi lanskap adalah hasil interaksi antara elemen-elemen spasial, yaitu, aliran energi, material, dan organisme diantara komponen-komponen ekosistem (Forman dan Godron 1986). Fungsi berhubungan dengan struktur, tetapi lebih menyangkut proses-proses biologi, kimia, dan fisik yang terjadi di dalam lanskap. Keanekaragaman dan distribusi spesies, proses-proses populasi (migrasi, kelahiran dan kematian), genetika populasi, dan interaksi tanaman-hama dan musuh alami semuanya dipengaruhi oleh struktur lanskap (Fry 1995).
Perubahan Lanskap
Perubahan lanskap merupakan perubahan yang terjadi pada struktur dan fungsi lanskap dari waktu ke waktu (Forman dan Godron 1986). Faktor penting yang menyebabkan perubahan tersebut adalah karena seringkali terjadi gangguan dalam skala yang luas pada ekosistem tersebut, seperti kebakaran, penebangan pohon dan banjir (Landis dan Menalled 1998). Perubahan lanskap ini mengakibatkan kehilangan spesies dan rusaknya jejaring makanan (Kruess dan Tscharntke 1994).
Hubungan Struktur Lanskap dan Serangga
membuat iklim mikro pada lahan pertanian sesuai bagi perkembangan musuh alami (Landis et al. 2000; Tscharntke et al. 2007; Rusch et al. 2010). Namun, habitat alami tidak selamanya berpengaruh positif terhadap pengendalian hayati. Menurut Tscharntke et al. (2016), hal tersebut diduga disebabkan oleh lima hal, yaitu 1) tidak adanya musuh alami yang efektif mengendalikan populasi hama di daerah tersebut, 2) habitat alami lebih mendukung perkembangan serangga hama dibandingkan musuh alami, 3) tanaman budi daya lebih banyak menyediakan sumber daya yang dibutuhkan musuh alami dibandingkan yang disediakan oleh habitat alami, 4) habitat alami yang tersedia tidak sesuai dalam hal jumlah, jarak, komposisi atau konfigurasi bagi perkembangan musuh alami, dan 5) praktik budi daya yang dilakukan oleh petani mengancam keberadaan musuh alami.
Sejumlah studi menunjukkan bahwa lanskap kompleks, yang dicirikan dengan besarnya proporsi habitat alami, mampu meningkatkan keanekaragaman dan kelimpahan musuh alami (Finke dan Snyder 2010; Holzschuh et al. 2010). Sebaliknya, kelimpahan dan keanekaragaman musuh alami lebih rendah pada lanskap sederhana dibandingkan dengan lanskap kompleks. Hal ini disebabkan landskap sederhana tidak mampu menyediakan kebutuhan musuh alami, seperti inang alternatif dan pakan bagi imagonya (Ge’neau et al. 2012). Lanskap kompleks disamping dapat meningkatkan keanekaragaman dan kelimpahan musuh alami pada lahan pertanian, juga mampu meningkatkan keefektifan musuh alami dalam mengendalikan serangga hama (Clough et al. 2007). Parasitoid merupakan salah satu kelompok serangga yang kinerjanya dipengaruhi kompleksitas lanskap (Brewer et al. 2008; Zhao et al. 2013). Kinerja dan kelimpahan parasitoid meningkat pada lanskap kompleks yang dicirikan dengan tingginya proporsi habitat alami (Schmidt dan Tscharntke 2005; Steingrover et al. 2010). Kemampuan parasitoid dalam mengendalikan populasi hama di lapangan sangat dipengaruhi oleh kompleksitas komunitas parasitoid. Keanekaragaman parasitoid yang rendah menyebabkan banyak spesies hama tidak terparasit (Kruess dan Tscharntke 1994). Rendahnya keanekaragaman parasitoid ini disebabkan oleh rendahnya daya dukung habitat terhadap perkembangan sejumlah parasitoid yang ada di lapangan (Menalled et al 1999). Habitat alami ini berfungsi sebagai sumber pakan, inang alternatif, tempat berlindung, dan membuat iklim mikro pada lahan pertanian sesuai bagi perkembangan musuh alami (Landis et al. 2000; Gagic et al. 2011).
Pada ekosistem pertanian, kompleksitas komunitas parasitoid dipengaruhi oleh sejumlah faktor yang terdapat pada skala petakan maupun lanskap (Clough et al 2007). Pada skala petakan, kelimpahan dan kekayaan spesies serangga dipengaruhi oleh teknik budi daya tanaman seperti cara pengolahan tanah, penggunaan pestisida dan pupuk anorganik serta keanekaragaman spesies tanaman (Landis et al 2000; Thorbek dan Bilde 2004). Pada skala lanskap, kelimpahan dan kekayaan spesies serangga dipengaruhi oleh fragmentasi habitat dan struktur lanskap (Purtauf et al 2005; Tscharntke et al 2007).
parameter yang dapat mengukur dampak keanekaragaman spesies terhadap jasa ekosistem, seperti produksi tanaman, hal ini disebabkan keanekaragaman fungsional memberikan informasi mengenai karakteristik fungsional suatu spesies. Menurut Petchey dan Gaston (2002), setiap spesies memiliki fungsi yang berbeda pada suatu ekosistem karena adanya perbedaan karakteristik dari setiap spesies. Keanekaragaman fungsional dapat diukur dengan melihat nilai community weighted mean (CWM) karakter suatu spesies (Ricotta dan Moretti 2011; Persson et al. 2015). Menurut Ricotta dan Moretti (2011), karakteristik morfologi suatu spesies yang digunakan untuk mengukur CWM adalah karakteristik spesies yang dapat diukur seperti karakteristik morfologi.
Struktur lanskap pertanian memiliki keanekaragaman dan komposisi habitat yang sangat bervariasi, mulai dari sangat sederhana hingga kompleks. Variasi ini terjadi oleh karena adanya pengaruh faktor internal dan eksternal, seperti iklim, tanah, politik, sosial ekonomi dan budaya (Abdoellah et al. 2004). Di daerah Jawa Barat, kompleksitas habitat sangat dipengaruhi oleh rendahnya kepemilikan lahan pertanian. Rata-rata luas kepemilikan lahan pertanian adalah sekitar 4.000 m2 (BPS 2014). Setiap pemilik lahan memiliki hak mutlak untuk memutuskan tanaman pertanian yang dibudidayakan dan hal ini dijamin oleh negara melalui Undang-Undang nomor 12 tahun 1992 tentang Sistem Budi daya Tanaman (Pemerintah Republik Indonesia 1992). Faktor-faktor tersebut pada akhirnya akan menyebabkan perubahan intensitas penggunaan lahan dan selanjutnya akan mengakibatkan fragmentasi habitat (Kruess dan Tscharntke 2000).
Hymenoptera Parasitika
Hymenoptera memiliki lebih dari 153 000 spesies yang telah diidentifikasi terdiri dari 132 famili dan 8 432 genus (Aguiar et al. 2013). Dengan demikian Hymenoptera merupakan kelompok serangga yang mempunyai keanekaragaman spesies yang tinggi setelah ordo Coleoptera dan Lepidoptera. Namun, Grissell (1999) berargumentasi bahwa jika semua spesies yang belum diidentifikasi turut diperhitungkan maka Hymenoptera merupakan ordo yang memiliki keanekaragaman spesies paling banyak. Penelitian yang dilakukan di daerah beriklim sedang (Gaston 1991) dan di daerah tropis (Stork 1991) menunjukkan bahwa Hymenoptera merupakan ordo yang memiliki keanekaragaman spesies paling banyak. Sharkey (2007) menduga jumlah spesies ordo Hymenoptera mencapai 1 000 000. Hymenoptera merupakan kelompok serangga yang paling banyak berinteraksi dengan organisme lain dalam suatu ekosistem dan memiliki nilai ekonomis yang tinggi. Kelompok ini terdiri dari 39 famili yang bersifat parasitoid, dua famili (Agaonidae dan Tanaostigmatidae) sebagai fitofag, dan beberapa famili sebagai pembuat puru, predator, dan penyerbuk (LaSalle dan Gauld 1993).
sub tropis dibandingkan dengan daerah beriklim sedang (temperate region) (Sharkey 2007).
Apocrita memiliki pertulangan sayap yang sederhana dan ruas pertama abdomen mengalami penggentingan. Berdasarkan struktur ovipositornya subordo ini dibagi menjadi 2 kelompok, yaitu aculeate dan parasitica (Goulet dan Huber 1993). Aculeata adalah kelompok Hymenoptera yang struktur ovipositornya mengalami modifikasi sebagai alat penyengat dan kebanyakan spesiesnya adalah predator dan penyerbuk (LaSalle dan Gauld 1993). Kelompok aculeate terdiri dari 2 superfamili dan 23 famili (Aguiar et al. 2013).
Parasitica adalah kelompok Hymenoptera yang struktur ovipositornya tidak berkembang menjadi alat penyengat. Kelompok parasitica terdiri dari 16 superfamili dan 82 famili (Aguiar et al. 2013). Sebagian besar spesies dari kelompok ini merupakan parasitoid (Goulet dan Huber 1993). Parasitoid adalah serangga yang memarasit serangga lain dan mampu melengkapi siklus hidupnya dalam satu inang (Driesche dan Bellows 1996). Parasitoid merupakan kelompok musuh alami yang telah banyak digunakan dalam pengendalian hayati serangga hama. Umumnya program pengendalian hayati tersebut menggunakan parasitoid yang berasal dari ordo Hymenoptera superfamili Ichneumonoidea dan Chalcidoidea (Driesche dan Bellows 1996; Greathead 1986). Superfamili Ichneumonoidea dan Chalcidoidea merupakan superfamili yang memiliki keanekaragaman spesies tertinggi di antara Hymenoptera parasitika lainnya.
Superfamili Ichneumonoidea
Superfamili Ichneumonoidea memiliki ciri-ciri pronotum berbentuk menyerupai segitiga, dan meluas sampai tegula. Antena berbentuk filiform dan umumnya terdiri dari 16 segmen. Trochanters belakang tersegmentasi menjadi dua. Ovipositor naik di depan apeks abdomen, tidak bisa ditarik, dan umumnya sepanjang atau lebih panjang dari tubuh. Venasi biasanya normal. Sayap depan tidak memiliki costal cell (Goulet dan Huber 1993). Superfamili Ichneumonoidea terdiri dari 3 famili, yaitu Ichneumonidae, Braconidae dan Praeichneumonidae (Aguiar et al. 2013).
Ichneumonidae merupakan kelompok parasitoid yang anggotanya banyak memarasit spesies serangga hama. Subfamili parasitoid dari famili Ichneumonidae dikelompokkan berdasarkan tipe inang yang diparasit, yaitu subfamili Ephialtinae merupakan ektoparasitoid larva atau pupa berbagai ordo serangga di dalam jaringan tanaman, subfamili Typhoninae merupakan ektoparasitoid larva Lepidoptera yang hidup terbuka, subfamili Gelinae merupakan ektoparasitoid serangga dalam kokon, subfamili Brachinae, Porizontinae dan Ophioninae merupakan endoparasitoid larva Lepidoptera, subfamili Ichneumoninae merupakan endoparasitoid pupa Lepidoptera, subfamili Scolobatinae merupakan endoparasitoid sawfly dan subfamili Diplazontinae merupakan endoparasitoid larva Syrphidae (Driesche dan Bellows 1996).
Rogadinae merupakan endoparasitoid larva Lepidoptera dan Coleoptera, subfamili Euphorinae merupakan endoparasitoid imago Coleoptera dan nimfa Hemiptera, subfamili Cheloninae merupakan endoparasitoid telur – larva Lepidoptera, subfamili Alysiinae merupakan endoparasitoid telur dan larva Diptera, subfamili Braconinae merupakan ektoparasitoid larva Lepidoptera yang hidup tersembunyi (Driesche dan Bellows 1996).
Superfamili Chalcidoidea
Superfamili Chalcidoidea memiliki ciri-ciri antena berbentuk genikulat dengan jumlah segmen maksimal 13 segmen. Umunya tubuh berwarna hitam, namun beberapa berwarna hijau metalik atau biru (Goulet dan Huber 1993). Superfamili Chalcidoidea terdiri dari 23 famili (Aguiar et al. 2013), namun hanya 6 famili yang anggotanya banyak digunakan dalam kegiatan pengendalian hayati, yaitu famili Aphelinidae, Chalcididae, Encyrtidae, Eulophidae, Mymaridae dan Trichogrammatidae (Driesche dan Bellows 1996).
Aphelinidae merupakan parasitoid Hemiptera dan parasitoid telur dari berbagai ordo serangga (Goulet dan Huber 1993). Beberapa spesies dari famili ini merupakan agensia pengendali hayati yang efektif mengendalikan hama, contoh Aphelinus asychis efektif mengendalikan 3 spesies aphid (Tatsumi dan Takada 2005), Encarsia diaspidicola merupakan agensia hayati yang efektif mengendalikan Pseudaulacaspis pentagona (Neumann et al. 2010).
Chalcididae terdiri dari 93 genus dan 1474 spesies (Aguiar et al. 2013). Beberapa spesies dari famili ini telah banyak digunakan dalam kegiatan pengendalian hayati seperti Opius sp dan Brachymeria lasus (Waterhause 1998).
Encyrtidae terdiri dari 495 genus dan 4061 spesies (Aguiar et al. 2013). Beberapa spesies dari famili ini telah banyak digunakan dalam kegiatan pengendalian hayati, seperti Diaphorencyrtus aligarhensis digunakan untuk mengendalikan Diaphorina citri, Copidosoma sp. digunakan untuk mengendalikan Trichoplusia ni (Waterhause 1998).
Eulophidae terdiri dari 336 genus dan 4972 spesies (Aguiar et al. 2013). Eulophidae merupakan salah satu famili penting dari superfamili Chalcidoidea. Kisaran ini parasitoid dari famili ini cukup luas, mencakup telur laba-laba, Hemiptera, Thysanoptera, Coleoptera, Lepidoptera, dan Diptera (Goulet dan Huber 1993; Driesche dan Bellows 1996). Beberapa spesies dari famili telah banyak digunakan dalam kegiatan pengendalian hayati, seperti Hemiptarsenus sp. digunakan untuk mengendalikan Ophiomyia phaseoli, Euplectrus sp digunakan untuk mengendalikan Trichoplusia ni (Waterhause 1998).
Trichogrammatidae terdiri dari 100 genus dan 884 spesies (Aguiar et al. 2013). Semua anggota famili ini adalah parasitoid telur serangga lain (Pinto dan Stouthamer 1994). Beberapa spesies dari famili ini seperti Trichogramma minutum dan T. pretiosum telah digunakan dalam pengendalian berbagai jenis Lepidoptera melalui pelepasan massal (Driesche dan Bellows 1996).
Pemencaran dan Persebaran Serangga
sebaran individu suatu spesies dalam ruang, pemencaran serangga dibedakan menjadi acak, kelompok dan teratur (Poole 1974) (Gambar 2.3). Pola sebaran serangga dapat ditentukan dengan menggunakan indeks dispersi (Iδ), yaitu rasio ragam terhadap rata-rata (σ2/μ), nilai-κ Binomial negatif dan indeks Morisita (Iγ) (Poole 1974; Southwood dan Henderson 2000).
Gambar 2.3 Ilustrasi pola pemencaran serangga (Poole 1974)
Pola pemencaran hama perlu diketahui untuk menetapkan pola pengambilan contoh yang sesuai baik untuk perencanaan penelitian maupun monitoring dinamika populasi hama (Ruiz-Cardenas et al. 2009). Menurut Ruesink dan Kogan (1975) pola sebaran hama penting diketahui untuk menetapkan pola pengambilan contoh dan jumlah contoh yang sesuai. Sedangkan pola sebaran musuh alami perlu diketahui untuk memastikan efektifitas musuh alami tersebut. Pola sebaran serangga di lapangan terdiri dari merata, acak, dan kelompok (Ruesink dan Kogan 1975). Pada umumnya, pola sebaran serangga adalah mengelompok, seperti Diaphorina citri Kuwayama (Costa et al. 2010), penggerek buah kopi (Wiryadiputra 2014), kelompok telur Ostrinia furnacalis (Lopez 2014), dan parasitoid Cotesia flavipes (Dinardo-Miranda 2014).
Pola pemencaran musuh musuh alami perlu diketahui untuk melakukan konservasi terhadap musuh alami tersebut. Dengan memahami pergerakan parasitoid maka akan diketahui habitat yang berperan sebagai source maupun sink bagi parasitoid tersebut sehingga dapat diketahui dengan pasti lokasi penempatan refugia dan tanaman penghasil nektar serta serbuk sari bagi parasitoid (Gurr et al. 2003; Gurr et al. 2004; Wratten et al. 2004). Sebaran dan gerakan serangga dapat dipelajari dengan cara menandai serangga tersebut.
Pergerakan serangga dapat dipelajari dengan sejumlah teknik, seperti tags, mutilasi, pewarna, serbuk, isotop stabil, genetik, radioisotop, dan protein (Hagler dan Jackson 2001; Lavandero et al. 2004a,b). Metode yang digunakan harus mempertimbangkan beberapa hal, antara lain tidak berpengaruh pada serangga yang ditandai, murah, tahan lama, dan mudah untuk diterapkan. Setiap teknik penandaan serangga mungkin memiliki efek yang berbeda pada setiap serangga, oleh karena itu perlu dilakukan studi pendahuluan sebelum suatu teknik penandaan digunakan dalam studi pemencaran serangga (Hagler dan Jackson 2001).
3
KOMPOSISI DAN KONFIGURASI LANSKAP PERTANIAN
DI KAWASAN BOGOR, CIANJUR DAN SUKABUMI
JAWA BARAT
Pendahuluan
Lanskap terdiri dari berbagai elemen yang sangat memengaruhi keanekaragaman, komposisi, kelimpahan, dan sebaran serangga. Oleh karena itu, kuantifikasi di tingkat kelas penggunaan lahan merupakan prasyarat dalam menganalisis lanskap (Turner 1987). Lebih lanjut Turner (2001) menjelaskan bahwa dalam menganalisis lanskap harus dipahami berapa dan bagaimana elemen lanskap tersebut disusun. Kuantifikasi lanskap perlu dilakukan untuk memahami komposisi dan konfigurasi lanskap. Sejumlah penelitian telah menunjukkan bahwa komposisi dan konfigurasi lanskap memengaruhi kelimpahan dan kekayaan spesies serangga (Hunter 2002, Marino dan Landis 1996). Peningkatan fragmentasi habitat pada skala lokal dapat menyebabkan kepunahan spesies beberapa serangga (Landis et al. 2000). Menurut Kruess dan Tscharntke (1994) fragmentasi habitat adalah peristiwa yang menyebabkan habitat yang luas dan berkelanjutan diperkecil atau dibagi menjadi dua atau lebih fragmen. Perangkat spasial remote sensing GIS memiliki kapasitas untuk mengkuantifikasi pola penggunaan lahan dan memahami keanekaragaman spasial (Turner 1990; Sinha dan Sharma 2006).
Intensifikasi dan ekstensifikasi pertanian telah mengakibatkan perubahan struktur spasial, keanekaragaman habitat dan komposisi habitat. Lanskap pertanian yang berada di sekitar Gunung Salak dan Gunung Gede Pangrango terdapat pada dataran rendah hingga sedang dan memiliki tingkat kompleksitas yang beragam. Lanskap sederhana dicirikan dengan rendahnya proporsi habitat alami, sedangkan lanskap kompleks memiliki proporsi habitat alami yang lebih tinggi. Karakteristik lanskap seperti konfigurasi dan komposisi elemen lanskap memengaruhi keanekaragaman hayati (Forman dan Godron 1986; Cushman et al. 2010). Untuk mempelajari hubungan antara kompleksitas lanskap dengan keanekaragaman serangga maka struktur lanskap harus dikuantifikasi.
Tujuan penelitian ini adalah untuk mengetahui komposisi dan konfigurasi lanskap pertanian di wilayah Bogor, Cianjur dan Sukabumi, Jawa Barat.
Bahan dan Metode
Lokasi dan Waktu
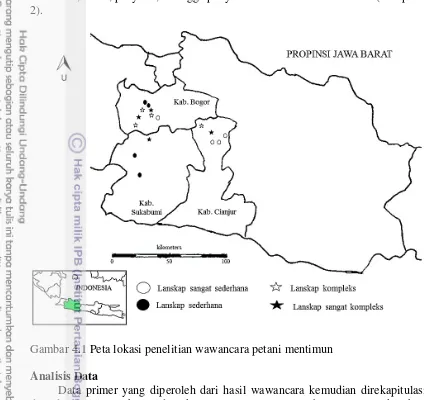
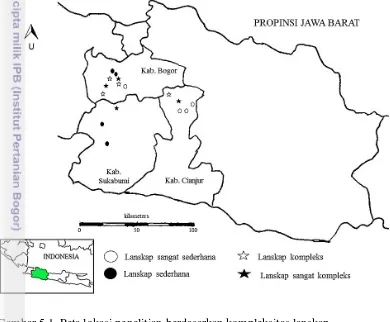
Penelitian dilaksanakan di lanskap pertanian yang berada di kawasan Bogor, Sukabumi dan Cianjur, Jawa Barat (Gambar 3.1). Penelitian ini dilaksanakan mulai Oktober 2014 sampai dengan Februari 2015.
Gambar 3.1 Lokasi penelitian di kawasan Bogor, Sukabumi dan Cianjur (16 lokasi)
Penentuan Lokasi Penelitian
Pengumpulan Data Spasial Lanskap
[image:40.595.50.486.151.783.2]Pengumpulan data spasial lanskap diawali dengan menentukan titik koordinat lokasi penelitian menggunakan global positioning system (GPS). Lokasi penelitian dan informasi geografisnya disajikan pada Tabel 3.1. Titik koordinat tersebut selanjutnya digunakan untuk menduga posisi masing-masing lokasi pertanian melalui Google earth sehingga diperoleh foto udara untuk setiap lokasi penelitian (Gambar 3.2). Selanjutnya dilakukan groundcheck untuk memastikan komoditas yang ditanam serta menentukan batas areal pertanian dengan areal lain dengan menggunakan GPS. Metode groundcheck yang dilakukan mengadopsi metode yang dilakukan oleh Thies et al. (2003) yaitu dengan cara menggambar kondisi lanskap pada radius 100 m, 200 m, 300 m, 400 m dan 500 m dalam bentuk peta berdasarkan penggunaan lahan dan jenis komoditas yang ditanam.
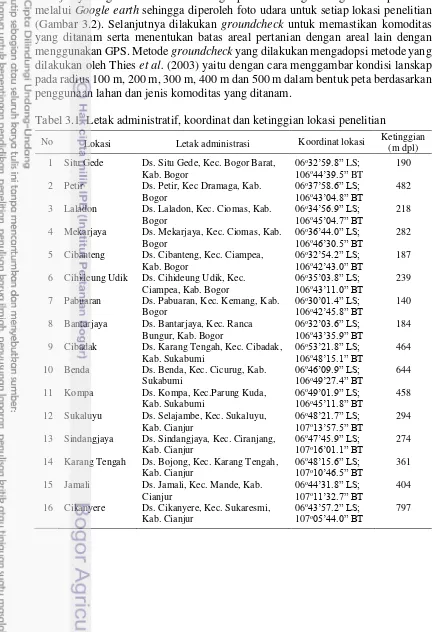
Tabel 3.1 Letak administratif, koordinat dan ketinggian lokasi penelitian
No Lokasi Letak administrasi Koordinat lokasi Ketinggian (m dpl) 1 Situ Gede Ds. Situ Gede, Kec. Bogor Barat,
Kab. Bogor
06o32’59.8” LS; 106o44’39.5” BT
190
2 Petir Ds. Petir, Kec Dramaga, Kab. Bogor
06o37’58.6” LS; 106o43’04.8” BT
482
3 Laladon Ds. Laladon, Kec. Ciomas, Kab. Bogor
06o34’56.9” LS; 106o45’04.7” BT
218
4 Mekarjaya Ds. Mekarjaya, Kec. Ciomas, Kab. Bogor
06o36’44.0” LS; 106o46’30.5” BT
282
5 Cibanteng Ds. Cibanteng, Kec. Ciampea, Kab. Bogor
06o32’54.2” LS; 106o42’43.0” BT
187
6 Cihideung Udik Ds. Cihideung Udik, Kec. Ciampea, Kab. Bogor
06o35’03.8” LS; 106o43’11.0” BT
239
7 Pabuaran Ds. Pabuaran, Kec. Kemang, Kab. Bogor
06o30’01.4” LS; 106o42’45.8” BT
140
8 Bantarjaya Ds. Bantarjaya, Kec. Ranca Bungur, Kab. Bogor
06o32’03.6” LS; 106o43’35.9” BT
184
9 Cibadak Ds. Karang Tengah, Kec. Cibadak, Kab. Sukabumi
06o53’21.8” LS; 106o48’15.1” BT
464
10 Benda Ds. Benda, Kec. Cicurug, Kab. Sukabumi
06o46’09.9” LS; 106o49’27.4” BT
644
11 Kompa Ds. Kompa, Kec.Parung Kuda, Kab. Sukabumi
06o49’01.9” LS; 106o45’11.8” BT
458
12 Sukaluyu Ds. Selajambe, Kec. Sukaluyu, Kab. Cianjur
06o48’21.7” LS; 107o13’57.5” BT
294
13 Sindangjaya Ds. Sindangjaya, Kec. Ciranjang, Kab. Cianjur
06o47’45.9” LS; 107o16’01.1” BT
274
14 Karang Tengah Ds. Bojong, Kec. Karang Tengah, Kab. Cianjur
06o48’15.6” LS; 107o10’46.5” BT
361
15 Jamali Ds. Jamali, Kec. Mande, Kab. Cianjur
06o44’31.8” LS; 107o11’32.7” BT
404
16 Cikanyere Ds. Cikanyere, Kec. Sukaresmi, Kab. Cianjur
06o43’57.2” LS; 107o05’44.0” BT
Gambar 3. 2 Foto udara lokasi penelitian yang digunakan sebagai pedoman dalam melakukan groundcheck. (A) Sukaluyu, (B) Kompa, (C) Bantarjaya, dan (D) Jamali.
Analisis Data
Data yang diperoleh dari hasil groundcheck dipadukan dengan foto udara yang diperoleh dari Google earth dan digunakan untuk membuat peta penggunaan lahan dengan bantuan perangkat lunak Quantum GIS (Quantum GIS Development Team 2014). Peta penggunaan lahan tersebut selanjutnya digunakan untuk melakukan analisis habitat dengan bantuan perangkat lunak FRAGSTATS versi 4 (McGarigal et al. 2014). Ukuran-ukuran lanskap dalam analisis habitat diperoleh berdasarkan metode McGarigal et al. (2014), meliputi:
Class Area (CA), jumlah keseluruhan area semua patch pada kelas yang sama (ha).
Total Landscape Area (TLA), jumlah keseluruhan luasan patch dalam lanskap (ha).
Mean Patch Size (MPS), rata-rata luas patch pada kelas yang sama(ha). Nilai MPS yang kecil menunjukkan bahwa suatu patch semakin terfragmentasi.
Total Edge (TE), merupakan total perimeter/panjang dari edge (m).
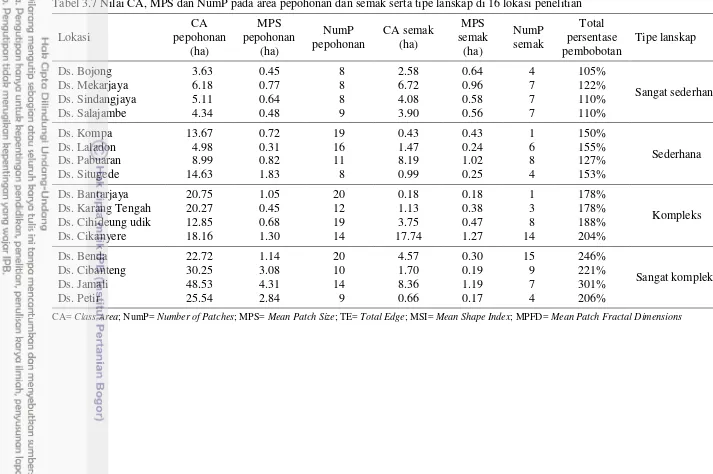
Lokasi penelitian dikelompokkan berdasarkan kompleksitas lanskap dan jarak ke habitat alami. Berdasarkan kompleksitas lanskap, lokasi penelitian dikelompokkan menjadi empat tipe lanskap, yaitu sangat sederhana, sederhana, kompleks dan sangat kompleks. Pengelompokan ini dilakukan berdasarkan skor dan bobot nilai CA, NumP dan MPS habitat alami (Tabel 3.2). Persentase bobot ditentukan berdasarkan pengaruh parameter lanskap terhadap kompleksitas lanskap. Berdasarkan jarak lokasi pertanaman mentimun dari habitat pepohonan, lokasi penelitian dikelompokkan menjadi dua, yaitu lokasi penelitian yang berjarak jauh
(≥ 400 m) dan dekat (≤ 200 m) dari habitat alami.
Tabel 3.2 Skor dan bobot parameter lanskap yang digunakan pada pengelompokan lanskap
Skor
Parameter lanskap
Pepohonan Semak
CA (ha) NumP MPS (ha) CA (ha) NumP MPS (ha) 1 4 – 12 8 – 11 0.3 – 1.3 0.2 – 4.6 1 – 4 0.2 – 0.4 2 13 – 21 12 – 15 1.4 – 2.3 4.7 – 8.9 5 – 8 0.5 – 0.7 3 22 – 30 16 – 19 2.4 – 3.3 9.0 – 13.4 9 – 12 0.8 – 1.0 4 31 – 39 20 – 23 3.4 – 4.3 13.5 – 17.7 13 – 16 1.1 – 1.3
Bobot 33% 25% 25% 7% 5% 5%
CA= Class Area; NumP= Number of Patches; MPS= Mean Patch Size
Hasil
Komposisi Lanskap
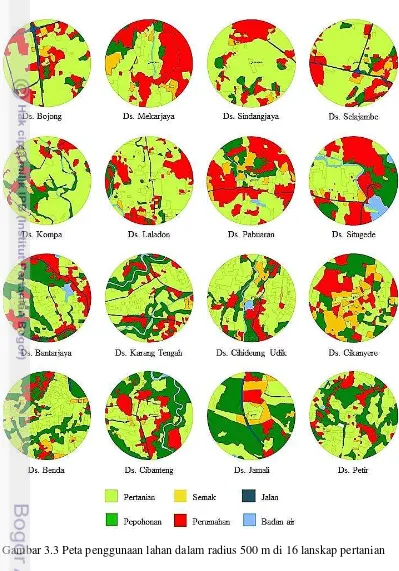
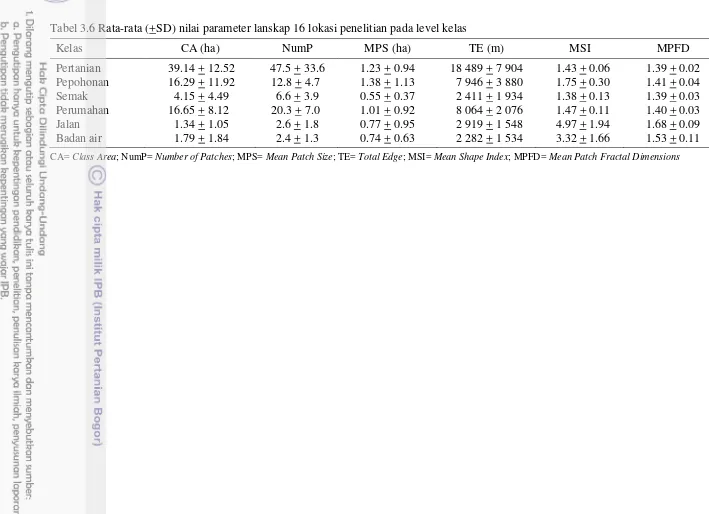
Lanskap pertanian di wilayah Bogor, Sukabumi dan Cianjur umumnya digunakan sebagai lahan pertanian, semak, pepohonan, perumahan, jalan dan sumber air (Gambar 3.3). Secara umum, penggunaan lahan didominasi oleh lahan pertanian (16.20% – 68.56%) (Tabel 3.3). Proporsi area pepohonan di lokasi penelitian bervariasi antara 4.60% - 61.40%. Proporsi luas area pepohonan tertinggi ditemukan di lokasi penelitian Desa Jamali, sedangkan proporsi terendah ditemukan di Desa Bojong (Tabel 3.3). Proporsi luas area pepohonan berkurang seiring dengan bertambah luasnya area pertanian (AdjR2= 0.39, P= 0.006) (Gambar 3.4).
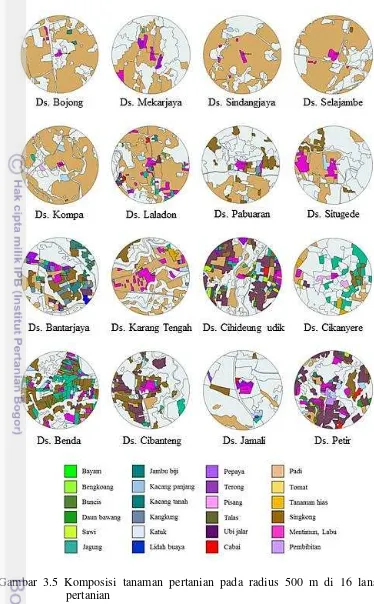
3 – 15 tanaman (Tabel 3.3). Lokasi penelitian yang berada di Desa Sindangjaya memiliki keragaman tanaman pertanian yang rendah karena daerah tersebut hanya ditanami oleh 3 jenis tanaman pertanian dan keragaman tertinggi ditemukan di daerah Petir (15 tanaman budi daya) (Tabel 3.4).
[image:43.595.111.510.150.721.2]
Tabel 3.3 Proporsi penggunaan lahan dalam radius 500 m (78.56 ha) di 16 lanskap pertanian
No Lokasi Proporsi luas (%)
A B C D E F
1 Ds. Bojong 64.31 4.60 3.27 23.03 4.78 0.00 2 Ds. Mekarjaya 40.74 7.87 8.56 42.06 0.00 0.77 3 Ds. Sindangjaya 65.92 6.51 5.20 21.92 0.45 0.00 4 Ds. Salajambe 65.87 5.53 4.97 20.17 3.46 0.00 5 Ds. Kompa 68.53 17.41 0.54 10.65 1.31 1.55 6 Ds. Laladon 68.56 6.34 1.87 21.82 0.62 0.78 7 Ds. Pabuaran 35.09 11.45 10.42 40.91 0.62 1.51 8 Ds. Situgede 35.38 18.64 1.26 34.86 1.30 8.56 9 Ds. Bantarjaya 48.16 25.41 0.23 21.81 2.40 1.99 10 Ds. Karang Tengah 52.93 25.81 1.44 14.07 2.29 3.45 11 Ds. Cihideung udik 61.25 16.37 4.78 13.95 0.83 2.83 12 Ds. Cikanyere 30.55 23.12 22.60 23.72 0.00 0.00 13 Ds. Benda 56.18 28.98 5.83 8.58 0.44 0.00 14 Ds. Cibanteng 39.41 35.33 2.16 18.98 1.11 3.00 15 Ds. Jamali 28.52 49.08 10.58 9.07 2.60 0.15 16 Ds. Petir 52.48 35.52 0.85 13.60 0.00 0.55 A= Pertanian; B= Pepohonan; C= Semak; D= Perumahan; E= Jalan; F= Badan air
Tabel 3.4 Luas area pertanian dalam radius 500 m dan jumlah jenis tanaman yang ditanami pada area tersebut
No Lokasi Luas area pertanian
(ha)
Jumlah komoditas pertanian
1 Ds. Bojong 50.64 4
2 Ds. Mekarjaya 31.99 5
3 Ds. Sindangjaya 51.76 3
4 Ds. Salajambe 51.72 5
5 Ds. Kompa 53.81 6
6 Ds. Laladon 53.83 9
7 Ds. Pabuaran 27.55 5
8 Ds. Situgede 27.78 5
9 Ds. Bantarjaya 37.01 13
10 Ds. Karang Tengah 41.56 10
11 Ds. Cihideung udik 48.09 10
12 Ds. Cikanyere 23.99 9
13 Ds. Benda 44.05 11
14 Ds. Cibanteng 28.44 8
15 Ds. Jamali 12.80 8
16 Ds. Petir 41.21 15
Struktur Lanskap
Lanskap di lokasi penelitian ditentukan berdasarkan dua tingkat hierarki mulai dari tingkat yang lebih luas yaitu skala lanskap dan dilanjutkan dengan tingkat yang lebih sempit yaitu skala kelas.
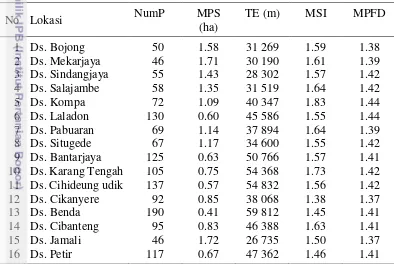
Skala Lanskap. Hasil analisis spasial lanskap menunjukkan bahwa lokasi penelitian di Desa Benda memiliki jumlah patch (NumP) yang paling banyak, yaitu 190 patch, kemudian diikuti oleh Desa Cihideung udik (137 patch) dan Desa Laladon (130 patch) (Tabel 3.5). Hal ini menunjukkan bahwa lanskap pertanian di lokasi tersebut membentuk kelompok-kelompok kecil (terpecah-pecah) sehingga lanskap tersebut mengalami fragmentasi. Jumlah patch terkecil ditemukan di lokasi penelitian yang berada di Desa Mekarjaya dan Jamali yaitu 46 patch, diikuti dengan Desa Bojong (50 patch) dan Sindangjaya (55 patch). Lanskap pe